1. Introduction
Conventional breeding of bananas and plantains (
Musa spp.) is challenged by poor seed set, which is attributed to a combination of factors [
1]. This slows down genetic gain because breeders cannot quickly create a wide progeny base for meaningful selections. The breeding potential of the available
Musa spp. gene pool has therefore not been fully exploited as some cultivars have been rendered “infertile” [
2,
3]. Besides, among Matooke and plantains, seed set is highly variable between genotypes and seasons [
2,
4,
5]. High temperature, high solar radiation, low rainfall amounts, and low relative humidity (RH) have been observed to increase seed set when plantain and Matooke flowers are pollinated manually [
6,
7].
However, weather attributes only explain a small percentage of variability in data. In a cross of the plantain ‘Bobby Tannap’ and the wild banana ‘Calcutta 4′, a simple linear regression for seed set and maximum RH could only explain 31.7% of variability [
6]. Correlation coefficients of weather attributes and seed set were also mostly below 0.5 for many Matooke genotypes [
7]. This suggests that other factors are yet to be identified when relating weather effects on seed set in banana. It is also not clear when weather most affects seed set and the weather attribute that accounts for most of the variability. Batte et al. [
8] reported that the month of pollination had a non-significant effect on pollination success, but seed set differed between years in Matooke. Moreover, variability between years is smaller than the variability between seasons or months. Therefore, differences in pollination success rates between months were expected, though only differences of total seed set between years were noticed [
8]. Since the onset of seasons varies between years, taking into account the variability of months within years could have brought out month effects in the analysis. Cultivar differences also exist [
4,
5]. Thus, analyses per cultivar could have brought out month effects much more accurately.
Embryos arising from bunches pollinated in a wet season were also reported to have a higher germination rate [
9]. These observations call for an in-depth investigation of weather effects on seed set. However, it is noteworthy that seed fertility is a desirable trait during breeding but not in the final hybrids for consumers. Banana breeders therefore have to understand sterility in order to use it to their advantage. The aim of this study was therefore to assess the effects of weather on seed set before pollination, at the time of pollination and after pollination in selected East African highland cooking bananas (EAHBs). An understanding of the stages when weather is most influential on fertility and the responsible weather attributes can help in designing manipulative experiments to increase seed set and thus improve breeding efficiency.
2. Materials and Methods
Female fertile ‘Enzirabahima’ (
Musa AAA group, Matooke subgroup) and ‘Mshale’ and ‘Nshonowa’ (
Musa AA group, Mchare subgroup) were pollinated with pollen fertile ‘Calcutta 4′ (
M. acuminata spp.
burmannicoides) from January 2016 to January 2019. Matooke is grown throughout the Great Lakes region of East Africa including Uganda, Rwanda, Burundi, DR. Congo, Kenya, and Tanzania [
10,
11]. On the other hand, Mchare is more widely adapted to different agro-ecological zones covering Madagascar, Comoros islands, Pemba and Zanzibar islands, around Mount Kilimanjaro and Mount Kenya, to as far as a few spots in central Uganda [
11]. Matooke and Mchare banana types belong to the same genetic complex [
11] and can be collectively referred to as EAHBs. All pollinations were made at the National Agricultural Research Laboratories (NARL)–Kawanda station located at latitude 0° 25′ N, longitude 32° 32′ E at an elevation of 1177 m above sea level. The soils at Kawanda are sandy-loams of the deep ferrallitic clay type with a pH range of 5.5–6.0 [
12]. Daily weather attributes in the study period are summarized in
Table 1.
The pollination technique was a modification of that described by Vuylsteke et al. [
13] by applying pollen germination media (PGM) to enhance stigma receptivity [
14]. PGM used in 2016 was prepared using 30 g/liter glucose in tap water only. On the other hand, complete PGM used from January 2017 to January 2019 was prepared with 30 g glucose, 0.25 g MgSO
4.7H
2O, 0.25 g KNO
3, 0.4 g Ca(NO
3)
2, and 0.1 g H
3BO
3 per liter [
14]. The use of complete glucose PGM yielded higher seed, but it was not statistically different. The PGM was applied to pollen dusted on stigmas in a fine mist using a hand spray pump as described by Waniale et al. [
15]. At full maturity, when fingers had just started ripening (yellowing), bunches were harvested, kept in a ripening room, and seed hand extracted from fruit pulp, washed, air-dried, and counted. Fingers per bunch and hands per bunch were also counted and recorded. Seed set was then standardized to seed set per 100 fruits per bunch as follows:
Table 1.
