
Functional Analysis of Glucose-6-Phosphate Translocator PoGPT1 in Embryogenic Callus Formation of Tree Peony
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
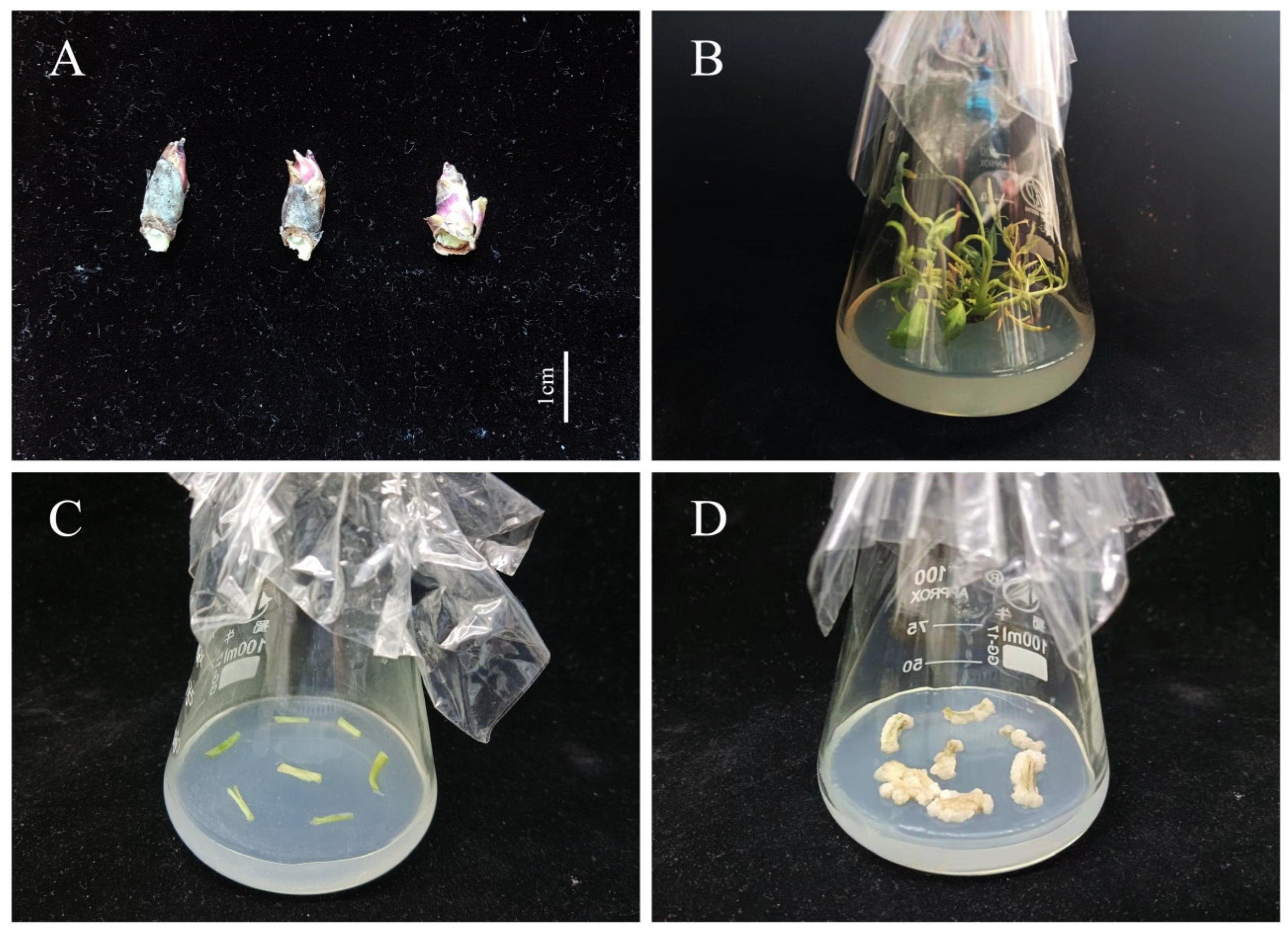
2.1. Plant Materials and Culture Conditions
2.2. Cloning and Analysis of GPT1
2.3. Subcellular Localization Analysis
2.4. Plasmid Construction and Transformation of Tree Peony Callus
2.5. Determination of Plant Hormone Content
2.6. Morphological and Anatomical Observation
2.7. Real-Time Quantitative PCR (RT-qPCR) Analysis
2.8. Statistical Analysis
3. Results
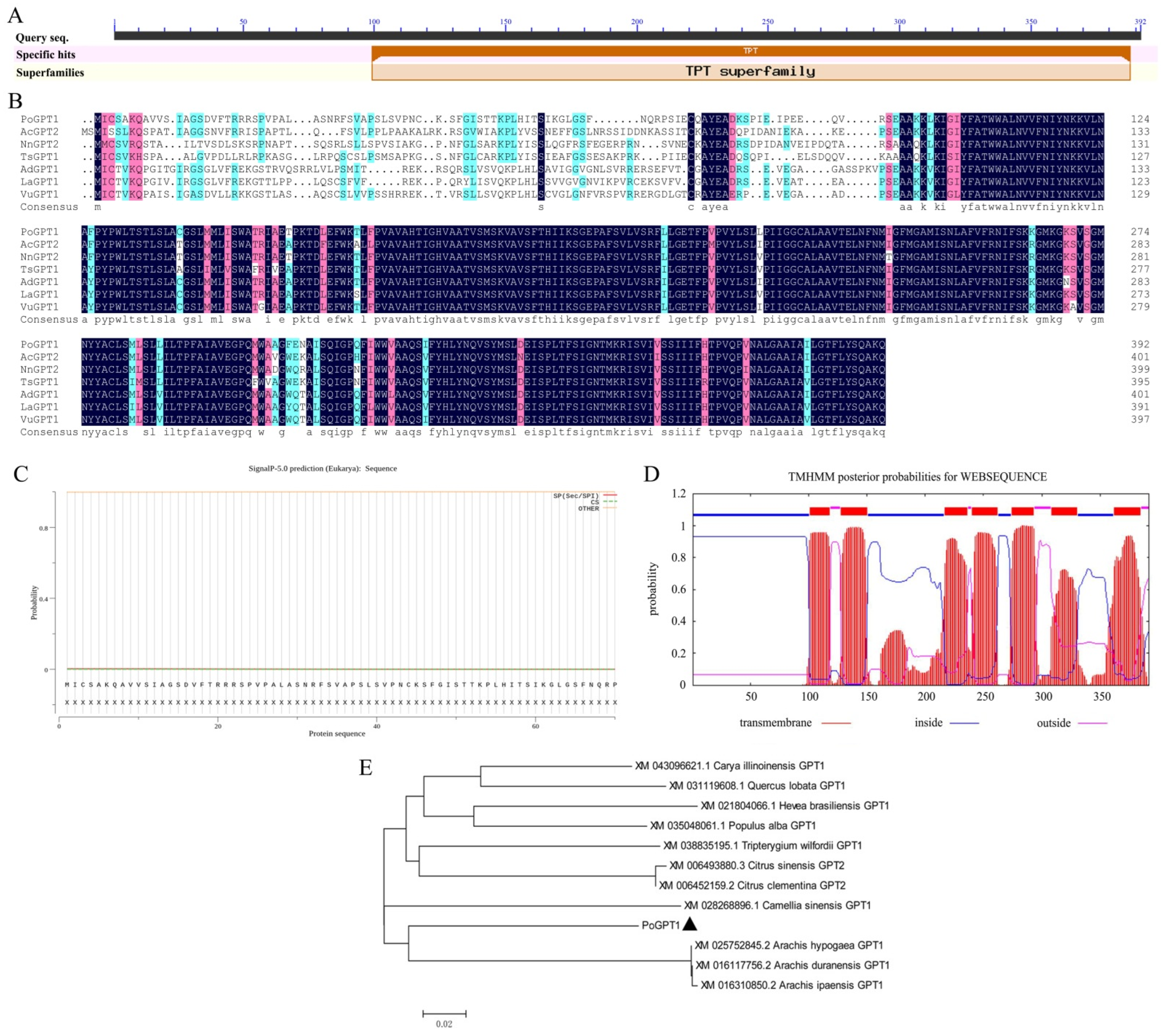
3.1. Sequence Analysis of PoGPT1
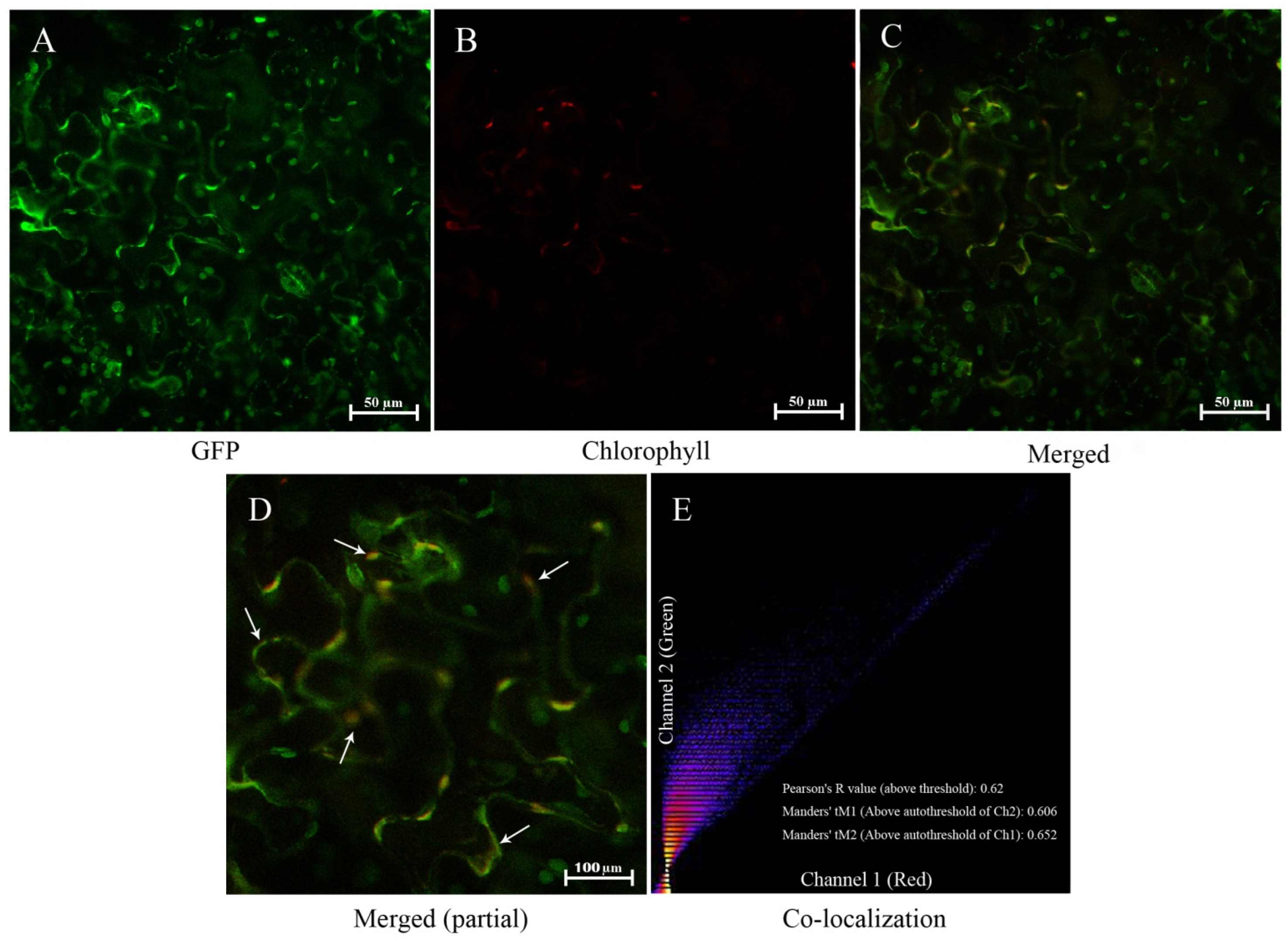
3.2. Subcellular Localization of PoGPT1
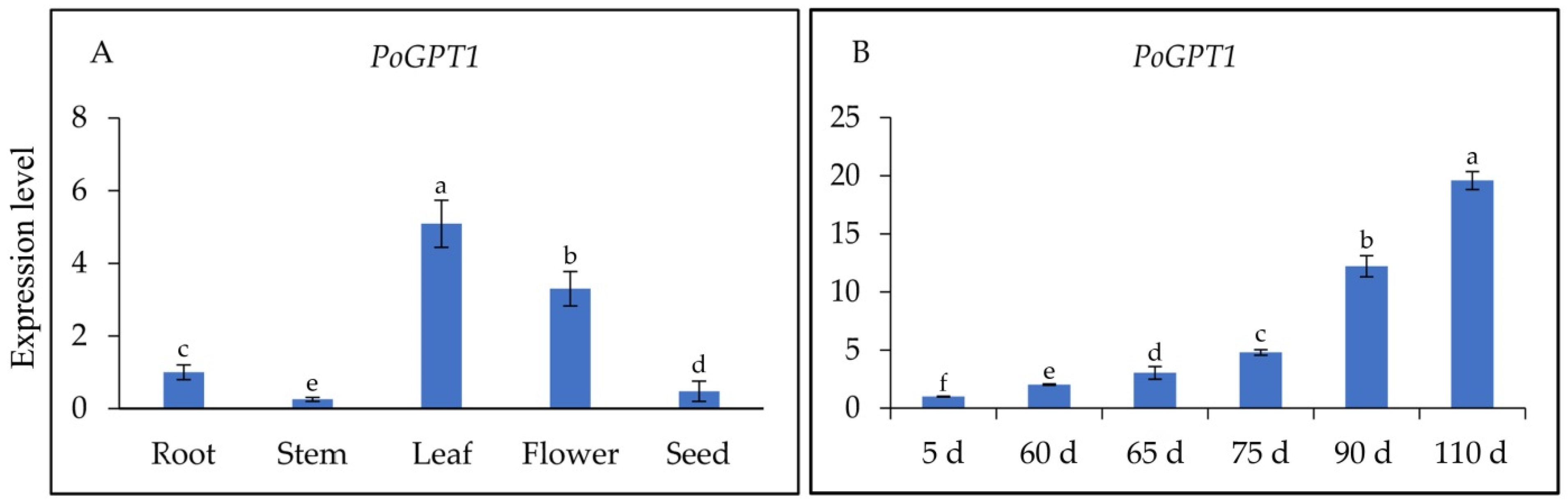
3.3. Spatiotemporal Expression Analysis of PoGPT1
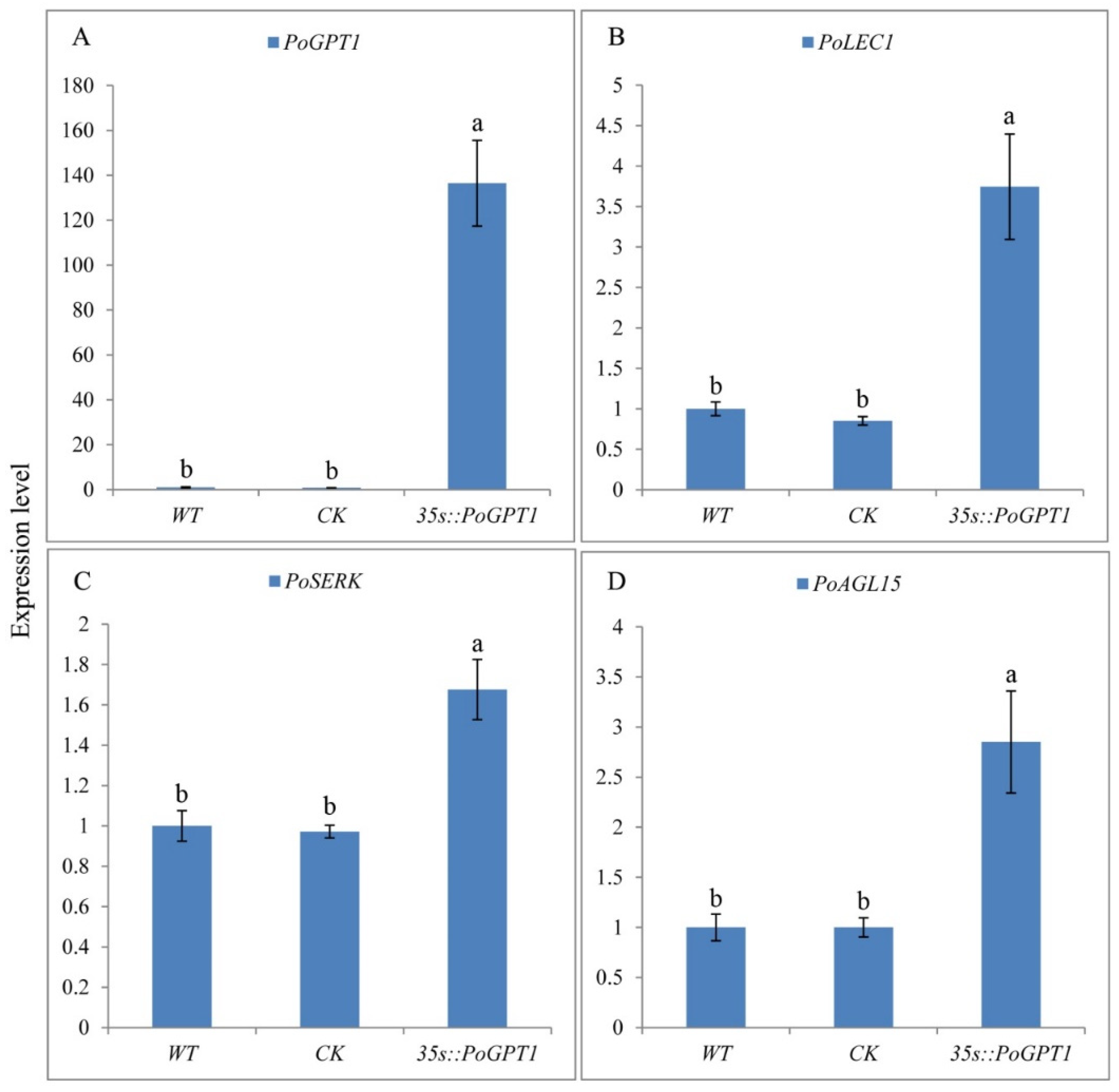
3.4. RT-qPCR Analysis of PoGPT1 and Genes Associated with Somatic Embryogenesis
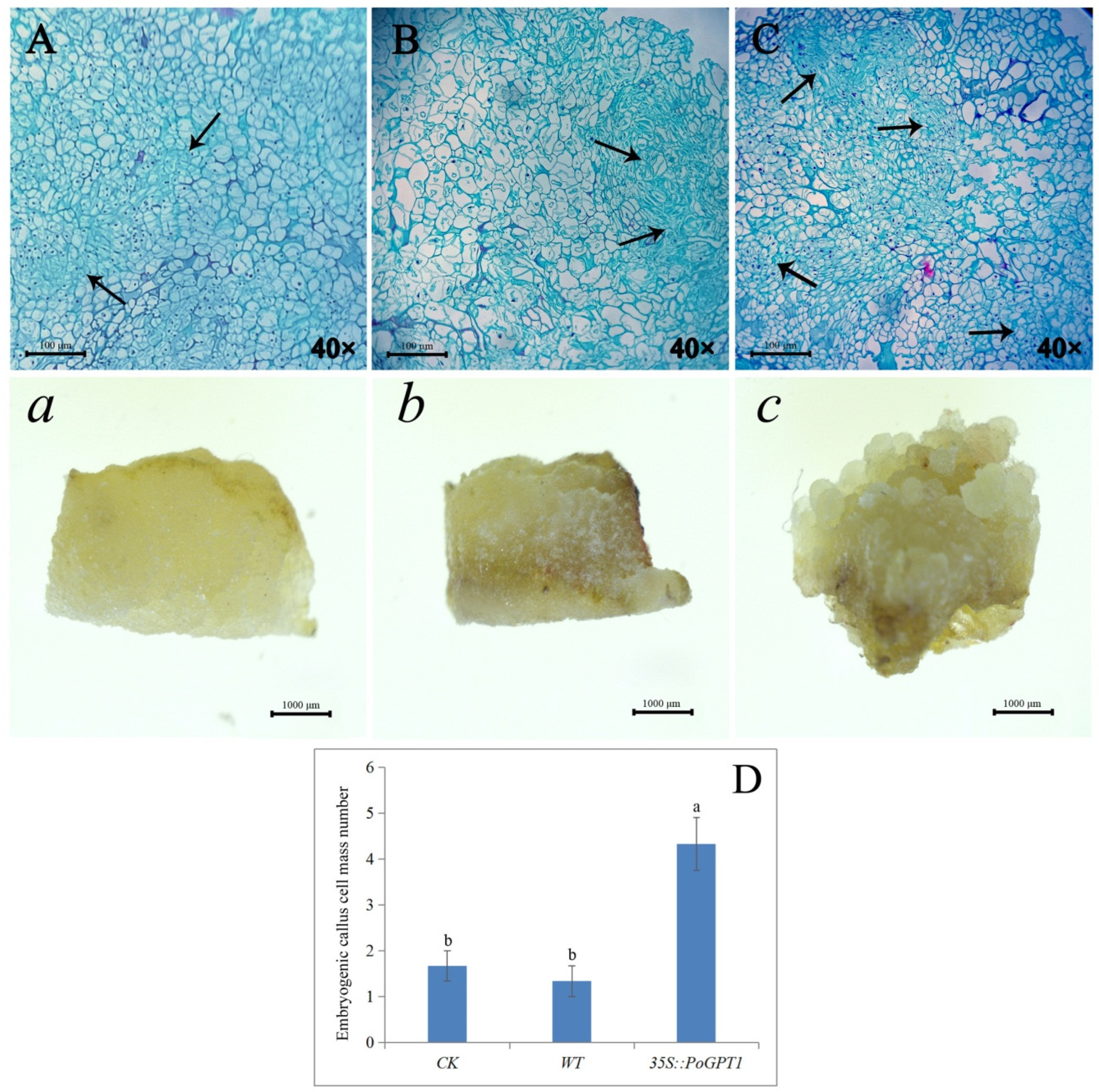
3.5. Anatomical Observations
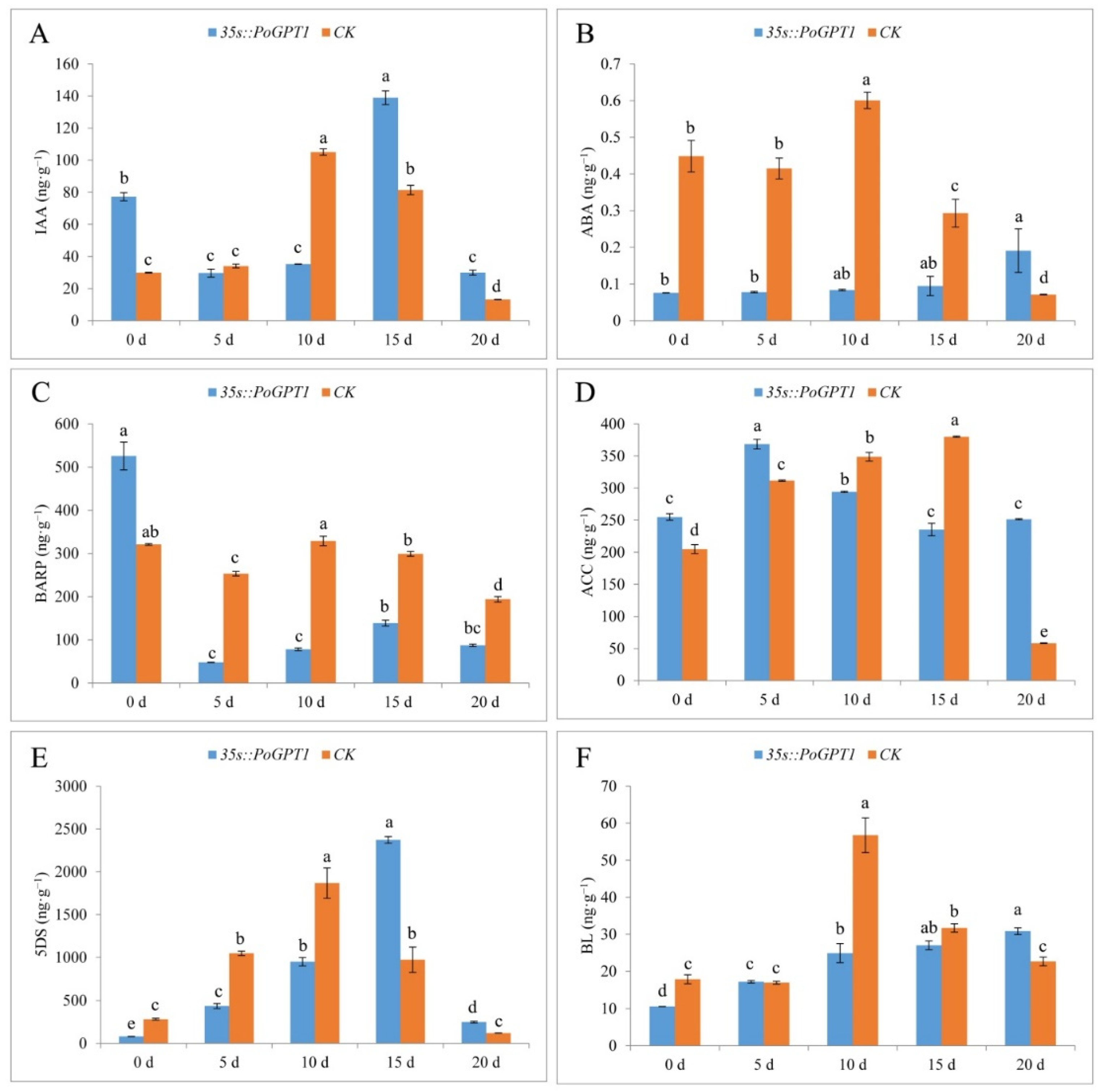
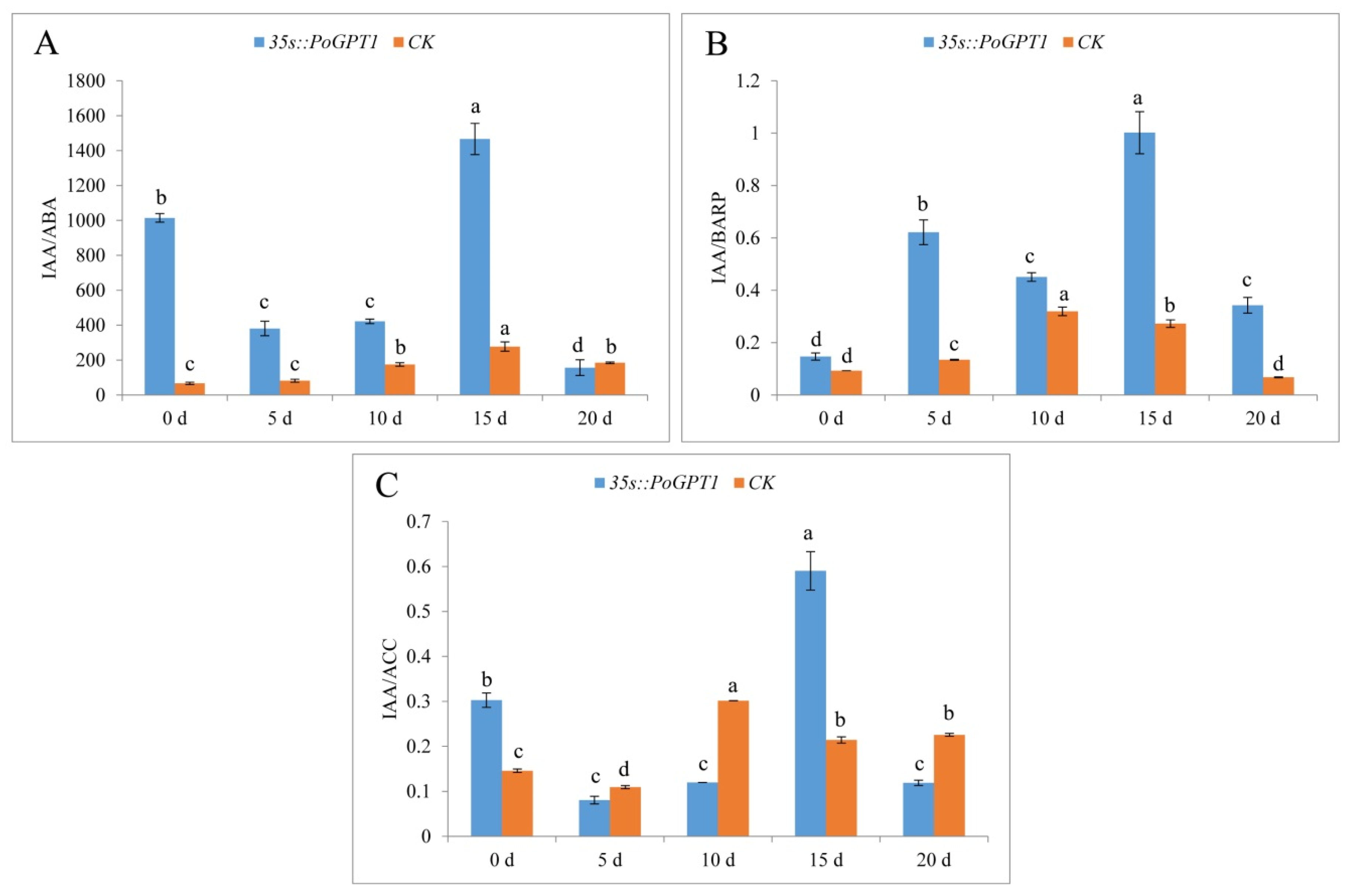
3.6. Endogenous Plant Hormone Contents
4. Discussion
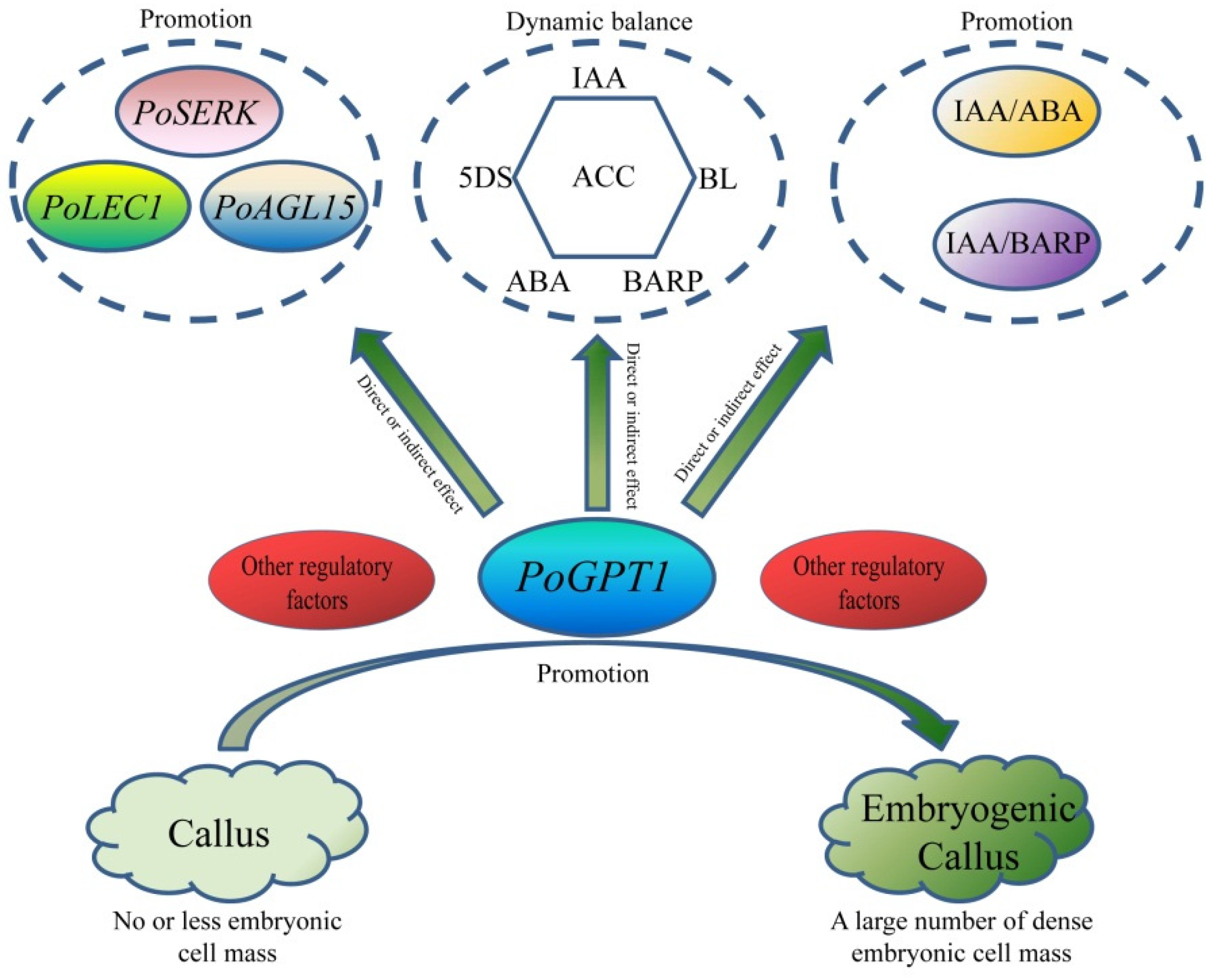
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Wang, Y.J.; Dong, C.L.; Xue, Z.Y.; Jin, Q.J.; Xu, Y.C. De novo transcriptome sequencing and discovery of genes related to copper tolerance in Paeonia ostii. Gene 2016, 576, 126–135. [Google Scholar] [CrossRef]
