
QTL Analysis of the Content of Some Bioactive Compounds in Brassica rapa L. Grown under Light Culture Conditions

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mapping Populations of Brassica rapa L. Doubled Haploid Lines
- Population DH 30—obtained by crossing Japanese root turnip (Kairyou Hakata) and oilseed yellow sarson (YS-143).
- Population DH 38—is a result of hybridization of pakchoi (Nai Bai Cai) and oilseed yellow sarson (YS-143).
2.2. Growing Plants under Controlled Conditions
2.3. Biochemical Analyses
2.4. Statistical Processing of the Obtained Results and QTL Analysis
3. Results and Discussion
3.1. The Timing of Start of Bolting
3.2. Analysis of the Content of Some Phytochemicals
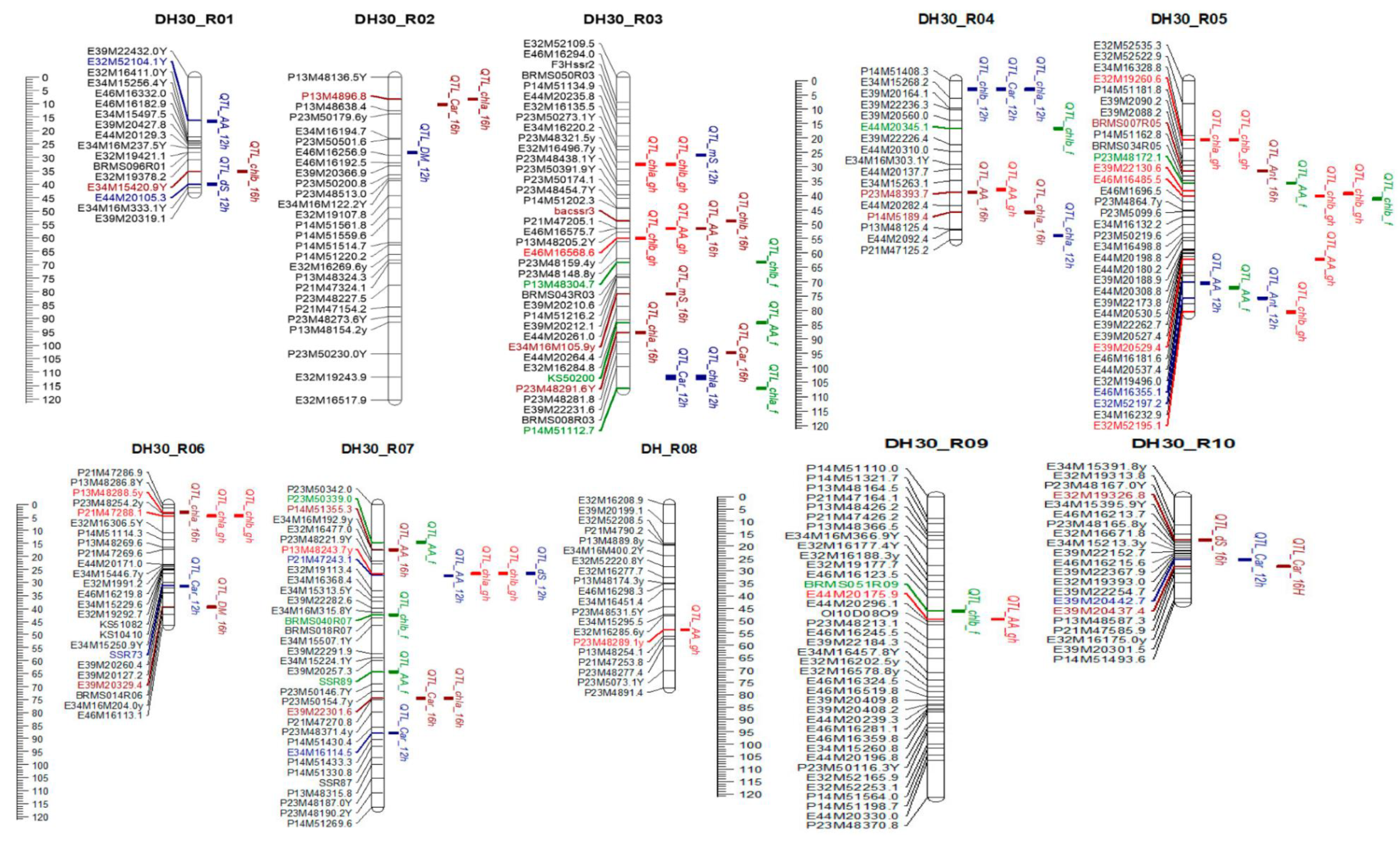
3.3. Quantitative Trait Loci Mapping (QTL Analysis)
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Artemyeva, A.M.; Solovieva, A.E.; Kocherina, N.V.; Rudneva, E.N.; Volkova, E.N.; Chesnokov, Y.V. DNA Marked Brassica rapa L. Double Haploid Lines and Identified QTLs that Control Economically Valuable Traits for Use in Breeding Cabbage Leaf Crops; Catalogue of the VIR Global Collection, Bull. 810; VIR: St. Petersburg, Russia, 2012; 174p. (In Russian) [Google Scholar]
- Artemyeva, A.M.; Solovieva, A.E.; Kocherina, N.V.; Berensen, F.A.; Rudneva, E.N.; Chesnokov, Y.V. Mapping of chromosome loci determined manifestation of morphological and biochemical traits of quality in Brassica rapa L. crops. Russ. J. Plant Physiol. 2016, 63, 259–272. [Google Scholar] [CrossRef]
- Favela-González, K.M.; Hernández-Almanza, A.Y.; De la Fuente-Salcido, N.M. The value of bioactive compounds of cruciferous vegetables (Brassica) as antimicrobials and antioxidants: A review. J. Food Biochem. 2020, 44, e13414. [Google Scholar] [CrossRef] [PubMed]
- Artemyeva, A.M.; Solovieva, A.Y.; Chesnokov, Y.V. Estimating morphological and biochemical characters of quality in doubled haploid lines Brassica rapa L. Vestn. Russ. Agric. Sci. 2014, 3, 38–41, (In Russian with English abstract). [Google Scholar]
- Artemyeva, A.M.; Solovieva, A.E.; Kocherina, N.V.; Chesnokov, Y.V. QTL analysis of biochemical traits of quality in Brassica rapa L. Veg. Crop. Russ. 2014, 1, 10–13, (In Russian with English abstract). [Google Scholar] [CrossRef] [Green Version]
- El-Mogy, M.M.; Mahmoud, A.W.M.; El-Sawy, M.B.Y.; Parmar, A. Pre-harvest foliar application of mineral nutrients to retard chlorophyll degradation and preserve bio-active compounds in broccoli. Agronomy 2019, 9, 711. [Google Scholar] [CrossRef] [Green Version]
- Artemyeva, A.M.; Rudneva, E.N.; Kocherina, N.V.; Chesnokov, Y.V. QTL analysis of morphological traits of quality in Brassica rapa L. Veg. Crop. Russ. 2015, 2, 14–17, (In Russian with English abstract). [Google Scholar] [CrossRef] [Green Version]
- Verma, K.; Tripathi, M.K.; Tiwari, S.; Tripathi, N. Analysis of genetic diversity among Brassica juncea genotypes using morphophysiological and SSR markers. Int. J. Curr. Microbiol. Appl. Sci. 2021, 10, 1108–1117. [Google Scholar] [CrossRef]
- Qi, X.; An, H.; Hall, T.E.; Di, C.; Blischak, P.D.; McKibben, M.T.W.; Hao, Y.; Conant, G.C.; Pires, J.C.; Barker, M.S. Genes derived from ancient polyploidy have higher genetic diversity and are associated with domestication in Brassica rapa. New Phytol. 2021, 230, 372–386. [Google Scholar] [CrossRef]
