
Adaptive Morphophysiological Features of Neottia ovata (Orchidaceae) Contributing to Its Natural Colonization on Fly Ash Deposits

 ,
, 
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
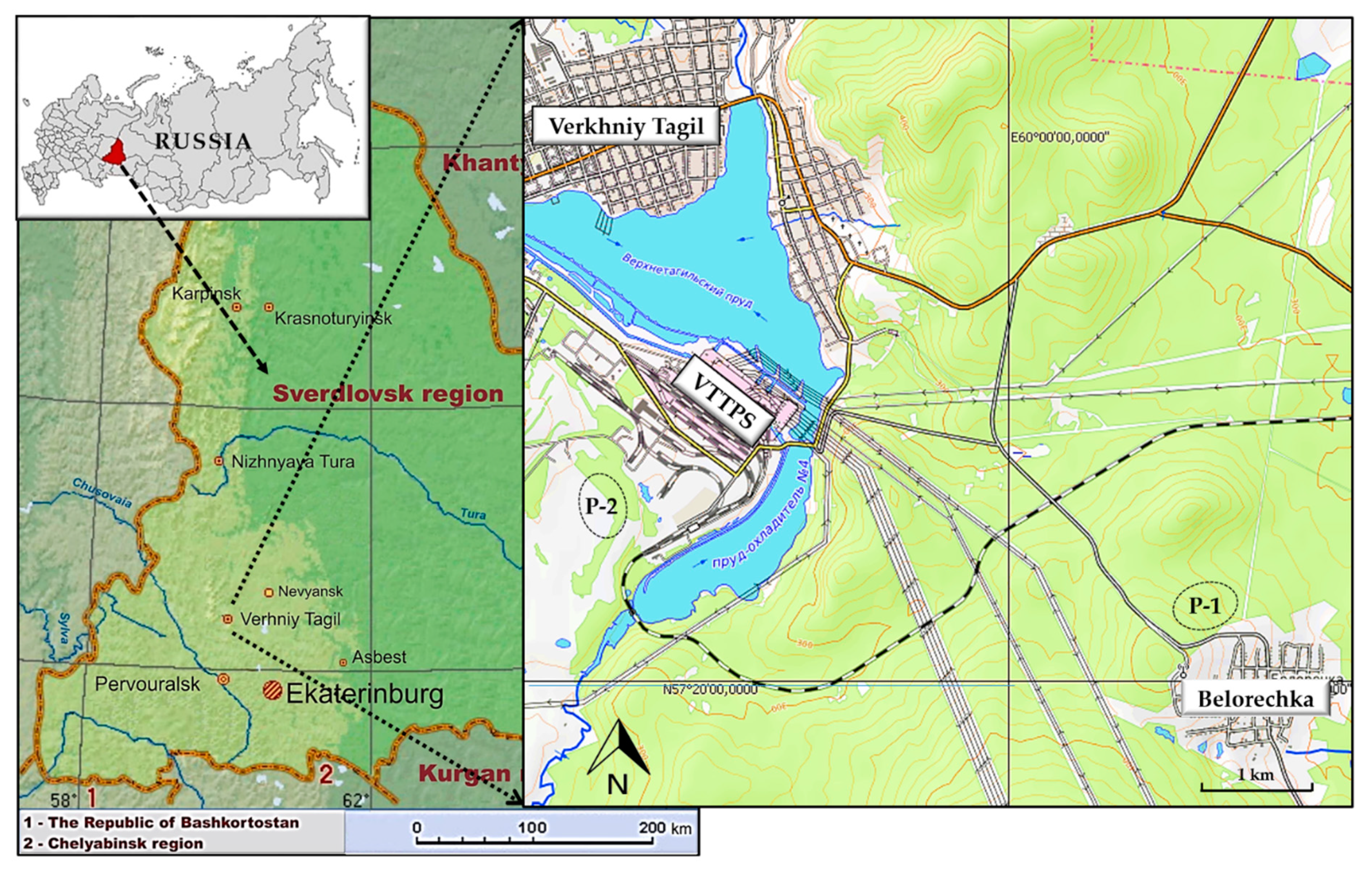
2.1. Study Area
2.2. Plant and Soil Substrate Sample Collection, Preparation and Analysis
2.3. Morphological, Anatomical Parameters and Mycorrhiza Assay
2.4. Physiological and Biochemical Parameters Assay
2.5. Statistical Analysis
3. Results
3.1. Brief Description of Studied Populations
3.2. Soil and Plant Composition
3.2.1. Physicochemical Characteristics and Metal Content in Soil Substrates
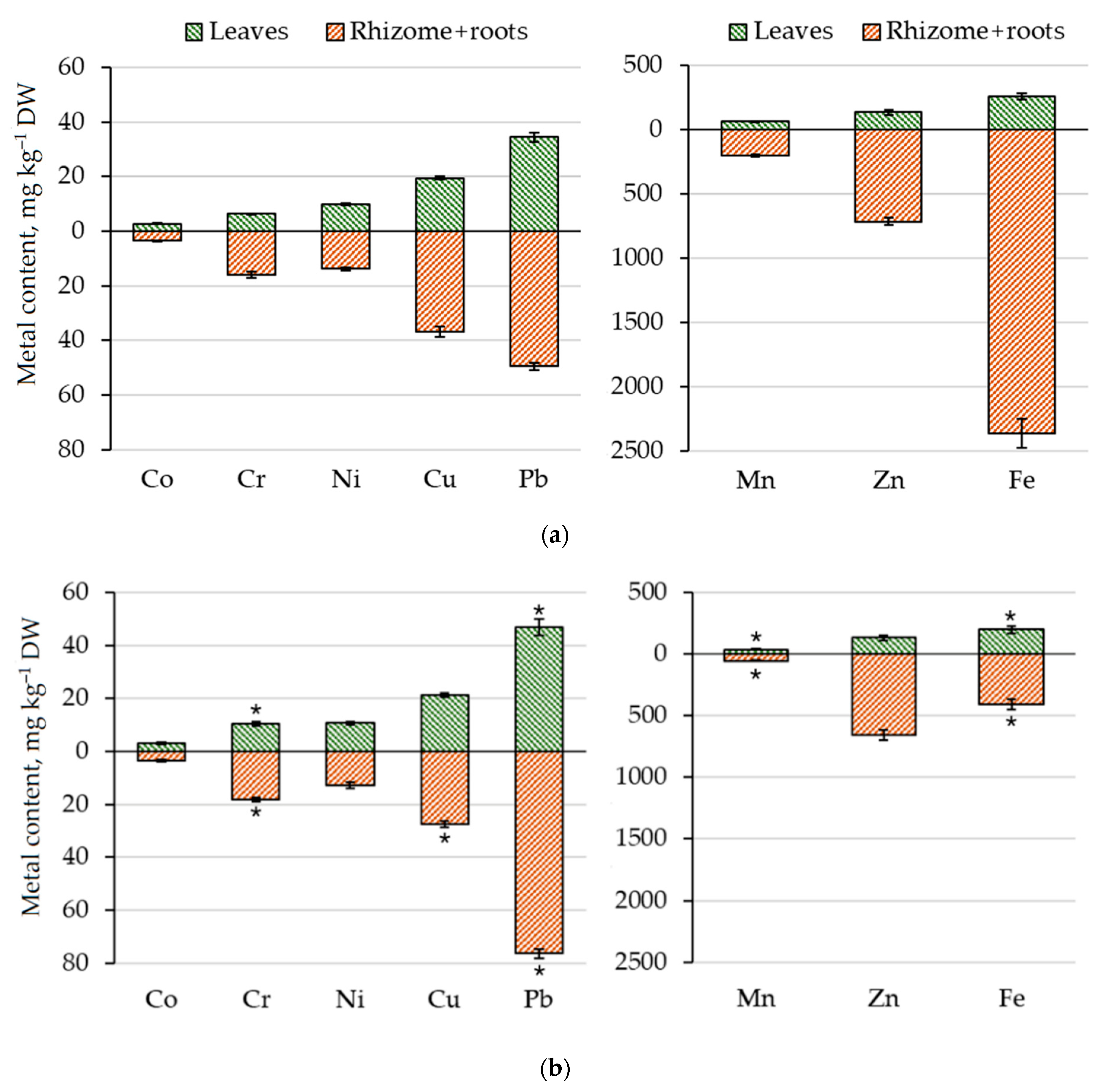
3.2.2. Macronutrient and Metal Content in N. ovata
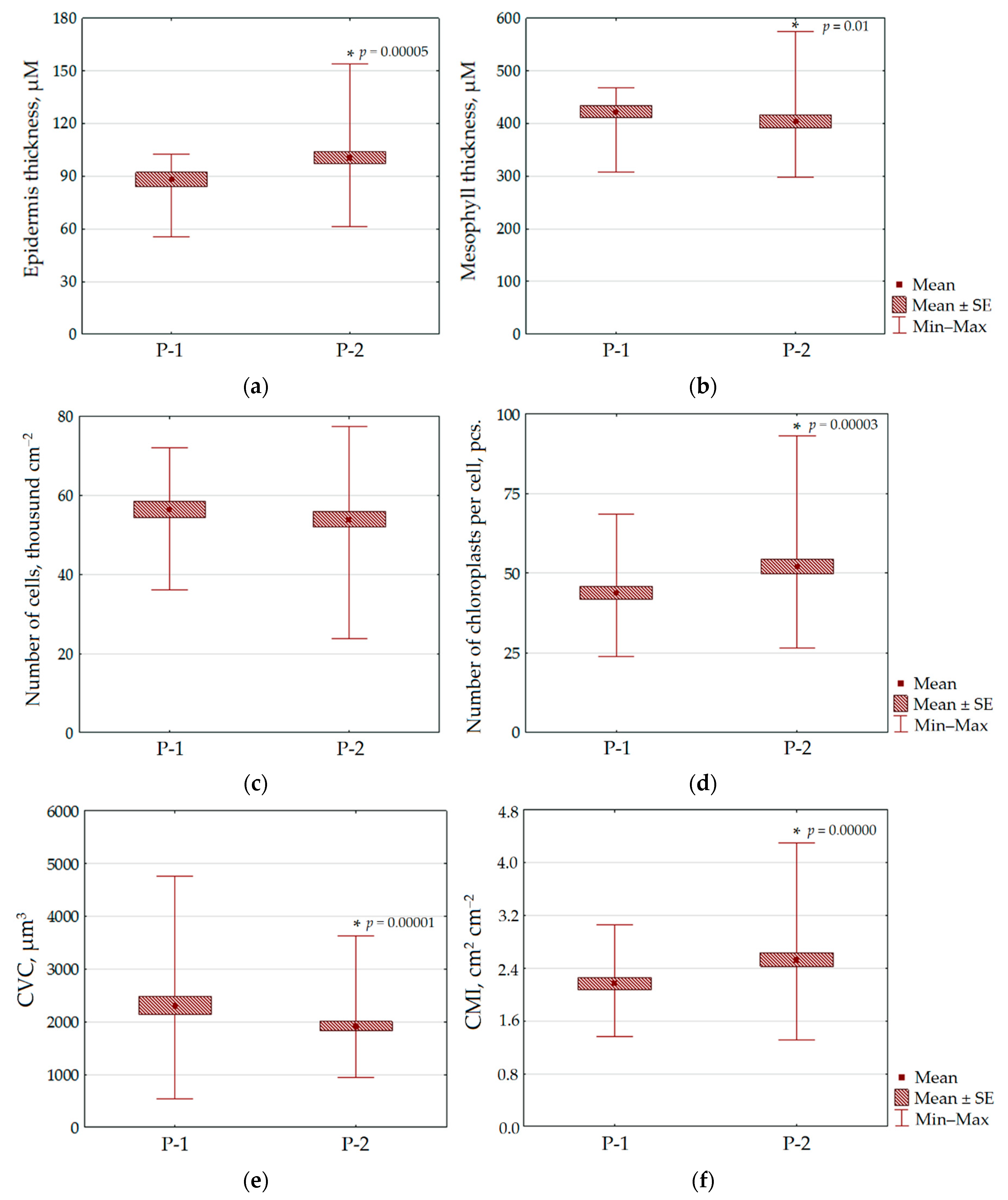
3.3. Morphological and Anatomical Characteristics of N. ovata
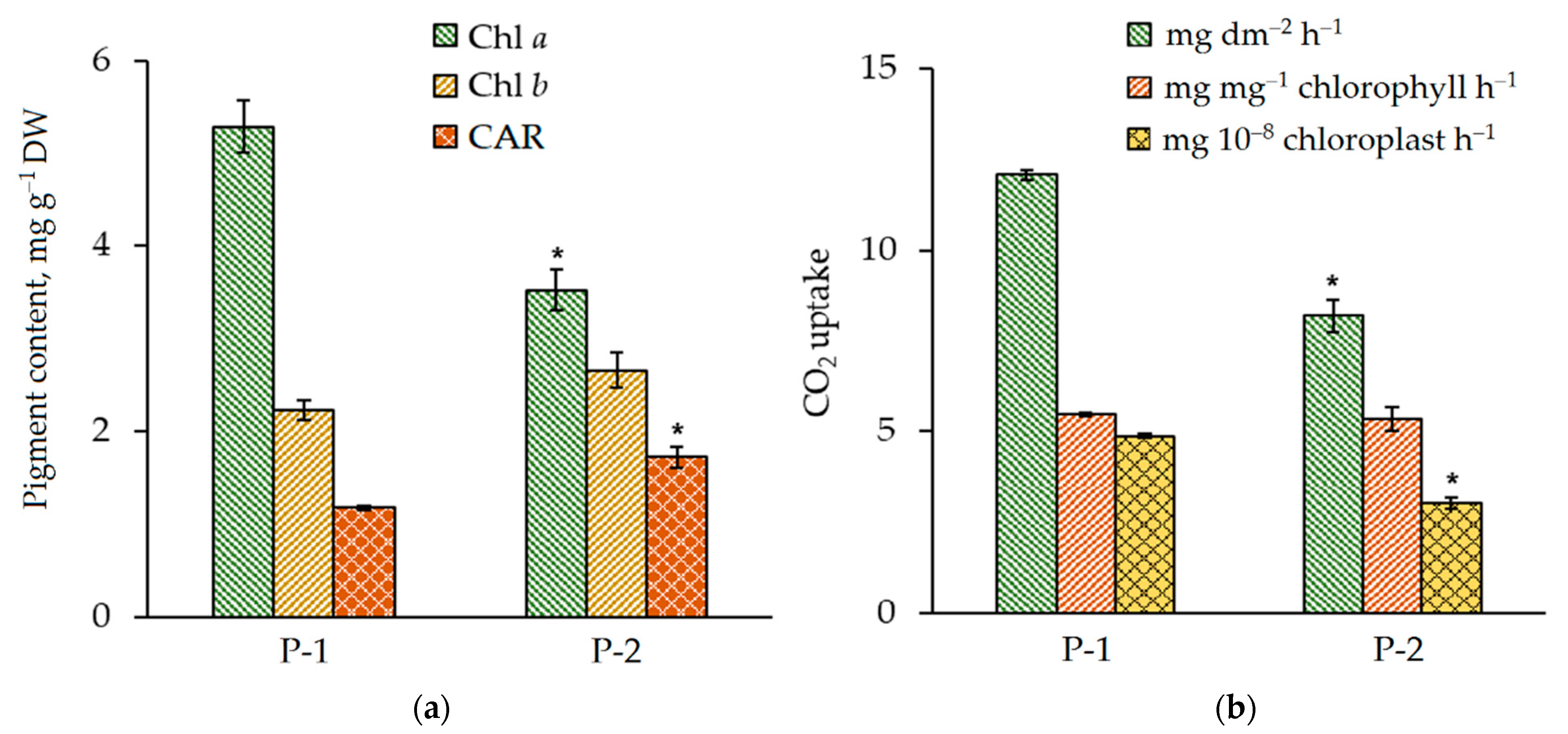
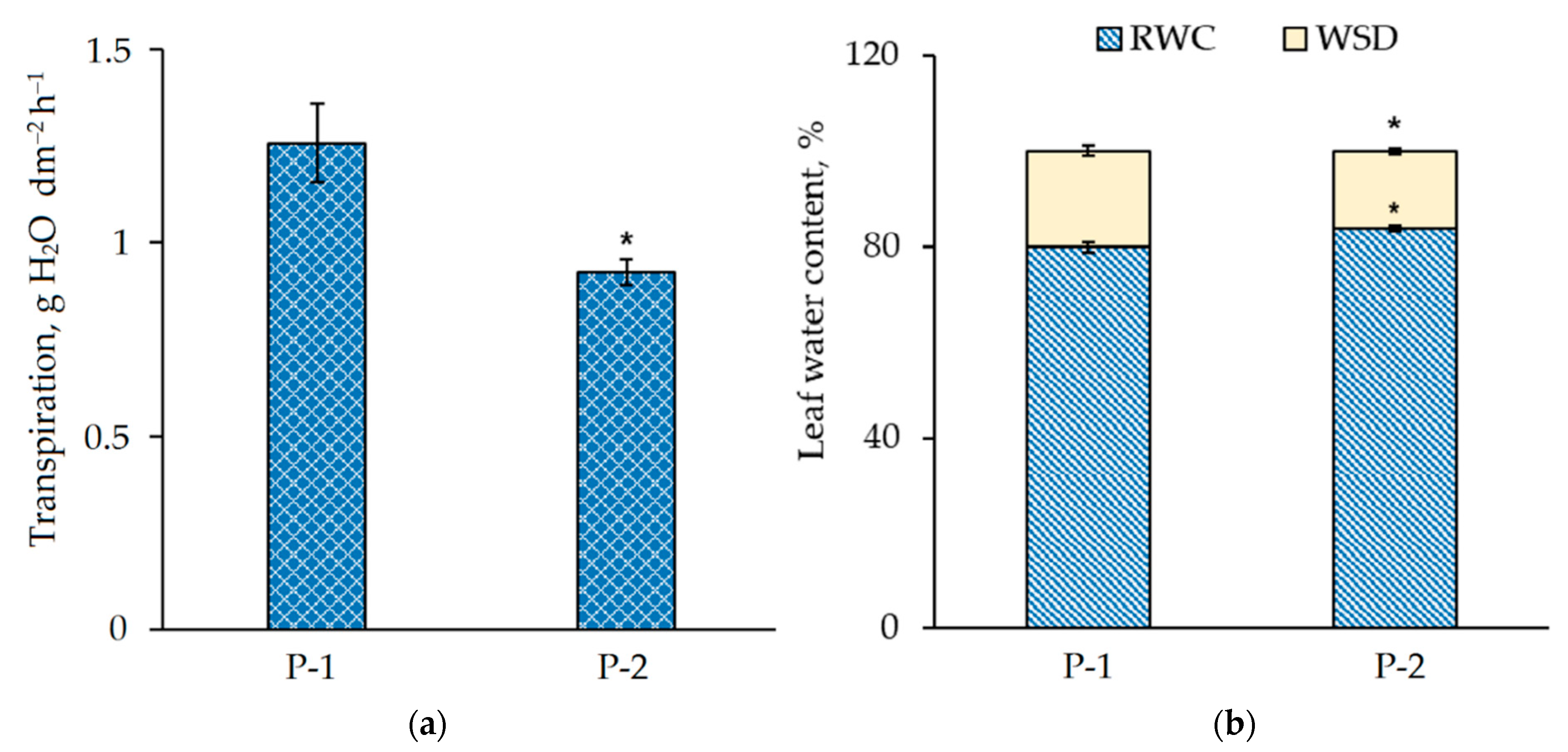
3.4. Physiological and Biochemical Parameters of N. ovata
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Chase, M.W.; Cameron, K.M.; Freudenstein, J.V.; Pridgeon, A.M.; Salazar, G.A.; van den Berg, C.; Schuiteman, A. An updated classification of Orchidaceae. Bot. J. Linn. Soc. 2015, 177, 151–174. [Google Scholar] [CrossRef] [Green Version]
- Christenhusz, M.J.M.; Byng, J.W. The number of known plants species in the world and its annual increase. Phytotaxa 2016, 261, 201–217. [Google Scholar] [CrossRef] [Green Version]
- Vakhrameeva, M.G.; Tatarenko, I.V. Ecological characteristics of orchids of European Part of Russia. Acta Univ. Wratislav. 2001, 79, 49–54. [Google Scholar]
- Vakhrameeva, M.G.; Tatarenko, I.V.; Varlygina, T.I.; Torosyan, G.K.; Zagulskii, M.N. Orchids of Russia and Adjacent Countries (within the Borders of the Former USSR); A.R.G. Gantner Verlag: Ruggell, Liechtenstein, 2008. [Google Scholar]
- Kindlmann, P.; Jersakova, J. Floral display, reproductive success, and conservation of terrestrial orchids. Selbyana 2005, 26, 136–144. [Google Scholar] [CrossRef]