- Wu, F.; Zhang, C.; Wang, X.Q.; Guo, J.; Dong, L. Ethylene-influenced development of tree peony cut flowers and characterization of genes involved in ethylene biosynthesis and perception. Postharvest Biol. Technol. 2017, 125, 150–160. [Google Scholar] [CrossRef]
- Zhang, H.F.; Li, X.F.; Wu, K.; Wang, M.K.; Liu, P.; Wang, X.S.; Deng, R.X. Antioxidant Activities and Chemical Constituents of Flavonoids from the Flower of Paeonia ostii. Molecules 2017, 22, 5. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Xue, J.Q.; Xue, Y.Q.; Liu, R.; Ren, X.X.; Wang, S.L.; Zhang, X.X. Transcriptome sequencing and identification of key callus browning-related genes from petiole callus of tree peony (Paeonia suffruticosa cv. Kao) cultured on media with three browning inhibitors. Plant Physiol. Biochem. 2020, 149, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.S.; Zhang, W.B.; Chang, Y.T.; Ma, Y.; Deng, Y.Y.; Fan, K.K.; Zhang, X.; Jiang, Z.H.; Hu, T. A Preliminary Investigation on the Functional Validation and Interactions of PoWOX Genes in Peony (Paeonia ostii). Horticulturae 2022, 8, 266. [Google Scholar] [CrossRef]
- Shang, W.Q.; Wang, Z.; He, S.L.; Liu, Y.P.; Fu, Z.Z. Research on the relationship between phenolic acids and rooting of tree peony (Paeonia suffruticosa) plantlets in vitro. Sci. Hortic. 2017, 224, 53–60. [Google Scholar] [CrossRef]
- Du, Y.M.; Cheng, F.Y.; Zhong, Y. Induction of direct somatic embryogenesis and shoot organogenesis and histological study in tree peony (Paeonia sect. Moutan). Plant Cell Tissue Organ Cult. 2020, 141, 557–570. [Google Scholar] [CrossRef]