- Collard, B.C.Y.; Jahufer, M.Z.Z.; Brouwer, J.B.; Pang, E.C.K. An introduction to markers, quantitative trait loci (QTL) mapping and marker-assisted selection for crop improvement: The basic concepts. Euphytica 2005, 142, 169–196. [Google Scholar] [CrossRef]
- Chesnokov, Y.V. Plant Genetic Resources and Modern Methods of DNA-Fingerprinting; VIR: St. Petersburg, Russia, 2007; 80p. (In Russian) [Google Scholar]
- Witzel, K.; Kurina, A.B.; Artemyeva, A.M. Opening the treasure chest: The current status of research on Brassica oleracea and B. rapa vegetables from ex situ germplasm collections. Front. Plant Sci. 2021, 12, 643047. [Google Scholar] [CrossRef] [PubMed]
- Pink, D.; Bailey, L.; McClement, S.; Hand, P.; Mathas, E.; Buchanan-Wollaston, V.; Astley, D.; King, G.; Teakle, G. Double haploids, markers and QTL analysis in vegetable brassicas. Euphytica 2008, 164, 509–514. [Google Scholar] [CrossRef]
- Lou, P.; Woody, S.; Brenham, K.; VanBuren, R.; Colle, M.; Edger, P.P.; Sartor, R.; Zheng, Y.; Levendoski, N.; Lim, J.; et al. Genetic and genomic resources to study natural variation in Brassica rapa. Plant Direct 2020, 4, e00285. [Google Scholar] [CrossRef]
- Cockram, J.; Mackay, I. Genetic mapping populations for conducting high-resolution trait mapping in plants. In Plant Genetics and Molecular Biology; Varshney, R., Pandey, M., Chitikineni, A., Eds.; Advances in Biochemical Engineering/Biotechnology; Springer: Cham, Switzerland, 2018; Volume 164, pp. 109–138. [Google Scholar] [CrossRef]
- Chesnokov, Y.V. Mapping of Quantitative Trait Loci in Plants; VIR: St. Petersburg, Russia, 2009; 100p. (In Russian) [Google Scholar]
- Şahin, Ö.; Kavuncu, O. A Study on relationships between various quantitative characteristics for using availability quantitative trait loci mapping. Selcuk J. Agric. Food Sci. 2019, 33, 42–44. [Google Scholar] [CrossRef]
- Jan, S.A.; Shinwari, Z.K.; Rabbani, M.A. Determining genetic divergence among Brassica rapa ecotypes through electrophoretic mobility of total seed proteins. J. Anim. Plant Sci. 2016, 26, 1758–1764. [Google Scholar]
- Ullah, N.; Khan, J.; Khan, M.W.; Raza, H.; Alam, M.; Ullah, H.; Hussain, I.; Jan, I.U.; Rahman, Z.; Ali, F. Genetic variability for biochemical traits among advanced lines of Brassica. Pure Appl. Biol 2017, 6, 1–8. [Google Scholar] [CrossRef]
- Lou, P.; Zhao, J.; He, H.; Hanhart, C.; Pino del Carpio, D.; Verkerk, R.; Custers, J.; Koornneef, M.; Bonnema, G. Quantitative trait loci for glucosinolates accumulation in Brassica rapa leaves. New Phytol. 2008, 179, 1017–1032. [Google Scholar] [CrossRef]
- Zhao, J.; Paulo, M.J.; Jamar, D.; Lou, P.; van Eeuwijk, F.; Bonnema, G.; Vreugdenhil, D.; Koornneef, M. Association mapping of leaf traits, flowering time, and phytate content in Brassica rapa. Genome 2007, 50, 963–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagheri, H.; El-Soda, M.; Kim, H.K.; Fritsche, S.; Jung, C.; Aarts, M.G. Genetic analysis of health-related secondary metabolites in Brassica rapa recombinant inbred lines population. Int. J. Mol. Sci. 2013, 14, 15561–15577. [Google Scholar] [CrossRef] [Green Version]
