
Effects of Bacillus amyloliquefaciens QSB-6 on the Growth of Replanted Apple Trees and the Soil Microbial Environment

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Fertilizer Production
2.2. Test Materials
2.3. Field Experiment
2.3.1. Soil Physical and Chemical Properties
2.3.2. Microbial Culture Methods
2.3.3. Determination of Soil Enzyme Activities
2.3.4. Quantitative Determination of Soil Phenolic Acids by HPLC
2.3.5. DNA Extraction from Four Species of Fusarium and Quantitative PCR
2.3.6. Terminal-restriction fragment length polymorphism (T-RFLP) Analysis
2.4. Statistical Analysis
3. Results
3.1. Effect of Strain QSB-6 on the Biomass of Replanted Apple Saplings
3.2. Effect of Strain QSB-6 on Soil Microorganisms
3.3. Effect of Strain QSB-6 on Soil Enzyme Activities
3.4. Effect of Strain QSB-6 on Soil Phenolic Acids
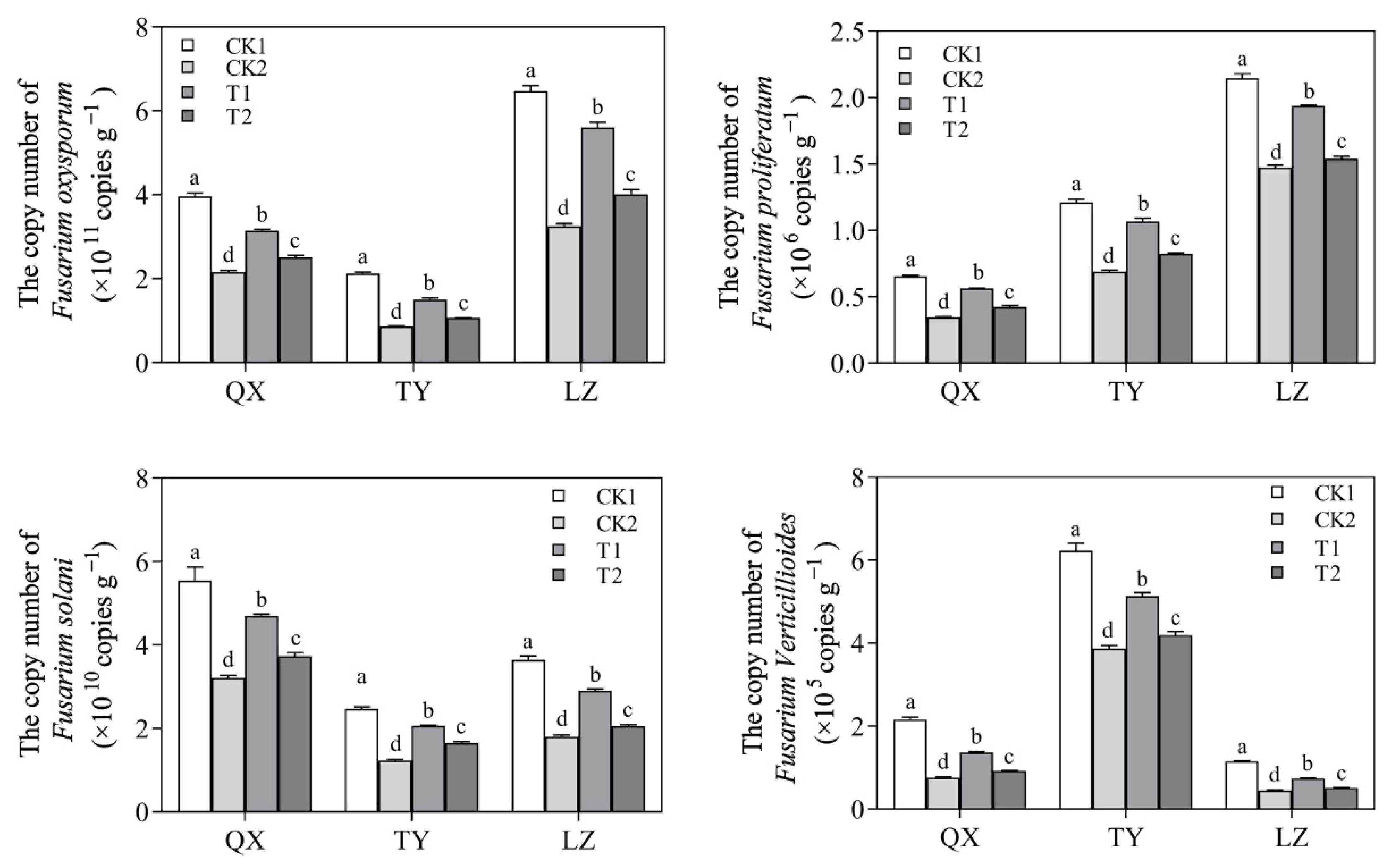
3.5. Inhibitory Effect of Strain QSB-6 on Four Species of Fusarium in Rhizosphere Soil
3.6. Effect of Strain QSB-6 on the Soil Microbial Community
4. Discussion
4.1. Effect of QSB-6 Fertilizer on the Soil Microbial Community
4.2. Effect of QSB-6 Fertilizer on Soil Phenolic Acids
4.3. Effect of QSB-6 Fertilizer Treatment on Soil Enzyme Activity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, X.S.; Han, M.Y.; Su, G.L.; Liu, F.Z.; Guo, G.N.; Jiang, Y.M.; Mao, Z.Q.; Peng, F.T.; Shu, H.R. Discussion on today’s world apple industry trends and the suggestions on sustainable and efficient development of apple industry in China. J. Fruit Sci. 2021, 27, 598–604. (In Chinese) [Google Scholar]
- Tewoldemedhin, Y.T.; Mazzola, M.; Labuschagne, I.; McLeod, A. A multi-phasic approach reveals that apple replant disease is caused by multiple biological agents, with some agents acting synergistically. Soil Biol. Biochem. 2011, 43, 1917–1927. [Google Scholar] [CrossRef]
- Tilston, E.L.; Deakin, G.; Bennett, J.; Passey, T.; Harrison, N.; Fernández, F.; Xu, X. Effect of fungal, oomycete and nematode interactions on apple root development in replant soil. CABI Agric. Biosci. 2020, 1, 14. [Google Scholar] [CrossRef]
- Balbín-Suárez, A.; Jacquiod, S.; Rohr, A.D.; Liu, B.; Flachowsky, H.; Winkelmann, T.; Smalla, K. Root exposure to apple replant disease soil triggers local defense response and rhizoplane microbiome dysbiosis. FEMS Microbiol. Ecol. 2021, 97, fiab031. [Google Scholar] [CrossRef] [PubMed]
- Bezemer, T.M.; Lawson, C.S.; Hedlund, K.; Edwards, A.R.; Brook, A.J.; Igual, J.M.; Van der Putten, W.H. Plant species and functional group effects on abiotic and microbial soil properties and plant-soil feedback responses in two grasslands. J. Ecol. 2006, 94, 893–904. [Google Scholar] [CrossRef]
- Zhang, B.; Li, X.; Wang, F.; Li, M.; Zhang, J.; Gu, L.; Zhang, Z. Assaying the potential autotoxins and microbial community associated with Rehmannia glutinosa replant problems based on its ‘autotoxic circle’. Plant Soil 2016, 407, 307–322. [Google Scholar] [CrossRef] [Green Version]
- Cavael, U.; Lentzsch, P.; Schwärzel, H.; Eulenstein, F.; Tauschke, M.; Diehl, K. Assessment of Agro-Ecological Apple Replant Disease (ARD) Management Strategies: Organic Fertilisation and Inoculation with Mycorrhizal Fungi and Bacteria. Agronomy 2021, 11, 272. [Google Scholar] [CrossRef]
- Hofmann, A.; Wittenmayer, L.; Arnold, G.; Schieber, A.; Merbach, W. Root exudation of phloridzin by apple seedlings (Malus×domestica Borkh.) with symptoms of apple replant disease. J. Appl. Bot. Food Qual. 2009, 82, 193–198. [Google Scholar]