- Ren, X.X.; Liu, Y.; Jeong, B.R. Callus induction and browning suppression in tree peony Paeonia ostii ‘Fengdan’. Hortic. Environ. Biotechnol. 2020, 61, 591–600. [Google Scholar] [CrossRef]
- Fu, Z.Z.; Xu, M.L.; Wang, H.J.; Li, Y.M.; Wang, L.M.; Gao, J.; Zhang, J.; Yuan, X.; Zhang, H.C. Analysis of the transcriptome and related physiological indicators of tree peony (Paeonia suffruticosa Andr.) plantlets before and after rooting in vitro. Plant Cell Tissue Organ Cult. 2021, 147, 529–543. [Google Scholar] [CrossRef]
- Zhu, X.T.; Li, X.Q.; Ding, W.J.; Jin, S.H.; Wang, Y. Callus induction and plant regeneration from leaves of peony. Hortic. Environ. Biotechnol. 2018, 59, 575–582. [Google Scholar] [CrossRef]
- Wen, S.S.; Chen, L.; Cheng, F.Y.; Tian, R.N. Correction to: Micropropagation of tree peony (Paeonia sect. Moutan): A review. Plant Cell Tissue Organ Cult. 2020, 141, 15. [Google Scholar] [CrossRef]
- Zhang, R.L.; Wang, X.B.; Shao, L.M.; Li, D.Q.; Xia, Y.P.; Zhang, J.P. Tissue culture and genetic transformation system construction of Paeonia lactiflora and Paeonia suffruticosa. Plant Physiol. J. 2021, 57, 235–247. [Google Scholar] [CrossRef]
- Karami, O.; Rahimi, A.; Mak, P.; Horstman, A.; Boutilier, K.; Compier, M.; Zaal, B.; Offringa, R. An Arabidopsis AT-hook motif nuclear protein mediates somatic embryogenesis and coinciding genome duplication. Nat. Commun. 2020, 12, 2508. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Liu, M.; Li, M.Q.; Li, P.; Zhou, R.; Jiang, F.L.; Wu, Z. Changes in Endogenous Hormone Contents and Related Gene Expression during Somatic Embryogenesis of Garlic. Acta Bot. Boreali-Occident. Sin. 2021, 41, 1175–1187. [Google Scholar] [CrossRef]
