
Monitoring the Green Vegetation Period of Two Narcissus Taxa by Non-Destructive Analysis of Selected Physiological and Morphological Properties

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Plant Growing Conditions
2.3. Measurement and Analysis
- Non-destructive plant-physiology examinations: 15 plants per taxon
- Biomass (weight of plant parts): ‘Dutch Master’: 60 plants, N. poeticus 90 plants per taxon (4 and 6 sampling dates, respectively).
- Leaf and stem length: 50 plants per taxon
2.3.1. Measurement of Plant Physiology Parameters
| F0 | F50 μs, fluorescence intensity at 50 μs |
| Fm | maximal fluorescence intensity |
| FV = Fm − F0 | maximal variable fluorescence |
| Fm/F0 | ratio between the extremes |
| Fv/F0 | ratio between variable and minimal fluorescence |
| Fv/Fm | maximum quantum yield of PSII in a dark-adapted state |
2.3.2. Measurement of Chlorophyll Content
2.3.3. Measuring of the Leaf and Scape Length
2.3.4. Measuring of the Weight of Plant Organs
2.4. Data Processing, Statistical Methods
3. Results and Discussion
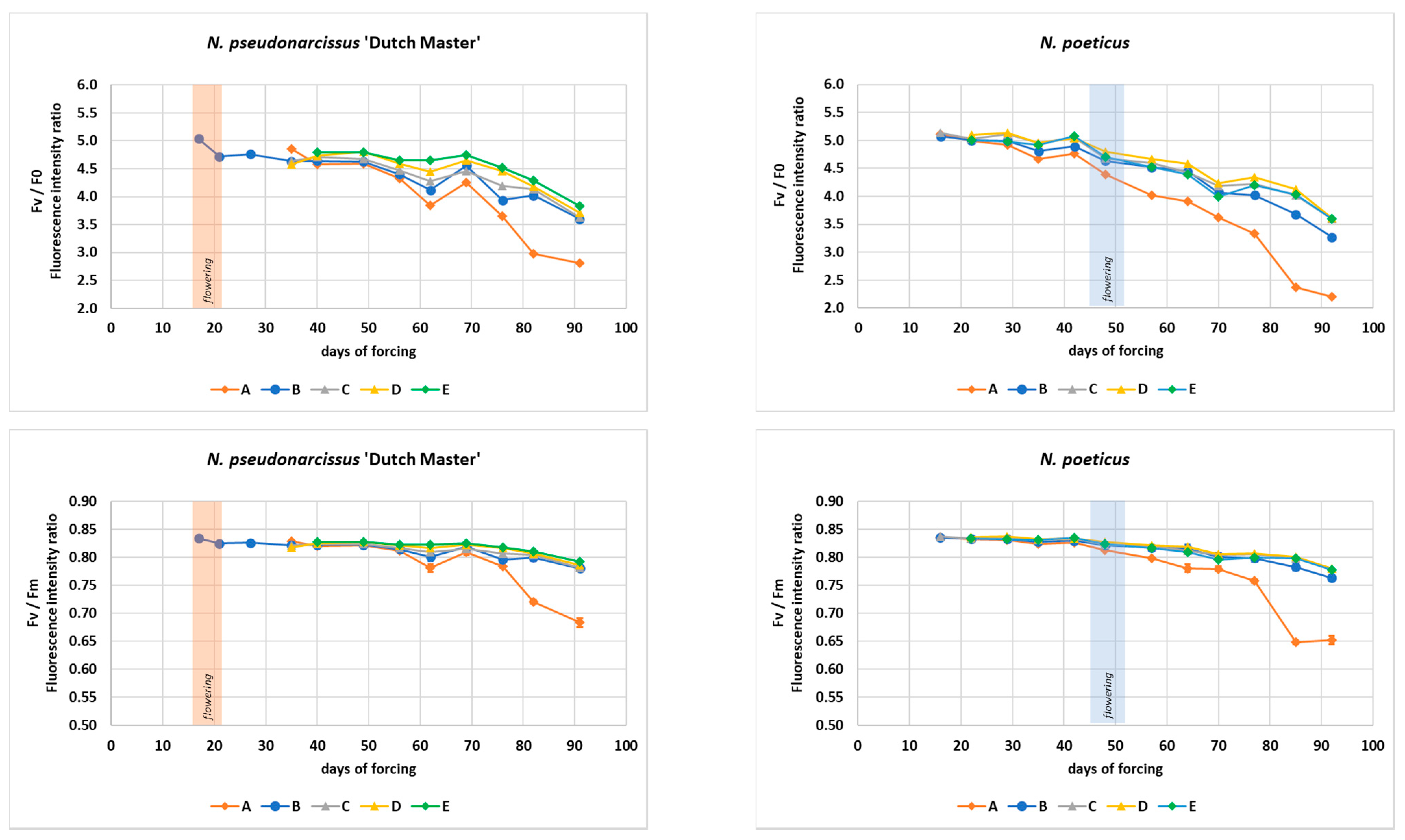
3.1. Plant Physiology Processes
| Fm/F0: |
| N. poeticus: y = −0.0002x2 + 0.0037x + 6.1345, R2 = 0.9696 |
| N. pseudonarcissus ‘Dutch Master’: y = −0.0001x2 − 0.0008x + 5.9995, R2 = 0.9134. |
| Fv/F0: |
| N. poeticus: y = −0.0002x2 + 0.0037x + 5.1345, R2 = 0.9696 |
| N. pseudonarcissus ‘Dutch Master’: y = −0.0001x2 − 0.0008x + 4.9995, R2 = 0.9134 |
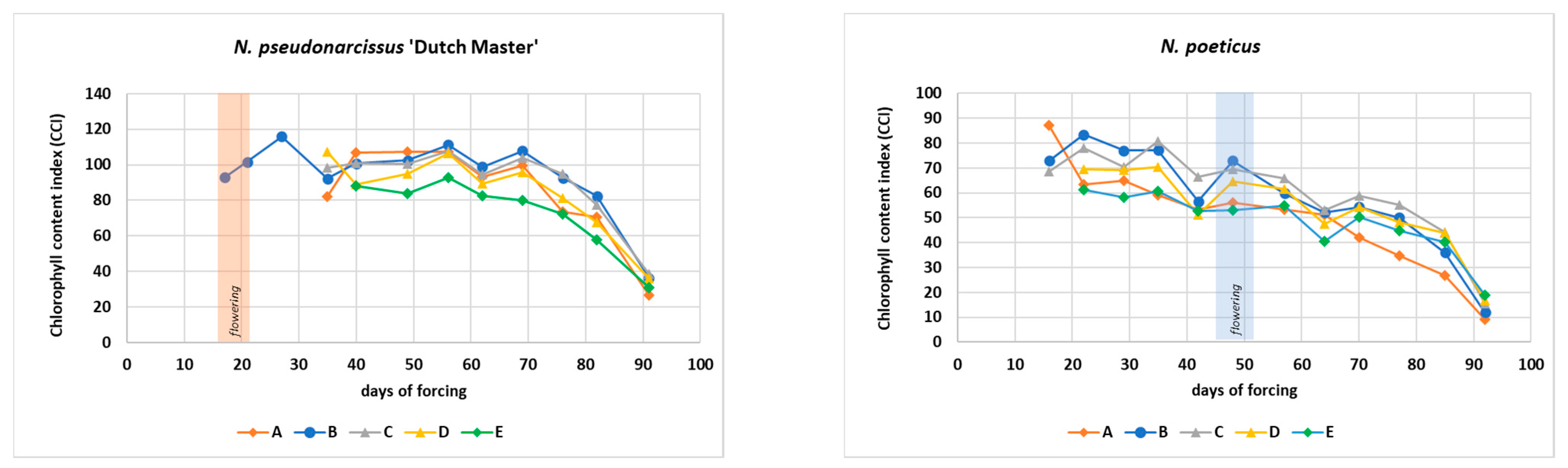
3.2. Chlorophyll Content
3.3. Fresh and Dry Weight of Plant Organs
3.3.1. Fresh Weight of the Leaves and Scape
3.3.2. Dry Mass of the Plant Parts
3.4. Leaf and Scape Length
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dole, J.M.; Wilkins, H.F. Floriculture. Principles and Species, 2nd ed.; Pearson/Prentice Hall: Upper Saddle River, NJ, USA, 2005; pp. 688–696. ISBN 0-13-046250-0. [Google Scholar]
- Tarakemeh, A.; Azizi, M.; Rowshan, V.; Salehi, H.; Spina, R.; Dupire, F.; Arouie, H.; Laurain-Mattar, D. Screening of Amaryllidaceae alkaloids in bulbs and tissue cultures of Narcissus papyraceus and four varieties of N. tazetta. J. Pharm. Biomed. Anal. 2019, 172, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Breiterová, K.; Koutová, D.; Maříková, J.; Havelek, R.; Kuneš, J.; Majorošová, M.; Opletal, L.; Hošťálková, A.; Jenčo, J.; Řezáčová, M.; et al. Amaryllidaceae