
Nitrogen Effect on Growth-Related Parameters and Evaluation of Portulaca oleracea as a Phytoremediation Species in a Cr(VI)-Spiked Soil


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Establishment, Measurements and Soil and Plant Analysis
2.2. Quality Assurance and Statistical Analyses
3. Results
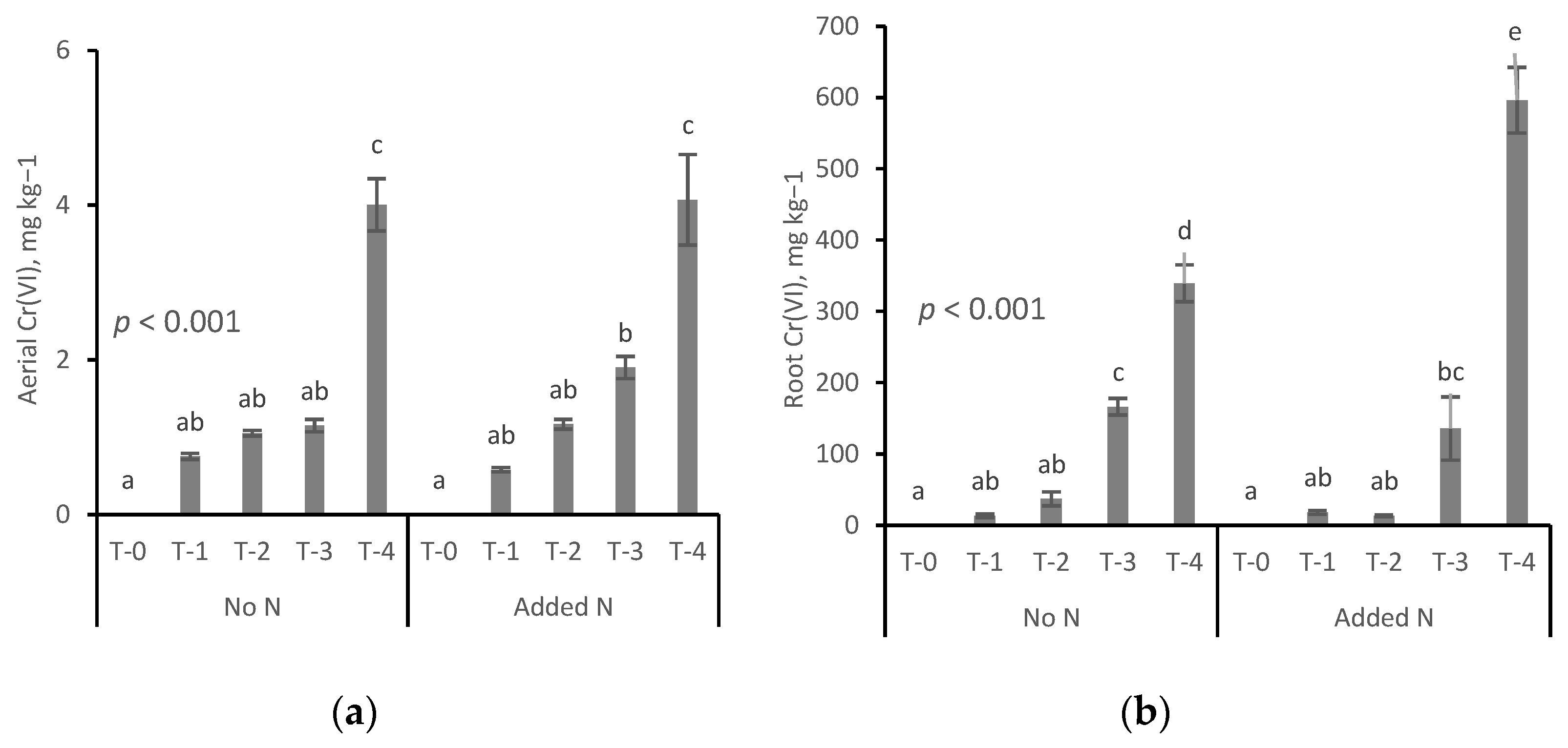
3.1. Cr(VI) Concentration in Plant Tissues
3.2. Effects of Cr(VI) on Parameters Relative to Photosynthesis and Plant Growth
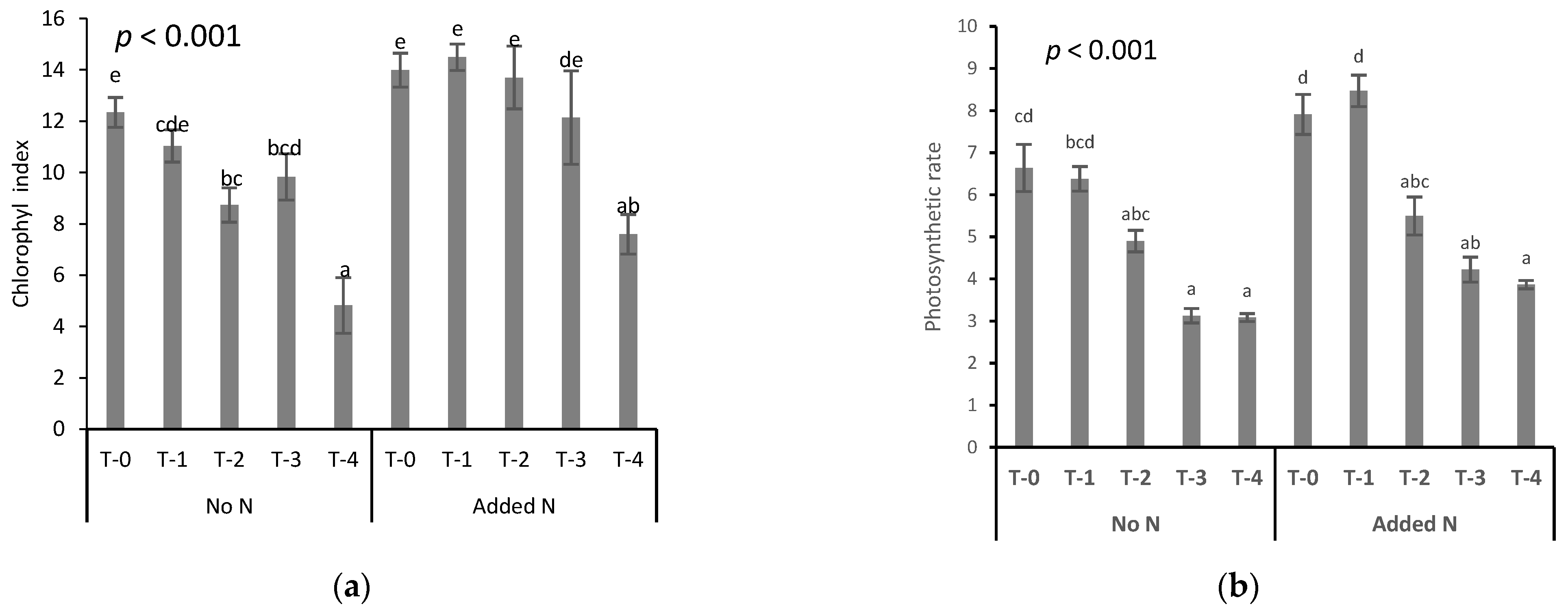
3.2.1. Chlorophyll Content Index and Photosynthetic Rate
3.2.2. Plant Height and Aerial Fresh Weight
3.2.3. Aerial Dry Weight and Root Dry Weight and Leaf Area per Plant
3.2.4. Leaf Area per Plant, Leaf Weight/Total Aerial Weight Ratio and Aerial Tissue Dry Matter Content (Aerial Dry Weight/Aerial Fresh Weight Ratio)
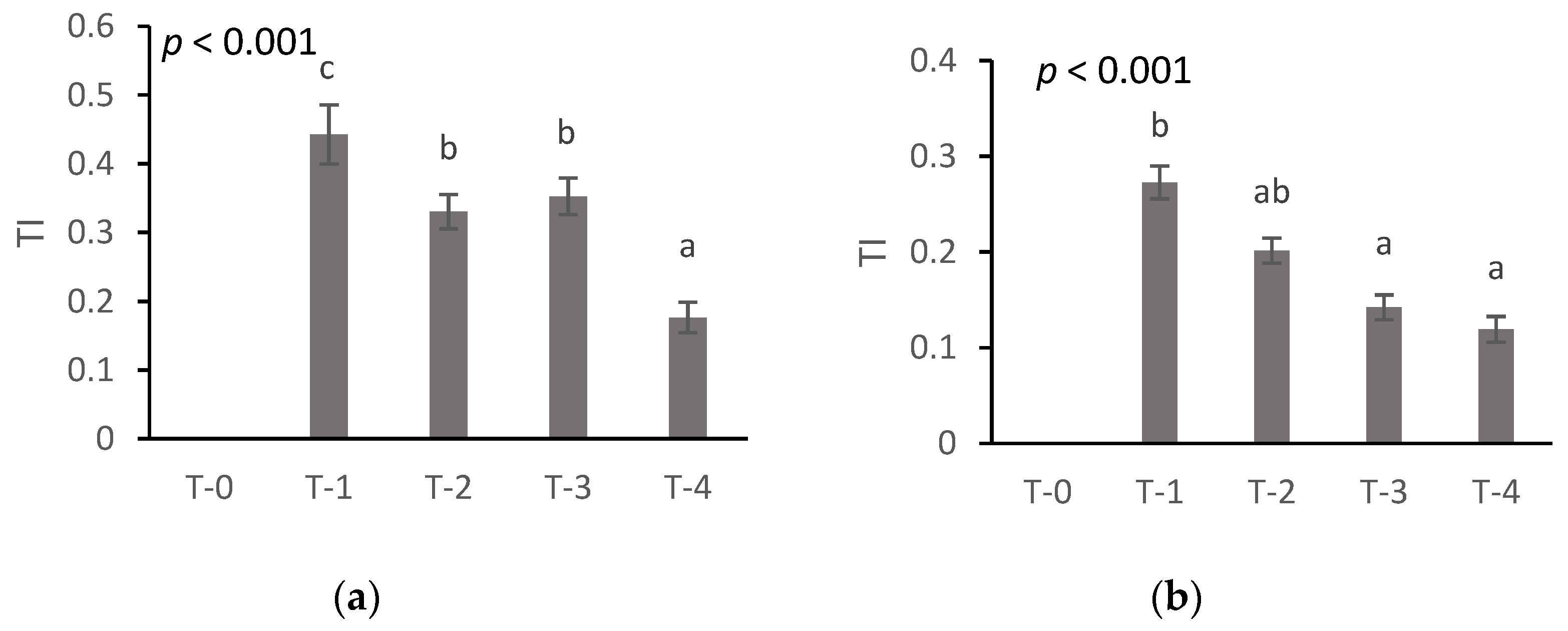
3.2.5. Tolerance Index
3.3. Phosphorus in Plant Tissues
4. Discussion
5. Conclusions
- Cr(VI) contents in root tissues were orders of magnitude higher than the concentrations found in aerial plant tissues.
- All physiological and growth parameters measured were severely affected and nitrogen in all cases resulted even partially in Cr(VI) stress alleviation.
