
Macroelements and Microelements in the Soil and Their Relationship with the Content of Steviol Glucosides in Stevia rebaudiana Bert from Five Regions of Colombia

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
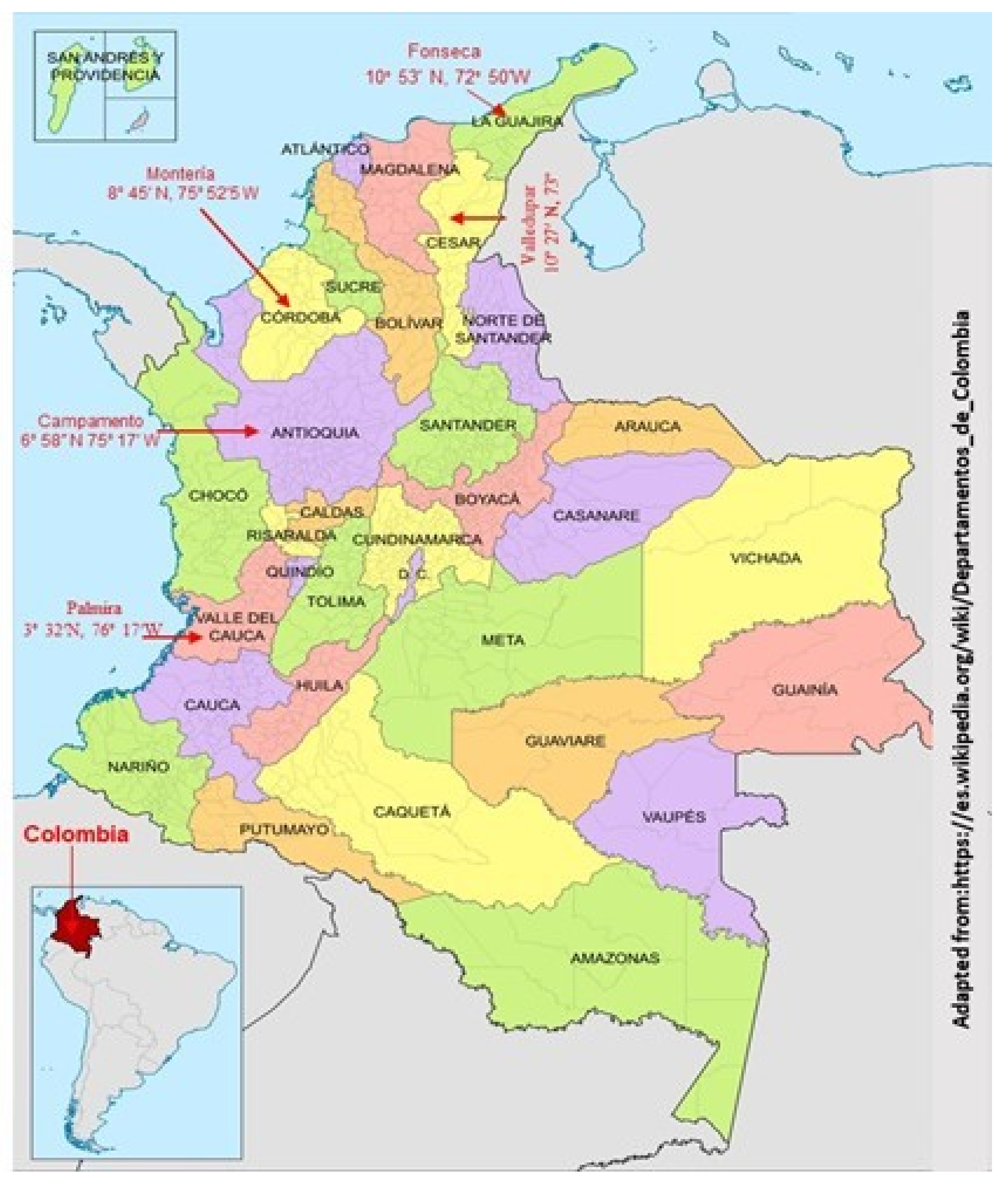
2.1. Location
2.2. Experimental Design
2.3. Collection of Soil Samples
2.4. Collection of Plant Material
2.5. Extraction of Glycosides from Steviol, Stevioside and Rebaudioside and Analysis by HPLC
2.6. Statistical Analysis
3. Results
3.1. Chemical Characterization of Macroelements in Soils and Nutritional Content in Stevia Leaves in Five Municipalities of Colombia and Statistical Analysis between Localities
3.2. Characterization of Micro Elements in Soils and Stevia Leaves
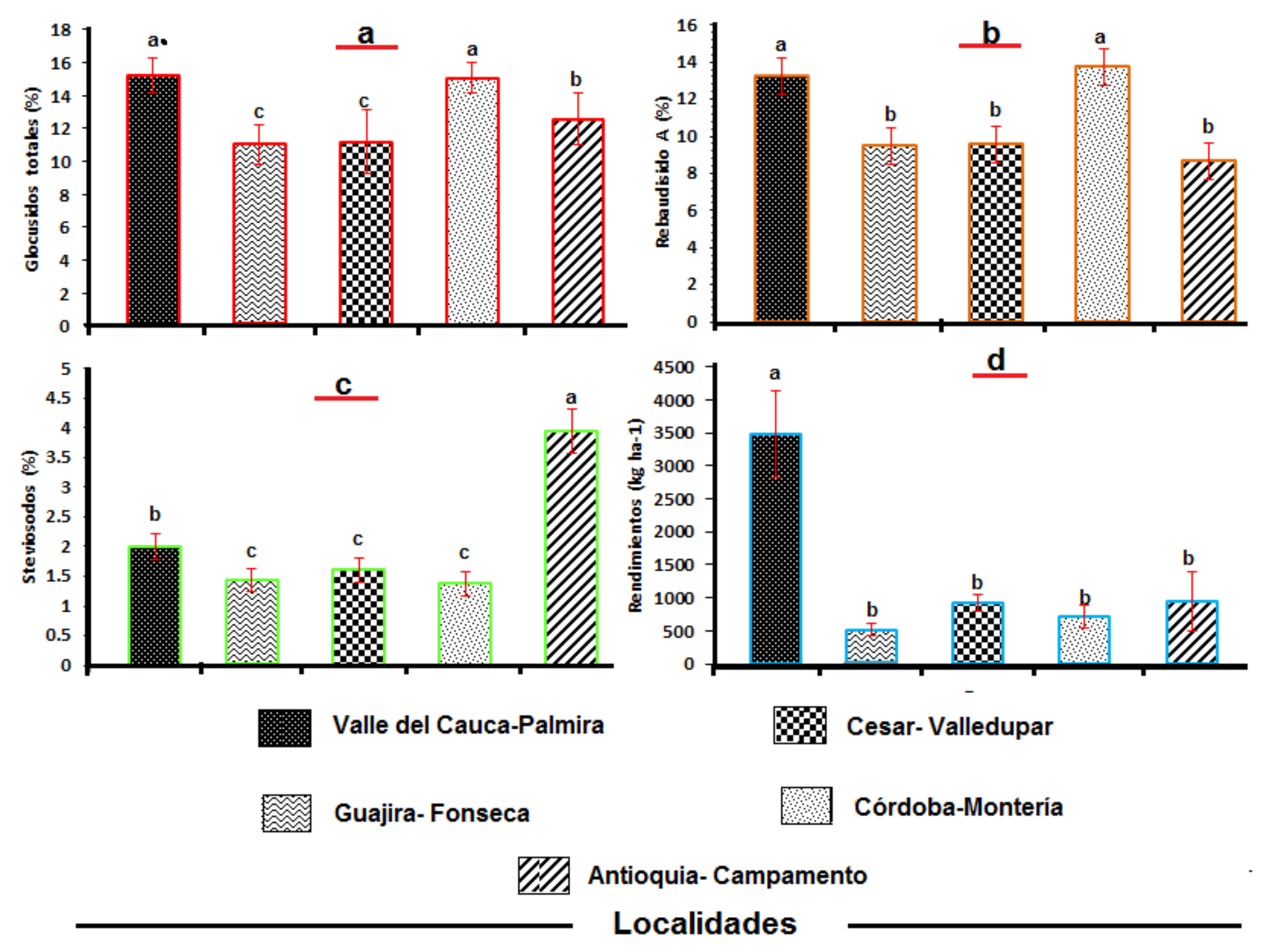
3.3. Total Glycoside Content and Stevia Yields at Five Locations
3.4. Effect of Macro and Microelements on the Content of Total Glycosides and Leaf Yields in Stevia
4. Discussion
4.1. Nutritional Characterization of Macro Elements in Soils and Stevia Leaves
4.2. Nutritional Characterization of Micro Elements in Soils and Stevia Leaves
4.3. Correlation between Macro and Micro FOLIAR Elements with Total Glycosides, Rebaudioside A, Stevioside, and Stevia Leaf Yield in Five Environments of Colombia
4.4. Correlation between Macro and Micro Nutritional Elements and Total Glycosides
4.5. Correlation between Macro and Microelements and Rebaudioside A, Steviosides
4.6. Correlation between Macro and Micro Elements and Performance
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reichardt, K.; Timm, L.C. How Plants Absorb Nutrients from the Soil. In Soil, Plant and Atmosphere; Springer: Cham, Switzerland, 2020; pp. 313–330. [Google Scholar]
- Pritchard, S.G.; Amthor, J.S. Crops and Environmental Change; Food Products Press: New York, NY, USA, 2005; p. 421. [Google Scholar]
- Chatsudthipong, V.; Muanprasat, C. Stevioside and related compounds: Therapeutic benefits beyond sweetness. Pharmacol. Ther. 2009, 121, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Geuns, J.M.C. Molecules of interest stevioside. Phytochemistry 2003, 64, 913–921. [Google Scholar] [CrossRef]
- Espitia, M.; Montoya, R.; Atencio, L. Rendimiento de Stevia rebaudiana Bert. bajo tres arreglos poblacionales en el Sinú Medio. Rev. UDCA Actual. Divulg. Cient. 2009, 12, 151–161. [Google Scholar]
