
The Effect of Microbial Fertilizer on the Growth, Rhizospheric Environment and Medicinal Quality of Fritillaria taipaiensis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Inoculum
2.2. Plant Materials
2.3. Reference Drugs
2.4. Cultivation Management and Sample Collection
2.5. Analysis of Mycorrhizal Colonization
2.6. Determination of Physiological and Biochemical Indices of Leaves
2.7. Count of Rhizosphere Microorganism
2.8. Determination of Enzyme Activity in Rhizospheric Soil
2.9. Quality Analysis of Bulb
2.10. Data Analysis
3. Results
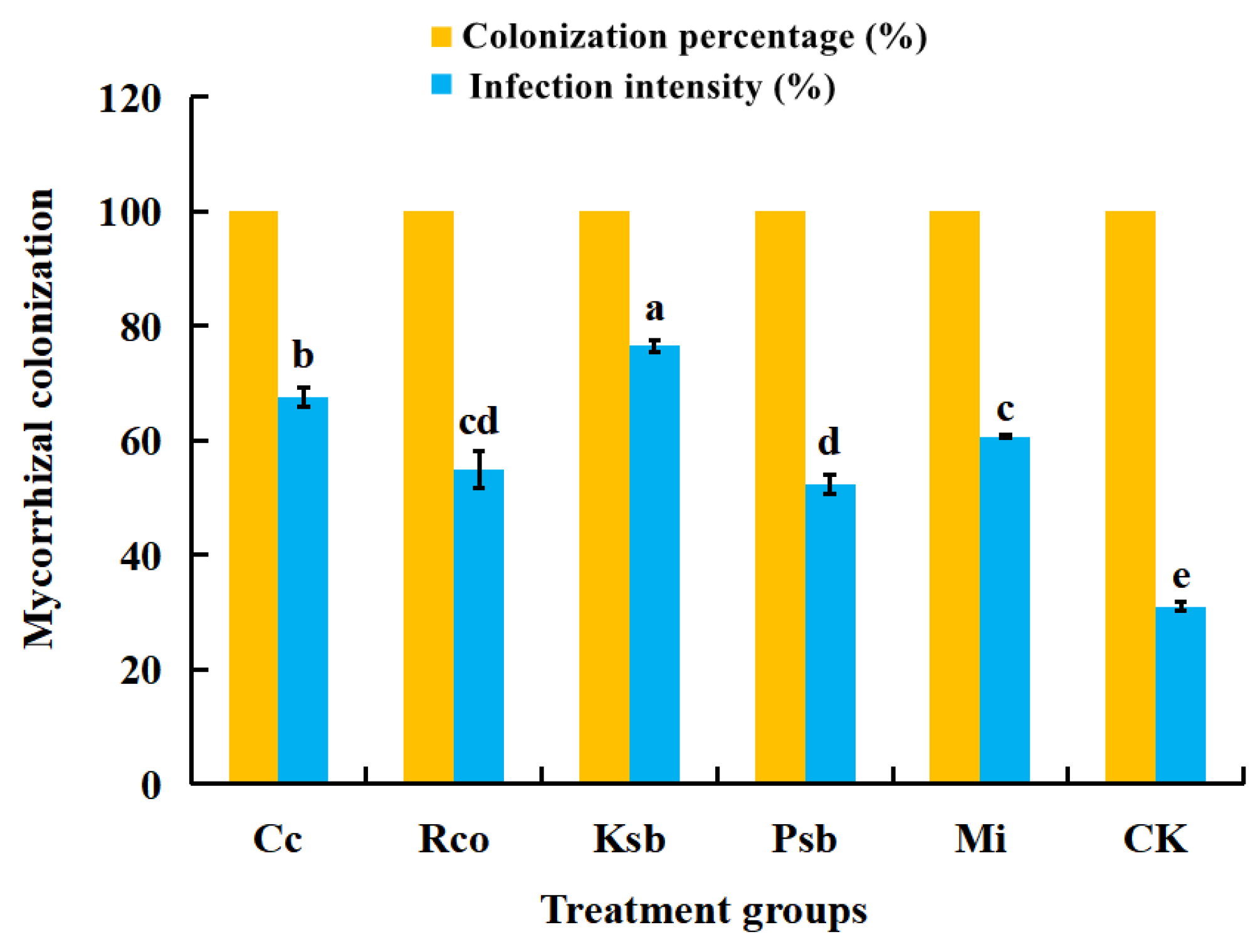
3.1. Effects of Different Microbial Inoculum Treatments on Mycorrhizal Colonization in the Root System of F. taipaiensis
3.2. Effects of Different Microbial Inoculum Treatments on Photosynthetic Pigment Content of F. taipaiensis Leaves
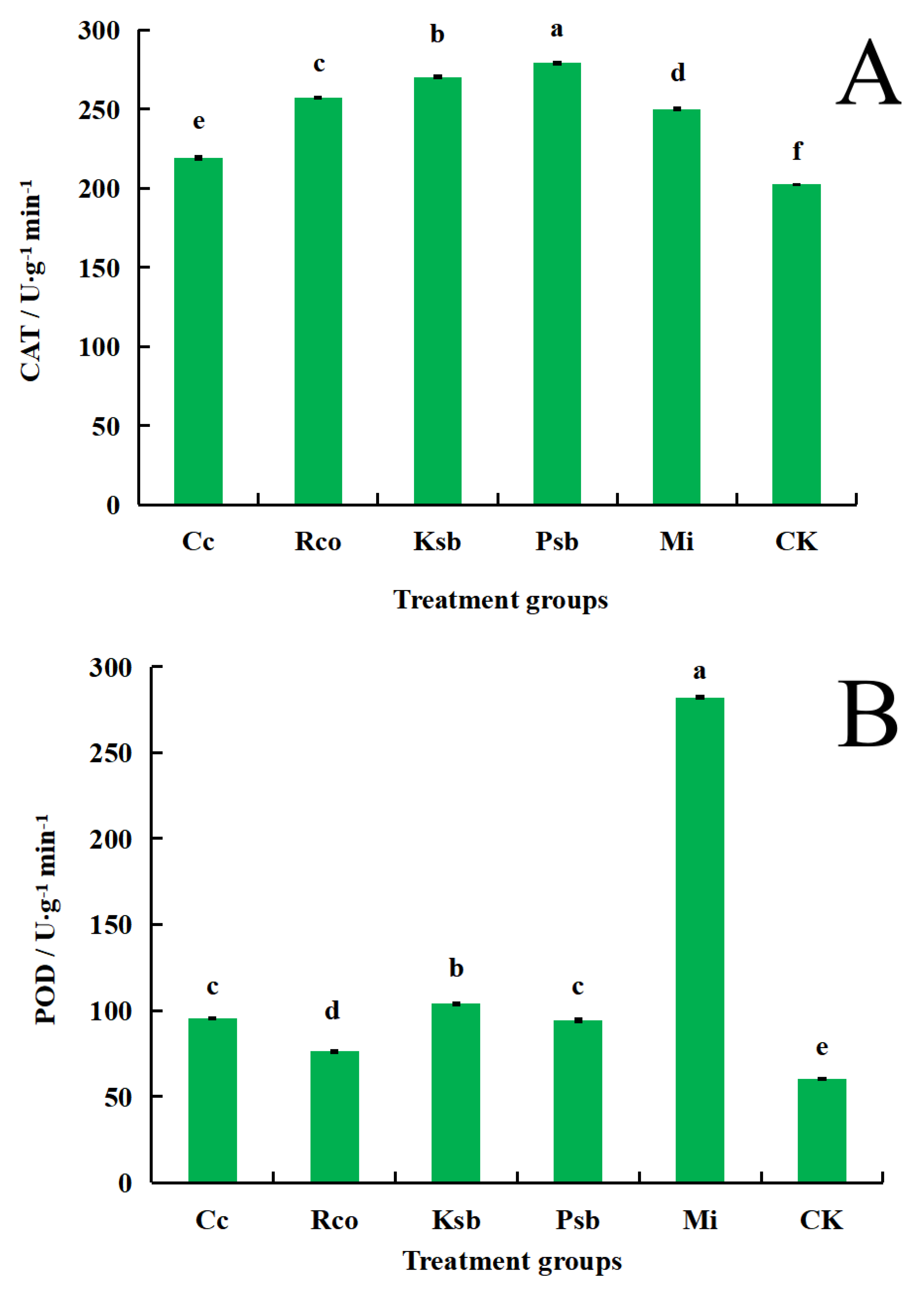
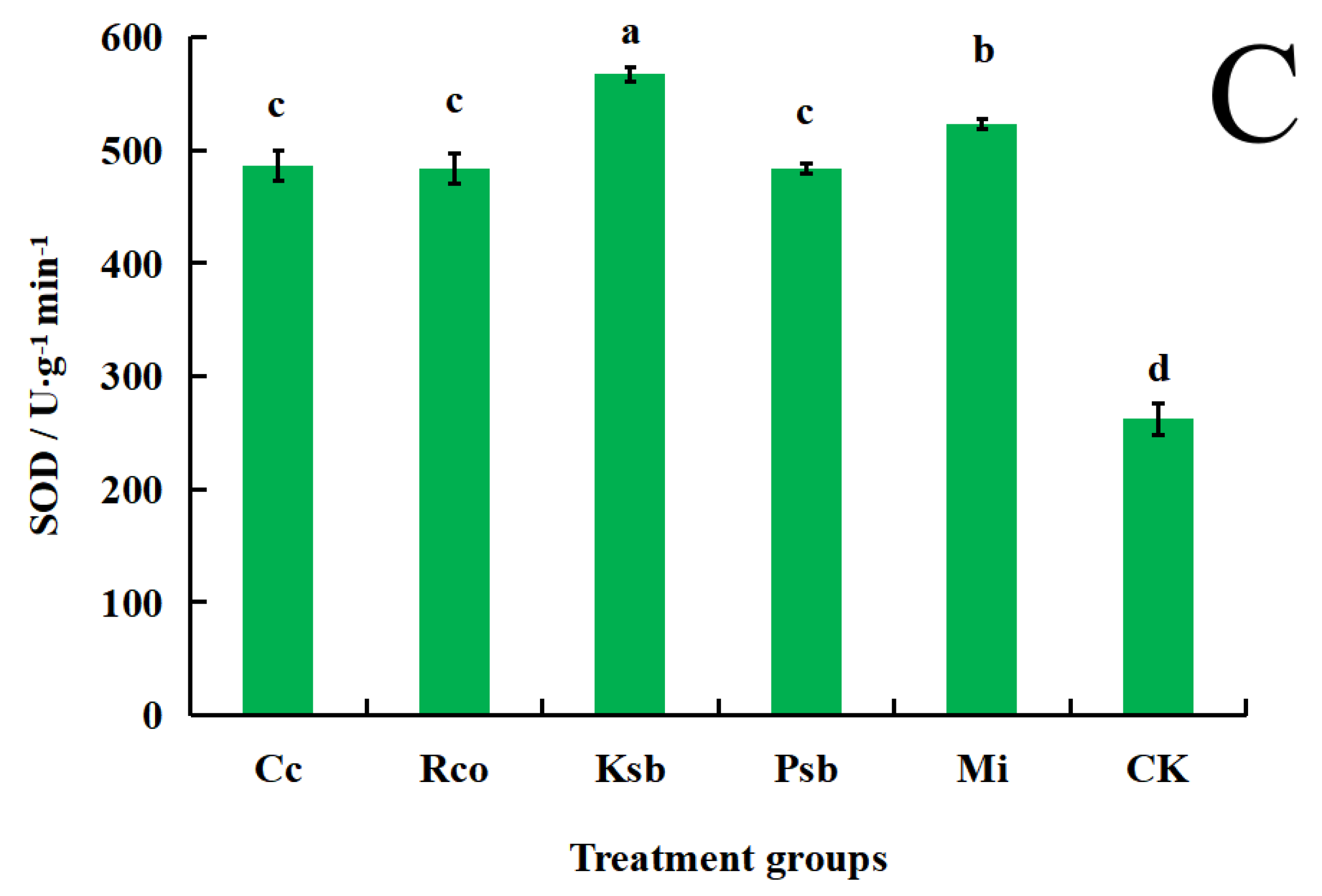
3.3. Effects of Different Microbial Inoculum Treatments on the Activities of Protective Enzymes in the Leaves of F. taipaiensis
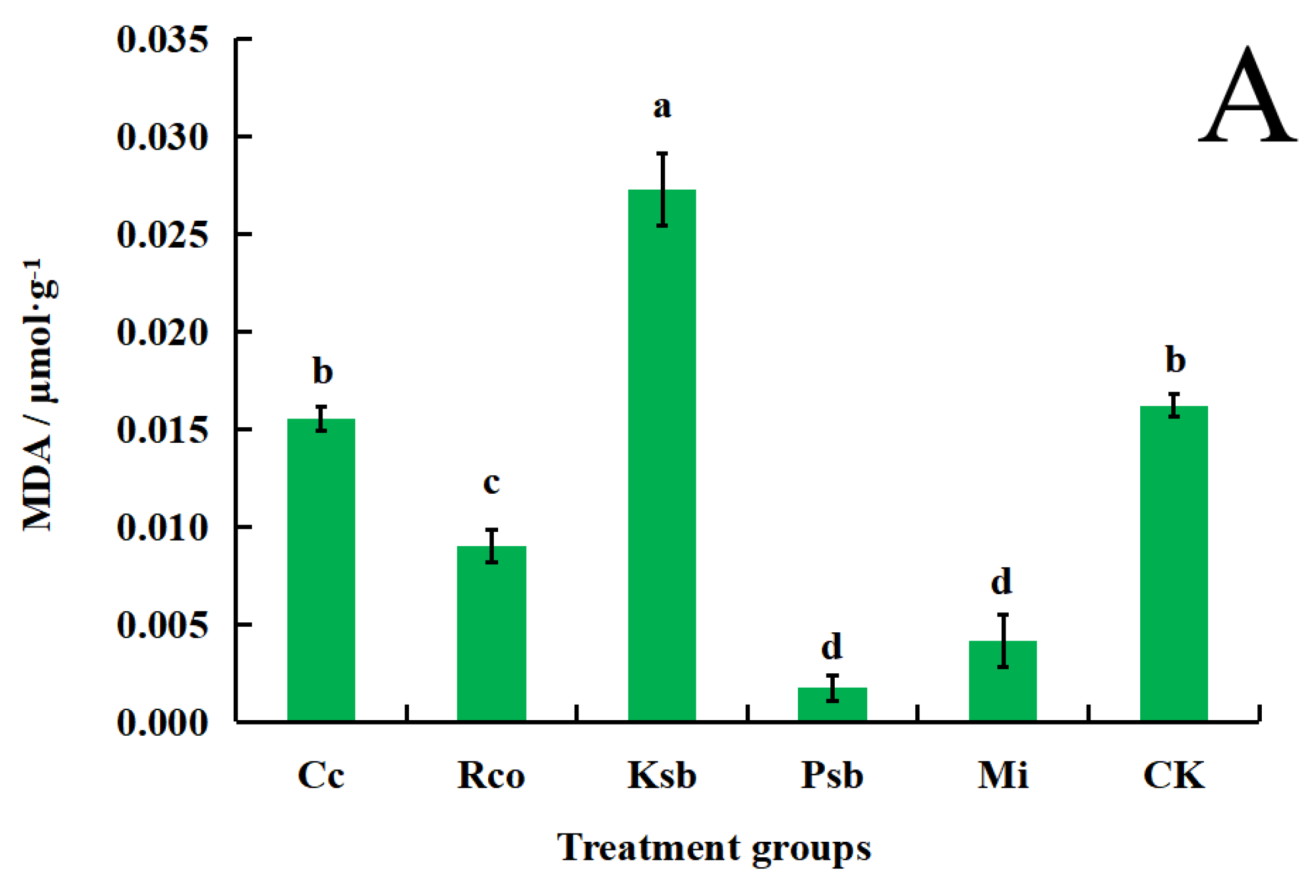
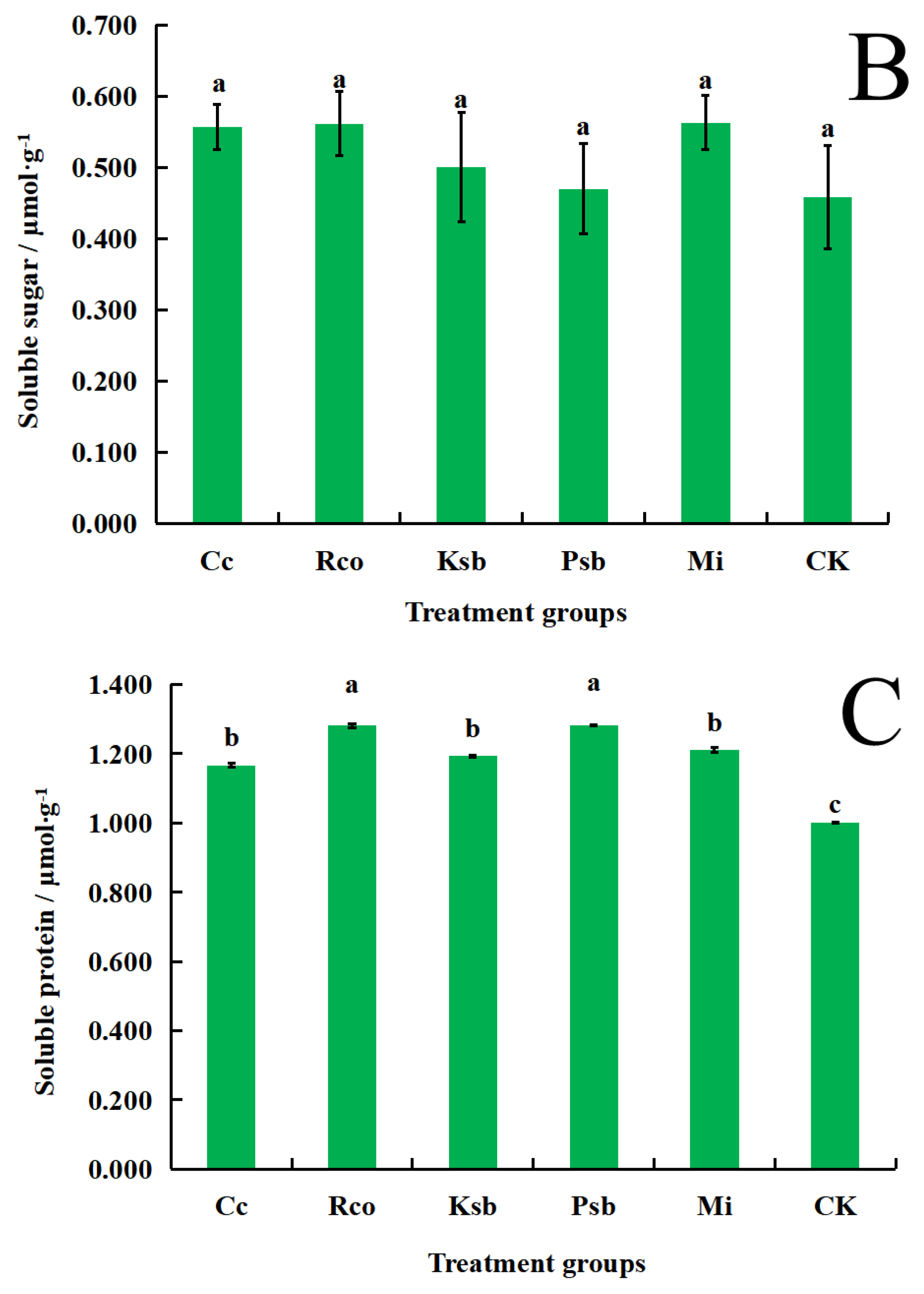
3.4. Effects of Different Microbial Inoculum Treatments on Malondialdehyde, Soluble Sugar, and Soluble Protein Content in the Leaves of F. taipaiensis
3.5. Effects of Different Microbial Inoculum Treatments on the Number of Culturable Bacteria, Fungi, and Actinomycetes in the Rhizospheric Soil of F. taipaiensis
3.6. Effects of Different Microbial Inoculum Treatments on Enzyme Activity in Rhizospheric Soil of F. taipaiensis
3.7. Effects of Different Microbial Inoculum Treatments on Nucleosides in the Bulb of F. taipaiensis
3.8. Effects of Different Microbial Inoculum Treatments on Alkaloids in the Bulb of F. taipaiensis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shen, L.; Zhou, N.; Fu, S.-Z.; Yi, D.-Y.; Jia, H.; Chen, H.-Y.; Wu, Y.-M. Pharmacognostical study on cultivated Fritillaria taipaiensis. J. Chin. Med. Mater. 2014, 37, 45–49. [Google Scholar] [CrossRef]
- Huang, Y.B.; Liu, H.M.; Fang, C.X.; Yu, Y.; Chen, H.Z.; Zhang, S.Y.; Li, S.M.; Hui, A.O. Comparative study on the pharmacodynamic differences of the Anti-tussive and Anti-inflammatory effects of the alkaloids from different varieties of Fritillariae cirrhosae Bulbs. Tradit. Chin. Drug Res. Clin. Pharmacol. 2018, 29, 19–22. [Google Scholar] [CrossRef]
- Zhou, N.; Guo, D.Q.; Shen, L.; Chen, Q.Y.; Tan, Y. Comparative contents of four alkaloids in bulbs of Fritillaria taipaiensis and Fritillaria unibracteata. Food Sci. 2014, 35, 133–136. [Google Scholar] [CrossRef]
- Chinese Pharmacopoeia Commission. Chuanbeimu. In Pharmacopoeia of the People's Republic of China, 1st ed.; Zhao, Y.Y., Fan, Z.X., Huang, K.., Li, Z., Gao, Y.M., Eds.; China Medical Science Press: Beijing, China, 2015; p. 36. [Google Scholar]
- Wang, L.Z. Quality Study on Fritillarias Cirrhosae Bulbus. Master's Thesis, Peking Union Medical College, Beijing, China, 2013. [Google Scholar]