- Under Cr(VI) stress purslane plants selectively accumulated phosphorus in aerial plant tissues.
- Cr(VI) stress resulted in lower water content in aerial plant tissues.
- Added N did not result in increased Cr(VI) content in aerial biomass compared to same Cr(VI)-amended treatments without N; however, the fact that added N improved plant’s growth and physiological functions even when exposed to high Cr(VI) soil concentrations, means that sufficient N fertilization may be a satisfactory treatment to increased purslane tolerance against Cr(VI) toxicity.
- On the same lines, added N makes purslane a species to be further considered for phytoremediation of Cr(VI)-laden soils; however, we acknowledge that more research is necessary before conclusive decisions may be drawn.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garrett, R.G. Natural sources of metals to the environment. Hum. Ecol. Risk Assess. 2000, 6, 945–963. [Google Scholar] [CrossRef]
- Shanker, A.K.; Cervantes, C.; Loza-Tavera, H.; Avudainayagam, S. Chromium toxicity in plants. Environ. Int. 2005, 31, 739–753. [Google Scholar] [CrossRef]
- Lilli, M.A.; Moraetis, D.; Nikolaidis, N.P.; Karatzas, G.P.; Kalogerakis, N. Characterization and mobility of geogenic chromium in soils and river bed sediments of Asopos basin. J. Hazard. Mater. 2015, 281, 12–19. [Google Scholar] [CrossRef]
- Megremi, I.; Vasilatos, C.; Vassilakis, E.; Economou-Eliopoulos, M. Spatial diversity of Cr distribution in soil and groundwater sites in relation with land use management in a Mediterranean region: The case of C. Evia and Assopos-Thiva Basins, Greece. Sci. Total Environ. 2019, 651, 656–667. [Google Scholar] [CrossRef]
- Jobby, R.; Jha, P.; Yadav, A.K.; Desai, N. Biosorption and biotransformation of hexavalent chromium [Cr(VI)]: A comprehensive review. Chemosphere 2018, 207, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Shamshad, S.; Rafiq, M.; Khalid, S.; Bibi, I.; Niazi, N.K.; Dumat, C.; Rashid, M.I. Chromium speciation, bioavailability, uptake, toxicity and detoxification in soil-plant system: A review. Chemosphere 2017, 178, 513–533. [Google Scholar] [CrossRef] [PubMed]
- US EPA. Chromium Compounds: Hazard Summary. Available online: https://www.epa.gov/sites/production/files/2016-09/documents/chromium-compounds.pdf (accessed on 15 June 2021).
