
The Isolation and Identification of Anthocyanin-Related GSTs in Chrysanthemum


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Extraction of RNA and DNA
2.3. cDNA Synthesis and qRT-PCR
2.4. Phylogenetic Analysis of CmGSTs
2.5. Isolation and Sequence Analysis of CmGST1
2.6. Vector Construction
2.7. Subcellular Location Analysis
2.8. Measurement of Anthocyanin
2.9. Transient Overexpression on Tobacco Leaves
2.10. Gene Cloning and Analysis of the Promoter of CmGST1
2.11. Dual-Luciferase Report Assay
3. Results
3.1. Identification and Phylogenetic Analysis of CmGSTs
3.2. Anthocyanin Analysis in Different Developmental Stages of ‘Monalisa Pink’ Flower
3.3. Expression Patterns of CmGSTs of Phi Subgroup in Developmental Stages of ‘Monalisa Pink’ Flower Petals
3.4. Sequence Alignment of CmGST1 and CmGST13 in Chrysanthemum
3.5. Subcellular Localization of CmGST1
3.6. Transient Overexpression of CmGST1 in Tobacco Leaves
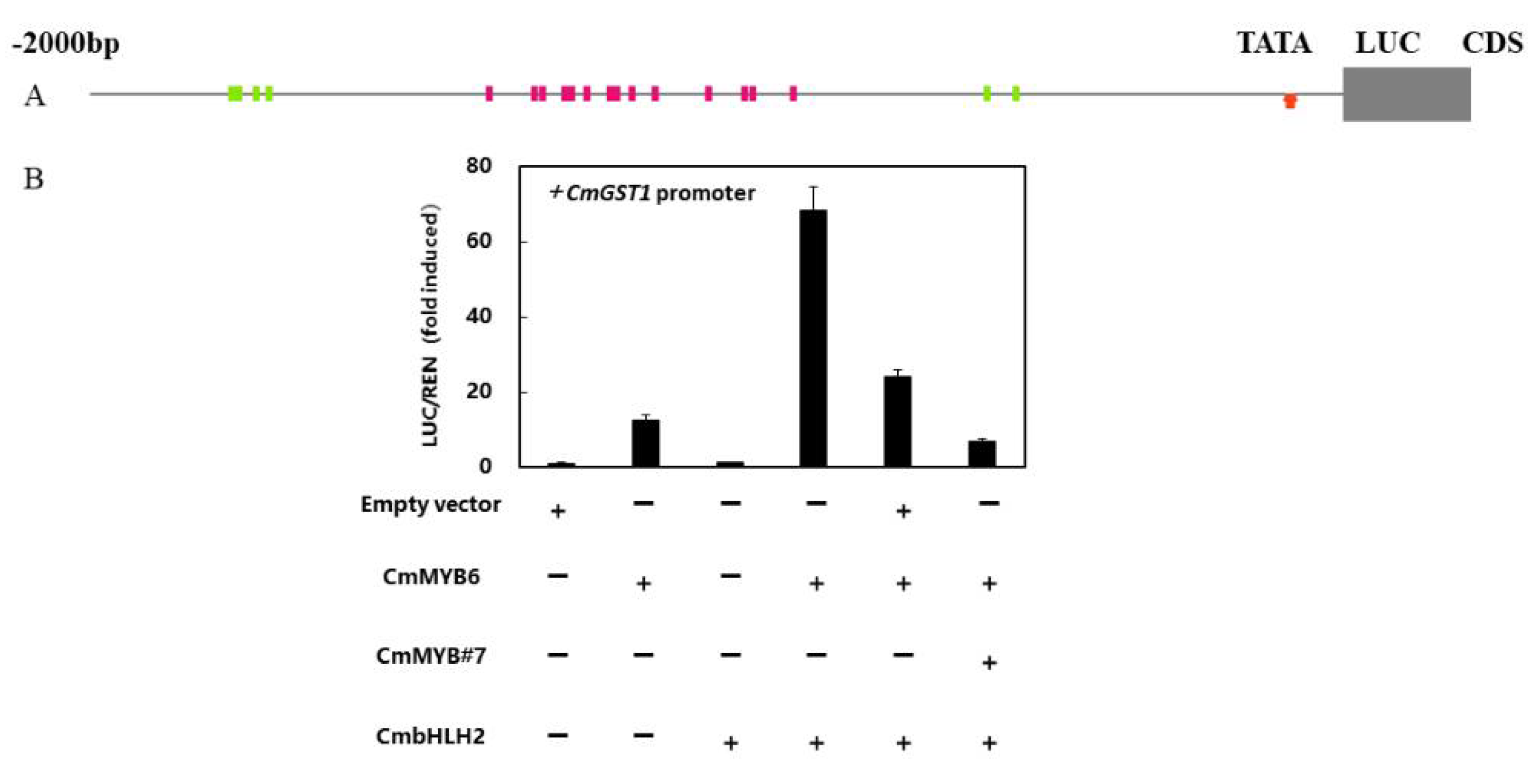
3.7. CmGST1 Promoter Was Regulated by Anthocyanin Related Transcription Factors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, X.F.; Xiang, L.L.; Yin, X.R.; Grierson, D.; Li, F.; Chen, K.S. The identification of a MYB transcription factor control ling anthocyanin biosynthesis regulation in Chrysanthemum flowers. Sci. Hortic. 2015, 194, 78–285. [Google Scholar] [CrossRef]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef] [PubMed]
- Bieza, K.; Lois, R. An Arabidopsis mutant tolerant to lethal ultraviolet-B levels shows constitutively elevated accumulation of flavonoids and other phenolics. Plant Physiol. 2001, 126, 1105–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veeriah, S.; Kautenburger, T.; Habermann, N.; Sauer, J.; Dietrich, H.; Will, F.; Pool-Zobel, B.L. Apple flavonoids inhibit growth of HT29 human colon cancer cells and modulate expression of genes involved in the biotransformation of xenobiotics. Mol. Arcinogen. 2006, 5, 164–174. [Google Scholar] [CrossRef]
- Winkel-Shirley, B. Flavonoid biosynthesis. A colorful model for genetics, biochemistry, cell biology, and biotechnology. Plant Physiol. 2001, 126, 485–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Dixon, R.A. The ‘ins’ and ‘outs’ of flavonoid transport. Trends Plant Sci. 2010, 15, 72–80. [Google Scholar] [CrossRef] [Green Version]
- Brazier-Hicks, M.; Evans, K.M.; Cunningham, O.D.; Hodgson, D.R.W.; Steel, P.G.; Edwards, R. Catabolism of glutathi one conjugates in Arabidopsis thaliana. Role in metabolic reactivation of the herbicide safener fenclorim. J. Biol. Chem. 2008, 283, 21102–21112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ketterer, B.; Meyer, D.J. Glutathione transferases: A possible role in the detoxication and repair of DNA and lipid hydroper oxides. Mutat. Res. 1989, 214, 33–40. [Google Scholar] [CrossRef]