alkaloids of different structural types from Narcissus L. cv. Professor Einstein and their cytotoxic activity. Plants 2020, 9, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okudo, H.; Sochacki, D. Botanical and horticultural aspects of major ornamental geophytes. In Ornamental Geophytes; Kamenetsky, R., Okubo, H., Eds.; CRC Press: Boca Raton, FL, USA, 2013; pp. 77–122. ISBN 978-1-4398-4924-8. [Google Scholar]
- Hanks, G.R. Commercial production of Narcissus bulbs. In Narcissus and Daffodil. The Genus Narcissus; Hanks, G.R., Ed.; Taylor & Francis: London, UK, 2002; pp. 54–130. ISBN 0-415-27344-7. [Google Scholar]
- Gul, F.; Shahri, W.; Tahir, I. Morphological and some biochemical aspects of flower development and senescence in Narcissus poeticus cv. Pheasant’s Eye. Int. J. Adv. Res. Sci. Eng. Technol. 2018, 7, 1745–1752. [Google Scholar]
- Le Nard, M.; De Hertogh, A.A. Bulb growth and development of flowering. In The Physiology of Flower Bulbs; De Hertogh, A.A., Le Nard, M., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 29–43. ISBN 0-444-87498-4. [Google Scholar]
- Hanks, G.R. Narcissus. In The Physiology of Flower Bulbs; De Hertogh, A.A., Le Nard, M., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 462–558. ISBN 0-444-87498-4. [Google Scholar]
- Kudo, G.; Ida, T.I.; Tani, T. Linkages between phenology, pollination, photosynthesis, and reproduction in deciduous forest understory plants. Ecology 2008, 2, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Veatch-Blohm, M.E.; Chen, D.; Hassett, M. Narcissus Cultivar differences in response to saline irrigation when application began either pre- or postemergence. HortScience 2013, 3, 8. [Google Scholar] [CrossRef] [Green Version]
- Veatch-Blohm, M.E.; Sawch, D.; Elia, N.; Pinciotti, D. Salinity tolerance of three commonly planted Narcissus cultivars. HortScience 2014, 9, 7. [Google Scholar] [CrossRef] [Green Version]
- Veatch-Blohm, M.E.; Roche, B.M.; Sweeney, T. The effect of bulb weight on salinity tolerance of three common Narcissus cultivars. Sci. Hortic. 2019, 248, 62–69. [Google Scholar] [CrossRef]
- Peng, C.; Xiaohong, M.; Qiang, S.; Junjie, G.; Yimin, S. Photosynthetic characteristics of two new cultivars of Narcissus. J. Shanghai Jiaotong Univ. Agric. Sci. 2013, 1, 28–33. [Google Scholar]