- Jain, P.; Kachhwaha, S.; Kothari, S.L. Improved micropropagation protocol and enhancement in biomass and chlorophyll content in Stevia rebaudiana (Bert.) Bertoni by using high copper levels in the culture medium. Sci. Hortic. 2008, 119, 315–319. [Google Scholar] [CrossRef]
- Jarma, O.A.J.; Combatt, C.; Cleves, C. Aspectos nutricionales y metabolismo de Stevia rebaudiana (Bertoni): Una revisión. Agron. Colomb. 2010, 28, 199–208. [Google Scholar]
- Lei, M.; Yan, S. Effects of potassium fertilizer on physiological and biochemical index of Stevia rebaudiana Bertoni. Energy Procedia 2011, 5, 581–586. [Google Scholar]
- Brandle, J.E.; Telmer, P.G. Steviol glycoside biosynthesis. Phytochemistry 2007, 68, 1855–1863. [Google Scholar] [CrossRef] [PubMed]
- Totté, N.; Charon, L.; Rohmer, M.; Compernolle, F.; Baboeuf, I.; Geuns, J. Biosynthesis of the Diterpenoid Steviol, an Entkaurene Derivative from Stevia rebaudiana Bertoni, Via the Methylerythritol Phosphate Pathway. Tetrahedron Lett. 2000, 41, 6407–6410. [Google Scholar] [CrossRef]
- Joyard, J.; Ferro, M.; Masselon, C.; Siegneuri-Berny, D.; Salvi, D.; Garin, J.; Rolland, N. Chloroplast proteomics and the compartmentation of plastidial isoprenoid biosintetic pathways. Mol. Plant 2009, 1154–1436. [Google Scholar] [CrossRef]
- Michelet, L.; Krieger, L.A. Reactive oxygen intermediates produced by photosynthetic electron transport are enhanced in short-day grown plants. Biochim. Biophys. Acta-Biomembr. 2011, 1817, 1306–1313. [Google Scholar] [CrossRef] [Green Version]
- Ceunen, S.; Geuns, J.M.C. Influence of photoperiodism on the spatio-temporal accumulation of steviol glycosides in Stevia rebaudiana (Bertoni). Plant Sci. 2013, 198, 72–82. [Google Scholar] [CrossRef]
- Barbet, M.C.; Giuliano, S.; Alletto, L.; Daydé, J.; Berger, M. Towards a semi-perennial culture of Stevia rebaudiana (Bertoni) Bertoni under temperate climate: Effects of genotype, environment and plant age on steviol glycoside content and composition. Genet. Res. Crop Evol. 2016, 63, 685–694. [Google Scholar] [CrossRef]
- Woelwer, R.U. Improved HPLC method for the evaluation of the major steviol glycosides in leaves of Stevia rebaudiana. Eur. Food Res. Technol. 2010, 231, 581–588. [Google Scholar] [CrossRef]
- Serfaty, M.; Ibdah, M.; Fischer, R.; Chaimovitsh, D.; Saranga, Y.; Dudai, N. Dynamics of yield components and stevioside production in Stevia rebaudiana grown under different planting times, plant stands and harvest regime. Prod. Ind. Crop. Prod. 2013, 50, 731–736. [Google Scholar] [CrossRef]