- El-Soda, M.; Boer, M.P.; Bagheri, H.; Hanhart, C.J.; Koornneef, M.; Aarts, M.G. Genotype–environment interactions affecting preflowering physiological and morphological traits of Brassica rapa grown in two watering regimes. J. Exp. Bot. 2014, 65, 697–708. [Google Scholar] [CrossRef] [Green Version]
- Kochetov, A.A.; Makarova, G.A.; Mirskaya, G.V.; Sinyavina, N.G. Agrophysical approach to the creation of new forms of cultivated plants. Agrophysics 2012, 1, 40–44, (In Russian with English abstract). [Google Scholar]
- Panova, G.G.; Udalova, O.R.; Kanash, E.V.; Galushko, A.S.; Kochetov, A.A.; Priyatkin, N.S.; Arkhipov, M.V.; Chernousov, I.N. Fundamentals of Physical Modeling “ideal” agroecosystems. Tech. Phys. 2020, 65, 1563–1569. [Google Scholar] [CrossRef]
- He, Y.; Chen, T.; Zeng, X. Genetic and epigenetic understanding of the seasonal timing of flowering. Plant Commun. 2020, 1, 100008. [Google Scholar] [CrossRef]
- Wang, S.L.; An, H.R.; Tong, C.G.; Jang, S. Flowering and flowering genes: From model plants to orchids. Hortic. Environ. Biotechnol. 2021, 62, 135–148. [Google Scholar] [CrossRef]
- Lou, P.; Zhao, J.; Kim, J.S.; Shen, S.; Del Carpio, D.P.; Song, X.; Jin, M.; Vreugdenhil, M.; Wand, X.; Koornneef, M.; et al. Quantitative trait loci for flowering time and morphological traits in multiple populations of Brassica rapa. J. Exp. Bot. 2007, 58, 4005–4016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egorova, K.V.; Sinyavina, N.G.; Kochetov, A.A.; Chesnokov, Y.V. Assessment of significant for breeding morphological traits in the double haploid population of Brassica rapa L. in controlled conditions of a regulated agroecosystem. Veg. Crop. Russ. 2020, 4, 28–31, (In Russian with English abstract). [Google Scholar] [CrossRef]
- Interstate Standard 12038-84. Agricultural seeds. Methods for Determination of Germination; Standartinform Publishing: Moscow, Russia, 2004. (In Russian) [Google Scholar]
- Ermakov, E.I.; Zheltov, Y.I.; Milto, N.E.; Kucherov, V.I. Soil for Growing Plants “Agrophyte”. Russian Federation Patent 2081555, 13 July 1997. (In Russian). [Google Scholar]
- State Standard of Russian Federation 8756. Fruit and Vegetable Products. Methods for Determination of Sugars; Standartinform Publishing: Moscow, Russia, 2010. (In Russian) [Google Scholar]
- State Standard of Russian Federation 24556-89. Products of Fruits and Vegetables Processing. Methods for Determination of Vitamin C; Standartinform Publishing: Moscow, Russia, 2003. (In Russian) [Google Scholar]
- Interstate Standart NF V05-129-1984. Fruit and Vegetable Products. Conventionnal Determination of Authocyanins. Detection and Identification; AFNOR Editions: Paris, France, 1984. [Google Scholar]
- Andersen, O.M.; Markham, K.R. Flavonoids: Chemistry, Biochemistry and Applications; CRC Press: Boca Raton, FL, USA, 2006; 1256p. [Google Scholar]
- Trineeva, O.V.; Safonova, E.F.; Slivkin, A.I.; Voropaeva, S.V. Method of Spectrophotometric Quantitative Determination in the Leaves of Stinging Nettle in the Joint Presence of Chlorophyll, Carotenoids and Hydroxycinnamic Acids. Russian Federation Patent 2531940, 27 October 2014. (In Russian). [Google Scholar]
- Van Ooijen, J.W. MapQTL® 6, Software for the Mapping of Quantitative Trait Loci in Experimental Populations of Diploid Species; Kyazma BV: Wageningen, The Netherlands, 2009; Available online: https://www.kyazma.nl/index.php/mc.MapQTL/ (accessed on 22 November 2021).