- Yantao, W.; Xiaohong, M.; Yimin, S. Biological characters and photosynthesis characteristics of Holland Narcissus. J. Shanghai Jiaotong Univ. Agric. Sci. 2009, 6, 619–623. [Google Scholar]
- Bacon, M.A. Water use efficiency in plant biology. In Water Use Efficiency in Plant Biology; Bacon, M.A., Ed.; Blackwell Publishing: Oxford, UK, 2004; pp. 1–26. ISBN 1-4051-1434-7. [Google Scholar]
- Jones, H. What is water use efficiency? In Water Use Efficiency in Plant Biology; Bacon, M.A., Ed.; Blackwell Publishing: Oxford, UK, 2004; pp. 27–41. ISBN 1-4051-1434-7. [Google Scholar]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazár, D. Parameters of photosynthetic energy partitioning. J. Plant Physiol. 2015, 175, 131–147. [Google Scholar] [CrossRef] [PubMed]
- Sestak, Z.; Shiffel, P. Leaf-age related differences in chlorophyll fluorescence (Review). Photosynthetica 1998, 3–4, 347–369. [Google Scholar]
- Prietro, P.; Penuleas, J.; Lluisa, J.; Adensio, A.; Estiarte, M. Effects of long-term experimental night-time warming and drought on photosynthesis, Fv/Fm and stomatal conductance in the dominant species of a Mediterranean shrubland. Acta Physiol. Plant. 2009, 31, 729–739. [Google Scholar] [CrossRef]
- Sharma, D.K.; Andersen, S.B.; Ottosen, C.; Rosenquist, E. Wheat cultivars selected for high Fv/Fm under heat stress maintain high photosynthesis, total chlorophyll, stomatal conductance, transpiration and dry matter. Physiol. Plant. 2015, 153, 284–298. [Google Scholar] [CrossRef] [PubMed]
- Doğru, A.; Çakirlar, H. Effects of leaf age on chlorophyll fluorescence and antioxidant enzymes activity in winter rapeseed leaves under cold acclimation conditions. Braz. J. Bot. 2020, 43, 11–20. [Google Scholar] [CrossRef]
- Haadi-e-vincheh, M.; Naderi, D.; Golparvar, A. Growth and physiological characteristics of Narcissus pseudonarcissus at different nitrogen levels. Int. J. Farming Allied Sci. 2013, 2, 1325–1329. [Google Scholar]
- Padilla, F.M.; de Souza, R.; Peña-Fleitas, M.T.; Gallardo, M.; Giménez, C.; Thompson, R.B. Different responses of various chlorophyll meters to increasing nitrogen supply in sweet pepper. Front. Plant Sci. 2018, 9, 1752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demír, S.; Çelikel, F.G. The effects of ethephon foliar spray on plant height of Narcissus cv. ‘Ice Follies’. Anadolu J. Agr. Sci. 2018, 33, 184–190. [Google Scholar]