- Kumar, P.P.; Mahajan, M.; Prasad, R.V.; Pathania, V.; Singh, B.; Singh, P.A. Harvesting regimes to optimize yield and quality in annual and perennial Stevia rebaudiana under sub-temperate conditions. Pharmacogn. Res. 2015, 2, 258–263. [Google Scholar]
- IGAC. Métodos analíticos del laboratorio de suelos, 6th ed.; Subdirección de Agrología: Bogotá, Colombia, 2006; p. 460. [Google Scholar]
- Empresa Brasileira de Pesquisa Agropecuária-EMBRAPA. Centro Nacional de Pesquisa de Solos. Manual de métodos de análise de solos, 2nd ed.; EMBRAPA-CNPS: Rio de Janeiro, Brazil, 1997; p. 212. [Google Scholar]
- Montoro, P.; Molfetta, I.; Maldini, M.; Ceccarini, L.; Piacente, S.; Pizza, C.; Macchia, M. Determination of six steviol glycosides of Stevia rebaudiana (Bertoni) from different geographical origin by LC–ESI–MS/MS. Food Chem. 2013, 14, 745–753. [Google Scholar] [CrossRef]
- Kuncoro, P.H.; Kog, K.; Satta, N.; Muto, Y. A study on the effect of compaction on transport properties of soil gas and water. I: Relative gas diffusivity, air permeability, and saturated hydraulic conductivity. Soil Tillage Res. 2014, 143, 172. [Google Scholar] [CrossRef]
- Fyfe, W.S.; Leonardos, O.H.; Theodoro, S.H. The use of rocks to improve family agriculture in Brazil. Anais da Academia Brasileira de Ciências 2006, 78, 721–730. [Google Scholar]
- Kobus, M.; Moryson, A.; Gramza, M. Directions on the use of stevia leaves (Stevia rebaudiana) as an additive in food products. Acta Sci. Pol. Technol. Aliment. 2015, 14, 5–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, R.G.; Gonzalez, R.H.; Ramýrez, O.R.; Cerrill, S.M.A.; Juarez, A.S. Seasonal trends of macro and micro minerals in 10 browse species that grow in northeastern Mexico. Anim. Feed Sci. Technol. 2006, 128, 155–164. [Google Scholar] [CrossRef]
- Tadhani, M.R.S. Preliminary Studies on Stevia rebaudiana Leaves: Proximal Composition, Mineral Analysis and Phytochemical Screening. J. Med. Sci. 2006, 6, 321–326. [Google Scholar]
- Romero, F.J.C.; Rodríguez, M.M.N.; Gutiérrez, C.M.C.; Escalante, J.A.S.; Peña, C.B.; Cueto, J.A. Growth and secondary metabolites of Stevia pilosa Lag. in three edaphoclimatic conditions in the state of Hidalgo, Mexico. Revista Chapingo Serie Ciencias Forestales y del Ambiente 2020, 26, 173–187. [Google Scholar] [CrossRef]