- Edwards, R.; Dixon, D.P. Plant glutathione transferases. Gluthione Transferases Gamma-Glutamyl Transpeptidases 2005, 401, 169–186. [Google Scholar]
- Kitamura, S.; Shikazono, N.; Tanaka, A. TRANSPARENT TESTA 19 is involved in the accumulation of both anthocyanins and proanthocyanidins in Arabidopsis. Plant J. 2004, 37, 104–114. [Google Scholar] [CrossRef]
- Kitamura, S.; Akita, Y.; Ishizaka, H.; Narumi, I.; Tanaka, A. Molecular characterization of an anthocyanin-related glutathi one S-transferase gene in cyclamen. J. Plant Physiol. 2012, 169, 636–642. [Google Scholar] [CrossRef] [PubMed]
- Mueller, L.A.; Goodman, C.D.; Silady, R.A.; Walbot, V. AN9, a petunia glutathione S-transferase required for anthocyanin sequestration, is a flavonoid-binding protein. Plant Physiol. 2000, 123, 1561–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conn, S.; Curtin, C.; Bezier, A.; Franco, C.; Zhang, W. Purification, molecular cloning, and characterization of glutathione S-transferases (GSTs) from pigmented Vitis vinifera L. cell suspension cultures as putative anthocyanin transport proteins. J. Exp. Bot. 2008, 59, 3621–3634. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Li, H.; Huang, J.R. Arabidopsis TT19 functions as a carrier to transport anthocyanin from the cytosol to tonoplasts. Mol. Plant. 2012, 5, 387–400. [Google Scholar] [CrossRef]
- Jiang, S.; Chen, M.; He, N.B.; Chen, X.L.; Wang, N.; Sun, Q.G.; Zhang, T.H.; Xu, H.F.; Fang, H.C.; Wang, Y.C.; et al. MdGSTF6, activated by MdMYB1, plays an essential role in anthocyanin accumulation in apple. Hortic. Res. 2019, 6, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Dong, W.Q.; Zhu, Y.C.; Allan, A.C.; Wang, K.L.; Xu, C.J. PpGST1, an anthocyanin-related glutathione S-transfer ase gene, is essential for fruit coloration in peach. Plant Biotechnol. J. 2020, 18, 1284–1295. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.W.; Xu, L.F.; Xu, H.; Yang, P.P.; He, G.R.; Tang, Y.C.; Song, M.; Ming, J. LhGST is an anthocyanin-related glutathi one S-transferase gene in Asiatic hybrid lilies (Lilium spp.). Plant Cell Rep. 2021, 40, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Shao, D.N.; Li, Y.J.; Zhu, Q.H.; Zhang, X.Y.; Liu, F.; Xue, F. GhGSTF12, a glutathione S-transferase gene, is essential for anthocyanin accumulation in cotton (Gossypium hirsutum L.). Plant Sci. 2021, 305, 110827. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.; Puryear, J.; Cairney, J. A simple and efficient method for isolating RNA from pine trees. Mol. Biotechnol. 2001, 19, 201–203. [Google Scholar] [CrossRef]
- Liu, X.F.; Yin, X.R.; Allan, A.C.; Kui, L.W.; Shi, Y.N.; Huang, Y.J.; Ferguson, I.B.; Xu, C.J.; Chen, K.S. The role of MrbHLH1 and MrMYB1 in regulating anthocyanin biosynthetic genes in tobacco and Chinese bayberry (Myrica rubra) during anthocyanin biosynthesis. Plant Cell Tissue Organ Cult. 2013, 115, 285–298. [Google Scholar] [CrossRef]