- He, G.M.; Cheng, F.Y.; Li, P. Preliminary studies on culture in vitro of ovule and immature embryo of two tree peony cultivars. Acta Hortic. Sin. 2006, 1, 185. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, D.J.; He, S.L.; Mao, C.C.; Meng, X.Y.; He, D. Effects of different treatments on physiological biochemical properties of early somatic embryogenesis of Paeonia suffruticosa Andr. J. Henan Agric. Sci. 2018, 47, 105–111. [Google Scholar] [CrossRef]
- Shen, M.M.; Wang, Q.; Yu, X.N.; Teixeira da Silva, J.A. Micropropagation of herbaceous peony (Paeonia lactiflora Pall.). Sci. Hortic. 2012, 148, 30–38. [Google Scholar] [CrossRef]
- Flügge, U.-I. Phosphate translocators in plastids. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 27–45. [Google Scholar] [CrossRef] [Green Version]
- Baune, M.C.; Lansing, H.; Fischer, K.; Meyer, T.; Charton, L.; Linka, N.; von Schaewen, A. The Arabidopsis plastidial glucose-6-phosphate transporter GPT1 is dually targeted to peroxisomes via the endoplasmic reticulum. Plant Cell 2020, 32, 1703–1726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kammerer, B.; Fischer, K.; Hilpert, B.; Schubert, S.; Gutensohn, M.; Weber, A.; Flügge, U.-I. Molecular characterization of a carbon transporter in plastids from heterotrophic tissues: The glucose 6-phosphate/phosphate antiporter. Plant Cell 1998, 10, 105–117. [Google Scholar] [CrossRef]