- Fu, S.Z.; Yuan, D.M.; Ou, Y.J.; Li, J.D.; Lu, Q.F. Discussion on the development history and countermeasures of Fritillaria Taipaiensis P.Y.Li in chongqing. Lishizhen Med. Mater. Med. Res. 2014, 25, 476–477. [Google Scholar] [CrossRef]
- Zhou, N.; Ding, B.; Guo, D.Q.; Zhang, D.Q.; Li, S.S. Comparative study on the microstructure of stem and leaf between wild and cultivated Fritillaria Taipaiensis P. Y. Li. J. Chin. Med. Mater. 2015, 38, 77–80. [Google Scholar] [CrossRef]
- Fu, S.Z. Ecological types and protection countermeasures of wild Fritillaria Taipaiensis P. Y. Li in three gorges area of Yangtze River. Lishizhen Med. Mater. Med. Res. 2012, 23, 2018–2019. [Google Scholar] [CrossRef]
- Dong, T.W. The Effect of Combined Application of Nitrogen Phosphorus Fertilizer and Microbial Fertilizer on Growth and Quality of Two-Year-old Fritillaria Taipaiensis P. Y. Li. Master’s Thesis, Northwest A&F University, Xianyang, China, 2018. [Google Scholar]
- Peng, R.; Ma, P.; Mo, R.; Sun, N. Analysis of the bioactive components from different growth stages of Fritillaria taipaiensis P. Y. Li. Acta Pharm. Sin. B 2013, 3, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Mu, M.J.; Zhou, X.Q.; Guo, D.Q.; Wang, Q.; Yang, M.; Zhang, H.; Zhou, N. Effect of growth years to the soil enzyme activities and heavy metal residue of Fritillaria taipaiensis P. Y. Li. Environ. Chem. 2019, 38, 1966–1972. [Google Scholar] [CrossRef]
- Xiao, Y.H.; Li, Q.; Liu, Z.X.; Li, Y.J.; Li, G.; Long, H. Advances in studies on rhizospheric microorganism of medicinal plants. Chin. Tradit. Herb. Drugs 2013, 44, 497–504. [Google Scholar] [CrossRef]
- Qiao, Q.M.; Cheng, M.G.; Wang, X.M. The potential of rhizosphere microbes in relieveing continuous cropping obstacle of medicinal plants. Chin. J. Soil Sci. 2009, 40, 957–961. [Google Scholar] [CrossRef]
- Geng, S.J.; Wang, B.; Liu, K.; Shang, H.Y.; Quan, J.J.; Lu, X.P.; Shen, Q.R. Effect of specific microbial fertilizer on the rhizosphere microbial flora of different continuous cropping obstacles soil. Jiangsu J. Agric. Sci. 2012, 28, 758–764. [Google Scholar] [CrossRef]
- Schübler, A.; Schwarzott, D.; Walker, C. A new fungal phylum, the Glomeromycota: Phylogenny and evolution. Mycol. Res. 2001, 105, 1413–1421. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Yano, K. Nitrogen delivery to maize via mycorrhizal hyphae depends on the form of N supplied plant. Cell Environ. 2005, 28, 1247–1254. [Google Scholar] [CrossRef]
- Etesami, H.; Jeong, B.R.; Glick, B.R. Contribution of Arbuscular Mycorrhizal Fungi, Phosphate–Solubilizing Bacteria, and Silicon to P Uptake by Plant. Front. Plant Sci. 2021, 12, 699618. [Google Scholar] [CrossRef]
- Mortimer, P.E.; Pérez-Fernández, M.; Valentine, A.J. Arbuscular mycorrhiza maintains nodule function during external NH4+ supply in Phaseolus vulgaris (L.). Mycorrhiza 2012, 22, 237–245. [Google Scholar] [CrossRef]