- Kelderer, M.; Manici, L.M.; Caputo, F.; Thalheimer, M. Planting in the ‘inter-row’to overcome replant disease in apple orchards: A study on the effectiveness of the practice based on microbial indicators. Plant Soil 2012, 357, 381–393. [Google Scholar] [CrossRef]
- Van Schoor, L.; Denman, S.; Cook, N.C. Characterisation of apple replant disease under South African conditions and potential biological management strategies. Sci. Hortic. 2009, 119, 153–162. [Google Scholar] [CrossRef]
- Singh, N.; Sharma, D.P.; Kaushal, R. Controlling replant disease of apple in Himachal Pradesh, India by rootstocks and soil agro-techniques. Pharma Innov. J. 2017, 6, 288–293. [Google Scholar]
- Leinfelder, M.M.; Merwin, I.A.; Fazio, G.; Robinson, T. Resistant rootstocks, preplant compost amendments, soil fumigation, and row repositioning for managing apple replant disease. HortScience 2004, 39, 841. [Google Scholar] [CrossRef] [Green Version]
- Garbeva, P.V.; Van Veen, J.A.; Van Elsas, J.D. Microbial diversity in soil: Selection of microbial populations by plant and soil type and implications for disease suppressiveness. Annu. Rev. Phytopathol. 2004, 42, 243–270. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.F.; Wang, H.Y.; Wang, M.; Li, H.; Xiang, L.; Pan, F.B.; Mao, Z.Q. Effects of soil texture on the growth of young apple trees and soil microbial community structure under replanted conditions. Hortic. Plant J. 2020, 6, 123–131. [Google Scholar] [CrossRef]
- Wang, G.S.; Yin, C.M.; Pan, F.B.; Wang, X.B.; Xiang, L.; Wang, Y.F.; Mao, Z.Q. Analysis of the fungal community in apple replanted soil around Bohai Gulf. Hortic. Plant J. 2018, 4, 175–181. [Google Scholar] [CrossRef]
- Liu, Z. The Isolation and Identification of Fungi Pathogen from Apple Replant Disease in Bohai Bay Region and the Screening of the Antagonistic Trichoderma Strains. Master’s Thesis, Shandong Agricultural University, Tai’an, China, 2013. [Google Scholar]
- Yin, C.M.; Xiang, L.; Wang, G.S.; Wang, Y.F.; Shen, X.; Chen, X.S.; Mao, Z.Q. Phloridzin promotes the growth of Fusarium moniliforme (Fusarium verticillioides). Sci. Hortic. 2017, 214, 187–194. [Google Scholar] [CrossRef]
- Duan, Y.N.; Chen, R.; Zhang, R.; Jiang, W.T.; Chen, X.S.; Yin, C.M.; Mao, Z.Q. Isolation, identification, and antibacterial mechanisms of Bacillus amyloliquefaciens QSB-6 and its effect on plant roots. Front. Microbiol. 2021, 12, 2727. [Google Scholar] [CrossRef]
- Trapero Casas, A.; Jimenez-Diaz, R.M. Fungal wilt and root rot disease of chickpea in Southern Spain. Phytopathology 1985, 75, 1146–1151. [Google Scholar] [CrossRef] [Green Version]
- Anjaiah, V.; Cornelis, P.; Koedam, N. Effect of genotype and root colonization in biological control of Fusarium wilts in pigeonpea and chickpea by Pseudomonas aeruginosa PNA1. Can. J. Microbiol. 2003, 49, 85–91. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Beale, M.H.; Ward, J.L.; Strange, R.N. Chickpea wilt: Identification and toxicity of 8-O-methyl-fusarubin from Fusarium acutatum. Photochemistry 2005, 66, 1536–1539. [Google Scholar] [CrossRef] [PubMed]
- Clements, J.; Cowgill, W.; Embree, C.; Gonzalez, V.; Hoying, S.; Kushad, M.; Autio, W. Rootstock Tolerance to Apple Replant Disease for Improved Sustainability of Apple Production. In Proceedings of the XXVIII International Horticultural Congress on Science and Horticulture for People (IHC2010): International Symposium on the Challenge for a Sustainable Production, Protection and Consumption of Medit, Lisbon, Portugal, 22–27 August 2010; pp. 521–528. [Google Scholar]
- Wang, Y.F.; Pan, F.B.; Wang, G.S.; Zhang, G.; Wang, Y.; Chen, X.S.; Mao, Z.Q. Effects of biochar on photosynthesis and antioxidative system of Malus hupehensis Rehd. seedlings under replant conditions. Sci. Hortic. 2014, 175, 9–15. [Google Scholar] [CrossRef]