- Kumar, P.; Tokas, J.; Singal, H.R. Amelioration of Chromium VI Toxicity in Sorghum (Sorghum bicolor L.) using Glycine Betaine. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kierczak, J.; Pietranik, A.; Pędziwiatr, A. Ultramafic geoecosystems as a natural source of Ni, Cr, and Co to the environment: A review. Sci. Total Environ. 2021, 755. [Google Scholar] [CrossRef] [PubMed]
- Antoniadis, V.; Levizou, E.; Shaheen, S.M.; Ok, Y.S.; Sebastian, A.; Baum, C.; Prasad, M.N.V.; Wenzel, W.W.; Rinklebe, J. Trace elements in the soil-plant interface: Phytoavailability, translocation, and phytoremediation—A review. Earth Sci. Rev. 2017, 171, 621–645. [Google Scholar] [CrossRef]
- Ertani, A.; Mietto, A.; Borin, M.; Nardi, S. Chromium in Agricultural Soils and Crops: A Review. Water Air Soil Pollut. 2017, 228. [Google Scholar] [CrossRef]
- Ahemad, M. Enhancing phytoremediation of chromium-stressed soils through plant-growth-promoting bacteria. J. Genet. Eng. Biotechnol. 2015, 13, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Sharma, I.; Pati, P.K.; Bhardwaj, R. Effect of 28-homobrassinolide on antioxidant defence system in Raphanus sativus L. under chromium toxicity. Ecotoxicology 2011, 20, 862–874. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Chaudhary, A.; Rizwan, M.; Anwar, H.T.; Adrees, M.; Farid, M.; Irshad, M.K.; Hayat, T.; Anjum, S.A. Alleviation of chromium toxicity by glycinebetaine is related to elevated antioxidant enzymes and suppressed chromium uptake and oxidative stress in wheat (Triticum aestivum L.). Environ. Sci. Pollut. Res. 2015, 22, 10669–10678. [Google Scholar] [CrossRef] [PubMed]
- Antoniadis, V.; Zanni, A.A.; Levizou, E.; Shaheen, S.M.; Dimirkou, A.; Bolan, N.; Rinklebe, J. Modulation of hexavalent chromium toxicity on Origanum vulgare in an acidic soil amended with peat, lime, and zeolite. Chemosphere 2018, 195, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Srivastava, S.; Prakash, S.; Srivastava, M.M. Fate of trivalent chromium in presence of organic acids: A hydroponic study on the tomato plant. Chem. Speciat. Bioavailab. 1998, 10, 147–150. [Google Scholar] [CrossRef]
- Ding, M.; Shi, X. Molecular mechanisms of Cr(VI)-induced carcinogenesis. Mol. Cell. Biochem. 2002, 234–235, 293–300. [Google Scholar] [CrossRef]
- Skeffington, R.A.; Shewry, P.R.; Peterson, P.J. Chromium Uptake and Transport in Barley Seedlings (Hordeum vulgare L.). Planta 1976, 132, 209–214. [Google Scholar] [CrossRef]
- Prasad, S.; Yadav, K.K.; Kumar, S.; Gupta, N.; Cabral-Pinto, M.M.S.; Rezania, S.; Radwan, N.; Alam, J. Chromium contamination and effect on environmental health and its remediation: A sustainable approaches. J. Environ. Manag. 2021, 285, 112174. [Google Scholar] [CrossRef]
- Nikalje, G.C.; Saini, N.; Suprasanna, P. Halophytes and Heavy Metals: Interesting Partnerships. In Plant-Metal Interactions; Srivastava, S., Srivastava, A., Suprasanna, P., Eds.; Springer Nature: Cham, Switzerland, 2019; pp. 99–118. ISBN 9783030207328. [Google Scholar]
- Karkanis, A.C.; Petropoulos, S.A. Physiological and growth responses of several genotypes of common purslane (Portulaca oleracea L.) under Mediterranean semi-arid conditions. Not. Bot. Horti Agrobot. Cluj-Napoca 2017, 45, 569–575. [Google Scholar] [CrossRef] [Green Version]