- Khiraoui, A.; Bakha, M.; Amchra, F.; Ourouadi, S.; Boulli, A.; Faiz, C.A.; Hasib, A. Nutritional and biochemical properties of natural sweeteners of six cultivars of Stevia rebaudiana Bertoni leaves grown in Morocco. J. Rev. Mater. Environ. Sci. 2017, 8, 1015–1022. [Google Scholar]
- Hatfield, J.L.; Prueger, J.H. Temperature extremes: Effect on plant growth and development. Rev. Weather. Clim. Extrem. 2015, 10, 4–10. [Google Scholar] [CrossRef] [Green Version]
- Samra, J.; Arora, L. Mineral nutrition. The mango: Botany, production and uses. Rev. CAB Int. 1997, 76, 175–201. [Google Scholar]
- Razaq, M.; Zhang, P.; Shen, H.L.S. Influence of nitrogen and phosphorus on the growth and root morphology of Acer mono. PLoS ONE 2017, 12, e0171321. [Google Scholar] [CrossRef] [Green Version]
- Zheng, R.; Su, S.; Xiao, H.; Tian, H.Q. Calcium: A Critical Factor in Pollen Germination and Tube Elongation. Int. J. Mol. Sci. 2019, 2, 420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, S.R.; Welch, R.M.; Mayland, H.F.; Grunes, D.L. Magnesium in plants: Uptake, distribution, function, and utilization by man and animals. In Metal Ions in Biological Systems; Sigel, H., Sigel, A., Eds.; Marcel Dekker Inc.: New York, NY, USA, 1990; pp. 33–56. [Google Scholar]
- Alloway, B.J. Zinc in Soils and Crop Nutrition; Revista Crop International Zinc Association: Brussels, Belgium, 2008; p. 139. [Google Scholar]
- Arbelo, C.D.; Armas, C.M.; Guerra, J.A.; Rodríguez, A. Salinidad y alcalinidad en suelos de las zonas de Tenerife (Islas canarias). Rev. Crop. Int. Edafol. 2006, 13, 171–179. [Google Scholar]
- Crichton, R.R.; Wilmet, S.; Legssyer, R.; Ward, R.J. Molecular and cellular mechanisms of iron homeostasis and toxicity in mammalian cells. J. Inorg. Biochem. 2002, 91, 9–18. [Google Scholar] [CrossRef]
- Das, K.; Shivananda, T.N.; Dang, R.; Sur, P. Interaction between phosphorus and zinc on the biomass yield and yield attributes of the medicinal plant stevia (Stevia rebaudiana). Sci. World J. 2005, 5, 390–395. [Google Scholar] [CrossRef] [Green Version]
- Nader, R.A.; Botros, W.A.; Khaled, A.E.; Mohamed, A.G.; Shimaa, G.H.; Hayssam, M.A.; Mohamed, S.E. Comparison of uridine diphosphateglycosyltransferase UGT76G1 genes from some varieties of Stevia rebaudiana Bertoni. Sci. Rep. 2019, 9, 8559. [Google Scholar]