- Winkelmann, T.; Smalla, K.; Amelung, W.; Baab, G.; Grunewaldt-Stöcker, G.; Kanfra, X.; Schloter. M. Apple replant disease: Causes and mitigation strategies. Curr. Issues Mol. Biol. 2018, 30, 89–106. [Google Scholar]
- Raymaekers, K.; Ponet, L.; Holtappels, D.; Berckmans, B.; Cammue, B.P. Screening for novel biocontrol agents applicable in plant disease management—A review. Biol. Control 2020, 144, 104240. [Google Scholar] [CrossRef]
- Chowdhury, S.K.; Majumdar, S.; Mandal, V. Application of Bacillus sp. LBF-01 in Capsicum annuum plant reduces the fungicide use against Fusarium oxysporum. Biocatal. Agric. Biotechnol. 2020, 27, 101714. [Google Scholar] [CrossRef]
- Azabou, M.C.; Gharbi, Y.; Medhioub, I.; Ennouri, K.; Barham, H.; Tounsi, S.; Triki, M.A. The endophytic strain Bacillus velezensis OEE1: An efficient biocontrol agent against Verticillium wilt of olive and a potential plant growth promoting bacteria. Biol. Control 2020, 142, 104168. [Google Scholar] [CrossRef]
- Bubici, G.; Kaushal, M.; Prigigallo, M.I.; Gómez-Lama Cabanás, C.; Mercado-Blanco, J. Biological control agents against Fusarium wilt of banana. Front. Microbiol. 2019, 10, 616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, G.K.H.; Wang, L.; Chen, J.; Ji, P. Biological control of Fusarium wilt on watermelon by fluorescent pseudomonads. Biocontrol Sci. Technol. 2020, 30, 212–227. [Google Scholar] [CrossRef]
- Patil, S.; Sriram, S. Biological control of Fusarium wilt in crop plants using non-pathogenic isolates of Fusarium species. Indian Phytopathol. 2020, 73, 11–19. [Google Scholar] [CrossRef]
- Moutassem, D.; Belabid, L.; Bellik, Y. Efficiency of secondary metabolites produced by Trichoderma spp. in the biological control of Fusarium wilt in chickpea. J. Crop Prot. 2020, 9, 217–231. [Google Scholar]
- Khedher, S.B.; Mejdoub-Trabelsi, B.; Tounsi, S. Biological potential of Bacillus subtilis V26 for the control of Fusarium wilt and tuber dry rot on potato caused by Fusarium species and the promotion of plant growth. Biol. Control 2021, 152, 104444. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, R.; Zhao, L.; Wang, H.; Chen, X.; Mao, Z.; Yin, C. Indigenous arbuscular mycorrhizal fungi enhance resistance of apple rootstock ‘M9T337’to apple replant disease. Physiol. Mol. Plant Pathol. 2021, 116, 101717. [Google Scholar] [CrossRef]
- Ab Rahman, S.F.S.; Singh, E.; Pieterse, C.M.; Schenk, P.M. Emerging microbial biocontrol strategies for plant pathogens. Plant Sci. 2018, 267, 102–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumbar, B.; Mahmood, R.; Nagesha, S.N.; Nagaraja, M.S.; Prashant, D.G.; Kerima, O.Z.; Chavan, M. Field application of Bacillus subtilis isolates for controlling late blight disease of potato caused by Phytophthora infestans. Biocatal. Agric. Biotechnol. 2019, 22, 101366. [Google Scholar] [CrossRef]
- Haddoudi, I.; Cabrefiga, J.; Mora, I.; Mhadhbi, H.; Montesinos, E.; Mrabet, M. Biological control of Fusarium wilt caused by Fusarium equiseti in Vicia faba with broad spectrum antifungal plant-associated Bacillus spp. Biol. Control 2021, 160, 104671. [Google Scholar] [CrossRef]
- Zheng, T.W.; Liu, L.; Nie, Q.W.; Hsiang, T.; Sun, Z.X.; Zhou, Y. Isolation, identification and biocontrol mechanisms of endophytic bacterium D61-A from Fraxinus hupehensis against Rhizoctonia solani. Biol. Control 2021, 158, 104621. [Google Scholar] [CrossRef]
- Anusha, B.G.; Gopalakrishnan, S.; Naik, M.K.; Sharma, M. Evaluation of Streptomyces spp. and Bacillus spp. for biocontrol of Fusarium wilt in chickpea (Cicer arietinum L.). Arch. Phytopathol. Plant Prot. 2019, 52, 417–442. [Google Scholar] [CrossRef]