- Sasamori, M.H.; Endres, D.J.; Droste, A. Asymbiotic culture of Cattleya intermedia Graham (Orchidaceae): The influence of macronutrient salts and sucrose concentrations on survival and development of plantlets. Acta Bot. Bras. 2015, 29, 292–298. [Google Scholar] [CrossRef] [Green Version]
- Einzmann, H.J.R.; Schickenberg, N.; Zotz, G. Variation in root morphology of epiphytic orchids along small–scale and large–scale moisture gradients. Acta Bot. Bras. 2020, 34, 66–73. [Google Scholar] [CrossRef]
- Brodmann, J.; Twele, R.; Francke, W.; Hölzler, G.; Zhang, Q.H.; Ayasse, M. Orchids mimic green–leaf volatiles to attract prey–hunting wasps for pollination. Curr. Biol. 2008, 18, 740–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brys, R.; Jacquemyn, H.; Hermy, M. Pollination efficiency and reproductive patterns in relation to local plant density, population size, and floral display in the rewarding Listera ovata (Orchidaceae). Bot. J. Linn. Soc. 2008, 157, 713–721. [Google Scholar] [CrossRef]
- Jakubska–Busse, A.; Kadej, M. The pollination of Epipactis Zinn, 1757 (Orchidaceae) species in central Europe—the significance of chemical attractants, floral morphology and concomitant insects. Acta Soc. Bot. Pol. 2011, 80, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Ehlers, B.K.; Pedersen, H. Genetic variation in three species of Epipactis (Orchidaceae): Geographic scale and evolutionary inferences. Biol. J. Linn. Soc. 2000, 69, 411–430. [Google Scholar] [CrossRef]
- Hens, H.; Jäkäläniemi, A.; Tali, K.; Efimov, P.; Kravchenko, A.V.; Kvist, L. Genetic structure of a regionally endangered orchid, the dark red helleborine (Epipactis atrorubens) at the edge of its distribution. Genetica 2017, 145, 209–221. [Google Scholar] [CrossRef] [Green Version]
- Jurkiewicz, A.; Turnau, K.; Mesjasz–Przybylowicz, J.; Przybylowicz, W.; Godzik, B. Heavy metal localisation in mycorrhizas of Epipactis atrorubens (Hoffm.) Besser (Orchidaceae) from zink mine tailings. Protoplasma 2001, 218, 117–124. [Google Scholar] [CrossRef]