- Petropoulos, S.A.; Karkanis, A.; Martins, N.; Ferreira, I.C.F.R. Edible halophytes of the Mediterranean basin: Potential candidates for novel food products. Trends Food Sci. Technol. 2018, 74, 69–84. [Google Scholar] [CrossRef] [Green Version]
- Nikalje, G.C.; Suprasanna, P. Coping with metal toxicity—Cues from halophytes. Front. Plant Sci. 2018, 9, 1–11. [Google Scholar] [CrossRef]
- Petropoulos, S.; Levizou, E.; Ntatsi, G.; Fernandes, Â.; Petrotos, K.; Akoumianakis, K.; Barros, L.; Ferreira, I. Salinity effect on nutritional value, chemical composition and bioactive compounds content of Cichorium spinosum L. Food Chem. 2017, 214, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Lutts, S.; Lefèvre, I. How can we take advantage of halophyte properties to cope with heavy metal toxicity in salt-affected areas? Ann. Bot. 2015, 115, 509–528. [Google Scholar] [CrossRef]
- Lefèvre, I.; Marchal, G.; Meerts, P.; Corréal, E.; Lutts, S. Chloride salinity reduces cadmium accumulation by the Mediterranean halophyte species Atriplex halimus L. Environ. Exp. Bot. 2009, 65, 142–152. [Google Scholar] [CrossRef]
- Corrêa, R.C.; Di Gioia, F.; Ferreira, I.C.; Petropoulos, S.A. Wild greens used in the Mediterranean diet. In The Mediterranean Diet: An Evidence-Based Approach; Preedy, V., Watson, R., Eds.; Academic Press: London, UK, 2020; pp. 209–228. ISBN 9788578110796. [Google Scholar]
- Correa, R.C.G.; Di Gioia, F.; Ferreira, I.; SA, P. Halophytes for Future Horticulture: The Case of Small-Scale Farming in the Mediterranean Basin. In Halophytes for Future Horticulture: From Molecules to Ecosystems towards Biosaline Agriculture; Grigore, M.-N., Ed.; Springer Nature: Cham, Switzerland, 2020; pp. 1–28. ISBN 9783030178543. [Google Scholar]
- Rodriguez, E.; Santos, C.; Azevedo, R.; Moutinho-Pereira, J.; Correia, C.; Dias, M.C. Chromium (VI) induces toxicity at different photosynthetic levels in pea. Plant Physiol. Biochem. 2012, 53, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, M.; Ganesh, K.S. Effect of Chromium on Growth, Biochemicals and Nutrient Accumulation of Paddy (Oryza sativa L.). Int. Lett. Nat. Sci. 2014, 23, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Moral, R.; Navarro Pedreno, J.; Gomez, I.; Mataix, J. Effects of chromium on the nutrient element content and morphology of tomato. J. Plant Nutr. 1995, 18, 815–822. [Google Scholar] [CrossRef]
- Kumar, S.; Joshi, U.N. Nitrogen metabolism as affected by hexavalent chromium in sorghum (Sorghum bicolor L.). Environ. Exp. Bot. 2008, 64, 135–144. [Google Scholar] [CrossRef]
- Alyazouri, A.; Jewsbury, R.; Tayim, H.; Humphreys, P.; Al-Sayah, M.H. Applicability of Heavy-Metal Phytoextraction in United Arab Emirates: An Investigation of Candidate Species. Soil Sediment Contam. 2014, 23, 557–570. [Google Scholar] [CrossRef]
- Feller, U.; Kopriva, S.; Vassileva, V. Plant nutrient dynamics in stressful environments: Needs interfere with burdens. Agriculture 2018, 8, 97. [Google Scholar] [CrossRef] [Green Version]
- Haneklaus, S.H.; Bloem, E.; Schnug, E. Hungry plants—A short treatise on how to feed crops under stress. Agriculture 2018, 8, 43. [Google Scholar] [CrossRef] [Green Version]