Daily weather attributes between January 2016 and January 2019 at the National Agricultural Research Laboratories.
Table 1.
Daily weather attributes between January 2016 and January 2019 at the National Agricultural Research Laboratories.
| Weather Attribute and Data Source | Maximum | Minimum | Average |
|---|
| Temp at 9 a.m. (°C)–NARL | 22.1 | 14.5 | 17.9 |
| Temp at 3 p.m. (°C)–NARL | 35.0 | 21.0 | 28.8 |
| Temp Max at 2 m (°C)–NASA | 32.2 | 20.5 | 26.0 |
| Temp Min at 2 m (°C)–NASA | 24.3 | 15.9 | 20.5 |
| Temp range at 2 m (°C)–NASA | 13.0 | 1.6 | 5.5 |
| Average temp at 2 m (°C)–NASA | 27.6 | 19.5 | 23.2 |
| Earth skin temp (°C)–NASA | 28.6 | 21.0 | 24.2 |
| Solar radiation (kW-hr/m2/day)–NASA | 7.5 | 0.7 | 5.2 |
| Relative humidity at 9 a.m. (%)–NARL | 100.0 | 52.0 | 89.4 |
| Relative humidity at 3 p.m. (%)–NARL | 100.0 | 24.0 | 71.5 |
| Average RH at 2 m (%)–NASA | 89.4 | 41.3 | 70.2 |
| Rainfall (mm)–NARL | 103.6 | 0.0 | 1314.5 (Annual) |
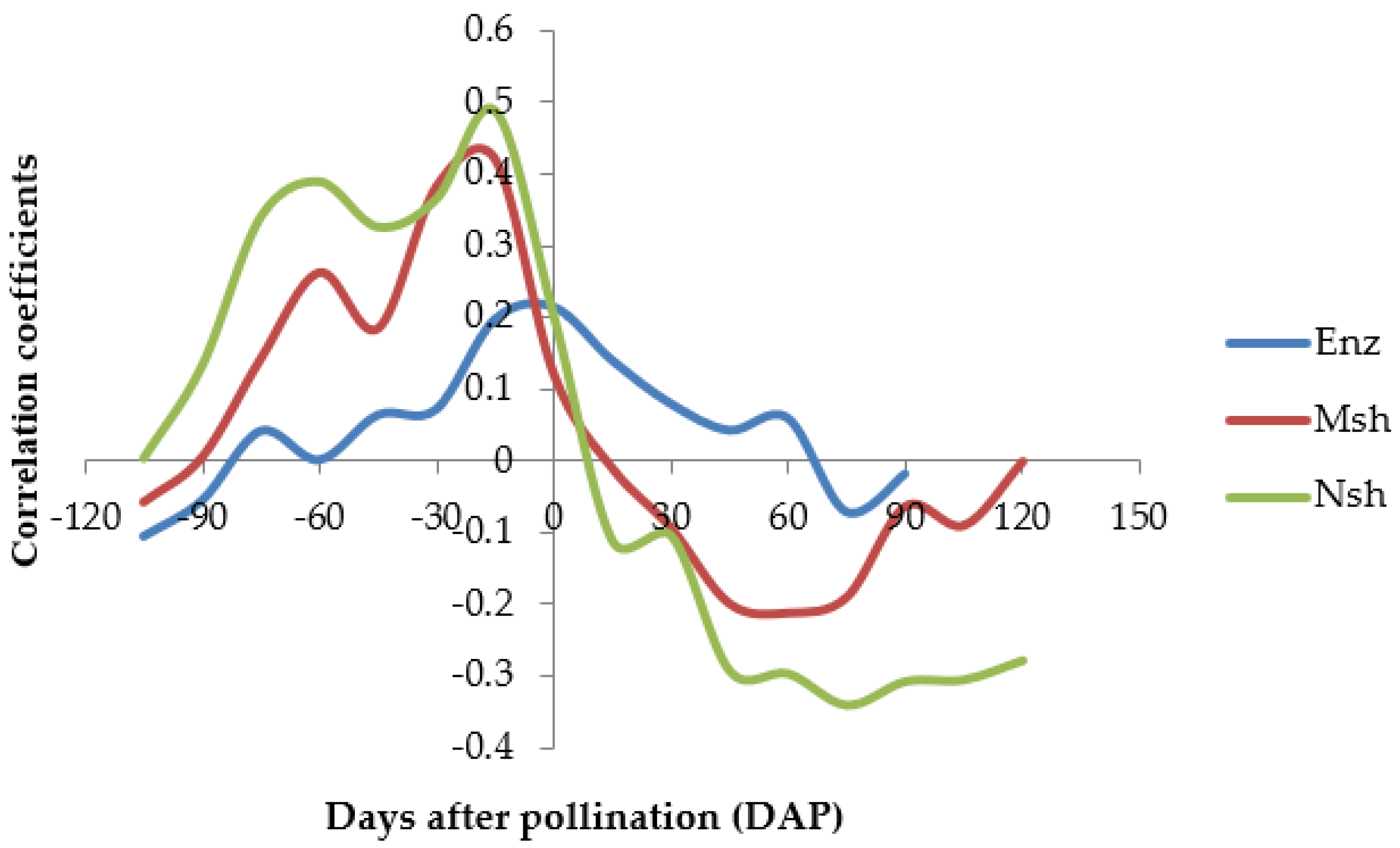
Pearson’s correlation analyses were performed between seed set and weather data at 105, 90, 75, 60, 45, 30, and 15 days before pollination (DBP), 0, 15, 30, 45, 60, 75, 90, 105 and 120 days after pollination (DAP). An assumption was made that one hand opens and is pollinated every day, though up to three hands can open each day [
17]. At the time of pollination (0 DAP), data for weather attributes was therefore averaged for days of bunch pollination to be equivalent to number of hands per bunch. The rest of the 15-day time intervals before and after pollination were 10-day averages of weather attributes counting forward from the designated date. For the cultivar ‘Enzirabahima’, correlation was not run beyond 90 DAP as this was close to its maturity period of 98 days from initial pollination. ‘Mshale’ and ‘Nshonowa’ matured in 131 and 135 DAP respectively. To make meaningful interpretation of results, a correlation analysis between weather attributes was conducted to determine inter-relationships and how they possibly interact to influence seed set. Weather data for inter-attribute relationship were 10-day averages of the weather attributes from initial date of pollination of all pollinated bunches. This included bunches pollinated without PGM though data is not presented as PGM significantly increases seed set [