- Shefferson, R.; Kull, T.; Tali, K. Mycorrhizal interactions of orchids colonizing Estonian mine tailings hills. Am. J. Bot. 2008, 95, 156–164. [Google Scholar] [CrossRef]
- Tešitelová, T.; Tešitel, J.; Jersáková, J.; Říhová, G.; Selosse, M.S. Symbiotic germination capability of four Epipactis species (Orchidaceae) is broader than expected from adult ecology. Am. J. Bot. 2012, 99, 1020–1032. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, Y.; Li, J.; Qin, J.; Zhang, W.; Hu, H. Physiological diversity of orchids. Plant Divers. 2018, 40, 196–208. [Google Scholar] [CrossRef]
- Swarts, D.N.; Dixon, W.D. Terrestrial orchid conservation in the age of extinction. Ann. Bot. 2009, 104, 543–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamowski, W. Expansion of native Orchids in anthropogenous habitats. Pol. Bot. Stud. 2006, 22, 35–44. [Google Scholar]
- Jermakowicz, E.; Brzosko, E. Demographic responses of boreal–montane orchid Malaxis monophyllos (L.) Sw. populations to contrasting environmental conditions. Acta Soc. Bot. Pol. 2016, 85, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Rewicz, A.; Bomanowska, A.; Shevera, M.; Kurowski, J.; Krason, K.; Zielinska, K. Cities and disturbed areas as man–made shelters for orchid communities. Not. Bot. Horti Agrobot. Cluj Napoca 2017, 45, 126–139. [Google Scholar] [CrossRef] [Green Version]
- Agostini, A.; Caltagirone, C.; Caredda, A.; Cicatelli, A.; Cogoni, A.; Farci, D.; Guarino, F.; Garau, A.; Labra, M.; Lussu, M.; et al. Heavy metal tolerance of orchid populations growing on abandoned mine tailings: A case study in Sardinia Island (Italy). Ecotoxicol. Environ. Saf. 2020, 189, 110018. [Google Scholar] [CrossRef] [PubMed]
- Filimonova, E.I.; Lukina, N.V.; Glazyrina, M.A. Orchidaceae in the technogenic ecosystems of the Urals. Ecosystems 2014, 11, 68–75. (In Russian) [Google Scholar]