- Subpiramaniyam, S. Portulaca oleracea L. for phytoremediation and biomonitoring in metal-contaminated environments. Chemosphere 2021, 280, 130784. [Google Scholar] [CrossRef] [PubMed]
- Sdouga, D.; Ben Amor, F.; Ghribi, S.; Kabtni, S.; Tebini, M.; Branca, F.; Trifi-Farah, N.; Marghali, S. An insight from tolerance to salinity stress in halophyte Portulaca oleracea L.: Physio-morphological, biochemical and molecular responses. Ecotoxicol. Environ. Saf. 2019, 172, 45–52. [Google Scholar] [CrossRef]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef]
- Alyazouri, A.; Jewsbury, R.; Tayim, H.; Humphreys, P.; Al-Sayah, M.H. Uptake of Chromium by Portulaca oleracea from Soil: Effects of Organic Content, pH, and Sulphate Concentration. Appl. Environ. Soil Sci. 2020, 2020. [Google Scholar] [CrossRef]
- Rahbarian, R.; Azizi, E.; Behdad, A.; Mirblook, A. Effects of Chromium on Enzymatic/Nonenzymatic Antioxidants and Oxidant Levels of Portulaca oleracea L. J. Med. Plants Prod. 2019, 1, 21–31. [Google Scholar]
- Renna, M.; Cocozza, C.; Gonnella, M.; Abdelrahman, H.; Santamaria, P. Elemental characterization of wild edible plants from countryside and urban areas. Food Chem. 2015, 177, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Brozou, E.; Ioannou, Z.; Dimirkou, A. Removal of Cr (VI) and Cr (III) From Polluted Water and Soil Sown with Beet (Beta vulgaris) or Celery (Apium graveolens) after the Addition of Modified Zeolites. Int. J. Waste Resour. 2018, 8. [Google Scholar] [CrossRef]
- Koutroubas, S.D.; Antoniadis, V.; Damalas, C.A.; Fotiadis, S. Sunflower growth and yield response to sewage sludge application under contrasting water availability conditions. Ind. Crops Prod. 2020, 154, 112670. [Google Scholar] [CrossRef]
- Jones, J.B.J.; Case, V.W. Chapter 15 Sampling, Handling, and Analyzing. In Soil Testing and Plant Analysis; Westerman, R.L., Ed.; Soil Science Society of America Inc.: Madison, WI, USA, 1990; Volume 3, pp. 389–427. ISBN 0891187936. [Google Scholar]
- Oliveira, H. Chromium as an Environmental Pollutant: Insights on Induced Plant Toxicity. J. Bot. 2012, 2012, 375843. [Google Scholar] [CrossRef]
- Appenroth, K.J.; Keresztes, Á.; Sárvári, É.; Jaglarz, A.; Fischer, W. Multiple effects of chromate on Spirodela polyrhiza: Electron microscopy and biochemical investigations. Plant Biol. 2003, 5, 315–323. [Google Scholar] [CrossRef] [Green Version]
- Kale, R.A.; Lokhande, V.H.; Ade, A.B. Investigation of chromium phytoremediation and tolerance capacity of a weed, Portulaca oleracea L. in a hydroponic system. Water Environ. J. 2015, 29, 236–242. [Google Scholar] [CrossRef]
- López-Bucio, J.; Hernández-Madrigal, F.; Cervantes, C.; Ortiz-Castro, R.; Carreón-Abud, Y.; Martínez-Trujillo, M. Phosphate relieves chromium toxicity in Arabidopsis thaliana plants by interfering with chromate uptake. BioMetals 2014, 27, 363–370. [Google Scholar] [CrossRef]
- Castro, R.O.; Trujillo, M.M.; López Bucio, J.; Cervantes, C.; Dubrovsky, J. Effects of dichromate on growth and root system architecture of Arabidopsis thaliana seedlings. Plant Sci. 2007, 172, 684–691. [Google Scholar] [CrossRef]
- Levizou, E.; Zanni, A.A.; Antoniadis, V. Varying concentrations of soil chromium (VI) for the exploration of tolerance thresholds and phytoremediation potential of the oregano (Origanum vulgare). Environ. Sci. Pollut. Res. 2019, 26, 14–23. [Google Scholar] [CrossRef]