- Toju, H.; Peay, K.G.; Yamamichi, M.; Narisawa, K.; Hiruma, K.; Naito, K.; Kiers, E.T. Core microbiomes for sustainable agroecosystems. Nat. Plants 2018, 4, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Tao, C.; Li, R.; Xiong, W.; Shen, Z.; Liu, S.; Wang, B.; Kowalchuk, G.A. Bio-organic fertilizers stimulate indigenous soil Pseudomonas populations to enhance plant disease suppression. Microbiome 2020, 8, 137. [Google Scholar] [CrossRef]
- MAFF; ADAS. The analysis of agricultural materials. In Reference Book 427: Methods 32 and 56; Ministry of Agriculture, Fisheries and Food: London, UK; Her Majesty’s Stationery Office: London, UK, 1986. [Google Scholar]
- Avery, B.W. Soil classification in the Soil Survey of England and Wales. J. Soil Sci. 1973, 243, 324–338. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhu, L.; Wang, J.; Xie, H.; Wang, J.; Wang, F.; Sun, F. Effects of fomesafen on soil enzyme activity, microbial population, and bacterial community composition. Environ. Monit. Assess. 2014, 186, 2801–2812. [Google Scholar] [CrossRef]
- Yin, C.M.; Wang, G.S.; Li, Y.Y.; Che, J.S.; Shen, X.; Chen, X.S.; Wu, S.J. A new method for analysis of phenolic acids in the soil-soil from replanted apple orchards was investigated. China Agric. Sci. 2013, 46, 4612–4619. (In Chinese) [Google Scholar]
- Xiang, L.; Wang, M.; Jiang, W.T.; Wang, Y.F.; Chen, X.S.; Yin, C.M.; Mao, Z.Q. Key indicators for renewal and reconstruction of perennial trees soil: Microorganisms and phloridzin. Ecotoxicol. Environ. Saf. 2021, 225, 112723. [Google Scholar] [CrossRef]
- Whelan, J.A.; Russell, N.B.; Whelan, M.A. A method for the absolute quantification of cDNA using real-time PCR. J. Immunol. Methods 2003, 278, 261–269. [Google Scholar] [CrossRef]
- Xu, H.J.; Bai, J.; Li, W.Y.; Zhao, L.X.; Li, Y.T. Removal of persistent DDT residues from soils by earthworms: A mechanistic study. J. Hazard. Mater. 2019, 365, 622–631. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.Y.; Wang, Y.K.; Chen, P.C.; Li, F.B.; Chen, M.J.; Hu, M. The effect of ammonium chloride and urea application on soil bacterial communities closely related to the reductive transformation of pentachlorophenol. J. Hazard. Mater. 2014, 272, 10–19. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, J.; Zhu, L.; Du, Z.; Wang, J.; Sun, X.; Zhou, T. Effects of 1-octyl-3-methylimidazolium nitrate on the microbes in brown soil. J. Environ. Sci. 2018, 67, 249–259. [Google Scholar] [CrossRef]
- Shyu, C.; Soule, T.; Bent, S.J.; Foster, J.A.; Forney, L.J. MiCA: A web-based tool for the analysis of microbial communities based on terminal-restriction fragment length polymorphisms of 16S and 18S rRNA genes. Microb. Ecol. 2007, 53, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Pan, F.; Han, X.; Song, F.; Zhang, Z.; Yan, J.; Xu, Y. Response of soil fungal community structure to long-term continuous soybean cropping. Front. Microbiol. 2019, 9, 3316. [Google Scholar] [CrossRef]
- Xu, F.; Chen, Y.; Cai, Y.; Gu, F.; An, K. Distinct roles for bacterial and fungal communities during the curing of vanilla. Front. Microbiol. 2020, 11, 2342. [Google Scholar] [CrossRef] [PubMed]
- Perkins, J.L. A Study of Diversity and Community Comparison Indices by Bioassay of Copper Using Colonized Benthic Macroinvertebrates; The University of Texas School of Public Health: Houston, TX, USA, 1981. [Google Scholar]
- Li, H.L.; Qiang, S.W. Mitigation of replant disease by mycorrhization in horticultural plants: A review. Folia Hortic. 2018, 30, 269–282. [Google Scholar] [CrossRef]