- Filimonova, E.I.; Lukina, N.V.; Glazyrina, M.A.; Borisova, G.G.; Maleva, M.G.; Chukina, N.V. Endangered orchid plant Epipactis atrorubens on serpentine and granite outcrops of Middle Urals, Russia: A comparative morphophysiological study. AIP Conf. Proc. 2019, 040016. [Google Scholar] [CrossRef]
- Filimonova, E.; Lukina, N.; Glazyrina, M.; Borisova, G.; Tripti; Kumar, A.; Maleva, M. A comparative study of Epipactis atrorubens in two different forest communities of the Middle Urals, Russia. J. For. Res. 2020, 31, 2111–2120. [Google Scholar] [CrossRef] [Green Version]
- Kotilínek, M.; Těšitelová, T.; Jersáková, J. Biological Flora of the British Isles: Neottia ovata. J. Ecol. 2015, 103, 1354–1366. [Google Scholar] [CrossRef]
- Vladimir Nikolaevich, B.; Nikolai Sergeevich, K. Red Book of the Sverdlovsk Region: Animals, Plants, Mushrooms; Mir LLC: Yekaterinburg, Russia, 2018. (In Russian) [Google Scholar]
- Wiegand, K.M.A. Revision of the Genus Listera. J. Torrey Bot. Soc. 1899, 26, 157–171. [Google Scholar] [CrossRef]
- Brzosko, E. The dynamics of Listera ovata populations on mineral islands in the Biebrza National Park. Acta Soc. Bot. Pol. 2002, 71, 243–251. [Google Scholar] [CrossRef] [Green Version]
- Gajic´, G.; Djurdjevic´, L.; Kostic, O.; Jaric´, S.; Mitrovic´, M.; Pavlovic, P. Ecological potential of plants for phytoremediation and ecorestoration of fly ash deposits and mine wastes. Front. Environ. Sci. 2018, 6, 124. [Google Scholar] [CrossRef] [Green Version]
- Pandey, V.C.; Prakash, P.; Bajpai, O.; Kumar, A.; Singh, N. Phytodiversity on fly ash deposits: Evaluation of naturally colonized species for sustainable phytorestoration. Environ. Sci. Pollut. Res. 2015, 22, 2776–2787. [Google Scholar] [CrossRef]
- Filimonova, E.I.; Lukina, N.V.; Glazyrina, M.A.; Veselkin, D.V.; Chibrik, T.S.; Stephanovich, G.S. Morphological features of juvenile Pinus sylvestris L. on the ash dumps in the Middle Urals. AIP Conf. Proc. 2019, 030008. [Google Scholar] [CrossRef]
- Mokronosov, A.T. Mesotructure and functional activity of the photosynthetic apparatus. In Mesotructure and Functional Activity of the Photosynthetic Apparatus; Mokronosov, A.T., Ed.; Ural. Gos. Univ.: Sverdlovsk, Russia, 1978; pp. 5–15. (In Russian) [Google Scholar]
