
SNPs Associated with Foliar Phylloxera Tolerance in Hybrid Grape Populations Carrying Introgression from Muscadinia

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Laboratory Assessment of Resistance to the Leaf Form of Phylloxera
2.3. SSR and Target SNP Marker Analysis
2.4. Genome Wide Association Study (GWAS)
3. Results
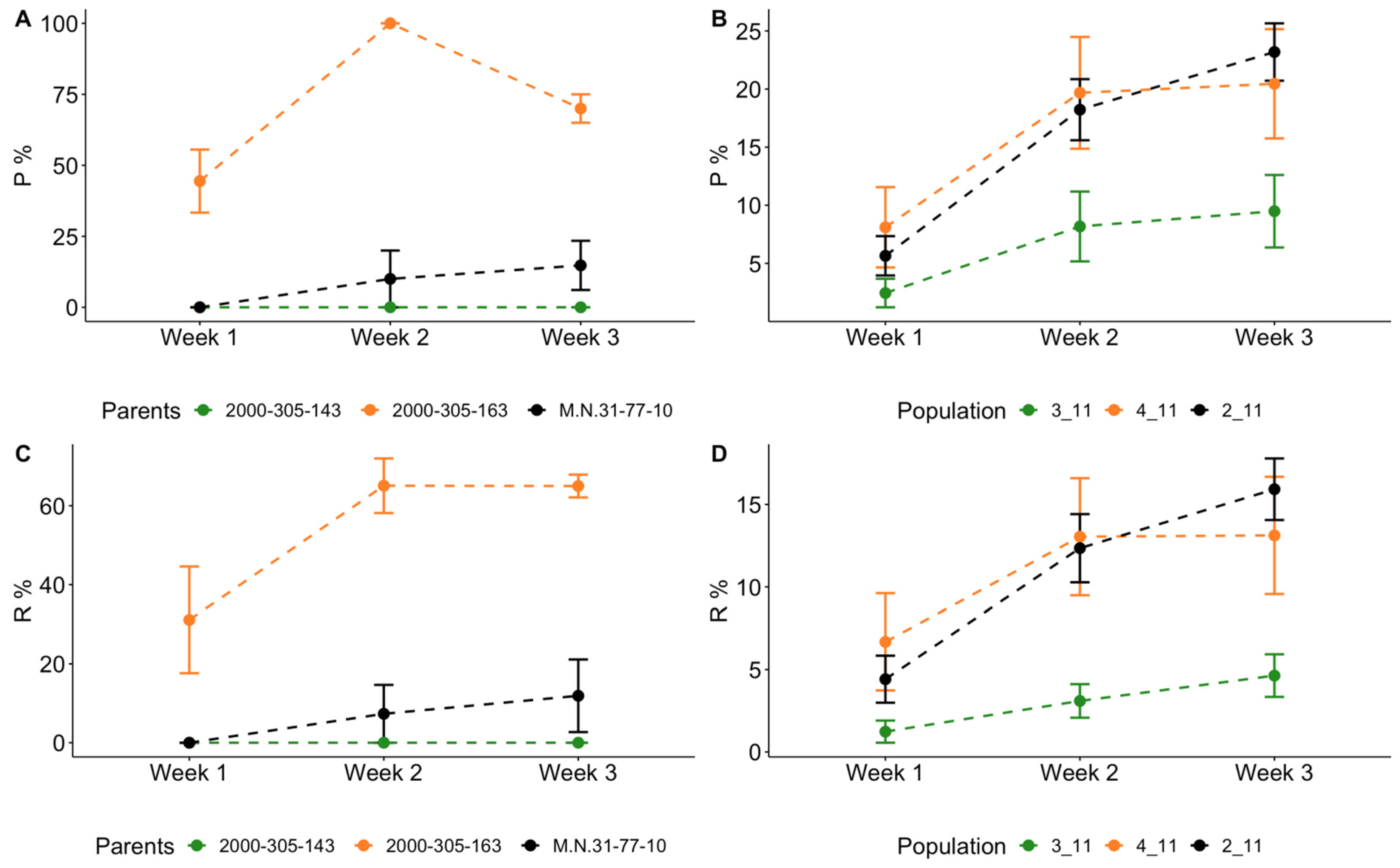