- Kocherina, N.V.; Artemyeva, A.M.; Chesnokov, Y.V. Using the technology of LOD score in mapping the quantitative trait loci in plants. Russ. Agric. Sci. 2011, 3, 14–17. [Google Scholar] [CrossRef]
- Ronald, P.S.; Penner, G.A.; Brown, P.D.; Brule-Babel, A. Identification of RAPD markers for percent hull in oat. Genome 1997, 40, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. Available online: https://www.wur.nl/en/show/mapchart.htm (accessed on 22 November 2021). [CrossRef] [PubMed] [Green Version]
- Barba, F.J.; Esteve, M.J.; Frígola, A. Bioactive components from leaf vegetable products. Stud. Nat. Prod. Chem. 2014, 41, 321–346. [Google Scholar] [CrossRef]
- Artemyeva, A.M.; Sinyavina, N.G.; Panova, G.G.; Chesnokov, Y.V. Biological features of Brassica rapa L. vegetable leafy crops when growing in an intensive light culture. Agric. Biol. 2021, 56, 103–120. [Google Scholar] [CrossRef]
- Kapusta-Duch, J.; Kopec, A.; Piatkowska, E.; Borczak, B.; Leszczynska, T. The beneficial effects of Brassica vegetables on human health. Ann. Natl. Inst. Hyg. 2012, 63, 389–395. [Google Scholar]
- Xiao, D.; Shen, H.R.; Zhao, J.J.; Wei, Y.P.; Liu, D.R.; Hou, X.L.; Bonnema, G. Genetic dissection of flowering time in Brassica rapa responses to temperature and photoperiod. Plant Sci. 2019, 280, 110–119. [Google Scholar] [CrossRef]
- Rahman, H.; Bennett, R.A.; Kebede, B. Molecular mapping of QTL alleles of Brassica oleracea affecting days to flowering and photosensitivity in spring Brassica napus. PLoS ONE 2018, 13, e0189723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandal, K.G.; Sinha, A.C. Nutrient management effects on light interception, photosynthesis, growth, dry-matter production and yield of Indian mustard (Brassica juncea). J. Agron. Crop Sci. 2004, 190, 119–129. [Google Scholar] [CrossRef]
- Kurina, A.B.; Kornyukhin, D.L.; Solovyeva, A.E.; Artemyeva, A.M. Genetic diversity of phenotypic and biochemical traits in VIR radish (Raphanus sativus L.) germplasm collection. Plants 2021, 10, 1799. [Google Scholar] [CrossRef] [PubMed]
- Chupakhina, G.N. Plant Ascorbic Acid System: Monograph; Immanuel Kant Baltic Federal University: Kaliningrad, Russia, 1997; 120p. (In Russian) [Google Scholar]
- Arrigoni, O.M.; De Tullio, C. Ascorbic acid: Much more than just an antioxidant. Biochim. Biophys. Acta Gen. Subj. 2002, 1569, 1–9. [Google Scholar] [CrossRef]
- Kim, Y.H.; Lee, J.S. Growth and contents of anthocyanins and ascorbic acid in lettuce as affected by supplemental UV-A LED irradiation with different light quality and photoperiod. Korean J. Hortic. Sci. Technol. 2016, 34, 596–606. [Google Scholar] [CrossRef]
- Welch, C.R.; Wu, Q.; Simon, J.E. Recent advances in anthocyanin analysis and characterization. Curr. Anal. Chem. 2008, 4, 75–101, (In Russian with English abstract). [Google Scholar] [CrossRef] [Green Version]
- Babak, O.G.; Nekrashevich, N.A.; Nikitinskaya, T.V.; Yatsevich, K.K.; Kilchevsky, A.V. Study of the Myb-factor polymorphism based on comparative genomics of vegetable Solanaceae crops (tomato, pepper, eggplant) to search for DNA markers that differentiate samples by the anthocyans accumulation. Dokl. Natl. Acad. Sci. Belarus 2020, 63, 721–729, (In Russian with English abstract). [Google Scholar] [CrossRef]