- Chatterjee, J.; Chatterjee, C. Phytotoxicity of cobalt, chromium and copper in cauliflower. Environ. Pollut. 2000, 109, 69–74. [Google Scholar] [CrossRef]
- Dwivedi, S.; Mishra, A.; Kumar, A.; Tripathi, P.; Dave, R.; Dixit, G.; Tiwari, K.K.; Srivastava, S.; Shukla, M.K.; Tripathi, R.D. Bioremediation potential of genus Portulaca L. collected from industrial areas in Vadodara, Gujarat, India. Clean Technol. Environ. Policy 2012, 14, 223–228. [Google Scholar] [CrossRef]
- Anandi, S.; Thangavel, P.; Subburam, V. Influence of aluminium on the restoration potential of a terrestrial vascular plant, Portulaca oleracea L. as a biomonitoring tool of fresh water aquatic environments. Environ. Monit. Assess. 2002, 78, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Deepa, R.; Senthilkumar, P.; Sivakumar, S.; Duraisamy, P.; Subbhuraam, C.V. Copper availability and accumulation by Portulaca oleracea Linn. stem cutting. Environ. Monit. Assess. 2006, 116, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, K.K.; Dwivedi, S.; Mishra, S.; Srivastava, S.; Tripathi, R.D.; Singh, N.K.; Chakraborty, S. Phytoremediation efficiency of Portulaca tuberosa rox and Portulaca oleracea L. naturally growing in an industrial effluent irrigated area in Vadodra, Gujrat, India. Environ. Monit. Assess. 2008, 147, 15–22. [Google Scholar] [CrossRef]
- Yang, Y.; Zhou, Z.; Bai, Y.; Jiao, W.; Chen, W. Trace Elements in Dominant Species of the Fenghe River, China: Their Relations to Environmental Factors. J. Environ. Qual. 2016, 45, 1252–1258. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, L.M.; Ma, L.Q.; Santos, J.A.G.; Guilherme, L.R.G.; Lessl, J.T. Effects of arsenate, chromate, and sulfate on arsenic and chromium uptake and translocation by arsenic hyperaccumulator Pteris vittata L. Environ. Pollut. 2014, 184, 187–192. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, L.M.; Gress, J.; De, J.; Rathinasabapathi, B.; Marchi, G.; Chen, Y.; Ma, L.Q. Sulfate and chromate increased each other’s uptake and translocation in As-hyperaccumulator Pteris vittata. Chemosphere 2016, 147, 36–43. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Chen, N.; Feng, C.; Li, M.; Gao, Y.; Hu, Y. Coupling enhancement of Chromium(VI) bioreduction in groundwater by phosphorus minerals. Chemosphere 2020, 240, 124896. [Google Scholar] [CrossRef]
- Dube, B.K.; Tewari, K.; Chatterjee, J.; Chatterjee, C. Excess chromium alters uptake and translocation of certain nutrients in citrullus. Chemosphere 2003, 53, 1147–1153. [Google Scholar] [CrossRef]
- Yao, Y.; Sun, H.; Xu, F.; Zhang, X.; Liu, S. Comparative proteome analysis of metabolic changes by low phosphorus stress in two Brassica napus genotypes. Planta 2011, 233, 523–537. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Treatments | Plant Height | Aerial Fresh Weight | |
|---|---|---|---|
| (cm) | (g pot−1) | ||
| No N | T-0 | 29.9 ± 1.09 f | 13.4 ± 2.18 |
| T-1 | 18.9 ± 0.57 d | 5.9 ± 0.58 | |
| T-2 | 16.3 ± 0.91 c | 4.4 ± 0.33 | |
| T-3 | 15.0 ± 0.41 bc | 4.5 ± 0.35 | |
| T-4 | 11.2 ± 0.46 a | 2.3 ± 0.31 | |
| Added N | T-0 | 31.2 ± 1.39 d | 29.5 ± 3.21 |
| T-1 | 22.0 ± 0.67 bc | 8.2 ± 0.52 | |
| T-2 | 15.7 ± 0.48 ab | 5.7 ± 0.30 | |
| T-3 | 15.7 ± 0.99 ab | 4.2 ± 0.38 | |