- Mokronosov, A.T.; Gavrilenko, V.F.; Zhigalova, T.V. Photosynthesis. Physiological and Environmental and Biochemical Aspects; Academia: Moscow, Russia, 2006. (In Russian) [Google Scholar]
- Ivanova, L.A.; P’yankov, V.I. Structural adaptation of the leaf mesophyll to shading. Russ. J. Plant Physiol. 2002, 49, 419–431. [Google Scholar] [CrossRef]
- Ivanova, L.A. Adaptive features of leaf structure in plants of different ecological groups. Russ. J. Ecol. 2014, 45, 107–115. [Google Scholar] [CrossRef]
- Terashima, I.; Miyazawa, S.; Hanba, Y.T. Why are sun leaves thicker than shade leaves?—Consideration based on analyses of CO2 diffusion in the leaf. J. Plant Res. 2001, 114, 93–105. [Google Scholar] [CrossRef]
- Slaton, M.R.; Smith, W.K. Mesophyll architecture and cell exposure to intercellular air space in alpine, desert, and forest species. Int. J. Plant Sci. 2002, 163, 937–948. [Google Scholar] [CrossRef]
- Nekrasova, O.; Radchenko, T.; Filimonova, E.; Lukina, N.; Glazyrina, M.; Dergacheva, M.; Uchaev, A.; Betekhtina, A. Natural forest colonisation and soil formation on ash dump in southern taiga. Folia For. Pol. Ser. A For. 2020, 62, 306–316. [Google Scholar] [CrossRef]
- Polley, J.R. Colorimetric determination of nitrogen in biological materials. Anal. Chem. 1954, 26, 1523–1524. [Google Scholar] [CrossRef]
- Fiske, C.H.; Subbarow, Y. The colorimetric determination of phosphorus. J. Biol. Chem. 1925, 66, 375–400. [Google Scholar] [CrossRef]
- Hellmuth, E.O. Measurement of Leaf Water Deficit with Particular Reference to the Whole Leaf Method. J. Ecol. 1970, 58, 409–417. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. [34] Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Pandey, V.C.; Abhilashb, P.C.; Upadhyaya, R.N.; Tewari, D.D. Application of fly ash on the growth performance and translocation of toxic heavy metals within Cajanus cajan L.: Implication for safe utilization of fly ash for agricultural production. J. Hazard. Mater. 2009, 166, 255–259. [Google Scholar] [CrossRef]