- Šamec, D.; Urlic, B.; Salopek-Sondi, B. Kale (Brassica oleracea var. acephala) as a superfood: Review of the scientific evidence behind the statement. Crit. Rev. Food Sci. Nutr. 2019, 59, 2411–2422. [Google Scholar] [CrossRef] [PubMed]
- Esteban, R.; Barrutia, O.; Artetxe, U.; Fernandez-Marin, B.; Hernandez, A.; Garcia-Plazaola, J.I. Internal and external factors affecting photosynthetic pigment composition in plants: A meta-analytical approach. New Phytol. 2015, 206, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Lambers, H.; Chapin, F.S.; Pons, T.L. Plant Physiological Ecology; Springer: New York, NY, USA, 2008; p. 623, (In Russian with English abstract). [Google Scholar] [CrossRef]
- Ivanov, L.A.; Ronzhina, D.A.; Yudina, P.K.; Zolotareva, N.V.; Kalashnikova, I.V.; Ivanova, L.A. Seasonal dynamics of the content of chlorophylls and carotenoids in the leaves of steppe and forest plants at the species and community level. Russ. J. Plant Physiol. 2020, 67, 453–462. [Google Scholar] [CrossRef]
- Park, J.E.; Park, Y.G.; Jeong, B.R.; Hwang, S.J. Growth and anthocyanin content of lettuce as affected by artificial light source and photoperiod in a closed-type plant production system. Kor. J. Hort. Sci. Technol. 2012, 30, 673–679. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, F.; Valentão, P.; Sousa, C.; Pereira, J.A.; Seabra, R.M.; Andrade, P.B. Chemical and antioxidative assessment of dietary turnip (Brassica rapa var. rapa L.). Food Chem. 2007, 105, 1003–1010. [Google Scholar] [CrossRef]
- Egorova, K.V.; Dubovitskaya, V.I.; Khomyakov, Y.V.; Chesnokov, Y.V. Study of photoperiod effect on microelements content in cabbage plants of Brassica rapa L. Agrophysica 2021, 2, 37–44, (In Russian with English abstract). [Google Scholar] [CrossRef]
- Raiola, A.; Errico, A.; Petruk, G.; Monti, D.M.; Barone, A.; Rigano, M.M. Bioactive compounds in Brassicaceae vegetables with a role in the prevention of chronic diseases. Molecules 2018, 23, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomas, M.; Zhang, L.; Zengin, G.; Rocchetti, G.; Capanoglu, E.; Lucini, L. Metabolomic insight into the profile, in vitro bioaccessibility and bioactive properties of polyphenols and glucosinolates from four Brassicaceae microgreens. Food Res. Int. 2021, 140, 110039. [Google Scholar] [CrossRef]
- Lander, E.S.; Botstein, D. Mapping mendelian factors underlying quantitative traits using RFLP linkage maps. Genetics 1989, 121, 185–199. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biopoligon of AFI 12 h Photoperiod | Biopoligon of AFI 16 h Photoperiod | VIR, Greenhouse * | VIR, Field * | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Trait | Mean ± SD | Xmin–Xmax | CV, % | Mean ± SD | Xmin–Xmax | CV, % | Mean ± SD | Xmin–Xmax | CV, % | Mean ± SD | Xmin–Xmax | CV, % |
| Days to start of bolting | 46.30 ± 7.63 a | 31.00–63.00 | 16.48 | 38.52 ± 8.51 ab | 26.00–54.00 | 22.08 | 61.80 ± 7.99 abc | 50.20–85.00 | 12.93 | 29.42 ± 4.16 abc | 15.00–37.00 | 14.13 |
| Dry matter (%) | 11.20 ± 3.18 | 7.42–17.96 | 28.39 | 12.41 ± 4.22 | 5.61–23.97 | 34.00 | 9.72 ± 4.15 | 5.84–19.32 | 46.67 | 10.44 ± 1.26 | 7.69–12.28 | 12.02 |
| Total sugar content (mg/100 g) | 1.30 ± 0.69 a | 0.11–2.63 | 54.08 | 1.88 ± 0.77 b | 0.74–3.53 | 40.96 | 0.65 ± 0.57 abc | 0.22–2.37 | 88.63 | 1.62 ± 0.65 c | 0.00–2.41 | 40.12 |