3.1. Laboratory Assessment of the Resistance of Hybrid Populations to Leaf Form of Phylloxera
3.2. Assessment of Parental Genotypes with SSR and SNP Markers Linked to the Phylloxera Resistance Loci in Muscadinia Background
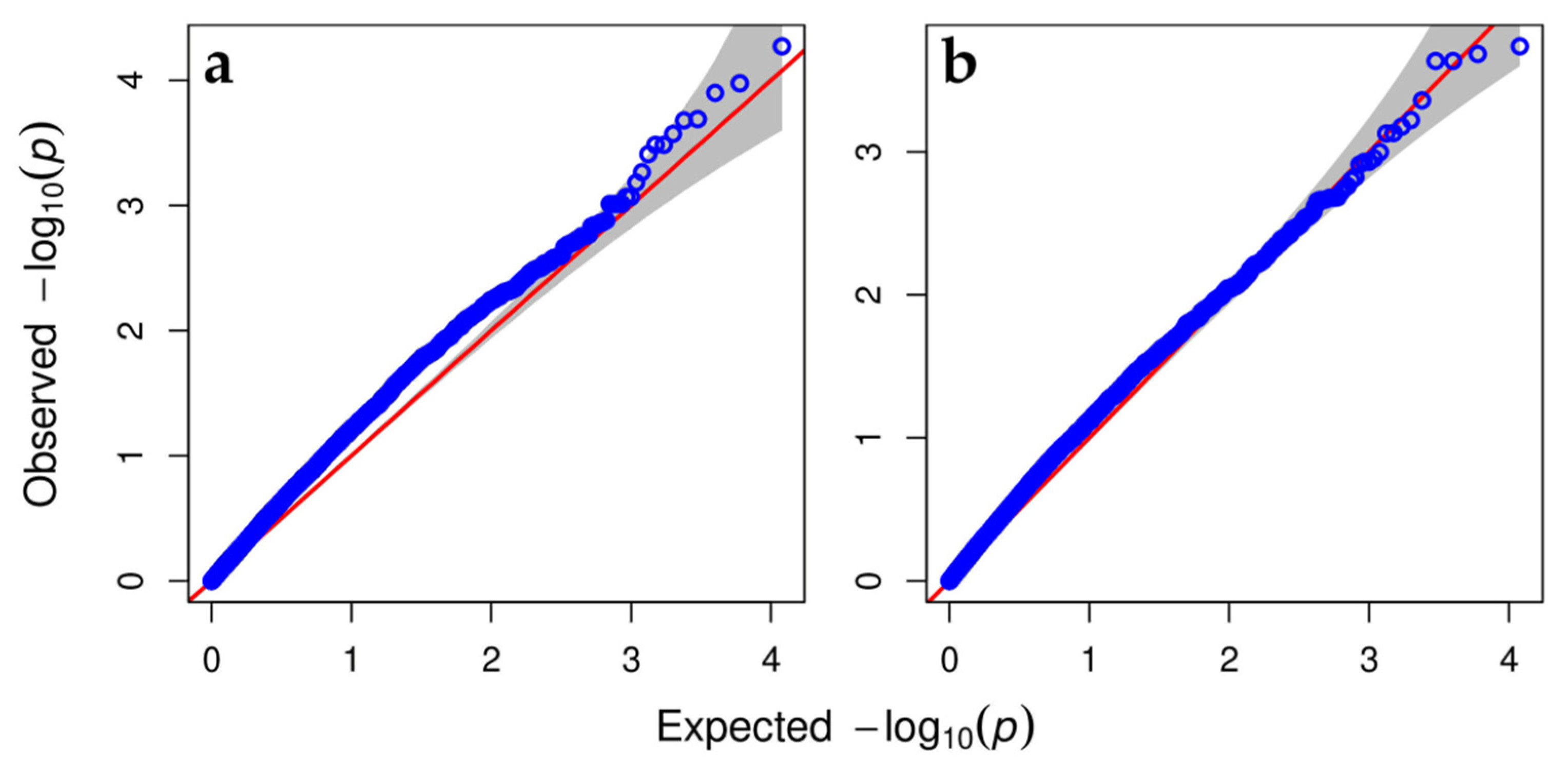
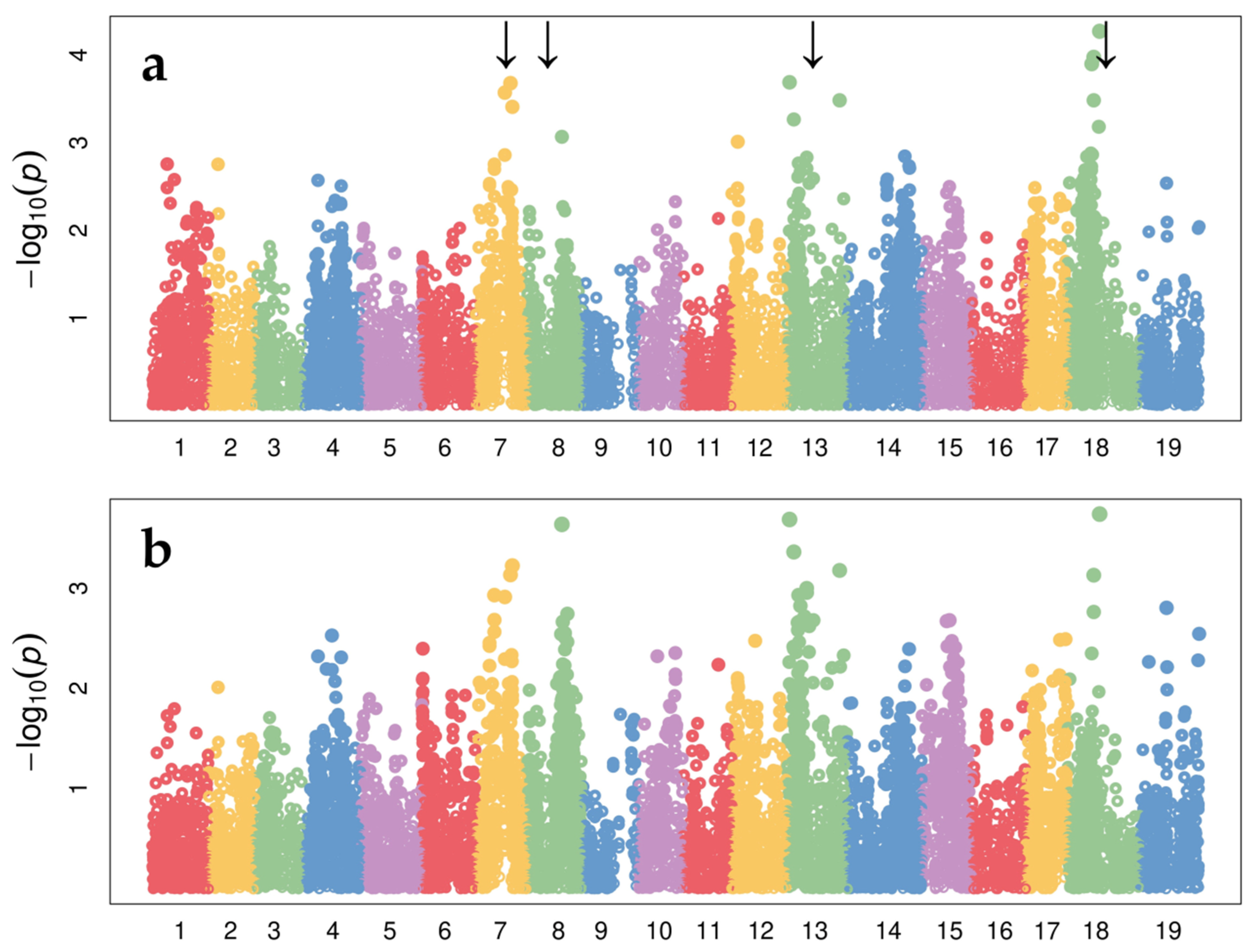
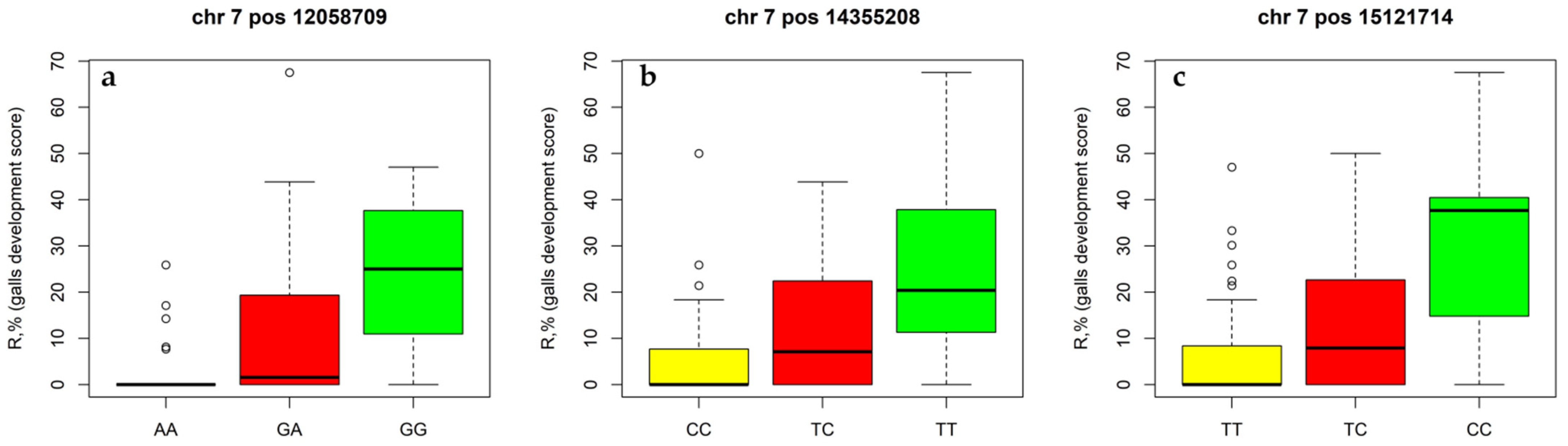
3.3. Genome-Wide Association Analysis (GWAS) of Variability in Resistance to the Leaf Form of Phylloxera in Three Studied Hybrid Populations
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- EPPO Global Database. Available online: https://gd.eppo.int/taxon/VITEVI/categorization (accessed on 12 November 2021).
- Ollat, N.; Cookson, S.J.; Lauvergeat, V.; Marguerit, E.; Barrieu, F.; Gambetta, G.; Goutouly, J.-P.; Tandonnet, J.-P.; Vivin, P.; Delrot, S. Grapevine Roots: The Dark Side. Acta Hortic. 2017, 1188, 213–226. [Google Scholar] [CrossRef]
- Skinkis, P.; Walton, V.M.; Kaiser, C. Grape Phylloxera Biology and Management in the Pacific Northwest. Oregon State University, OSU Extension Catalog 2009. Available online: https://catalog.extension.oregonstate.edu/sites/catalog/files/project/pdf/ec1463.pdf (accessed on 12 November 2021).