- Jarma, O.A.; Combatt, C.E.; Polo, S.J. Glycoside contents depending on the nutrient deficiencies in Stevia rebaudiana Bert. Revista UDCA de Actualidad y Divulgación Cientifica 2012, 15, 107–116. [Google Scholar]
- Ahmad, N.; Rab, A.; Ahmad, N. Light-induced biochemical variations in secondary metabolite production and antioxidant activity in callus cultures of Stevia rebaudiana (Bert). J. Photochem. Photobiol. B 2016, 154, 51–56. [Google Scholar] [CrossRef]
- De Lima, O.; Malavolta, E. Sintomas de desordens nutricionais em estévia Stevia rebaudiana (Bert.). J. Sci. Agric. 1997, 54, 53–61. [Google Scholar] [CrossRef]
- Sharma, S.; Walia, S.; Singh, B.; Kumara, R. Comprehensive review on agro technologies of low-calorie natural sweetener stevia (Stevia rebaudiana Bertoni): A boon to diabetic patients. J. Sci. Food Agric. 2016, 96, 1867–1879. [Google Scholar] [CrossRef]
- Tavarini, S.; Sgherri, C.; Ranieri, A.M.; Angelini, L.G. Effect of nitrogen fertilization and harvest time on steviol glycosides flavonoid composition, and antioxidant properties in Stevia rebaudiana Bertoni. J. Agric. Food Chem. 2015, 63, 7041–7050. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, T. From waste products to ecochemicals: Fifty years research of plant secondary metabolism. Phytochemistry 2007, 68, 2831–2846. [Google Scholar] [CrossRef] [PubMed]
- Pal, P.K.; Kumar, R.; Guleria, V.; Mahajan, M.; Prasad, R.; Pathania, V.; Gill, B.S.; Singh, D.; Chand, G.; Singh, B.; et al. Crop-ecology and nutritional variability influence growth and secondary metabolites of Stevia rebaudiana Bertoni. BMC Plant Biol. 2015, 15, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senbayram, M.; Gransee, A.; Wahle, V.; Thiel, H. Role of magnesium fertilisers in agriculture: Plant–soil continuum. Crop Pasture Sci. 2016, 66, 1219–1229. [Google Scholar] [CrossRef]
- Gerendás, J.; Führs, H. The significance of magnesium for crop quality. Plant Soil 2013, 368, 101–128. [Google Scholar] [CrossRef] [Green Version]
- Festa, R.A.; Thiele, D.J. Copper: An essential metal in biology. Curr. Biol. 2011, 21, 877–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Yang, Y.; Hou, M.; Huang, X.; Zhang, T.; Huang, S.; Yuan, H. Optimized Nitrogen Topdressing Strategies Enhance Steviol Glycoside Productivity in Stevia (Stevia rebaudiana Bertoni) Plants. J. Soil Sci. Plant Nutr. 2020, 20, 1133–1143. [Google Scholar] [CrossRef]