- Mathews, H.; Clendennen, S.K.; Caldwell, C.G.; Liu, X.L.; Connors, K.; Matheis, N.; Schuster, D.K.; Menasco, D.J.; Wagoner, W.; Lightner, J.; et al. Activation Tagging in Tomato Identifies a Transcriptional Regulator of Anthocyanin Biosynthesis, Modification, and Transport. Plant Cell 2003, 15, 1689–1703. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Yin, X.R.; Zeng, J.K.; Ge, H.; Song, M.; Xu, C.J.; Li, X.; Ferguson, I.B.; Chen, K.S. Activator- and repressor-type MYB transcription factors are involved in chilling injury induced flesh lignification in loquat via their interactions with the phenylpropanoid pathway. J. Exp. Bot. 2014, 65, 4349–4359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, J.K.; Li, X.; Xu, Q.; Chen, J.Y.; Yin, X.R.; Ferguson, I.B.; Chen, K.S. EjAP2-1, an AP2/ERF gene, is a novel regulator of fruit lignification induced by chilling injury, via interaction with EjMYB transcription factors. Plant Biotechnol. J. 2015, 13, 1325–1334. [Google Scholar] [CrossRef] [PubMed]
- Reinemer, P.; Prade, L.; Hof, P.; Neuefeind, T.; Huber, R.; Zettl, R.; Palme, K.; Schell, J.; Koelln, I.; Bartunik, H.D.; et al. Three-dimensional structure of glutathione S-transferase from Arabidopsis thaliana at 2.2 angstrom resolution: Structural characterization of herbicide-conjugating plant glutathione S-transferases and a novel active site architecture. J. Mol. Biol. 1996, 255, 289–309. [Google Scholar] [CrossRef] [PubMed]
- Xiang, L.L.; Liu, X.F.; Li, X.; Yin, X.R.; Grierson, D.; Li, F.; Chen, K.S. A Novel bHLH Transcription Factor Involved in Regulating Anthocyanin Biosynthesis in Chrysanthemums (Chrysanthemum morifolium Ramat). PLoS ONE 2015, 10, e0143892. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J. Flavonoid transport mechanisms: How to go, and with whom. Trends Plant Sci. 2015, 20, 576–585. [Google Scholar] [CrossRef]
- Cheng, J.; Liao, L.; Zhou, H.; Gu, C.; Wang, L.; Han, Y.P. A small indel mutation in an anthocyanin transporter causes var iegated colouration of peach flowers. J. Exp. Bot. 2015, 66, 7227–7239. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.F.; Dai, C.; Li, Y.P.; Feng, J.; Liu, Z.C.; Kang, C.Y. Reduced Anthocyanins in Petioles codes for a GST anthocyanin transporter that is essential for the foliage and fruit coloration in strawberry. J. Exp. Bot. 2018, 69, 2595–2608. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Dong, W.Q.; Wang, K.; Zhang, B.; Allan, A.C.; Lin, W.K.; Chen, K.S.; Xu, C.J. Differential Sensitivity of Fruit Pigmentation to Ultraviolet Light between Two Peach Cultivars. Front. Plant Sci. 2017, 8, 1552. [Google Scholar] [CrossRef]