18]. A principal component analysis (PCA) was run for weather attributes and seed set at the time of pollination to determine the attribute that accounts for most variability in the data. PCA was based on correlation matrix and 10-day averages of the weather attributes was considered at the time of pollination.
A multiple linear regression approach with groups was employed at the 15-day intervals considered from 105 DBP to 120 DAP. The response variable (Y) was seed set per 100 fruits per bunch and the weather attributes (
Table 1) were explanatory variables (X). The analyses were run for each of the three cultivars and bunch size (number of hands) was taken as the grouping factor. Significant weather attributes (
P < 0.05) from the accumulated analysis of variance (ANOVA) tables for all considered time intervals were then combined and reanalyzed in a multiple linear regression. Correlation, PCA, and multiple linear regression analysis were run using Genstat 19th edition developed by VSN International (VSNi).
4. Discussion
Pollination of bananas with PGM stimulates pollen to germinate faster [
14] and yields significantly more seed [
15,
18]. Therefore, this accounts for some error that was not accounted for in previous studies that related weather and seed set in
Musa spp. [
6,
12]. Previous studies also considered only the time of pollination to relate weather and seed set [
6,
12], yet weather influences seed set before and after pollination. The different weather requirements at different floral development stages reflect unique physiological processes. Vegetative growth is not critical as revealed by low association of weather with seed set. Having vigorous vegetative growth that results in big bunches may be important at this stage as big bunches are suggested to be more fertile [
15,
17,
18]. However, bunch size grouping was significant for only ‘Mshale’ in the current study.