- Jeong, C.Y.; Kim, J.H.; Lee, W.J.; Jin, J.Y.; Kim, J.; Hong, S.W.; Lee, H. At Myb56 regulates anthocyanin levels via the modulation of AtGPT2 expression in response to sucrose in Arabidopsis. Mol. Cells 2018, 41, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Knappe, S.; Flügge, U.-I.; Fischer, K. Analysis of the plastidic phosphate translocator gene family in Arabidopsis and identification of new phosphate translocator-homologous transporters, classified by their putative substratebinding site. Plant Physiol. 2003, 131, 1178–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunz, H.H.; Häusler, R.E.; Fettke, J.; Herbst, K.; Niewiadomski, P.; Gierth, M.; Bell, K.; Steup, M.; Flügge, U.-I.; Schneider, A. The role of plastidial glucose-6-phosphate/phosphate translocators in vegetative tissues of Arabidopsis thaliana mutants impaired in starch biosynthesis. Plant Biol. 2010, 12, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Dyson, B.C.; Allwood, J.W.; Feil, R.; Xu, Y.; Miller, M.; Bowsher, C.G.; Goodacre, R.; Lunn, J.E.; Johnson, G.N. Acclimation of metabolism to light in Arabidopsis thaliana: The glucose 6-phosphate/phosphate translocator GPT2 directs metabolic acclimation. Plant Cell Environ. 2015, 38, 1404–1417. [Google Scholar] [CrossRef] [Green Version]