- Utumi, M.M.; Monnerat, P.H.; Pereira, P.R.G.; Fontes, P.C.R.; Godinho, V.P.C. Macronutrient deficiencies in Stevia: Visual symptoms and effects on growth, chemical composition, and stevioside production. Pesquisa Agropecuaria Brasileira 1999, 34, 1039–1043. [Google Scholar]
- Geeta, G.K.; Midmore, D.J.; Resham, G. Effect of nutrient omission and pH on the biomass and concentration and content of steviol glycosides in stevia (Stevia rebaudiana (Bertoni) Bertoni) under hydroponic conditions. J. Appl. Res. Med. Aromat. Plants 2017, 7, 136–142. [Google Scholar]
- Humphries, J.M.; Stangoulis, J.C.R.; Graham, R.D. Manganese. In Handbook of Plant Nutrition; Barker, A.V., Pilbeam, D.J., Eds.; CRC PRESS: Boca Raton, FL, USA, 2007; Volume 351, p. 374. [Google Scholar]
- Bondarev, N.I.; Sukhanova, M.A.; Reshetnyak, O.V.; Nosov, A.M. Stevial Glycoside content in different organs of Stevia rebaudiana and its dynamics during ontogeny. Biol. Plant. 2003, 47, 261–264. [Google Scholar] [CrossRef]
- Cheeseman, J.M. The integration of activity in saline environments: Problems and perspectives. Funct. Plant Biol. 2013, 40, 759–774. [Google Scholar] [CrossRef]
- Landi, M.; Pardossi, A.; Remorini, D.; Guidi, L. Antioxidant and photosynthetic response of a purple-leaved and a green-leaved cultivar of sweet basil (Ocimum basilicum) to boron excess. Environ. Exp. Bot. 2013, 85, 64–75. [Google Scholar] [CrossRef]
- Marschner, P. Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: New York, NY, USA, 2012; p. 672. [Google Scholar]
- Siqueira-Silva, A.I.; da Silva, L.C.; Azevedo, A.A.; Oliva, M.A. Iron plaque formation and morphoanatomy of roots from species of resting subjected to excess iron. Ecotoxicol. Environ. Saf. 2012, 78, 265–275. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Location | IR 100% | IR 50% | T | P | SB | PH |
|---|---|---|---|---|---|---|
| Palmira (Valle) | 790 | 395 | 23.7 | 1065 | 1946 | 7.8 |
| Campamento (Antioquia) | 659 | 330 | 19.5 | 3973 | 1391 | 4.6 |
| Montería (Córdoba) | 834 | 417 | 28 | 1247 | 2180 | 6.4 |
| Valledupar (Cesar) | 1010 | 505 | 29 | 1324 | 2676 | 7.4 |
| Fonseca (Guajira) | 1097 | 548 | 28 | 839 | 2591 | 7.5 |
| Macronutrient Contents in the Soil | |||||||
|---|---|---|---|---|---|---|---|
| Locality | O M. | P | S | Na | K | Ca | Mg |
| mg kg | |||||||
| Palmira | 2.0 ± 0.5b | 86.1 ± 20.1b | 18.4 ± 4.5bc | 0.39 ± 0.20b | 0.54 ± 0.05c | 15.6 ± 1.0b | 6.5 ± 0.5b |