- Kabata–Pendias, A.; Mukherjee, A.B. Trace Elements from Soil to Human; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Academic Press: San Diego, CA, USA, 1995. [Google Scholar]
- Hejcman, M.; Schellberg, J.; Pavlu, V. Dactylorhiza maculata, Platanthera bifolia and Listera ovata survive N application under P limitation. Acta Oecol. 2010, 36, 684–688. [Google Scholar] [CrossRef]
- Van Arendonk, J.J.C.M.; Poorter, H. The chemical composition and anatomical structure of leaves of grass species differing in relative growth rate. Plant Cell Environ. 1994, 17, 963–970. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Golovko, T.; Tabalenkova, G. Pigments and productivity of the crop plants. In Photosynthetic Pigments: Chemical Structure, Biological Function and Ecology; Golovko, T.K., Gruszecki, W.I., Prasad, M.N.V., Strzalka, K.J., Eds.; Komi Scientific Centre of the Ural Branch of the Russian Academy of Sciences: Syktyvkar, Russia, 2014; pp. 207–220. [Google Scholar]
- Gruszecki, W.; Szymanska, R.; Fiedor, L. Carotenoids as photoprotectors. In Photosynthetic Pigments: Chemical Structure, Biological Function and Ecology; Golovko, T.K., Gruszecki, W.I., Prasad, M.N.V., Strzalka, K.J., Eds.; Komi Scientific Centre of the Ural Branch of the Russian Academy of Sciences: Syktyvkar, Russia, 2014; pp. 161–170. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | pH (H2O) | Electrical Conductivity (EC), µS cm−1 | Total Dissolved Solids (TDS), mg L−1 |
|---|---|---|---|
| NFC | 5.80 ± 0.04 1 | 162.8 ± 11.3 | 89.9 ± 6.1 |
| (5.62–6.34) 2 | (108.0–280.0) | (51.0–145.0) | |
| FAD | 6.19 ± 0.03 * (5.79–6.32) | 96.6 ± 7.2 * (46.0–146.0) | 48.0 ± 3.7 * (23.0–73.0) |
| Metal | Total Content, mg kg−1 DW | Available Content, mg kg−1 DW | ||
|---|---|---|---|---|
| NFC | FAD | NFC | FAD | |
| K | 948.8 ± 87.1 1 | 1256.2 ± 75.8 * | 211.6 ± 5.6 | 310.9 ± 3.8 * |