- Andriotis, V.M.E.; Pike, M.J.; Bunnewell, S.; Hills, M.J.; Smith, A.M. The plastidial glucose-6-phosphate/phosphate antiporter GPT1 is essential for morphogenesis in Arabidopsis embryos. Plant J. 2010, 64, 128–139. [Google Scholar] [CrossRef]
- Zhang, M.M.; Xu, X.W.; Zheng, Y.P.; Zhang, Y.; Deng, X.X.; Lou, S.; Wu, Q.P.; Xu, J.; Zhang, S.Q. Expression of a plastid-localized sugar transporter in the suspensor is critical to embryogenesis. Plant Physiol. 2020, 185, 1021–1038. [Google Scholar] [CrossRef]
- Zheng, Y.P.; Deng, X.X.; Qu, A.; Zhang, M.M.; Tao, Y.; Yang, L.Y.; Liu, Y.D.; Xu, J.; Zhang, S.Q. Regulation of pollen lipid body biogenesis by MAP kinases and downstream WRKY transcription factors in Arabidopsis. PLoS Genet. 2018, 14, e1007880. [Google Scholar] [CrossRef] [Green Version]
- Qu, A.L.; Xu, Y.; Yu, X.X.; Si, Q.; Xu, X.W.; Liu, C.H.; Yang, L.L.; Zheng, Y.P.; Zhang, M.M.; Zhang, S.Q.; et al. Sporophytic control of anther development and male fertility by glucose-6-phosphate/phosphate translocator 1 (OsGPT1) in rice. J. Genet. Genom. 2021, 48, 695–705. [Google Scholar] [CrossRef]
- Niewiadomski, P.; Knappe, S.; Geimer, S.; Fischer, K.; Schulz, B.; Unte, U.S.; Rosso, M.G.; Ache, P.; Flügge, U.-I.; Schneider, A. The Arabidopsis plastidic glucose 6-phosphate/phosphate translocator GPT1 is essential for pollen maturation and embryo sac development. Plant Cell 2005, 17, 760–775. [Google Scholar] [CrossRef]
- Pérez-Pascual, D.; Jiménez-Guillen, D.; Villanueva-Alonzo, H.; Souza-Perera, R.; Godoy-Hernández, G.; Zúñiga-Aguilar, J.J. Ectopic expression of the Coffea canephora SERK1 homolog-induced differential transcription of genes involved in auxin metabolism and in the developmental control of embryogenesis. Physiol. Plant 2018, 163, 530–551. [Google Scholar] [CrossRef]