| Monosaccharides (mg/100 g) | 0.82 ± 0.47 | 0.11–2.12 | 57.32 | 1.13 ± 0.62 | 0.20–2.33 | 54.87 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Disaccharides (mg/100 g) | 0.48 ± 0.47 | 0.00–1.38 | 97.92 | 0.74 ± 0.49 | 0.00–1.69 | 66.22 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Ascorbic acid (mg/100 g) | 63.98 ± 23.92 a | 28.16–113.52 | 37.39 | 101.36 ± 52.76 ab | 33.88–209.44 | 52.06 | 16.90 ± 2.24 abc | 12.32–20.00 | 13.23 | 56.89 ± 13.30 bc | 37.05–78.85 | 23.38 |
| Chlorophyll a (mg/100 g) | 85.71 ± 25.17 a | 55.17–147.16 | 29.37 | 58.66 ± 5.6 ab | 39.06–83.35 | 20.04 | 80.76 ± 22.65 b | 35.84–117.87 | 28.04 | 64.51 ± 11.33 a | 47.46–87.73 | 17.57 |
| Chlorophyll b (mg/100 g) | 25.04 ± 8.57 a | 10.73–45.91 | 34.22 | 16.29 ± 2.98 ab | 11.69–23.64 | 18.28 | 41.38 ± 12.80 abc | 20.02–61.99 | 30.93 | 27.72 ± 5.50 bc | 20.16–39.42 | 19.84 |
| Carotenoids (mg/100 g) | 30.32 ± 8.35 a | 16.59–50.36 | 27.52 | 21.02 ± 4.24 a | 12.88–28.77 | 20.17 | 23.60 ± 5.96 a | 14.04–31.42 | 25.27 | 18.91 ± 2.36 a | 16.42–23.56 | 12.46 |
| Anthocyanins (mg/100 g) | 1.60 ± 0.95 | 0.49–3.62 | 59.50 | 1.67 ± 1.43 | 0.58–6.34 | 85.33 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Biopoligon of AFI 12 h Photoperiod | Biopoligon of AFI 16 h Photoperiod | VIR, Greenhouse * | VIR, Field * | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Trait | Mean ± SD | Xmin–Xmax | CV, % | Mean ± SD | Xmin–Xmax | CV, % | Mean ± SD | Xmin–Xmax | CV, % | Mean ± SD | Xmin–Xmax | CV, % |
| Days to start of bolting | 45.44 ± 6.48 a | 31.00–55.00 | 14.27 | 33.78 ± 5.27 ab | 26.00–49.00 | 15.59 | 60.50 ± 5.69 abc | 47.00–73.00 | 9.40 | 30.62 ± 3.92 abc | 25.00–40.00 | 12.81 |
| Dry matter (%) | 9.47 ± 2.16 a | 6.26–15.77 | 22.84 | 10.25 ± 3.00 b | 7.10–22.51 | 29.24 | 7.66 ± 1.88 bc | 4.84–11.20 | 24.48 | 11.75 ± 2.93 ac | 6.76–19.04 | 24.96 |
| Total sugar content (mg/100 g) | 0.82 ± 0.56 a | 0.06–2.76 | 67.68 | 1.29 ± 0.57 b | 0.29–2.15 | 48.40 | 0.55 ± 0.47 bc | 0.18–2.68 | 86.62 | 1.32 ± 0.81 ac | 0.00–3.03 | 63.00 |
| Monosaccharides (mg/100 g) | 0.59 ± 0.30 a | 0.06–1.11 | 51.09 | 0.87 ± 0.46 a | 0.10–1.67 | 56.81 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Disaccharides (mg/100 g) | 0.23 ± 0.40 | 0.00–1.83 | 169.88 | 0.42 ± 0.47 | 0.00–1.79 | 112.69 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Ascorbic acid (mg/100 g) | 43.22 ± 19.92 a | 15.40–97.02 | 46.09 | 55.40 ± 27.13 b | 14.08–102.08 | 48.97 | 16.15 ± 3.24 abc | 8.00–20.00 | 20.08 | 55.37 ± 23.76 c | 20.90–138.70 | 42.91 |
| Chlorophyll a (mg/100 g) | 73.19 ± 20.95 a | 37.09–112.08 | 28.63 | 56.39 ± 14.42 ab | 31.01–105.65 | 25.57 | 83.47 ± 31.75 b | 41.54–150.44 | 38.03 | 73.42 ± 17.37 b | 50.99–140.21 | 23.57 |
| Chlorophyll b (mg/100 g) | 22.25 ± 6.38 a | 10.08–34.91 | 28.70 | 16.09 ± 4.42 b | 7.86–32.49 | 27.50 | 44.18 ± 18.54 abc | 23.41–86.77 | 41.97 | 32.63 ± 9.57 abc | 15.24–54.49 | 29.32 |
| Carotenoids (mg/100 g) | 27.37 ± 7.31 a | 13.22–39.97 | 26.72 | 19.94 ± 4.84 a | 13.14–35.76 | 24.27 | 22.33 ± 5.51 a | 13.28–31.90 | 24.70 | 17.99 ± 3.92 a | 9.73–26.61 | 21.78 |
| Anthocyanins (mg/100 g) | 1.58 ± 0.65 | 0.58–3.07 | 41.34 | 1.71 ± 1.00 | 0.49–4.32 | 58.23 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Trait | Group | Position | LOD | % Expl. | AFI | VIR * | ||
|---|---|---|---|---|---|---|---|---|
| 12 h | 16 h | Greenhouse | Field | |||||
| Ascorbic acid | R 01 | 16.20 | 2.35 | 45.1 | X | |||