- Miller, W.B. Production chain, forcing physiology, and flower production systems. In Ornamental Geophytes; Kamenetsky, R., Okubo, H., Eds.; CRC Press: Boca Raton, FL, USA, 2013; pp. 287–332. ISBN 978-1-4398-4924-8. [Google Scholar]
- Miller, W.B.; Olberg, M.W. Novel ethephon application methods for Narcissus. HortScience 2016, 10, 1245–1250. [Google Scholar] [CrossRef] [Green Version]
- Çelikel, F.G.; Demir, S. Effects of ethephon spray on plant quality and growth parameters of potted Narcissus tazetta. Acta Hortic. 2019, 1263, 439–448. [Google Scholar] [CrossRef]
- Cohen, D.; Sandler Ziv, D.; Fintea, C.; Ion, A.; Forer, I.; Rabinowitch, H.D.; Kamenetsky, R. New varieties of paperwhite Narcissus: Florogenesis and forcing requirements. Isr. J. Plant Sci. 2009, 4, 335–345. [Google Scholar] [CrossRef]
- Woźny, A.; Jerzy, M. Effect of light wavelength on growth and flowering of narcissi forced under short-day and low quantum irradiance conditions. J. Hortic. Sci. Biotechnol. 2007, 6, 924–928. [Google Scholar] [CrossRef]
- Rees, A.R. The Growth of Bulbs: Applied Aspects of the Physiology of Ornamental Bulbous Crop Plant, 1st ed.; Academic Press Inc.: London, UK, 1972; pp. 45–113. [Google Scholar]
- Slezák, K.A.; Mazur, J.; Jezdinský, A.; Kapczyńska, A. Bulb size interacts with lifting term in determining the quality of Narcissus poeticus L. propagation material. Agronomy 2020, 10, 975. [Google Scholar] [CrossRef]
- Addai, I.K. Influence of cultivar or nutrients application on growth, flower production and bulb yield of the common hyacinth. Am. J. Sci. Ind. Res. 2011, 2, 229–245. [Google Scholar] [CrossRef]
- Singh, S.K.; Reddy, K.R. Regulation of photosynthesis, fluorescence, stomatal conductance and water-use efficiency of cowpea (Vigna unguiculata [L.] Walp.) under drought. J. Photochem. Photobiol. B Biol. 2011, 1, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Daymond, A.J.; Tricker, P.J.; Hadley, P. Genotypic variation in photosynthesis in cacao is correlated with stomatal conductance and leaf nitrogen. Biol. Plant. 2011, 1, 99–104. [Google Scholar] [CrossRef]
- Linderson, M.L.; Mikkelsen, T.N.; Ibrom, A.; Lindroth, A.; Ro-Poulsen, H.; Pilegaard, K. Up-scaling of water use efficiency from leaf to canopy as based on leaf gas exchange relationships and the modeled in-canopy light distribution. Agric. For. Meteorol. 2012, 152, 201–211. [Google Scholar] [CrossRef]