- Tvorogova, V.E.; Lutova, L.A. Genetic regulation of zygotic embryogenesis in Angiosperm plants. Russ. J. Plant Physiol. 2018, 65, 1–14. [Google Scholar] [CrossRef]
- Zheng, Q.L.; Zheng, Y.M.; Ji, H.H.; Burnie, W.; Perry, S.E. Gene regulation by the AGL15 transcription factor reveals hormone interactions in somatic embryogenesis. Plant Physiol. 2016, 172, 2374–2387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.D. Observation and Transcriptome Analysis of Tree Peony Embryo Development. Master’s Thesis, Henan Agricultural University, Zhengzhou, China, 2019. [Google Scholar] [CrossRef]
- Honaas, L.; Kahn, E. A practical examination of RNA isolation methods for European pear (Pyrus communis). BMC Res. Notes 2017, 10, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sparkes, I.A.; Runions, J.; Kearns, A.; Hawes, C. Rapid, transient expression of fluorescent fusion proteins in tobacco plants and generation of stably transformed plants. Nat. Protoc. 2006, 1, 2019–2025. [Google Scholar] [CrossRef]
- Shen, P. The preliminary study on establishment of genetic transformation system of callus of Paeonia. Master’s Thesis, Henan Agricultural University, Zhengzhou, China, 2014. Available online: https://kns.cnki.net/KCMS/detail/detail.aspx?dbname=CMFD201801&filename=1017253701.nh (accessed on 20 June 2021).