- Li, S.Y.; Zuo, D.Y.; Cheng, H.L.; Ali, M.; Wu, C.F.; Ashraf, J.; Zhang, Y.P.; Feng, X.X.; Lin, Z.X.; Wang, Q.L.; et al. Glutathione S-transferases GhGSTF1 and GhGSTF2 involved in the anthocyanin accumulation in Gossypium hirsutum L. Int. J. Biol. Macromol. 2020, 165, 2565–2575. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Dixon, R.A. MATE Transporters Facilitate Vacuolar Uptake of Epicatechin 3′-O-Glucoside for Proanthocyanidin Biosynthesis in Medicago truncatula and Arabidopsis. Plant Cell 2010, 22, 991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poustka, F.; Irani, N.; Feller, A.; Lu, Y.H.; Pourcel, L.; Frame, K.; Grotewold, E. A trafficking pathway for Anthocyanins overlaps with the endoplasmic reticulum-to-vacuole protein-sorting route in Arabidopsis and contributes to the formation of vacuolar inclusions. Plant Physiol. 2007, 145, 1323–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.B.; Wang, L.; Deroles, S.; Bennett, R.; Davies, K. New insight into the structures and formation of anthocyanic vacuolar inclusions in flower petals. BMC Plant Biol. 2006, 6, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Gomez, C.; Terrier, N.; Torregrosa, L.; Vialet, S.; Fournier-Level, A.; Verries, C.; Souquet, J.M.; Mazauric, J.P.; Klein, M.; Cheynier, V. Grapevine MATE-Type Proteins Act as Vacuolar H+-Dependent Acylated Anthocyanin Transporters. Plant Physiol. 2009, 150, 402–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Sung, S.Y.; Kim, Y.S.; Jo, Y.D.; Kang, S.Y.; Kim, J.B.; Ahn, J.W.; Ha, B.K.; Kim, D.S. Isolation and characteri zation of differentially expressed genes in petals of chrysanthemum mutant cultivars developed by irradiation. Sci. Hortic. 2015, 189, 132–138. [Google Scholar] [CrossRef]
- Liu, Y.F.; Qi, Y.W.; Zhang, A.; Wu, H.X.; Liu, Z.D.; Ren, X.L. Molecular cloning and functional characterization of AcGST1, an anthocyanin-related glutathione S-transferase gene in kiwifruit (Actinidia chinensis). Plant Mol. Biol. 2019, 100, 451–465. [Google Scholar] [CrossRef]
- Xiang, L.L.; Liu, X.F.; Li, H.; Yin, X.R.; Grierson, D.; Li, F.; Chen, K.S. CmMYB#7, an R3 MYB transcription factor, acts as a negative regulator of anthocyanin biosynthesis in chrysanthemum. J. Exp. Bot. 2019, 70, 3111–3123. [Google Scholar] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Liu, X.; Li, F.; Xiang, L.; Chen, K. The Isolation and Identification of Anthocyanin-Related GSTs in Chrysanthemum. Horticulturae 2021, 7, 231. https://doi.org/10.3390/horticulturae7080231
Li Y, Liu X, Li F, Xiang L, Chen K. The Isolation and Identification of Anthocyanin-Related GSTs in Chrysanthemum. Horticulturae. 2021; 7(8):231. https://doi.org/10.3390/horticulturae7080231
Chicago/Turabian StyleLi, Yajing, Xiaofen Liu, Fang Li, Lili Xiang, and Kunsong Chen. 2021. "The Isolation and Identification of Anthocyanin-Related GSTs in Chrysanthemum" Horticulturae 7, no. 8: 231. https://doi.org/10.3390/horticulturae7080231