- Ma, Z.; Wang, Q.; Wang, X.; Chen, X.; Wang, Y.; Mao, Z. Effects of Biochar on Replant Disease by Amendment Soil Environment. Commun. Soil Sci. Plant Anal. 2021, 52, 673–685. [Google Scholar] [CrossRef]
- Di Prima, S.; Rodrigo-Comino, J.; Novara, A.; Iovino, M.; Pirastru, M.; Keesstra, S.; Cerdà, A. Soil physical quality of citrus orchards under tillage, herbicide, and organic managements. Pedosphere 2018, 28, 463–477. [Google Scholar] [CrossRef] [Green Version]
- Adekiya, A.O.; Ejue, W.S.; Olayanju, A.; Dunsin, O.; Aboyeji, C.M.; Aremu, C.; Akinpelu, O. Different organic manure sources and NPK fertilizer on soil chemical properties, growth, yield and quality of okra. Sci. Rep. 2020, 10, 16083. [Google Scholar] [CrossRef]
- Hill, P.; Krištůfek, V.; Dijkhuizen, L.; Boddy, C.; Kroetsch, D.; van Elsas, J.D. Land use intensity controls actinobacterial community structure. Microb. Ecol. 2011, 61, 286–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatti, A.A.; Haq, S.; Bhat, R.A. Actinomycetes benefaction role in soil and plant health. Microb. Pathog. 2017, 111, 458–467. [Google Scholar] [CrossRef]
- Caputo, F.; Nicoletti, F.; Picione, F.D.L.; Manici, L.M. Rhizospheric changes of fungal and bacterial communities in relation to soil health of multi-generation apple orchards. Biol. Control 2015, 88, 8–17. [Google Scholar] [CrossRef]
- Franke-Whittle, I.H.; Manici, L.M.; Insam, H.; Stres, B. Rhizosphere bacteria and fungi associated with plant growth in soils of three replanted apple orchards. Plant Soil 2015, 395, 317–333. [Google Scholar] [CrossRef]
- Spath, M.; Insam, H.; Peintner, U.; Kelderer, M.; Kuhnert, R.; Franke-Whittle, I.H. Linking soil biotic and abiotic factors to apple replant disease: A greenhouse approach. J. Phytopathol. 2015, 163, 287–299. [Google Scholar] [CrossRef]
- Bai, Y.; Wang, G.; Cheng, Y.; Shi, P.; Yang, C.; Yang, H.; Xu, Z. Soil acidification in continuously cropped tobacco alters bacterial community structure and diversity via the accumulation of phenolic acids. Sci. Rep. 2019, 9, 12499. [Google Scholar] [CrossRef] [Green Version]
- Liang, B.W.; Ma, C.Q.; Fan, L.M.; Wang, Y.Z.; Yuan, Y.B. Soil amendment alters soil physicochemical properties and bacterial community structure of a replanted apple orchard. Microbiol. Res. 2018, 216, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Ma, C.; Fan, L.; Wang, Y.; Yuan, Y. Compost amendment alters soil fungal community structure of a replanted apple orchard. Arch. Agron. Soil Sci. 2021, 67, 739–752. [Google Scholar] [CrossRef]
- Zhao, W.S.; Guo, Q.G.; Su, Z.H.; Wang, P.P.; Dong, L.H.; Hu, Q.; Lu, X.Y.; Zhang, X.Y.; Li, S.Z.; Ma, P. Rhizosphere soil fungal community structure of healthy potato plants and Verticillium dahliae and their utilization characteristics of carbon sources. Sci. Agric. Sin. 2021, 54, 296–309. [Google Scholar]
- Gadhave, K.R.; Devlin, P.F.; Ebertz, A.; Ross, A.; Gange, A.C. Soil inoculation with Bacillus spp. modifies root endophytic bacterial diversity, evenness, and community composition in a context-specific manner. Microb. Ecol. 2018, 76, 741–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Z.; Huang, J.; Yang, C.; Xu, Y.; Shen, Q.; Chen, W. Screening of suitable carriers for Bacillus amyloliquefaciens strain QL-18 to enhance the biocontrol of tomato bacterial wilt. Crop Prot. 2015, 75, 96–103. [Google Scholar] [CrossRef]
- Malusá, E.; Sas-Paszt, L.; Ciesielska, J. Technologies for beneficial microorganisms inocula used as biofertilizers. Sci. World J. 2012, 2012, 491206. [Google Scholar] [CrossRef]