- Wang, S.; Chen, A.; Xie, K.; Yang, X.; Luo, Z.; Chen, J.; Zeng, D.; Ren, Y.; Yang, C.; Wang, L.; et al. Functional analysis of the OsNPF4.5 nitrate transporter reveals a conserved mycorrhizal pathway of nitrogen acquisition in plants. Proc. Natl. Acad. Sci. USA 2020, 117, 16649–16659. [Google Scholar] [CrossRef]
- Wang, Y.; Xing, D.; Song, L.L.; Han, S.Y.; Chen, T.S. Effects of arbuscular mycorrhizal fungi on nutrient exchange in mulberry plant in rocky desertification areas. Chin. J. Tropical. Crop. 2020, 41, 7–14. [Google Scholar] [CrossRef]
- Balestrini, R.; Brunetti, C.; Cammareri, M.; Caretto, S.; Cavallaro, V.; Cominelli, E.; De Palma, M.; Docimo, T.; Giovinazzo, G.; Grandillo, S.; et al. Strategies to Modulate Specialized Metabolism in Mediterranean Crops: From Molecular Aspects to Field. Int. J. Mol. Sci. 2021, 22, 2887. [Google Scholar] [CrossRef]
- Yu, M.; Xie, W.; Zhang, X.; Zhang, S.; Wang, Y.; Hao, Z.; Chen, B. Arbuscular Mycorrhizal Fungi Can Compensate for the Loss of Indigenous Microbial Communities to Support the Growth of Liquorice (Glycyrrhiza uralensis Fisch.). Plants 2019, 9, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santander, C.; Aroca, R.; Ruiz-Lozano, J.M.; Olave, J.; Cartes, P.; Borie, F.; Cornejo, P. Arbuscular mycorrhiza effects on plant performance under osmotic stress. Mycorrhiza 2017, 27, 639–657. [Google Scholar] [CrossRef]
- Sanmartín, N.; Pastor, V.; Pastor-Fernández, J.; Flors, V.; Pozo, M.J.; Sánchez-Bel, P. Role and mechanisms of callose priming in mycorrhiza-induced resistance. J. Exp. Bot. 2020, 71, 2769–2781. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wang, B.W.; Yan, X.F. Effect of arbuscular mycorrhiza on camptothecin content in Camptotheca acuminata seedlings. Acta. Ecologica. Sinica 2006, 4, 1057–1062. [Google Scholar] [CrossRef]
- Guo, Q.S.; Cheng, L.T.; Liu, Z.Y. Study on influence of arbuscular mycorrhizal fungi Pinellia ternata yield and chemical composition. China J. Chin. Mater. Med. 2010, 3, 333–338. [Google Scholar] [CrossRef]
- Huang, J.-H.; Tan, J.-F.; Jie, H.-K.; Zeng, R.-S. Effects of inoculating arbuscular mycorrhizal fungi on Artemisia annua growth and its officinal components. J. Appl. Ecol. 2011, 22, 1443–1449. [Google Scholar] [CrossRef]
- Saini, I.; Aggarwal, A.; Kaushik, P. Inoculation with Mycorrhizal Fungi and Other Microbes to Improve the Morpho-Physiological and Floral Traits of Gazania rigens (L.) Gaertn. Agriculture 2019, 9, 51. [Google Scholar] [CrossRef] [Green Version]
- Vosnjak, M.; Likar, M.; Osterc, G. The Effect of Mycorrhizal Inoculum and Phosphorus Treatment on Growth and Flowering of Ajania (Ajania pacifica (Nakai) Bremer et Humphries). Plant Hortic. 2021, 7, 178. [Google Scholar] [CrossRef]
- Mongkolthanaruk, W. Classification of Bacillus Beneficial Substances Related to Plants, Humans and Animals. J. Microbiol. Biotechnol. 2012, 22, 1597–1604. [Google Scholar] [CrossRef] [Green Version]