- Tomás, M.; Medrano, H.; Pou, A.; Escalona, J.M.; Martorell, S.; Ribas-Carbó, M.; Flexas, J. Water-use efficiency in grapevine cultivars grown under controlled conditions: Effects of water stress at the leaf and whole-plant level. Aust. J. Grape Wine Res. 2012, 2, 164–172. [Google Scholar] [CrossRef]
- Medrano, H.; Tomás, M.; Martorell, S.; Flexas, J.; Hernández, E.; Rosselló, J.; Pou, A.; Escalona, J.M.; Bota, J. From leaf to whole-plant water use efficiency (WUE) in complex canopies: Limitations of leaf WUE as a selection target. Crop. J. 2015, 3, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Vidal, P.; Marinangeli, P.; Medrano, H.; Gulías, J. Water-use efficiency and growth capacity of two Amaryllidaceae species under three different water regimes. Acta Hortic. 2013, 1000, 223–227. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Tang, C.F.; Zhou, S.B.; Wang, Y.P.; Zhang, D.; Wu, G.W.; Chang, L.L. Comparison of the photosynthetic characteristics of four Lycoris species with leaf appearing in autumn under field conditions. Photosynthetica 2012, 50, 570–576. [Google Scholar] [CrossRef] [PubMed]
- Quanchao, L.; Yang, L.; Yaoyu, X.; Ling, L.; Shaobing, H.; Xiaojing, C. Responses of physiological property and chlorophyll fluorescence characteristics of narcissus to salt stress. J. Fujian Agric. For. Univ. (Nat. Sci. Ed.) 2019, 2, 161–167. [Google Scholar]
- Mauromicale, G.; Ierna, A.; Marchese, M. Chlorophyll fluorescence and chlorophyll content in field-grown potato as affected by nitrogen supply, genotype, and plant age. Photosynthetica 2006, 1, 76–82. [Google Scholar] [CrossRef]
- Xu, Z.Z.; Zhou, G.S.; Wang, Y.L.; Han, G.X.; Li, Y.J. Changes in chlorophyll fluorescence in maize plants with imposed rapid dehydration at different leaf ages. J. Plant Growth. Regul. 2008, 27, 83–92. [Google Scholar] [CrossRef]
- Campbell, P.K.E.; Huemmrich, K.F.; Middleton, E.M.; Ward, L.A.; Julitta, T.; Daughtry, C.S.T.; Burkart, A.; Russ, A.L.; Kustas, W.P. Diurnal and seasonal variations in chlorophyll fluorescence associated with photosynthesis at leaf and canopy scales. Remote Sens. 2019, 11, 488. [Google Scholar] [CrossRef] [Green Version]