The variable start of weather influence on seed set before pollination suggests genotypic differences. This may be the corresponding period of bunch initiation at the growing apex of the three EAHBs. About 11 leaves are said to be inside the pseudostem when bunch initiation happens [
19]. High temperature has been attributed to proper bunch development [
20], which in turn results in high seed set. It is ironic and unclear why low minimum temperature was required for seed set in Mchare 60 DBP. Bunch initiation time for ‘Enzirabahima’ happens about 45 DBP as high solar radiation and low RH at 3 p.m. had an effect on seed set. The magnitude of association was lower in ‘Enzirabahima’ compared to Mchare which suggests more variables involved in seed set for ‘Enzirabahima’.
The non-significant month effects for pollination success observed by Batte et al. [
8] could have been a result of differences in maturity of the EAHBs. These differences may portray same weather conditions having different effects on fertility because of differences in developmental stages even if flowering is at the same time. The month pollination success effects could have therefore been confounded within pooled cultivars as obseverved by Batte et al. [
8]. Some weather effects could not be detected after pooling the three EAHBs (Data not presented). Non-significant month effects for pollination success could have also been as a result of differences in weather of the same months in different years over the 21-year period. Moreover, since pollination success is determined by the presence or absence of seed rather than number of seeds, month effects may have been masked. It may therefore be better to consider number of seed rather than pollination success when evaluating weather effects on fertility in
Musa. Ssebuliba et al. [
7] considered three years and found differences in seed production of EAHBs in different months but years were non-significant.
Bunch initiation time may follow the order of time to maturity with late maturing genotypes having the longest time between likely bunch initiation and time of flowering. The lower association of weather and seed set in ‘Enzirabahima’ 30 DBP may reflect less critical developmental processes. This was also observed in Mchare between 45 and 30 DBP. Peak weather involvement in seed set was about 15 DBP and this may be the most critical stage of bunch developmental stage. This is the likely stage when embryo sac differentiation takes place because low temperature leads to embryo sac malformation [
19], and consequently low seed set. The bunch is likely to be in the vertical position, but it has not cleared the pseudostem, and ovule differentiation is active with meiosis taking place in both diploids and triploids [
21].
Pollinations are usually done between 7 a.m. and 10 a.m. and pollen germinates within three hours. Maximum temperature which is usually after 2 p.m. is therefore less likely to be involved in pollen germination. High temperature has been reported to overcome self-incompatibility in citrus [
22]. In this current study, increase of seed set as a result of high average and maximum temperature at the time pollination may be responsible for overcoming pollen tube inhibition, especially in ‘Enzirabahima’ and ‘Nshonowa’. This may partly be in agreement with observations made by Soares et al. [
23] who observed a necrosis in the prolongation zone of the fruit, it forms much sooner in triploids than in diploids. The necrosis is suggested to arrest pollen tubes before reaching the ovules. However, these results also suggest that pollen tube inhibition are cultivar dependent in diploids as ‘Mshale’ did not need high temperature at the time of pollination. For this reason, temperature is one of the most important weather attributes at the time of pollination for highly sterile genotypes.
The high RH at 9 a.m. required by ‘Mshale’ for increased seed set suggests its requirement for pollen hydration. This may explain observations made by Shepherd [
17] of delayed pollen tube entry into the style by an hour. He also observed increased seed set when pollinated bunches of ‘Gros Michel’ were kept in a moist environment soon after pollination. This favors pollen hydration and consequently germination on the stigma. This particular observation also brings up the question of bagging materials used under different weather conditions. It is a common practice to use polyethylene bags during rainy seasons and cotton bags during the hot seasons as polyethylene scorches the bunch in hot conditions. The way forward would be to use polyethylene bags to create a humid environment around the bunch, especially for pollinations in hot seasons. Creating a shade to avoid scorching of the bunch maybe necessary when polyethylene bags are used.
From 15 to 75 DAP, high temperature in the morning could be critical for physiological processes of embryogenesis and initial seed development in EAHBs. Low solar radiation, low average daily temperature at 2 m, coupled with high RH suggest critical physiological processes necessary for seed development. Since average temperature is negatively correlation with RH, it may be hard to tell whether both temperature and RH play a role in seed set after pollination. Nonetheless, it is likely that these conditions slow down photosynthetic rate which ultimately slows fruit filling rate. These conditions also slow the evapotranspiration rate which results in slow movement of materials in the plant especially from the leaves and roots to the stem and bunch. This may favor resource allocation to the seed rather than the fruit pulp. Interestingly, Shepherd [
17] found that fruit circumference is negatively correlated with seed set in ‘Gros Michel’.