| (735.8–1162.1) 2 | (1080.8–1450.6) | (198.0–225.4) | (290.6–326.3) | |
| Ca | 16,779.6 ± 856.1 | 8369.1 ± 437.2 * | 6644.8 ± 534.3 | 4439.9 ± 480.9 * |
| (14,690.2–18,876.0) | (7286.3–10676.0) | (5395.9–7823.9) | (3098.7–6351.4) | |
| Mg | 5321.0 ± 329.9 | 2708.0 ± 602.6 * | 1264.1 ± 47.2 | 792.4 ± 139.6 * |
| (4515.1–6127.2) | (999.8–4368.0) | (1149.0–1379.2) | (411.7–1243.8) | |
| Fe | 36,273.4 ± 1472.1 | 33,087.7 ± 3008.1 | 4810.1 ± 214.8 | 2592.9 ± 358.0 * |
| (32,670.0–39877.8) | (24,256.6–41777.9) | (4288.5–5332.0) | (1601.5–3705.0) | |
| Zn | 269.9 ± 25.4 | 192.1 ± 10.2 * | 158.4 ±14.4 | 92.7 ± 4.7 * |
| (208.8–332.6) | (159.0–224.1) | (123.3–193.3) | (77.8–111.6) | |
| Mn | 2043.4 ± 153.4 | 572.7 ± 114.6 * | 1388.2 ± 123.8 | 439.9 ± 93.0 * |
| (1694.0–2393.2) | (270.8–957.0) | (1086.4–1690.2) | (166.6–712.5) | |
| Pb | 152.6 ± 7.8 | 180.0 ± 5.1 * | 69.8 ± 7.2 | 86.2 ± 3.0 * |
| (133.8–171.3) | (158.8–196.7) | (58.2–81.3) | (74.3–98.2) | |
| Cu | 110.3 ± 5.4 | 69.2 ± 8.4 * | 83.1 ± 5.4 | 39.5 ± 3.7 * |
| (98.1–122.4) | (42.1–98.2) | (70.2–96.5) | (26.8–51.9) | |
| Ni | 26.2 ± 4.1 | 22.9 ± 4.9 | 8.5 ± 0.6 | 4.5 ± 0.6 * |
| (16.8–35.5) | (9.1–37.5) | (6.9–10.0) | (2.8–7.8) | |
| Cr | 41.7 ± 1.9 | 63.3 ± 3.1 * | 4.8 ± 0.4 | 8.1 ± 0.5 * |
| (37.6–45.9) | 49.5–72.8 | (4.1–5.6) | (5.9–10.1) | |
| Co | 5.4 ± 0.5 | 7.5 ± 0.9 | 3.4 ± 0.4 | 4.5 ± 0.4 |
| (3.9–7.2) | (4.3–11.5) | (2.3–4.3) | (3.1–6.1) | |
| Macronutrient | Leaves, mg g−1 DW | Rhizome + Roots, mg g−1 DW | ||
|---|---|---|---|---|
| P-1 | P-2 | P-1 | P-2 | |
| N | 42.84 ± 1.30 1 | 23.08 ± 2.32 * | 29.05 ± 3.92 | 28.15 ± 3.13 |
| P | 4.48 ± 0.04 | 5.48 ± 0.15 * | 3.96 ± 0.03 | 2.92 ± 0.25 * |
| K | 33.00 ± 0.61 | 46.97 ± 0.73 * | 9.13 ± 0.36 | 12.70 ± 1.37 * |
| Ca | 19.17 ± 0.28 | 18.13 ± 0.98 | 16.41 ± 0.62 | 15.57 ± 0.85 |
| Mg | 2.22 ± 0.08 | 1.54 ± 0.08 * | 1.58 ± 0.09 | 1.32 ± 0.05 |
| Metal | BCF(Aboveground) | BCF(Underground) | TF(Aboveground/Underground) | |||
|---|---|---|---|---|---|---|
| P-1 | P-2 | P-1 | P-2 | P-1 | P-2 | |