- Kloepper, J.W.; Ryu, C.-M.; Zhang, S. Induced Systemic Resistance and Promotion of Plant Growth by Bacillus spp. Phytopathology 2004, 94, 1259–1266. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Tang, W.; Yang, F.; Meng, J.; Chen, W.; Li, X. Influence of biochar application on potassium-solubilizing Bacillus mucilaginosus as potential biofertilizer. Prep. Biochem. Biotechnol. 2017, 47, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.M.; Li, N.K.; Zhao, G.Y.; Zhang, S.F. Research progress on the phosphate-solubilizing and potassium-solubilizing microorganism. J. Hebei Univ. 2015, 35, 442–448. [Google Scholar] [CrossRef]
- Gundala, P.B.; Chinthala, P.; Sreenavasulu, B. A new facultative alkaliphilic potassium solubilizing Bacillus sp. SVUNM9 isolated from mica cores of Nellore District, Andhra Pradesh, India: Research and reviews. J. Microbiol. Biotechn. 2013, 2, 1–7. [Google Scholar]
- Bhatt, K.; Maheshwari, D.K. Zinc solubilizing bacteria (Bacillus megaterium) with multifarious plant growth promoting activities alleviates growth in Capsicum annuum L. 3 Biotech 2020, 10, 36. [Google Scholar] [CrossRef]
- Kang, S.-M.; Radhakrishnan, R.; You, Y.-H.; Joo, G.-J.; Lee, I.-J.; Lee, K.-E.; Kim, J.-H. Phosphate Solubilizing Bacillus megaterium mj1212 Regulates Endogenous Plant Carbohydrates and Amino Acids Contents to Promote Mustard Plant Growth. Indian J. Microbiol. 2014, 54, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Zhang, L.; Zhou, J.; George, T.S.; Feng, G. Arbuscular mycorrhizal fungi enhance mineralisation of organic phosphorus by carrying bacteria along their extraradical hyphae. New Phytol. 2020, 230, 304–315. [Google Scholar] [CrossRef]
- Feitosa de Souza, T.A.; Rodriguez-Echeverría, S.; Alves de Andrade, L.; Freitas, H. Arbuscular mycorrhizal fungi in Mimosa tenuiflora (Willd.) Poir from Brazilian semi-arid. Braz. J. Microbiol. 2016, 47, 359–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Delden, S.H.; Nazarideljou, M.J.; Marcelis, L.F.M. Nutrient solutions for Arabidopsis thaliana: A study on nutrient solution composition in hydroponics systems. Plant Methods 2020, 16, 72. [Google Scholar] [CrossRef]
- Philips, J.M.; Hayman, D.S. Improved procedures for clearing and attaining parasitic and vesicular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–162. [Google Scholar] [CrossRef]
- Trouvelot, A.; Kough, J.L.; Gianinazzi, P.V. Mesure du taux de mycorhization VA d'un système radiculaire. Recherche de méthodes d'estimation ayant une signification fonctionnelle. In Physiological and Genetical Aspects of Mycorrhizae, 1st ed.; Gianinazzi-Pearson, V., Gianinazzi, S., Eds.; INRA Publications: Paris, France, 1986; pp. 154–156. [Google Scholar]
- Zhang, Z.L.; Qu, W.J.; Li, X.F. Plant Physiology Experiment Instruction, 1st ed.; Higher Education Press: Beijing, China, 2009. [Google Scholar]