- Smith, R.S. Legume inoculant formulation and application. Can. J. Microbiol. 1992, 38, 485–492. [Google Scholar] [CrossRef]
- Vijayaraghavan, P.; Arun, A.; Al-Dhabi, N.A.; Vincent, S.G.P.; Arasu, M.V.; Choi, K.C. Novel Bacillus subtilis IND19 cell factory for the simultaneous production of carboxy methyl cellulase and protease using cow dung substrate in solid-substrate fermentation. Biotechnol. Biofuels 2016, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Zhang, Z.; Ling, N.; Yuan, Y.; Zheng, X.; Shen, B.; Shen, Q. Bacillus subtilis SQR 9 can control Fusarium wilt in cucumber by colonizing plant roots. Biol. Fertil. Soils 2011, 47, 495–506. [Google Scholar] [CrossRef]
- Weller, D.M. Pseudomonas biocontrol agents of soilborne pathogens looking back over 30 years. Phytopathology 2007, 97, 250–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazurier, S.; Corberand, T.; Lemanceau, P.; Raaijmakers, J.M. Phenazine antibiotics produced by fluorescent pseudomonads contribute to natural soil suppressiveness to fusarium wilt. ISME J 2009, 3, 977–991. [Google Scholar] [CrossRef]
- Mendes, R.; Kruijt, M.; De Bruijn, I.; Dekkers, E.; Van der Voort, M.; Schneider, J.H.; Raaijmakers, J.M. Deciphering the rhizosphere microbiome for disease-suppressive bacteria. Science 2011, 332, 1097–1100. [Google Scholar] [CrossRef]
- Shen, Z.; Wang, D.; Ruan, Y.; Xue, C.; Zhang, J.; Li, R.; Shen, Q. Deep 16S rRNA pyrosequencing reveals a bacterial community associated with banana Fusarium wilt disease suppression induced by bio-organic fertilizer application. PLoS ONE 2014, 9, e98420. [Google Scholar]
- Qin, S.; Yeboah, S.; Cao, L.; Zhang, J.; Shi, S.; Liu, Y. Breaking continuous potato cropping with legumes improves soil microbial communities, enzyme activities and tuber yield. PLoS ONE 2017, 12, e0175934. [Google Scholar] [CrossRef]
- Liu, H.; Li, J.; Carvalhais, L.C.; Percy, C.D.; Prakash Verma, J.; Schenk, P.M.; Singh, B.K. Evidence for the plant recruitment of beneficial microbes to suppress soil-borne pathogens. New Phytol. 2021, 229, 2873–2885. [Google Scholar] [CrossRef]
- Li, R.; Shen, Z.; Sun, L.; Zhang, R.; Fu, L.; Deng, X.; Shen, Q. Novel soil fumigation method for suppressing cucumber Fusarium wilt disease associated with soil microflora alterations. Appl. Soil Ecol. 2016, 101, 28–36. [Google Scholar] [CrossRef]
- Gao, X.B.; Zhao, F.X.; Xiang, S.H.E.N.; Hu, Y.L.; Hao, Y.H.; Yang, S.Q.; Mao, Z.Q. Effects of cinnamon acid on respiratory rate and its related enzymes activity in roots of seedlings of Malus hupehensis Rehd. Agric. Sci. China 2020, 9, 833–839. [Google Scholar] [CrossRef]
- Yin, C.; Xiang, L.; Wang, G.; Wang, Y.; Shen, X.; Chen, X.; Mao, Z. How to plant apple trees to reduce replant disease in apple orchard: A study on the phenolic acid of the replanted apple orchard. PLoS ONE 2016, 11, e0167347. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Jin, Y.; Zhu, W.; Tang, J.; Hu, S.; Zhou, T.; Chen, X. Baicalin released from Scutellaria baicalensis induces autotoxicity and promotes soilborn pathogens. J. Chem. Ecol. 2011, 36, 329–338. [Google Scholar] [CrossRef]
- Zhou, X.; Yu, G.; Wu, F. Responses of soil microbial communities in the rhizosphere of cucumber (Cucumis sativus L.) to exogenously applied p-hydroxybenzoic acid. J. Chem. Ecol. 2012, 38, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Hannula, S.E.; Morriën, E.; De Hollander, M.; Van Der Putten, W.H.; Van Veen, J.A.; De Boer, W. Shifts in rhizosphere fungal community during secondary succession following abandonment from agriculture. ISME J. 2017, 11, 2294–2304. [Google Scholar] [CrossRef]