- Kralik, P.; Ricchi, M. A basic guide to real time PCR in microbial diagnostics: Definitions, parameters, and everything. Front. Microbiol. 2017, 8, 108. [Google Scholar] [CrossRef] [Green Version]
- Forootan, A.; Sjöback, R.; Björkman, J.; Sjögreen, B.; Linz, L.; Kubista, M. Methods to determine limit of detection and limit of quantification in quantitative real-time PCR (qPCR). Biomol. Detect. Quantif. 2017, 12, 1–6. [Google Scholar] [CrossRef]
- Zimmerman, J.L. Somatic embryogenesis: A model for early development in higher plants. Plant Cell 1993, 5, 1411–1423. [Google Scholar] [CrossRef] [Green Version]
- Pasternak, T.P.; Prinsen, E.; Ayaydin, F.; Miskolczi, P.; Potters, G.; Asard, H.; Van Onckelen, H.A.; Dudits, D.; Fehér, A. The role of auxin, pH, and stress in the activation of embryogenic cell division in leaf protoplast-derived cells of alfalfa. Plant Physiol. 2002, 129, 1807–1819. [Google Scholar] [CrossRef] [Green Version]
- Rose, R.J. Somatic embryogenesis in the Medicago truncatula model: Cellular and molecular mechanisms. Front. Plant Sci. 2019, 10, 267. [Google Scholar] [CrossRef]
- Fehér, A. Somatic embryogenesis—stress-induced remodeling of plant cell fate. BBA-Gene Regul. Mech. 2015, 1849, 385–402. [Google Scholar] [CrossRef]
- Sweetlove, L.J.; Fernie, A.R. The spatial organization of metabolism within the plant cell. Annu. Rev. Plant Biol. 2013, 64, 723–746. [Google Scholar] [CrossRef] [PubMed]
- Renato, M.; Boronat, A.; Azcon-Bieto, J. Respiratory processes in non-photosynthetic plastids. Front. Plant Sci. 2015, 6, 496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths, C.A.; Paul, M.J.; Foyer, C.H. Metabolite transport and associated sugar signalling systems underpinning source/sink interactions. BBA-Gene Regul. Mech. 2016, 1857, 1715–1725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maillot, P.; Lebel, S.; Schellenbaum, P.; Jacques, A.; Walter, B. Differential regulation of SERK, LEC1-like and pathogenesis-related genes during indirect secondary somatic embryogenesis in grapevine. Plant Physiol. Biochem. 2009, 47, 743–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, P.; Joshi, S.; Tian, R.; Junior, R.D.; Chakrabarti, M.; Perry, S.E. The MADS-domain factor AGAMOUS-Like18 promotes somatic embryogenesis. Plant Physiol. 2022, 188, 1617–1631. [Google Scholar] [CrossRef]
- Jiménez, V.M.; Bangerth, F. Endogenous hormone concentrations and embryogenic callus development in wheat. Plant Cell Tissue Organ Cult. 2001, 67, 37–46. [Google Scholar] [CrossRef]
- Kępczyńska, E.; Orłowska, A. Profiles of endogenous ABA, bioactive GAs, IAA and their metabolites in Medicago truncatula Gaertn. non-embryogenic and embryogenic tissues during induction phase in relation to somatic embryo formation. Planta 2021, 253, 67. [Google Scholar] [CrossRef]
- Méndez-Hernández, H.A.; Ledezma-Rodriguez, M.; Avilez-Montalvo, R.N.; Juárez-Gómez, Y.L.; Skeete, A.O.; Avilez, J.; De-laPena, C.; Loyola-Vargas, V.M. Signaling overview of plant somatic embryogenesis. Front. Plant Sci. 2019, 10, 77. [Google Scholar] [CrossRef] [Green Version]
- Etienne, H.; Sotta, B.; Montoro, P.; Miginiac, E.; Carrona, M.P. Relations between exogenous growth regulators and endogenous indole-3-acetic acid and abscisic acid in the expression of somatic embryogenesis in Hevea brasiliensis (Müll. Arg.). Plant Sci. 1993, 88, 91–96. [Google Scholar] [CrossRef]
- Ivanova, A.; Velcheva, M.; Denchev, P.; Atanassov, A.; van Onckelen, H.A. Endogenous hormone levels during direct somatic embryogenesis in Medicago falcata. Physiol. Plant 1994, 92, 85–89. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Forward Primer Sequence (5′-3′) | Reverse Primer Sequence (5′-3′) |
|---|---|---|
| PoGPT1-Clone | ATGATTTGCTCTGCAAAACAAGC | TCACTGCTTTGCCTGGGAAT |
| PoGPT1-OE | ACGGGGGACTCTTGACCATGGATGATTTGCTCTGCAAAACAAGC | AAGTTCTTCTCCTTTACTAGTTCACTGCTTTGCCTGGGAAT |
| PoGPT1-GFP | GGGGTACCATGATTTGCTCTGCAAAACAAGC | GCTCTAGACTGCTTTGCCTGGGAATACAA |
| β-tubulin | TGAGCACCAAAGAAGTGGACGAAC | CACACGCCTGAACATCTCCTGAA |
| PoGPT1-RT | GCAGTTTCGTTCACCCATATCATCAAG | CAGCAAGAGCACATCCACCAATTATC |
| PoLEC1-RT | TCACGGCTGAGGACTTACTATGGG | TAATGTTCTCGGCACGCAAGGAAG |
| PoSERK-RT | GGCGAAGGAAACCACAAGAA | CCAACGAACCATCAGCAAGG |
| PoAGL15-RT | GGATAATTCACTGCCGTCTT | CCATGCCAATTTGACTTCC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Y.; Shang, W.; Wang, Z.; He, S.; Sun, Y.; Shi, L.; Shen, Y.; He, D. Functional Analysis of Glucose-6-Phosphate Translocator PoGPT1 in Embryogenic Callus Formation of Tree Peony. Horticulturae 2022, 8, 957. https://doi.org/10.3390/horticulturae8100957
Song Y, Shang W, Wang Z, He S, Sun Y, Shi L, Shen Y, He D. Functional Analysis of Glucose-6-Phosphate Translocator PoGPT1 in Embryogenic Callus Formation of Tree Peony. Horticulturae. 2022; 8(10):957. https://doi.org/10.3390/horticulturae8100957
Chicago/Turabian StyleSong, Yinglong, Wenqian Shang, Zheng Wang, Songlin He, Yuke Sun, Liyun Shi, Yuxiao Shen, and Dan He. 2022. "Functional Analysis of Glucose-6-Phosphate Translocator PoGPT1 in Embryogenic Callus Formation of Tree Peony" Horticulturae 8, no. 10: 957. https://doi.org/10.3390/horticulturae8100957