- Wright, S.F.; Upadhyaya, A. A survey of soils for aggregate stability and glomalin, a glyco protein produced by hyphae of arbuscular mycorrhizal fungi. Plant Soil 1998, 198, 97–107. [Google Scholar] [CrossRef]
- Lin, X.G. Principles and Methods of Soil Microbial Research, 1st ed.; Higher Education Press: Beijing, China, 2010. [Google Scholar]
- Yan, J.; Han, X.Z.; Wang, S.Q.; Li, X.H.; Zhu, W.W. Effects of different nitrogen forms on microbial quantity and enzymes activities in soybean field. J. Plant. Nutr. Fertil. 2010, 16, 341–347. [Google Scholar]
- Guan, S.Y. Soil Enzyme and Its Research Method, 1st ed.; Agricultural Press: Beijing, China, 1986. [Google Scholar]
- You, J.; Zhang, D.Q.; Pan, X.J.; Zhang, H.; Zhou, N.; Yu, J.J. Simultaneous determination of nine nucleosides contents using high performance liquid chromatography in Fritillaria taipaiensis P. Y. Li and Fritillaria unibracteata Hsiao et K. C. Hsia. Food Ferment. Ind. 2016, 42, 174–179. [Google Scholar] [CrossRef]
- Zhou, N.; Zhang, D.-Q.; Sun, Q.; Jiang, B.; Huang, Z.-C. Effects of fungal elicitors on the secondary metabolite steroidal saponin in Paris polyphylla var. yunnanensis. Acta Pharm. Sin. 2012, 47, 1237–1242. [Google Scholar] [CrossRef]
- Wei, Z.-X.; Guo, D.-Q.; Li, H.-F.; Ding, B.; Zhang, J.; Zhou, N.; Yu, J. Photosynthetic parameters and physiological indexes of Paris polyphylla var. yunnanensis influenced by arbuscular mycorrhizal fungi. China J. Chin. Mater. Med. 2015, 40, 3945–3952. [Google Scholar] [CrossRef]
- Wang, W.-L.; Du, J.-B.; Xu, F.-L.; Zhang, X.-H. Effect of fertilization levels on soil microorganism amount and soil enzyme activities. China J. Chin. Mater. Med. 2013, 38, 3851–3856. [Google Scholar] [CrossRef]
- Mu, M.-J.; Zhang, D.-G.; Zhang, H.; Yang, M.; Guo, D.-Q.; Zhou, N. Correlation between rhizospheric microorganisms distribution and alkaloid content of Fritillaria taipaiensis. China J. Chin. Mater. Med. 2019, 44, 2231–2235. [Google Scholar] [CrossRef]
- Li, W.; Sun, L.Y.; Zhang, C.P.; Xu, M.; Guo, Z.X. Distribution characteristic of arbuscular mycorrhizal fungi in the rhizosphere of endangered Lilium tsingtauense plants. J. Northeast Agric. Univ. 2016, 47, 30–37. [Google Scholar] [CrossRef]
- Ortíz-Castro, R.; Valencia-Cantero, E.; López-Bucio, J. Plant growth promotion by Bacillus megaterium involves cytokinin signaling. Plant Signal. Behav. 2008, 3, 263–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muchate, N.S.; Nikalje, G.; Rajurkar, N.S.; Suprasanna, P.; Nikam, T.D. Plant Salt Stress: Adaptive Responses, Tolerance Mechanism and Bioengineering for Salt Tolerance. Bot. Rev. 2016, 82, 371–406. [Google Scholar] [CrossRef]
- Kamali, S.; Mehraban, A. Effects of Nitroxin and arbuscular mycorrhizal fungi on the agro-physiological traits and grain yield of sorghum (Sorghum bicolor L.) under drought stress conditions. PLoS ONE 2020, 15, e0243824. [Google Scholar] [CrossRef]