- Poonguzhali, S.; Madhaiyan, M.; Sa, T. Cultivation-dependent characterization of rhizobacterial communities from field grown Chinese cabbage Brassica campestris ssp pekinensis and screening of traits for potential plant growth promotion. Plant Soil 2006, 286, 167–180. [Google Scholar] [CrossRef]
- Song, Y.; Song, C.; Yang, G.; Miao, Y.; Wang, J.; Guo, Y. Changes in labile organic carbon fractions and soil enzyme activities after marshland reclamation and restoration in the Sanjiang Plain in Northeast China. Environ. Manag. 2012, 50, 418–426. [Google Scholar] [CrossRef]
- Stpniewska, Z.; Wolińska, A.; Ziomek, J. Response of soil catalase activity to chromium contamination. J. Environ. Sci. 2009, 21, 1142–1147. [Google Scholar] [CrossRef]
- Hu, B.; Liang, D.; Liu, J.; Lei, L.; Yu, D. Transformation of heavy metal fractions on soil urease and nitrate reductase activities in copper and selenium co-contaminated soil. Ecotoxicol. Environ. Saf. 2014, 110, 41–48. [Google Scholar] [CrossRef]
- Wei, K.; Chen, Z.; Zhu, A.; Zhang, J.; Chen, L. Application of 31p NMR spectroscopy in determining phosphatase activities and p composition in soil aggregates influenced by tillage and residue management practices. Soil Tillage Res. 2014, 138, 35–43. [Google Scholar] [CrossRef]
- Yu, P.; Liu, S.; Han, K.; Guan, S.; Zhou, D. Conversion of cropland to forage land and grassland increases soil labile carbon and enzyme activities in northeastern China. Agric. Ecosyst. Environ. 2017, 245, 83–91. [Google Scholar] [CrossRef]
- Sabaté, D.C.; Petroselli, G.; Erra-Balsells, R.; Audisio, M.C.; Brandan, C.P. Beneficial effect of Bacillus sp. P12 on soil biological activities and pathogen control in common bean. Biol. Control 2020, 141, 104131. [Google Scholar] [CrossRef]
- Zhang, X.; Gao, J.; Zhao, F.; Zhao, Y.; Li, Z. Characterization of a salt-tolerant bacterium Bacillus sp. from a membrane bioreactor for saline wastewater treatment. J. Environ. Sci. 2014, 26, 1369–1374. [Google Scholar] [CrossRef]
- Rawat, J.; Sanwal, P.; Saxena, J. Potassium and its role in sustainable agriculture. In Potassium Solubilizing Microorganisms for Sustainable Agriculture; Springer: New Delhi, India, 2016; pp. 235–253. [Google Scholar]
- Yadav, R.; Ror, P.; Rathore, P.; Kumar, S.; Ramakrishna, W. Bacillus subtilis CP4, isolated from native soil in combination with arbuscular mycorrhizal fungi promotes biofortification, yield and metabolite production in wheat under field conditions. J. Appl. Microbiol. 2021, 131, 339–359. [Google Scholar] [CrossRef] [PubMed]
- Iovieno, P.; Morra, L.; Leone, A.; Pagano, L.; Alfani, A. Effect of organic and mineral fertilizers on soil respiration and enzyme activities of two Mediterranean horticultural soils. Biol. Fertil. Soils 2009, 45, 555–561. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Place | Ammonium-Nitrogen (mg kg−1) | Nitrate Nitrogen (mg kg−1) | Organic Matter (%) | Available Phosphorus (mg kg−1) | Soil Bulk Density (g cm−3) | Available Potassium (mg kg−1) | Soil pH | Soil Moisture Content (%) |
|---|---|---|---|---|---|---|---|---|
| LZ | 1.5 ± 0.0 b | 39.2 ± 1.1 b | 2.2 ± 0.1 a | 17.5 ± 0.0 c | 1.0 ± 0.1 a | 63.2 ± 8.8 c | 7.1 ± 0.0 a | 8.9 ± 0.2 a |
| QX | 1.6 ± 0.0 b | 42.8 ± 0.2 a | 2.2 ± 0.0 a | 19.2 ± 0.3 b | 1.2 ± 0.1 a | 108.4 ± 5.0 b | 6.7 ± 0.3 a | 10.3 ± 1.7 a |
| TY | 2.3 ± 0.2 a | 43.7 ± 0.2 a | 2.3 ± 0.0 a | 21.7 ± 0.5 a | 1.1 ± 0.1 a | 223.3 ± 11.7 a | 7.1 ± 0.0 a | 11.5 ± 1.1 a |
| Place | Soil Treatments | Plant Height (cm) | Ground Diameter (mm) | Numbers of Branches | Branch Length (cm) |
|---|---|---|---|---|---|
| LZ | CK1 | 150.7 ± 3.0 c | 13.8±0.1 c | 7.7 ± 0.9 b | 44.5 ± 6.6 c |