- Forneck, A.; Mammerler, R.; Tello, J.; Breuer, M.; Müller, J.; Fahrentrapp, J. First European Leaf-Feeding Grape Phylloxera (Daktulosphaira Vitifoliae Fitch) Survey in Swiss and German Commercial Vineyards. Eur. J. Plant Pathol. 2019, 154, 1029–1039. [Google Scholar] [CrossRef] [Green Version]
- Edwards, J.; Norng, S.; Powell, K.S.; Granett, J. Relationships between grape phylloxera abundance, fungal interactions and grapevine decline. Acta Hortic. 2007, 733, 151–157. [Google Scholar] [CrossRef]
- Usatov, V.; Kireyeva, L.; Volynkin, V.; Klimenko, V.; Oleinikov, N. Evaluation of Interspecific Populations of Grapevine in Breeding for Complex Resistance to Fungal Diseases and Phylloxera. Vitis-J. Grapevine Res. 1990, S, 278–294. [Google Scholar]
- Armengol, J.; Gramaje, D. Soilborne Fungal Pathogens Affecting Grapevine Rootstocks: Current Status and Future Prospects. Acta Hortic. 2016, 1136, 235–328. [Google Scholar] [CrossRef]
- Zhang, J.; Hausmann, L.; Eibach, R.; Welter, L.J.; Töpfer, R.; Zyprian, E.M. A Framework Map from Grapevine V3125 (Vitis Vinifera ‘Schiava Grossa’ × ‘Riesling’) × Rootstock Cultivar ‘Börner’ (Vitis Riparia × Vitis Cinerea) to Localize Genetic Determinants of Phylloxera Root Resistance. Theor. Appl. Genet. 2009, 119, 1039–1051. [Google Scholar] [CrossRef]
- Smith, H.M.; Clarke, C.W.; Smith, B.P.; Carmody, B.M.; Thomas, M.R.; Clingeleffer, P.R.; Powell, K.S. Genetic identification of SNP markers linked to a new grape phylloxera resistant locus in Vitis cinerea for marker-assisted selection. BMC Plant Biol. 2018, 18, 360. [Google Scholar] [CrossRef] [Green Version]
- Clark, M.D.; Teh, S.L.; Burkness, E.; Moreira, L.; Watson, G.; Yin, L.; Hutchison, W.D.; Luby, J.J. Quantitative Trait Loci Identified for Foliar Phylloxera Resistance in a Hybrid Grape Population: Foliar Phylloxera Resistance. Aust. J. Grape Wine Res. 2018, 24, 292–300. [Google Scholar] [CrossRef] [Green Version]
- Powell, K.S. Grape phylloxera: An overview. In Root Feeders: An Ecosystem Approach; Johnson, S.N., Murray, P.J., Eds.; CABI: Wallingford, CT, USA, 2008; pp. 96–114. ISBN 978-1-84593-461-3. [Google Scholar]
- Rubio, B.; Lalanne-Tisné, G.; Voisin, R.; Tandonnet, J.-P.; Portier, U.; Van Ghelder, C.; Lafargue, M.; Petit, J.-P.; Donnart, M.; Joubard, B.; et al. Characterization of Genetic Determinants of the Resistance to Phylloxera, Daktulosphaira Vitifoliae, and the Dagger Nematode Xiphinema Index from Muscadine Background. BMC Plant Biol. 2020, 20, 213. [Google Scholar] [CrossRef]
- Volynkin, V.; Vasylyk, I.; Volodin, V.; Grigoreva, E.; Karzhaev, D.; Lushchay, E.; Ulianich, P.; Volkov, V.; Risovannaya, V.; Blinova, S.; et al. The Assessment of Agrobiological and Disease Resistance Traits of Grapevine Hybrid Populations (Vitis Vinifera L. × Muscadinia Rotundifolia Michx.) in the Climatic Conditions of Crimea. Plants 2021, 10, 1215. [Google Scholar] [CrossRef]
- Volynkin, V.; Gorislavets, S.; Volodin, V.; Vasylyk, I.; Lushchay, E.; Likhovskoi, V.; Potokina, E. Immunogenic Breeding Program. Stage I-Phytopathological Screening of the Grape Gene Pool. E3S Web Conf. 2021, 254, 03003. [Google Scholar] [CrossRef]
- Bouquet, A. Introduction Dans l’espèce Vitis Vinifera L. d’un Caractère de Résistance à l’oidium (Uncinula Necator Schw. Burr.) Issu de l’espèce Muscadinia Rotundifolia (Michx.) Small. Vignevini 1986, 12, 141–146. [Google Scholar]
- Codes des Caracteres Descriptifs des Varietes et Especes de Vitis OIV 2009 Website of Organisation Internationale de la Vigne et du vin from. Available online: http://www.oiv.int/fr (accessed on 12 November 2021).