- Soltani Kazemi, J.; Aboutalebian, M.A.; Hamzei, J.; Meskarbashee, M. Mycorrhiza and seed priming effect to improve the balance of sodium and potassium and some changes in sodium and potassium and some changes in antioxidants in the leaves of maize (Zea mays L.) under soil salinity. J. Agr. 2018, 17, 18–27. [Google Scholar] [CrossRef]
- Nosheen, A.; Bano, A.; Yasmin, H.; Keyani, R.; Habib, R.; Shah, S.T.A.; Rabia, N. Protein quantity and quality of saf-flower seed improved by NP fertilizer and rhizobacteria (Azospirillum and Azotobacter spp.). Front. Plant Sci. 2016, 7, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azmat, A.; Yasmin, H.; Hassan, M.N.; Nosheen, A.; Naz, R.; Sajjad, M.; Ilyas, N.; Akhtar, M.N. Co-application of bio-fertilizer and salicylic acid improves growth, photosynthetic pigments and stress tolerance in wheat under drought stress. PeerJ 2020, 8, e9960. [Google Scholar] [CrossRef]
- Abdel-Salam, E.; Alatar, A.; El-Sheikh, M.A. Inoculation with arbuscular mycorrhizal fungi alleviates harmful effects of drought stress on damask rose. Saudi J. Biol. Sci. 2018, 25, 1772–1780. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.L.; Luo, X.G.; Li, B.Q.; Li, Z.F.; Yang, S.; Zhong, X.M.; Wu, H.L. Isolation of Bacillus mucilaginosus and its influence on cotton seed germination in the desert sand. Agric. Res. Arid. Area. 2015, 33, 77–82. [Google Scholar] [CrossRef]
- Haynes, K.M.; Preston, M.; McLaughlin, J.W.; Webster, K.; Basiliko, N. Dissimilar bacterial and fungal decomposer communities across rich to poor fen peatlands exhibit functional redundancy. Can. J. Soil Sci. 2015, 95, 219–230. [Google Scholar] [CrossRef] [Green Version]
- Gu, W.C.; Zhang, J.; Zhou, N.; Pan, X.J.; Ding, B.; Qi, J.S.; Yang, M. Effect of different arbuscular mycorrhizal fungi combinations and inoculation periods on rhizosphere soil physicochemical properties and microbial quantity of Paris polyphylla var. yunnanensis seedlings. Chin. J. Exp. Tradit. Med. Form. 2020, 26, 116–130. [Google Scholar] [CrossRef]
- Ou, H.; Guo, D.-Q.; Lin, J.-J.; Yan, Z.-Y.; Teng, Z.; Wang, G.-Z.; Zhou, N. Effects of Different AM Fungi on Quantity and Enzyme Activity of Rhizosphere Soil Microorganism of Paris polyphylla var. yunnanensis. J. Chin. Med. Mater. 2016, 39, 948–955. [Google Scholar]
- Liao, H.B.; Li, Y.X.; Shao, J.J.; Fang, F.; Guo, W.D.; Chen, W.R. Impacts of continuous cropping on Fritillaria thunbergii Miq. growth and rhizosphere soil properties. Chin. J. Ecol. 2011, 30, 2203–2208. [Google Scholar] [CrossRef]
- Yokoyama, D.; Imai, N.; Kitayama, K. Effects of nitrogen and phosphorus fertilization on the activities of four different classes of fine-root and soil phosphatases in Bornean tropical rain forests. Plant Soil 2017, 416, 463–476. [Google Scholar] [CrossRef]