| CK2 | 206.0 ± 7.6 a | 24.3±1.1 a | 11.0 ± 0.6 a | 73.8 ± 2.4 a | |
| T1 | 179.3 ± 0.9 b | 18.4±0.9 b | 7.7 ± 0.7 b | 54.3 ± 17 bc | |
| T2 | 188.0 ± 0.6 b | 20.2±0.9 b | 10.0 ± 0.6 ab | 58.0 ± 3.1 b | |
| QX | CK1 | 139.0 ± 5.9 c | 18.0 ± 0.1 d | 8.7 ± 1.5 b | 34.0 ± 1.7 b |
| CK2 | 191.3 ± 6.3 a | 28.4 ± 0.6 a | 15.0 ± 1.0 a | 53.6 ± 2.7 a | |
| T1 | 165.7 ± 2.3 b | 23.1 ± 0.0 c | 11.7 ± 1.2 ab | 38.1 ± 1.6 b | |
| T2 | 180.7 ± 1.7 a | 25.6 ± 1.0 b | 12.0 ± 1.5 ab | 48.8 ± 3.0 a | |
| TY | CK1 | 191.7 ± 12.1 c | 17.0 ± 1.5 b | 10.0 ± 1.5 b | 49.1 ± 4.7 b |
| CK2 | 254.3 ± 4.3 a | 25.8 ± 0.1 a | 16.3 ± 0.7 a | 68.0 ± 5.6 a | |
| T1 | 229.3 ± 0.7 b | 19.1 ± 0.2 b | 12.0 ± 1.0 ab | 55.7 ± 3.8 ab | |
| T2 | 251.7 ± 4.9 a | 23.5 ± 0.6 a | 15.0 ± 2.1 a | 66.1 ± 1.1 a |
| Place | Soil Treatments | The Number of Soil Bacteria (×105 CFU/g Soil) | The Number of Soil Fungi (×103 CFU/g Soil) | The Number of Soil Actinomycete (×104 CFU/g Soil) | The Ratio of Bacteria and Fungi | The Ratio of Actinomycete and Fungi |
|---|---|---|---|---|---|---|
| LZ | CK1 | 40.3 ± 1.5 b | 52.0 ± 2.1 a | 60.7 ± 3.5 c | 78.0 ± 5.8 b | 11.7 ± 1.0 c |
| CK2 | 25.7 ± 1.2 c | 31.0 ± 0.6 c | 52.0 ± 1.0 c | 82.7 ± 2.4 b | 16.8 ± 0.4b c | |
| T1 | 46.7 ± 1.5 b | 44.7 ± 1.8 b | 91.0 ± 1.2 b | 104.7 ± 3.7 b | 20.4 ± 0.9 b | |
| T2 | 89.0 ± 3.8 a | 34.0 ± 2.5 c | 140.3 ± 4.2 a | 266.4 ± 31.7 a | 41.9 ± 4.2 a | |
| QX | CK1 | 38.0 ± 1.5 c | 68.7 ± 0.9 a | 92.3 ± 1.2 b | 55.4 ± 3.0 c | 13.5 ± 0.1 c |
| CK2 | 27.0 ± 3.0 d | 21.7 ± 0.9 d | 75.0 ± 2.3 c | 123.9 ± 9.6 b | 34.6 ± 0.4 b | |
| T1 | 63.6 ± 1.8 b | 58.0 ± 2.1 b | 102.7 ± 4.1 b | 110.2 ± 6.0 b | 17.78 ± 1.2 c | |
| T2 | 107.3 ± 1.9 a | 32.0 ± 1.2 c | 154.7 ± 6.4 a | 336.7 ± 17.9 a | 48.54 ± 3.2 a | |
| TY | CK1 | 56.0 ± 0.6 c | 53.7 ± 2.0 a | 85.0 ± 2.5 c | 104.6 ± 3.7 c | 15.9 ± 0.3 b |
| CK2 | 24.3 ± 0.9 d | 27.3 ± 0.9 c | 65.3 ± 2.7 c | 89.0 ± 0.4 d | 24.0 ± 1.7 b | |
| T1 | 66.0 ± 2.1 b | 48.0 ± 1.2 b | 114.3 ± 7.7 b | 137.5 ± 3.1 b | 23.9 ± 2.2 b | |
| T2 | 114.7 ± 3.7 a | 31.3 ± 0.9 c | 169.0 ± 9.3 a | 365.9 ± 2.5 a | 54.1 ± 4.0 a |
| Microorganism | Treatment | Margalef’s Index | Shannon’s Index | Mcintosh’s Index | Brillouin’s Index |
|---|---|---|---|---|---|
| Fungi | CK1 | 10.3 ± 0.1 b | 3.6 ± 0.0 ab | 16.6 ± 0.8 a | 3.0 ± 0.0 a |
| CK2 | 9.9 ± 0.7 b | 3.5 ± 0.1 ab | 8.7 ± 0.9 c | 2.7 ± 0.1 bc | |
| T1 | 12.2 ± 0.2 a | 3.7 ± 0.0 a | 11.9 ± 0.4 b | 2.9 ± 0.1 ab | |
| T2 | 11.7 ± 0.3 a | 3.4 ± 0.1 b | 8.5 ± 0.6 c | 2.5 ± 0.1 c | |
| Bacterial | CK1 | 11.0 ± 0.2 b | 3.5 ± 0.0 b | 12.420.6 b | 2.8 ± 0.0 c |
| CK2 | 9.6 ± 0.1 d | 3.3 ± 0.0 c | 12.6 ± 0.5 b | 2.6 ± 0.0 d | |
| T1 | 12.1 ± 0.1 a | 3.7 ± 0.0 a | 12.6 ± 0.4 b | 3.0 ± 0.0 b | |
| T2 | 10.5 ± 0.0 c | 3.7 ± 0.0 a | 25.5 ± 1.2 a | 3.3 ± 0.0 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duan, Y.; Zhou, Y.; Li, Z.; Chen, X.; Yin, C.; Mao, Z. Effects of Bacillus amyloliquefaciens QSB-6 on the Growth of Replanted Apple Trees and the Soil Microbial Environment. Horticulturae 2022, 8, 83. https://doi.org/10.3390/horticulturae8010083
Duan Y, Zhou Y, Li Z, Chen X, Yin C, Mao Z. Effects of Bacillus amyloliquefaciens QSB-6 on the Growth of Replanted Apple Trees and the Soil Microbial Environment. Horticulturae. 2022; 8(1):83. https://doi.org/10.3390/horticulturae8010083
Chicago/Turabian StyleDuan, Yanan, Yifan Zhou, Zhao Li, Xuesen Chen, Chengmiao Yin, and Zhiquan Mao. 2022. "Effects of Bacillus amyloliquefaciens QSB-6 on the Growth of Replanted Apple Trees and the Soil Microbial Environment" Horticulturae 8, no. 1: 83. https://doi.org/10.3390/horticulturae8010083