Events that happen after pollination may be responsible for whether seed develops into good or bad seed, seed with or without an embryo and embryo germination rates. Vuylsteke et al. [
9] reported that high embryo germination rates in plantains are obtained following pollination in the wet season. Results from this current study seem to concur with Vuylsteke et al. [
9] as high precipitation after pollination favours seed set in EAHB. However, it has to be noted that high precipitation attributes have to come just after low precipitation. It could also be possible that high rate of fruit filling may have huge implications on seed fertility in
Musa. Matooke has low seed set with poor embryo recovery and germination [
3] and it matures much earlier than Mchare. Ssebuliba et al. [
3] also found differences among Matooke cultivars for percentage hard black seed, embryo recovery and embryo germination. These differences reflect genetic diversity in Matook and could be correlated with days to maturity.
Though maximum temperature and average temperature measurement accounted for the most variability in data, they may not be entirely responsible for seed set in
Musa. Temperature at 9 a.m. and RH at 9 a.m. also contributed to seed set as they were highly ranked in the second and third PCs for the three EAHB. This suggests that banana improvement programs also have to consider manipulating morning temperature and RH after pollination for increased seed set. Maximum and average temperature at 2 m generally had the highest coefficients in the first PCs which suggests careful selection of the pollination station. Shepherd [
17] observed that the pollination station situated at the coast had higher average seed per bunch compared to that in the sheltered valley. However, the few fertile bunches from the pollination station in the sheltered valley were highly fertile. This observation in ‘Gros Michel’ may point to the importance of high temperature before pollination as observed at the coast. Conversely, the importance of low temperature after pollination explains the observations of the pollination station in the sheltered valley. Taking into consideration temperature requirements before and after pollination will potentially increase seed set, embryo rescue, and germination rates.
Weather involvement in set seed in EAHBs after pollination suggests ovule abortion and seed development issues which result in fluffy seed. Banana breeding programs may have to devote some efforts into determining the appropriate time of harvest for maximum embryo rescue and germination rates. This may also suggest that using the sinking-floating criteria to screen for seeds with embryos may not be the best method. Only seeds that sink in water are said to contain embryos.
The regression model used could hardly account for variability in all considered development stages in ‘Enzirabahima’ thus there are other major factors that limit seed set. If these unknown factors were identified and addressed, weather effects on seed set in ‘Enzirabahima’ would become more apparent. The highest variability accounted for by regression in ‘Enzirabahima’ was at the time of pollination, which may suggest where the biggest limiting factor(s) occurs. Temperature at the time of pollination is the most limiting factor to seed set in ‘Enzirabahima’, as revealed by correlation and PCA. Using a regression approach was not the most effective way of pinpointing the most critical stage for weather involvement in seed set. ‘Nshonowa’ had the highest percentage variability explained by the regression model across all considered development stages. This suggested that weather effects on seed set in ‘Nshonowa’ are very paramount for seed set.
Combining significant weather attributes from accumulated regression ANOVA tables of considered development stages showed an overall increase in regression accuracy. This confirms weather involvement in seed set before, during, and after pollination. Ortiz and Vuylsteke [
6] only observed 31.7% of variability explained by a simple linear regression of maximum RH and seed set of a cross between plantain ‘Bobby Tannap’ and wild banana ‘Calcutta 4′. The low variability accounted for by the regression model was a result of considering only the time of pollination and only RH in regression model. Moreover, as observed in this study, maximum and average temperature are the most important for seed set rather than RH.
A plot of correlation coefficients of average temperature with seed set against floral developmental stages summarized the general trend required for seed set. However, the likelihood that nature provides these specific conditions is slim. Besides, pollination is a continuous process, and thus most bunches are pollinated when conditions are not favorable for seed set. This consequently resulted in the low correlation and PC coefficients. Moreover, as observed, bunch size has an effect on seed set. Thus, bulking different bunch sizes in the same analysis reduces precision.


 ,
, 

{kind=link}
{kind=link}