- Fu, X.F.; Zhang, G.P.; Zhang, X.W.; Ren, J.H. Effects of PSB and AMF on growth, microorganisms and soil enzyme activities in the rhizosphere of Taxus chinensis var.mairei Seedlings. Acta Bot. Boreal. Occident. Sin. 2016, 36, 353–360. [Google Scholar] [CrossRef]
- Jacobson, K.A.; Jarvis, M.F.; Williams, M. Purine and Pyrimidine (P2) Receptors as Drug Targets. J. Med. Chem. 2002, 45, 4057–4093. [Google Scholar] [CrossRef] [PubMed]
- Ralevic, V.; Burnstock, G. Receptors for purines and pyrimidines. Pharmacol. Rev. 1998, 50, 413–492. [Google Scholar] [CrossRef] [PubMed]
- Carver, J.; Walker, W.A. The role of nucleotides in human nutrition. J. Nutr. Biochem. 1995, 6, 58–72. [Google Scholar] [CrossRef]
- Chen, B.Y.; Ma, H.H.; Yang, T.; Cheng, Z.M.; Niu, X.X.; Ma, X.W. Effect of different microbial fertilizer dosage on the components of Fritillaria pallidiflora Schvek. Xinjiang Agric. Sci. 2017, 54, 871–877. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Carotenoid (mg·g−1) | Chlorophyll a (mg·g−1) | Chlorophyll b (mg·g−1) | Chlorophyll (a + b) (mg·g−1) |
|---|---|---|---|---|
| CK | 84.08 ± 0.39 d | 0.96 ± 0.01 d | 0.29 ± 0.01 d | 1.25 ± 0.05 d |
| Cc | 134.07 ± 0.93 b | 1.61± 0.02 b | 0.45 ± 0.01 b | 2.06 ± 0.03 b |
| Rco | 135.76 ± 0.59 b | 1.64 ± 0.01 ab | 0.45 ± 0.01 b | 2.09 ± 0.01 bc |
| Ksb | 119.92 ± 2.40 c | 1.44 ± 0.04 c | 0.40 ± 0.01 c | 1.84 ± 0.02 bc |
| Psb | 141.65 ± 0.56 a | 1.66 ± 0.01 a | 0.48 ± 0.01 a | 2.14 ± 0.01 a |
| Mi | 119.33 ± 0.28 c | 1.47 ± 0.01 c | 0.40 ± 0.01 c | 1.88 ± 0.01 c |
| Group. | Bacteria (lgCFU/g) | Fungi (lgCFU/g) | Actinomycetes (lgCFU/g) | Total Microorganisms (lgCFU/g) | Bacteria/Fungi | Actinomycetes/Fungi |
|---|---|---|---|---|---|---|
| CK | 5.36 ± 0.26 c | 3.55 ± 0.05 d | 4.40 ± 0.04 c | 5.42 ± 0.23 b | 80.10 ± 4.78 e | 7.31 ± 0.30 c |
| Cc | 5.88 ± 0.22 b | 3.72 ± 0.05 ab | 4.69 ± 0.08 a | 5.92 ± 0.20 a | 164.94 ± 7.69 c | 9.42 ± 0.39 a |
| Rco | 5.92 ± 0.11 ab | 3.67 ± 0.04 c | 4.50 ± 0.03 b | 5.94 ± 0.09 a | 186.36 ± 5.38 b | 6.77 ± 0.91 c |
| Ksb | 6.03 ± 0.09 a | 3.65 ± 0.01 c | 4.54 ± 0.03 b | 6.05 ± 0.09 a | 255.30 ± 8.86 a | 8.12 ± 0.61 b |
| Psb | 5.84 ± 0.05 b | 3.73 ± 0.05 a | 4.52 ± 0.03 b | 5.87 ± 0.05 a | 145.05 ± 6.17 d | 6.48 ± 0.45 c |
| Mi | 5.93 ± 0.14 ab | 3.83 ± 0.08 a | 4.74 ± 0.10 a | 5.97 ± 0.05 a | 135.48 ± 4.36 d | 8.29 ± 0.81 ab |
| Group | Protease (mg·kg−1) | Urease (g·kg−1) | Acid Phosphatase (g·kg−1) | Neutral Phosphatase (g·kg−1) | Alkaline Phosphatase (g·kg−1) | Catalase (g·kg−1) | Invertase (g·kg−1) |
|---|---|---|---|---|---|---|---|
| CK | 29.286 ± 0.104 d | 15.719 ± 0.077 d | 0.255 ± 0.016 d | 0.162 ± 0.012 c | 0.037 ± 0.013 d | 0.162 ± 0.004 f | 12.604 ± 0.062 e |
| Cc | 64.035 ± 0.051 b | 16.617 ± 0.086 d | 0.812 ± 0.069 b | 0.264 ± 0.008 a | 0.086 ± 0.019 b | 0.324 ± 0.011 d | 12.955 ± 0.001 e |
| Rco | 54.266 ± 0.028 c | 33.081 ± 0.034 b | 0.541 ± 0.018 c | 0.237 ± 0.030 a | 0.118 ± 0.009 a | 0.384 ± 0.002 b | 25.074 ± 0.087 c |