| K | 155.94 | 151.09 | 43.13 | 40.86 | 3.62 | 3.70 |
| Ca | 2.89 | 4.08 | 2.47 | 3.51 | 1.17 | 1.16 |
| Mg | 1.75 | 1.95 | 1.25 | 1.67 | 1.41 | 1.17 |
| Fe | 0.05 | 0.08 | 0.49 | 0.16 | 0.11 | 0.48 |
| Zn | 0.85 | 1.39 | 4.50 | 7.11 | 0.19 | 0.20 |
| Mn | 0.04 | 0.08 | 0.15 | 0.13 | 0.31 | 0.62 |
| Pb | 0.49 | 0.54 | 0.71 | 0.82 | 0.70 | 0.66 |
| Cu | 0.23 | 0.54 | 0.44 | 0.70 | 0.53 | 0.77 |
| Ni | 1.17 | 2.40 | 1.62 | 2.87 | 0.72 | 0.84 |
| Cr | 1.34 | 1.30 | 2.99 | 2.24 | 0.45 | 0.58 |
| Co | 0.84 | 0.82 | 1.04 | 0.94 | 0.81 | 0.88 |
| Parameters | Populations | |
|---|---|---|
| P-1 | P-2 | |
| Shoot length, cm | 60.2 ± 7.2 1 | 45.0 ± 2.1 * |
| (46.0–69.5) 2 | (23.0–69.0) | |
| Inflorescence length, cm | 18.8 ± 4.9 | 14.8 ± 1.1 |
| (13.0–28.5) | (7.2–30.0) | |
| Number of flowers, pcs. | 29.0 ± 8.3 | 23.7 ± 2.1 |
| (17.0–45.0) | (7.0–46.0) | |
| Upper leaf area, cm2 | 38.9 ± 8.7 | 48.1 ± 5.8 |
| (12.9–60.6) | (22.8–87.5) | |
| Lower leaf area, cm2 | 33.7 ± 11.9 | 53.6 ± 7.2 |
| (15.1–63.6) | (24.1–101.4) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maleva, M.; Borisova, G.; Chukina, N.; Sinenko, O.; Filimonova, E.; Lukina, N.; Glazyrina, M. Adaptive Morphophysiological Features of Neottia ovata (Orchidaceae) Contributing to Its Natural Colonization on Fly Ash Deposits. Horticulturae 2021, 7, 109. https://doi.org/10.3390/horticulturae7050109
Maleva M, Borisova G, Chukina N, Sinenko O, Filimonova E, Lukina N, Glazyrina M. Adaptive Morphophysiological Features of Neottia ovata (Orchidaceae) Contributing to Its Natural Colonization on Fly Ash Deposits. Horticulturae. 2021; 7(5):109. https://doi.org/10.3390/horticulturae7050109
Chicago/Turabian StyleMaleva, Maria, Galina Borisova, Nadezhda Chukina, Olga Sinenko, Elena Filimonova, Natalia Lukina, and Margarita Glazyrina. 2021. "Adaptive Morphophysiological Features of Neottia ovata (Orchidaceae) Contributing to Its Natural Colonization on Fly Ash Deposits" Horticulturae 7, no. 5: 109. https://doi.org/10.3390/horticulturae7050109