| Valledupar | 2.1 ± 0.7b | 94.6 ± 20.6a | 21.5 ± 9.6bc | 0.45 ± 0.20b | 0.91 ± 0.13b | 5.6 ± 0.6d | 2.2 ± 0.2d |
| Fonseca | 4.1 ± 1.1a | 68.2 ± 10.6b | 31.7 ± 9.2ab | 0.92 ± 0.20a | 1.26 ± 0.24a | 20.1 ± 2.2a | 3.9 ± 0.8c |
| Montería | 1.1 ± 0.3c | 12.9 ± 6.5c | 12.9 ± 5.1c | 0.12 ± 0.03c | 0.42 ± 0.24c | 13.0 ± 1.9c | 8.4 ± 1.3a |
| Campamento | 4.0 ± 0.3a | 96 ± 22a | 46.3 ± 20a | 0.06 ± 0.03c | 0.44 ± 0.09c | 3.8 ± 1.8d | 2.1 ± 0.3d |
| Macronutrient Contents in Stevia Leaves | |||||||
| Locality | N | P | S | Na | K | Ca | Mg |
| g kg | |||||||
| Palmira | 1.96 ± 0.1e | 2.26 ± 0.2bc | 0.83 ± 0.2a | 0.16 ± 0.1a | 18.7 ± 3.53b | 8.13 ± 1.1b | 4.25 ± 0.5a |
| Valledupar | 2.50 ± 0.1c | 3.66 ± 0.6a | 1.21 ± 0.2a | 0.10 ± 0.05a | 35.7 ± 6.92a | 7.64 ± 1.5bc | 3.40 ± 0.5b |
| Fonseca | 3.43 ± 0.4a | 3.55 ± 0.3a | 1.97 ± 2.0a | 0.15 ± 0.10a | 35.9 ± 7.31a | 9.80 ± 1.3a | 3.25 ± 0.7ab |
| Montería | 2.26 ± 0.1d | 2.88 ± 0.3b | 1.40+0.3a | 0.06 ± 0.01a | 22.88+4.82b | 6.00 ± 0.0c | 3.00 ± 0.0b |
| Campamento | 2.84 ± 0.2b | 1.76 ± 0.5c | 1.06 ± 0.5a | 0.09 ± 0.03a | 17.84 ± 9.57b | 3.47 ± 0.9d | 2.05 ± 0.9c |
| CorrCoef. | 0.74 ** | −0.12 ns | −0.12 ns | 0.54 ** | 0.11 ns | 0.41 * | 0.23 ns |
| Loc | 2.59 ** | 5.69 ** | 1.50 ns | 0.014 ns | 695.69 ** | 46.65 ** | 5.07 ** |
| RL | 0.19 ** | 0.25 ns | 0.49 ns | 0.002 ns | 159.29 ns | 16.05 ** | 3.00 ** |
| Loc * RL | 0.14 ** | 0.62 ** | 1.31 ns | 0.015 * | 28.77 ns | 2.82 * | 0.72 * |
| Micronutrient Contents in the Soil | |||||
|---|---|---|---|---|---|
| Cu | Fe | Zn | Mn | B | |
| mg kg | |||||
| Palmira | 0.38 ± 0.3c | 0.80 ± 0.1c | 1.53 ± 1.0b | 19.82 ± 8.4c | 0.25 ± 0.05bc |
| Valledupar | 0.42 ± 0.3bc | 10.78 ± 3.3c | 2.58 ± 0.12b | 39.20 ± 10.2b | 0.29 ± 0.03bc |
| Fonseca | 0.38 ± 0.4c | 1.87 ± 0.4c | 2.57 ± 0.8b | 26.35 ± 6.4c | 0.37 ± 0.11a |
| Montería | 2.62 ± 0.5a | 26.08 ± 6.2b | 2.83 ± 0.4b | 54.67 ± 2.3a | 0.30 ± 0.16ab |
| Campamento | 1.33 ± 0.5b | 50.43 ± 23.2a | 9.97 ± 2.2a | 21.23 ± 5.4c | 0.18 ± 0.04c |
| Micronutrient Contents in Stevia Leaves | |||||
| Cu | Fe | Zn | Mn | B | |
| mg kg | |||||
| Palmira | 11.25 ± 3.54a | 393.25 ± 5.2b | 87.5 ± 23.1a | 268.5 ± 35.2a | 16.88 ± 2.7c |
| Valledupar | 10.00 ± 0.00a | 1527.80 ± 868a | 49.9 ± 0.32a | 106.1 ± 16.9bc | 20.20 ± 3.1b |
| Fonseca | 11.25 ± 3.54a | 304.13 ± 133.8b | 87.0 ± 34.9a | 143.7 ± 39.9b | 23.75 ± 2.7a |