| T-4 | 13.5 ± 0.46 a | 3.5 ± 0.40 | |
| Treatment effect | p < 0.001 | p = 0.298 | |
| Cr effect | p < 0.001 | p < 0.001 | |
| Nitrogen effect | p = 0.038 | p < 0.001 | |
| Treatments | Aerial Dry Weight | Root Dry Weight | |
|---|---|---|---|
| (g pot−1) | (g pot−1) | ||
| No N | T-0 | 0.99 ± 0.167 b | 0.30 ± 0.089 a |
| T-1 | 0.53 ± 0.009 ab | 0.39 ± 0.053 ab | |
| T-2 | 0.42 ± 0.015 a | 0.56 ± 0.084 abc | |
| T-3 | 0.48 ± 0.049 ab | 0.83 ± 0.093 abc | |
| T-4 | 0.23 ± 0.015 a | 0.50 ± 0.127 abc | |
| Added N | T-0 | 2.73 ± 0.339 c | 0.38 ± 0.056 ab |
| T-1 | 0.76 ± 0.107 ab | 0.76 ± 0.169 abc | |
| T-2 | 0.50 ± 0.019 ab | 1.02 ± 0.126 c | |
| T-3 | 0.45 ± 0.051 a | 0.79 ± 0.095 abc | |
| T-4 | 0.36 ± 0.052 a | 0.96 ± 0.146 bc | |
| Treatment effect | p < 0.001 | p = 0.045 | |
| Cr effect | p < 0.001 | p = 0.053 | |
| Nitrogen effect | p < 0.001 | p = 0.021 | |
| Treatments | Leaf Area | Leaf/Aerial Weight Ratio | Aerial Tissue (Dry Weight/Fresh Weight Ratio) | |
|---|---|---|---|---|
| (cm2 Plant−1) | (g g−1) | (g g−1) | ||
| No N | T-0 | 114.5 ± 18.8 b | 0.33 ± 0.020 | 0.0723± 0.0032 a |
| T-1 | 59.6 ± 2.5 ab | 0.28 ± 0.021 | 0.0936 ± 0.0015 bc | |
| T-2 | 43.3 ±2.8 a | 0.27 ± 0.028 | 0.0884 ± 0.0041 bc | |
| T-3 | 48.3 ±4.1 a | 0.30 ± 0.022 | 0.0934 ± 0.0042 bc | |
| T-4 | 41.0 ± 3.7 a | 0.26 ± 0.025 | 0.0953 ± 0.0047 bc | |
| Added N | T-0 | 381.5 ± 19.0 c | 0.50 ± 0.070 | 0.0790 ± 0.0026 ab |
| T-1 | 120.9 ± 4.1 b | 0.31 ± 0.012 | 0.0875 ± 0.0009 bc | |
| T-2 | 98.3 ± 5.9 ab | 0.33 ± 0.018 | 0.0927 ± 0.004 bc | |
| T-3 | 68.4 ± 2.4 ab | 0.31 ± 0.027 | 0.0996 ± 0.004 c | |
| T-4 | 49.9 ± 1.9 a | 0.26 ± 0.024 | 0.0998 ± 0.008 c | |
| Treatment effect | p < 0.001 | p = 0.358 | p = 0.015 | |
| Cr effect | p < 0.001 | p < 0.001 | p = 0.002 | |
| Nitrogen effect | p < 0.001 | p = 0.007 | p = 0.324 | |
| Treatments | Phosphorus Content | Potassium Content | |
|---|---|---|---|
| No N | T-0 | 3.41 ab | 39.50 f |
| T-1 | 4.10 abc | 28.75 abc | |
| T-2 | 5.10 cd | 31.62 bcde | |
| T-3 | 4.72 bcd | 34.65 cdef | |
| T-4 | 6.10 d | 37.63 ef | |
| Added N | T-0 | 3.99 abc | 35.50 def |
| T-1 | 2.95 a | 24.04 a | |
| T-2 | 4.16 abc | 25.92 ab | |
| T-3 | 4.40 abc | 36.07 ef | |
| T-4 | 4.75 bcd | 29.17 abcd | |
| Treatment effect | p < 0.001 | p < 0.001 | |
| Cr effect | p < 0.001 | p < 0.001 | |
| Nitrogen effect | p = 0.034 | p = 0.002 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thalassinos, G.; Nastou, E.; Petropoulos, S.A.; Antoniadis, V. Nitrogen Effect on Growth-Related Parameters and Evaluation of Portulaca oleracea as a Phytoremediation Species in a Cr(VI)-Spiked Soil. Horticulturae 2021, 7, 192. https://doi.org/10.3390/horticulturae7070192
Thalassinos G, Nastou E, Petropoulos SA, Antoniadis V. Nitrogen Effect on Growth-Related Parameters and Evaluation of Portulaca oleracea as a Phytoremediation Species in a Cr(VI)-Spiked Soil. Horticulturae. 2021; 7(7):192. https://doi.org/10.3390/horticulturae7070192
Chicago/Turabian StyleThalassinos, Georgios, Elina Nastou, Spyridon A. Petropoulos, and Vasileios Antoniadis. 2021. "Nitrogen Effect on Growth-Related Parameters and Evaluation of Portulaca oleracea as a Phytoremediation Species in a Cr(VI)-Spiked Soil" Horticulturae 7, no. 7: 192. https://doi.org/10.3390/horticulturae7070192