| R 03 | 56.44 | 1.18 | 28.7 | X | ||||
| R 03 | 56.5 | 1.70 | 23.0 | X | ||||
| R 03 | 91.62 | 1.34 | 32.0 | X | ||||
| R 04 | 37.89 | 1.46 | 20.0 | X | ||||
| R 04 | 38.91 | 2.02 | 44.1 | X | ||||
| R 05 | 35.74 | 2.20 | 46.9 | X | ||||
| R 05 | 62.18 | 1.98 | 26.2 | X | ||||
| R 05 | 70.09 | 1.27 | 27.7 | X | ||||
| R 05 | 72.09 | 0.96 | 24.2 | X | ||||
| R 07 | 14.68 | 0.99 | 24.9 | X | ||||
| R 07 | 17.45 | 1.64 | 37.6 | X | ||||
| R 07 | 27.34 | 1.73 | 35.8 | X | ||||
| R 07 | 64.41 | 1.70 | 38.7 | X | ||||
| R 08 | 48.18 | 2.13 | 45.8 | X | ||||
| R 09 | 49.59 | 0.88 | 12.6 | X | ||||
| Anthocyanins | R 05 | 31.54 | 1.57 | 33.1 | X | |||
| R 05 | 75.77 | 1.83 | 34.4 | X | ||||
| Carotenoids | R 02 | 10.26 | 2.92 | 52.6 | X | |||
| R 03 | 31.63 | 2.68 | 33.7 | X | ||||
| R 03 | 73.54 | 1.17 | 28.7 | X | ||||
| R 03 | 73.55 | 0.39 | 5.9 | X | ||||
| R 03 | 102.83 | 1.07 | 23.9 | X | ||||
| R 03 | 111.89 | 1.40 | 27.6 | X | ||||
| R 04 | 3.00 | 2.23 | 40.1 | X | ||||
| R 04 | 16.74 | 1.11 | 27.4 | X | ||||
| R 04 | 22.27 | 0.82 | 11.8 | X | ||||
| R 05 | 20.70 | 0.96 | 24.2 | X | ||||
| R 05 | 20.70 | 0.83 | 11.9 | X | ||||
| R 06 | 4.37 | 1.86 | 24.9 | X | ||||
| R 06 | 31.31 | 1.03 | 21.1 | X | ||||
| R07 | 74.49 | 2.82 | 51.4 | X | ||||
| R 07 | 79.92 | 0.94 | 23.6 | X | ||||
| R 07 | 82.92 | 1.52 | 32.1 | X | ||||
| R 07 | 105.19 | 2.12 | 45.7 | X | ||||
| R 09 | 31.86 | 1.21 | 29.4 | X | ||||
| R 10 | 25.46 | 1.07 | 24.0 | X | ||||
| R 10 | 28.24 | 1.07 | 24.0 | X | ||||
| Chlorophyll a | R 02 | 8.26 | 2.44 | 46.5 | X | |||
| R 03 | 32.63 | 2.34 | 30.2 | X | ||||
| R 03 | 95.20 | 1.77 | 36.4 | X | ||||
| R 03 | 111.89 | 1.39 | 27.3 | X | ||||
| R 03 | 116.01 | 1.23 | 29.9 | X | ||||
| R 04 | 3.00 | 2.47 | 43.4 | X | ||||
| R 04 | 45.87 | 1.10 | 24.4 | X | ||||
| R 04 | 53.99 | 1.42 | 27.8 | X | ||||
| R 05 | 20.70 | 0.74 | 10.7 | X | ||||
| R 05 | 40.19 | 1.57 | 36.4 | X | ||||
| R 06 | 3.08 | 1.09 | 24.4 | X | ||||
| R 06 | 4.36 | 1.69 | 22.8 | X | ||||
| R 07 | 26.60 | 0.78 | 11.2 | X | ||||
| R 07 | 74.49 | 2.00 | 40.0 | X | ||||
| Chlorophyll b | R 01 | 35.22 | 3.17 | 55.6 | X | |||
| R 03 | 32.63 | 2.65 | 33.4 | X | ||||
| R 03 | 53.66 | 1.64 | 34.2 | X | ||||
| R 03 | 69.15 | 1.29 | 31.1 | X | ||||
| R 03 | 60.10 | 0.84 | 12.1 | X | ||||
| R 04 | 3.00 | 2.77 | 47.2 | X | ||||
| R 04 | 16.74 | 1.10 | 27.2 | X | ||||
| R 05 | 20.70 | 0.56 | 8.3 | X | ||||
| R 05 | 39.25 | 0.71 | 10.3 | X | ||||
| R 05 | 41.19 | 1.25 | 30.2 | X | ||||
| R 05 | 80.57 | 1.50 | 35.1 | X | ||||
| R 06 | 4.36 | 2.10 | 27.5 | X | ||||
| R 07 | 26.60 | 1.15 | 16.2 | X | ||||
| R 07 | 42.40 | 0.62 | 16.3 | X | ||||
| R 09 | 46.16 | 1.55 | 36.0 | X | ||||
| Dry matter | R 02 | 28.24 | 1.90 | 38.5 | X | |||
| R 06 | 39.32 | 1.98 | 39.7 | X | ||||
| Monosaccharides | R 03 | 29.12 | 1.42 | 29.1 | X | |||
| R 03 | 80.87 | 1.60 | 38.8 | X | ||||
| Disaccharides | R 01 | 40.02 | 3.01 | 51.8 | X | |||
| R 03 | 48.68 | 2.26 | 42.2 | X | ||||
| R 07 | 26.60 | 2.81 | 49.4 | X | ||||
| R 10 | 17.44 | 2.79 | 57.5 | X | ||||
| Trait | Group | Position | LOD | % Expl. | AFI | VIR * | ||
|---|---|---|---|---|---|---|---|---|
| 12 h | 16 h | Greenhouse | Field | |||||
| Ascorbic acid | R 02 | 42.75 | 0.62 | 8.6 | X | |||
| 55.56 | 1.81 | 32.7 | X | |||||
| R 05 | 24.52 | 3.10 | 35.9 | X | ||||