- Jouzi, H.; Shekafandeh, A. Effect of pre-planting bulbs treatment with soaking in NPK solution on growth and flowering of Narcissus pseudonarcissus cv. Dutch Master. Adv. Environ. Biol. 2011, 8, 2430–2433. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling | DASF * | Root Weight | Bulb Weight | Leaf Weight | Scape Weight | Fresh Weight | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| N. pseudonarcissus ‘Dutch Master’ | |||||||||||
| Start of forcing (SF) | 3 | 0.758 ± 0.028 | aA | 8.814 ± 0.412 | aA | 0.277 ± 0.021 | dB | 0.160 ± 0.007 | d | ||
| Flowering(F) | 18 | 0.620 ± 0.028 | bA | 5.607 ± 0.152 | bA | 1.259 ± 0.052 | bB | 0.727 ± 0.027 | bA | 0.338 ± 0.007 | A |
| F + 4 weeks | 50 | 0.573 ± 0.049 | bA | 5.698 ± 0.215 | bA | 1.684 ± 0.058 | aA | 0.805 ± 0.027 | aA | ||
| Lifting | 98 | 0.457 ± 0.034 | cA | 8.571 ± 0.320 | aA | 1.004 ± 0.072 | cA | 0.346 ± 0.034 | cA | ||
| N. poeticus | |||||||||||
| Start of forcing (SF) | 2 | 0.517 ± 0.032 | aB | 4.621 ± 0.217 | aB | 0.393 ± 0.021 | dA | ||||
| SF + 2 weeks | 16 | 0.526 ± 0.047 | a | 3.595 ± 0.168 | b | 1.058 ± 0.053 | b | ||||
| SF + 4 weeks | 29 | 0.545 ± 0.039 | a | 2.940 ± 0.149 | c | 1.518 ± 0.122 | a | 0.038 ± 0.007 | d | ||
| Flowering(F) | 46 | 0.575 ± 0.034 | aB | 2.848 ± 0.128 | cB | 1.563 ± 0.078 | aA | 0.464 ± 0.022 | aB | 0.147 ± 0.004 | B |
| F + 4 weeks | 74 | 0.454 ± 0.019 | bB | 3.580 ± 0.126 | bB | 1.454 ± 0.063 | aB | 0.272 ± 0.017 | cB | ||
| Lifting | 97 | 0.405 ± 0.026 | cA | 4.553 ± 0.222 | aB | 0.962 ± 0.077 | cA | 0.339 ± 0.041 | bA | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jezdinská Slezák, K.; Jezdinský, A.; Vachůn, M.; Sotolářová, O.; Pokluda, R.; Uher, J. Monitoring the Green Vegetation Period of Two Narcissus Taxa by Non-Destructive Analysis of Selected Physiological and Morphological Properties. Horticulturae 2021, 7, 585. https://doi.org/10.3390/horticulturae7120585
Jezdinská Slezák K, Jezdinský A, Vachůn M, Sotolářová O, Pokluda R, Uher J. Monitoring the Green Vegetation Period of Two Narcissus Taxa by Non-Destructive Analysis of Selected Physiological and Morphological Properties. Horticulturae. 2021; 7(12):585. https://doi.org/10.3390/horticulturae7120585
Chicago/Turabian StyleJezdinská Slezák, Katalin, Aleš Jezdinský, Miroslav Vachůn, Oldřiška Sotolářová, Robert Pokluda, and Jiří Uher. 2021. "Monitoring the Green Vegetation Period of Two Narcissus Taxa by Non-Destructive Analysis of Selected Physiological and Morphological Properties" Horticulturae 7, no. 12: 585. https://doi.org/10.3390/horticulturae7120585