| Ksb | 29.686 ± 0.154 d | 28.924 ± 0.001 c | 0.871 ± 0.036 b | 0.190 ± 0.014 bc | 0.087 ± 0.002 bc | 0.426 ± 0.025 a | 32.223 ± 0.074 b |
| Psb | 78.242 ± 0.019 a | 41.365 ± 0.027 a | 0.778 ± 0.056 b | 0.208 ± 0.035 ab | 0.089 ± 0.007 bc | 0.339 ± 0.064 c | 36.624 ± 0.039 a |
| Mi | 72.726 ± 0.047 a | 33.766 ± 0.004 b | 2.442 ± 0.071 a | 0.171 ± 0.023 bc | 0.039 ± 0.004 cd | 0.206 ± 0.024 e | 14.467 ± 0.003 d |
| Group | Uracil | Cytidine | Guanine | Uridine | Adenine | Guanosine | Thymidine | Adenosine | 2’-Deoxyadenosine | Total |
|---|---|---|---|---|---|---|---|---|---|---|
| CK | 38.385 ± 0.640 a | 24.730 ± 1.318 b | 23.745 ± 1.633 e | 223.737 ± 1.178 f | 51.757 ± 0.826 a | 232.013 ± 1.648 f | 28.042 ± 0.998 f | 310.870 ± 0.340 e | 26.373 ± 1.435 e | 951.932 ± 2.678 f |
| Cc | 16.355 ± 0.710 c | 48.862 ± 0.233 a | 30.296 ± 0.093 d | 339.488 ± 0.371 c | 28.650 ± 0.455 d | 321.895 ± 1.367 b | 98.703 ± 0.453 a | 377.707 ± 0.272 c | 106.974 ± 2.469 a | 1352.576 ± 0.423 c |
| Rco | 13.114 ± 0.316 d | 27.358 ± 1.425 b | 60.227 ± 1.633 b | 271.344 ± 0.490 e | 36.767 ± 0.074 b | 260.075 ± 1.633 e | 51.107 ± 0.540 e | 340.540 ± 0.306 d | 57.505 ± 4.764 d | 982.581 ± 1.799 e |
| Ksb | 17.628 ± 0.268 c | 25.236 ± 0.825 b | 87.911 ± 2.267 a | 343.810 ± 0.182 b | 24.047 ± 0.222 e | 316.040 ± 2.397 c | 84.102 ± 0.647 b | 399.131 ± 1.059 b | 91.504 ± 0.325 b | 1370.242 ± 0.185 b |
| Psb | 16.584 ± 0.682 c | 25.124 ± 0.816 b | 46.803 ± 0.801 c | 279.541 ± 0.468 d | 33.324 ± 0.946 c | 266.124 ± 1.676 d | 58.658 ± 1.788 d | 346.949 ± 0.301 d | 59.356 ± 4.563 d | 1110.430 ± 8.132 d |
| Mi | 23.143 ± 0.230 b | 52.328 ± 1.633 a | 24.280 ± 0.486 e | 362.572 ± 1.152 a | 51.382 ± 0.099 a | 364.171 ± 1.135 a | 76.347 ± 1.420 c | 417.247 ± 0.977 a | 78.514 ± 0.944 c | 1426.841 ± 6.054 a |
| Group | Peimisine | Total Alkaloids |
|---|---|---|
| CK | 0.26 ± 0.01 c | 0.57 ± 0.01 e |
| Cc | 0.39 ± 0.01 a | 1.04 ± 0.01 b |
| Rco | 0.40 ± 0.01 a | 0.84 ± 0.01 d |
| Ksb | 0.33 ± 0.01 b | 1.08 ± 0.01 ab |
| Psb | 0.33 ± 0.01 b | 1.10 ± 0.02 a |
| Mi | 0.32 ± 0.01 b | 0.90 ± 0.03 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, N.; Mu, M.; Yang, M.; Zhou, Y.; Ma, M. The Effect of Microbial Fertilizer on the Growth, Rhizospheric Environment and Medicinal Quality of Fritillaria taipaiensis. Horticulturae 2021, 7, 500. https://doi.org/10.3390/horticulturae7110500
Zhou N, Mu M, Yang M, Zhou Y, Ma M. The Effect of Microbial Fertilizer on the Growth, Rhizospheric Environment and Medicinal Quality of Fritillaria taipaiensis. Horticulturae. 2021; 7(11):500. https://doi.org/10.3390/horticulturae7110500
Chicago/Turabian StyleZhou, Nong, Maojun Mu, Min Yang, You Zhou, and Mingguo Ma. 2021. "The Effect of Microbial Fertilizer on the Growth, Rhizospheric Environment and Medicinal Quality of Fritillaria taipaiensis" Horticulturae 7, no. 11: 500. https://doi.org/10.3390/horticulturae7110500