| 34.64 | 2.07 | 36.5 | X | |||||
| 43.07 | 2.28 | 28.0 | X | |||||
| 43.50 | 0.93 | 11.6 | X | |||||
| R 06 | 25.18 | 2.59 | 31.1 | X | ||||
| 86.52 | 1.53 | 28.5 | X | |||||
| 93.29 | 1.53 | 19.8 | X | |||||
| R 07 | 50.20 | 1.76 | 32.0 | X | ||||
| 57.74 | 0.55 | 7.7 | X | |||||
| R 08 | 12.49 | 1.51 | 18.1 | X | ||||
| R 09 | 0.00 | 2.69 | 32.1 | X | ||||
| 69.54 | 1.14 | 14.0 | X | |||||
| 74.03 | 1.66 | 21.2 | X | |||||
| Anthocyanins | R 05 | 56.99 | 1.54 | 19.9 | X | |||
| R 09 | 34.66 | 2.64 | 39.7 | X | ||||
| 45.59 | 1.21 | 16.0 | X | |||||
| Carotenoids | R 02 | 44.68 | 1.04 | 13.9 | X | |||
| R 02 | 44.68 | 0.74 | 10.1 | X | ||||
| R04 | 47.16 | 0.88 | 12.0 | X | ||||
| R05 | 10.66 | 0.59 | 8.1 | X | ||||
| R06 | 62.48 | 1.29 | 17.0 | X | ||||
| 96.59 | 1.91 | 30.6 | X | |||||
| R 08 | 62.52 | 1.41 | 23.6 | X | ||||
| R09 | 13.6 | 2.18 | 26.9 | X | ||||
| R 10 | 25.56 | 2.28 | 27.9 | X | ||||
| Chlorophyll a | R 04 | 47.16 | 0.80 | 10.9 | X | |||
| 52.69 | 0.72 | 9.0 | X | |||||
| R 05 | 38.28 | 1.05 | 12.9 | X | ||||
| 40.74 | 1.39 | 23.4 | X | |||||
| 43.07 | 1.05 | 14.0 | X | |||||
| R 06 | 52.78 | 0.84 | 10.5 | X | ||||
| 57.27 | 1.26 | 16.6 | X | |||||
| 62.48 | 1.44 | 18.7 | X | |||||
| 83.67 | 1.22 | 14.8 | X | |||||
| 96.59 | 1.94 | 31.1 | X | |||||
| R 08 | 62.51 | 0.80 | 10.0 | X | ||||
| 62.52 | 1.24 | 21.2 | X | |||||
| R 09 | 2.00 | 1.01 | 13.5 | X | ||||
| 6.92 | 1.34 | 16.2 | X | |||||
| 13.60 | 1.94 | 24.3 | X | |||||
| 15.58 | 1.83 | 21.4 | X | |||||
| 28.74 | 2.01 | 23.2 | X | |||||
| 35.65 | 1.31 | 17.2 | X | |||||
| 67.28 | 1.00 | 17.4 | X | |||||
| R 10 | 17.43 | 0.98 | 12.1 | X | ||||
| 25.56 | 2.45 | 29.7 | X | |||||
| Chlorophyll b | R 05 | 10.665 | 0.65 | 8.9 | X | |||
| 43.07 | 1.10 | 14.6 | X | |||||
| 46.39 | 0.82 | 11.1 | X | |||||
| 50.83 | 4.46 | 44.4 | X | |||||
| R 06 | 76.37 | 1.37 | 17.9 | X | ||||
| 83.67 | 1.2 | 14.6 | X | |||||
| 96.59 | 1.56 | 25.9 | X | |||||
| R 09 | 15.58 | 1.53 | 18.2 | X | ||||
| 13.60 | 2.77 | 32.9 | X | |||||
| R 10 | 25.56 | 2.09 | 25.9 | X | ||||
| 27.95 | 2.13 | 26.4 | X | |||||
| 59.23 | 1.24 | 15.0 | X | |||||
| Dry matter | R 05 | 23.50 | 2.03 | 25.4 | X | |||
| 43.07 | 1.47 | 19.1 | X | |||||
| 55.58 | 1.72 | 28.1 | X | |||||
| Monosaccharide | R 05 | 22.81 | 1.27 | 18.3 | X | |||
| R 08 | 17.37 | 2.39 | 45.7 | X | ||||
| Disaccharides | R 02 | 89.36 | 1.23 | 17.7 | X | |||
| R 07 | 35.17 | 1.76 | 36.3 | X | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Egorova, K.V.; Sinyavina, N.G.; Artemyeva, A.M.; Kocherina, N.V.; Chesnokov, Y.V. QTL Analysis of the Content of Some Bioactive Compounds in Brassica rapa L. Grown under Light Culture Conditions. Horticulturae 2021, 7, 583. https://doi.org/10.3390/horticulturae7120583
Egorova KV, Sinyavina NG, Artemyeva AM, Kocherina NV, Chesnokov YV. QTL Analysis of the Content of Some Bioactive Compounds in Brassica rapa L. Grown under Light Culture Conditions. Horticulturae. 2021; 7(12):583. https://doi.org/10.3390/horticulturae7120583
Chicago/Turabian StyleEgorova, Ksenia V., Nadezhda G. Sinyavina, Anna M. Artemyeva, Natalia V. Kocherina, and Yuriy V. Chesnokov. 2021. "QTL Analysis of the Content of Some Bioactive Compounds in Brassica rapa L. Grown under Light Culture Conditions" Horticulturae 7, no. 12: 583. https://doi.org/10.3390/horticulturae7120583