- Angelini, E.; Clair, D.; Borgo, M.; Bertaccini, A.; Boudon-Padieu, E. Flavescence Dorée in France and Italy-Occurrence of Closely Related Phytoplasma Isolates and Their near Relationships to Palatinate Grapevine Yellows and an Alder Yellows Phytoplasma. Vitis 2001, 40, 79–86. [Google Scholar]
- Okonechnikov, K.; Golosova, O.; Fursov, M. Unipro UGENE: A Unified Bioinformatics Toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [Green Version]
- Lipka, A.E.; Tian, F.; Wang, Q.; Peiffer, J.; Li, M.; Bradbury, P.J.; Gore, M.A.; Buckler, E.S.; Zhang, Z. GAPIT: Genome association and prediction integrated tool. Bioinformatics 2012, 28, 2397–2399. [Google Scholar] [CrossRef] [Green Version]
- Forneck, A.; Powell, K.S.; Walker, M.A. Scientific Opinion: Improving the Definition of Grape Phylloxera Biotypes and Standardizing Biotype Screening Protocols. Am. J. Enol. Vitic. 2016, 67, 371–376. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Trivedi, A.K. Regulation of apoptosis by E3 ubiquitin ligases in ubiquitin proteasome system. Cell Biol. Int. 2020, 44, 721–734. [Google Scholar] [CrossRef]
- Wilmink, J.; Breuer, M.; Forneck, A. Effect of Temperature on Host Plant-specific Leaf- and Root-feeding Performances: A Comparison of Grape Phylloxera Biotypes C and G. Entomol. Exp. Appl. 2021, 169, 1113–1125. [Google Scholar] [CrossRef]
- Stranishevskaya, E.; Misiak, A. The Effects of Leaf Phylloxera on the Fruitfulness Parameters, Yield and Quality of the Grape Plant Viticulture and Winemaking. Vitic. Winemak. 2010, 40, 53–55. [Google Scholar]
- Savi, T.; García González, A.; Herrera, J.C.; Forneck, A. Gas Exchange, Biomass and Non-Structural Carbohydrates Dynamics in Vines under Combined Drought and Biotic Stress. BMC Plant Biol. 2019, 19, 408. [Google Scholar] [CrossRef]
- Volynkin, V.; Levchenko, S.; Vasylyk, I.; Likhovskoi, V. Analysis of F2-F6 generations from hybridization with Vitis rotundifolia at the Institute Magarach. Acta Hortic. 2020, 1289, 269–274. [Google Scholar] [CrossRef]
- Bouquet, A. Etude de la résistance au phylloxera radicicole des hybrides Vitis vinifera× Muscadinia rotundifolia. Vitis 1983, 32, 311–323. [Google Scholar]
- Blank, L.; Wolf, T.; Eimert, K.; Schröder, M.-B. Differential Gene Expression during Hypersensitive Response in Phylloxera -Resistant Rootstock ‘Börner’ Using Custom Oligonucleotide Arrays. J. Plant Interact. 2009, 4, 261–269. [Google Scholar] [CrossRef]
- Eitle, M.W.; Griesser, M.; Vankova, R.; Dobrev, P.; Aberer, S.; Forneck, A. Grape Phylloxera (D. Vitifoliae) Manipulates SA/JA Concentrations and Signalling Pathways in Root Galls of Vitis Spp. Plant Physiol. Biochem. 2019, 144, 85–91. [Google Scholar] [CrossRef]
- Eitle, M.W.; Cargnoni, M.; Acar, A.; Crespo Martinez, S.; Failla, O.; Kaul, H.-P.; Griesser, M.; Forneck, A. Phylloxeration Effects on the Sink Activity and Assimilation Rate in Phylloxera (Daktulosphaira Vitifoliae Fitch) Infested Grapevines (Vitis spp.). Acta Hortic. 2017, 1188, 291–298. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OIV Resistance, Points | Visual Assessment of the Leaf Surface | Definition |
|---|---|---|
| 1 | Very large number of normal, fertile leaf galls; more than 50% of the leaf surface occupied by galls | Very low degree of resistance |