| Montería | 9.94 ± 0.05a | 444.07 ± 235.4b | 55.7 ± 6.97a | 58.7 ± 5.76c | 9.23 ± 0.9d |
| Campamento | 9.93 ± 0.05a | 343.34 ± 144.4b | 69.4 ± 20.5a | 273.8 ± 85.8a | 5.73 ± 2.0e |
| Coef.Corr | −0.20 ns | −0.12 ns | −0.02 ns | −0.77 ** | 0.43 * |
| Loc | 4.26 ns | 2,589,780 ** | 6332.3 ns | 82,844.3 ** | 482.89 ** |
| RL | 0.0009 ns | 587,525.5 ns | 4608.4 ns | 5629.66 ns | 11.30 ns |
| Loc * RL | 6.251 ns | 261758.6 ns | 9935.5 ns | 4226.31 ns | 9.544 ns |
| Fv | GL | Sum of Squares | Middle Square |
|---|---|---|---|
| Loc | 4 | 15,470.69 | 3867.67 ** |
| Block | 4 | 875.83 | 218.95 ns |
| RL6 | 1 | 19.48 | 19.48 ns |
| Loc * RL | 46 | 353.70 | 88.42 ns |
| Error | 30 | 6328.13 | 210.93 ns |
| Total | 43 | 23,047.85 | 535.99 |
| Average | 129.99 | ||
| coefficient of variation (%) | 11.17 |
| Nutrients Foliares | G T | RebA | Stv | Yield |
|---|---|---|---|---|
| N | 19.86 ** | — | 9.281 ** | 596 ** |
| P | - | 13.74 ** | - | - |
| S | - | - | - | - |
| Ca | - | - | −3.826 | 349 ** |
| Mg | 25.97 ** | 21.06 ** | 4.07 ** | −831 ** |
| K | - | - | - | - |
| Zn | - | - | - | - |
| Mn | - | 0.06714 * | −1.607 ° | |
| B | −2.949 | - 2629 | - | 42.1 |
| Fe | - | - | - | −0.442 ** |
| Na | - | - | −11.68 ** | - |
| Cu | 3.32 * | 3.66 ** | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Combatt Caballero, E.; Hernández Burgos, J.; Jarma-Orozco, A.; Jaraba Navas, J.; Rodríguez Páez, L. Macroelements and Microelements in the Soil and Their Relationship with the Content of Steviol Glucosides in Stevia rebaudiana Bert from Five Regions of Colombia. Horticulturae 2021, 7, 547. https://doi.org/10.3390/horticulturae7120547
Combatt Caballero E, Hernández Burgos J, Jarma-Orozco A, Jaraba Navas J, Rodríguez Páez L. Macroelements and Microelements in the Soil and Their Relationship with the Content of Steviol Glucosides in Stevia rebaudiana Bert from Five Regions of Colombia. Horticulturae. 2021; 7(12):547. https://doi.org/10.3390/horticulturae7120547
Chicago/Turabian StyleCombatt Caballero, Enrique, Jaime Hernández Burgos, Alfredo Jarma-Orozco, Juan Jaraba Navas, and Luis Rodríguez Páez. 2021. "Macroelements and Microelements in the Soil and Their Relationship with the Content of Steviol Glucosides in Stevia rebaudiana Bert from Five Regions of Colombia" Horticulturae 7, no. 12: 547. https://doi.org/10.3390/horticulturae7120547