| 3 | Large number of normal, fertile leaf galls; 21–50% of the leaf surface occupied by galls | Low degree of resistance |
| 5 | Few, small, normal shaped, fertile leaf galls; 11–20% of the leaf surface occupied by galls | Medium degree of resistance |
| 7 | Few, uncompleted sterile leaf galls; up to 10% of the leaf surface occupied by galls | High resistance |
| 9 | Localized necrosis, punctures, no leaf galls | Very high resistance |
| Genotypes | LG7 | LG3 | LG10 | |||
|---|---|---|---|---|---|---|
| 7_4261424 40 cM | 7_13408919 75 cM | VVIQ67 88 cM | 3_5494608 | VMC3D7 | VVIN85 | |
| Dixie | C/C | T/T | 257/257 | C/C | 174/174 | 350/352 |
| Seibel 13-666 | C/T | T/T | 261/261 | C/C | 164/166 | 358/358 |
| Nimrang | C/C | T/T | 261/261 | C/T | 166/168 | 358/368 |
| M. no. 31-77-10 | C/C | T/T | 261/261 | C/T | 166/168 | 358/358 |
| Regent | C/T | T/T | 255/263 | C/T | 162/168 | 358/368 |
| 2000-305-143 | C/C | T/T | 257/263 | C/T | 168/168 | 368/368 |
| 2000-305-163 | C/C | T/T | 255/261 | C/T | 162/164 | 368/368 |
| Chromosome_SNP | p-Value, Intensity of Gall Formation | p-Value, % of Infested Leaves | Gene Symbol and Gene Region | Function |
|---|---|---|---|---|
| 7_14355208 | 0.0002 | 0.0003 | LOC100250982 (VIT_00005082001) (14352427-14362155) | Cell division cycle associated protein |
| 7_12058709 | 0.0003 | 0.0004 | LOC100267035 (12052165-12072010) | Chromatin modifica-tion-related protein EAF1 B |
| 7_15121714 | 0.0004 | 0.02 | LOC100262935 (VIT_00003711001) (15121373-15127749) | tRNA-dihydrouridine (20) synthase [NAD(P)+]-like |
| 8_14223679 | 0.0008 | 0.0002 | LOC100243094 (VIT_00025537001) (14214106-14227029) | E3 ubiqui-tin-protein ligase UPL4 |
| 8_14223697 | 0.0008 | 0.0002 | LOC100243094 (VIT_00025537001) (14214106-14227029) | E3 ubiqui-tin-protein ligase UPL4 |
| 13_21213131 | 0.0003 | 0.00008 | * LOC100261896 (VIT_00001464001) (21208653-21212237) | Uncharacterized LOC100261896 |
| 18_10506585 | 0.0001 | 0.0009 | LOC104882551 (VIT_00009690001) (10506198-10513987) | Uncharacterized LOC104882551 |
| 18_10506582 | 0.0003 | 0.003 | LOC104882551 (VIT_00009690001) (10506198-10513987) | Uncharacterized LOC104882551 |
| 18_12898048 | 0.03 | 0.00005 | LOC100260855 (VIT_00009966001) (12893538-12898481) | Uncharacterized LOC100260855 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vasylyk, I.; Gorislavets, S.; Matveikina, E.; Lushchay, E.; Lytkin, K.; Grigoreva, E.; Karzhaev, D.; Volkov, V.; Volodin, V.; Spotar, G.; et al. SNPs Associated with Foliar Phylloxera Tolerance in Hybrid Grape Populations Carrying Introgression from Muscadinia. Horticulturae 2022, 8, 16. https://doi.org/10.3390/horticulturae8010016
Vasylyk I, Gorislavets S, Matveikina E, Lushchay E, Lytkin K, Grigoreva E, Karzhaev D, Volkov V, Volodin V, Spotar G, et al. SNPs Associated with Foliar Phylloxera Tolerance in Hybrid Grape Populations Carrying Introgression from Muscadinia. Horticulturae. 2022; 8(1):16. https://doi.org/10.3390/horticulturae8010016
Chicago/Turabian StyleVasylyk, Irina, Svetlana Gorislavets, Elena Matveikina, Ekaterina Lushchay, Kirill Lytkin, Elizaveta Grigoreva, Dmitry Karzhaev, Vladimir Volkov, Vitalii Volodin, Gennady Spotar, and et al. 2022. "SNPs Associated with Foliar Phylloxera Tolerance in Hybrid Grape Populations Carrying Introgression from Muscadinia" Horticulturae 8, no. 1: 16. https://doi.org/10.3390/horticulturae8010016