
Hydrogen Gas Improves Seed Germination in Cucumber by Regulating Sugar and Starch Metabolisms



Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Hydrogen-Rich Water
2.2. Plant Material and Hydrogen-Rich Water Treatment
2.3. Determination of Morphological Indexes and Seed Vitality
2.4. Determination of Soluble Sugar, Soluble Protein, and Starch Contents
2.5. Determination of Total Amylase and α- and β-Amylase Activities
2.6. Total RNA Extraction
2.7. Determination of AMY, BMY, SS4, and SS3 Gene Expression
2.8. Statistical Analysis
3. Result
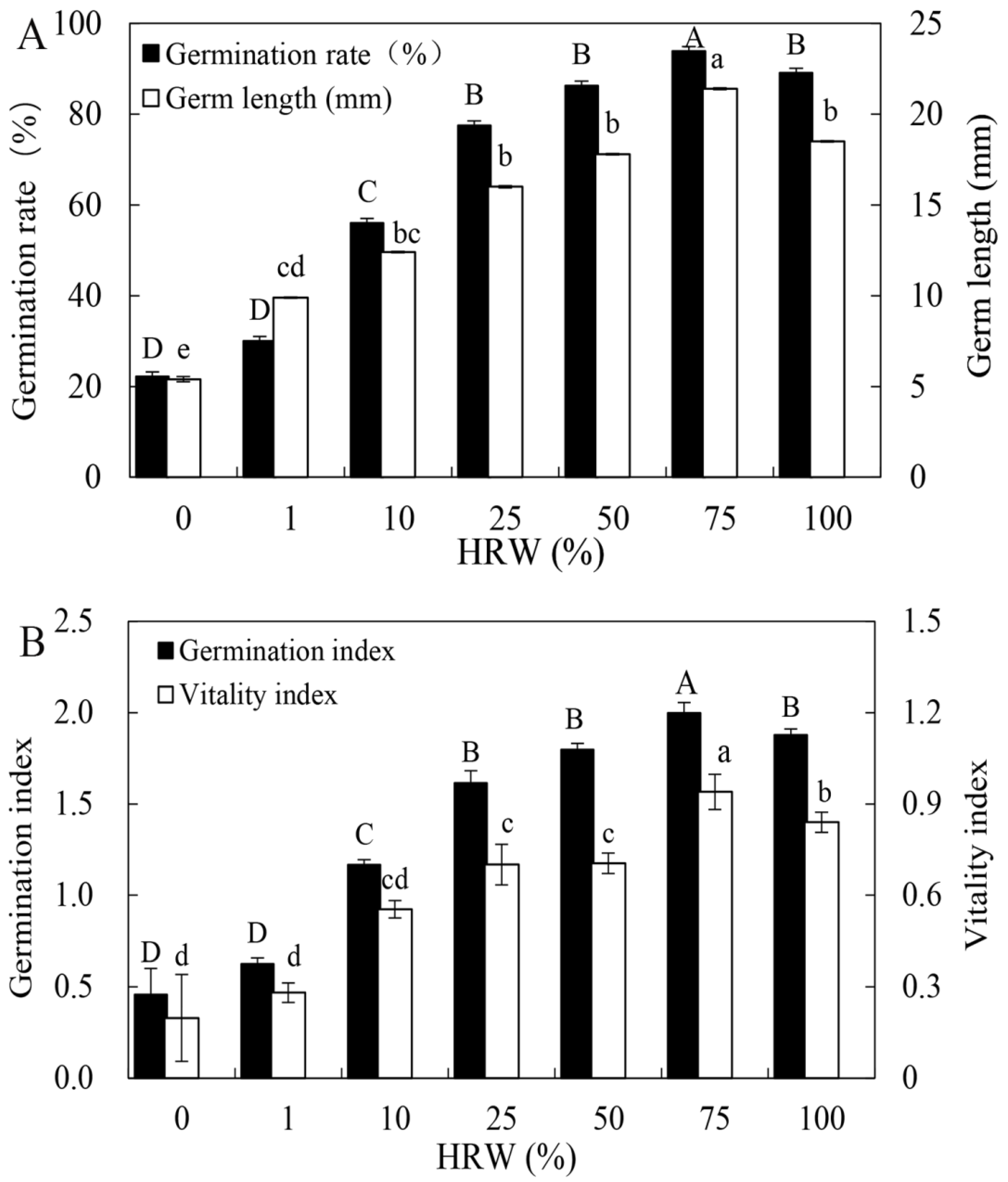
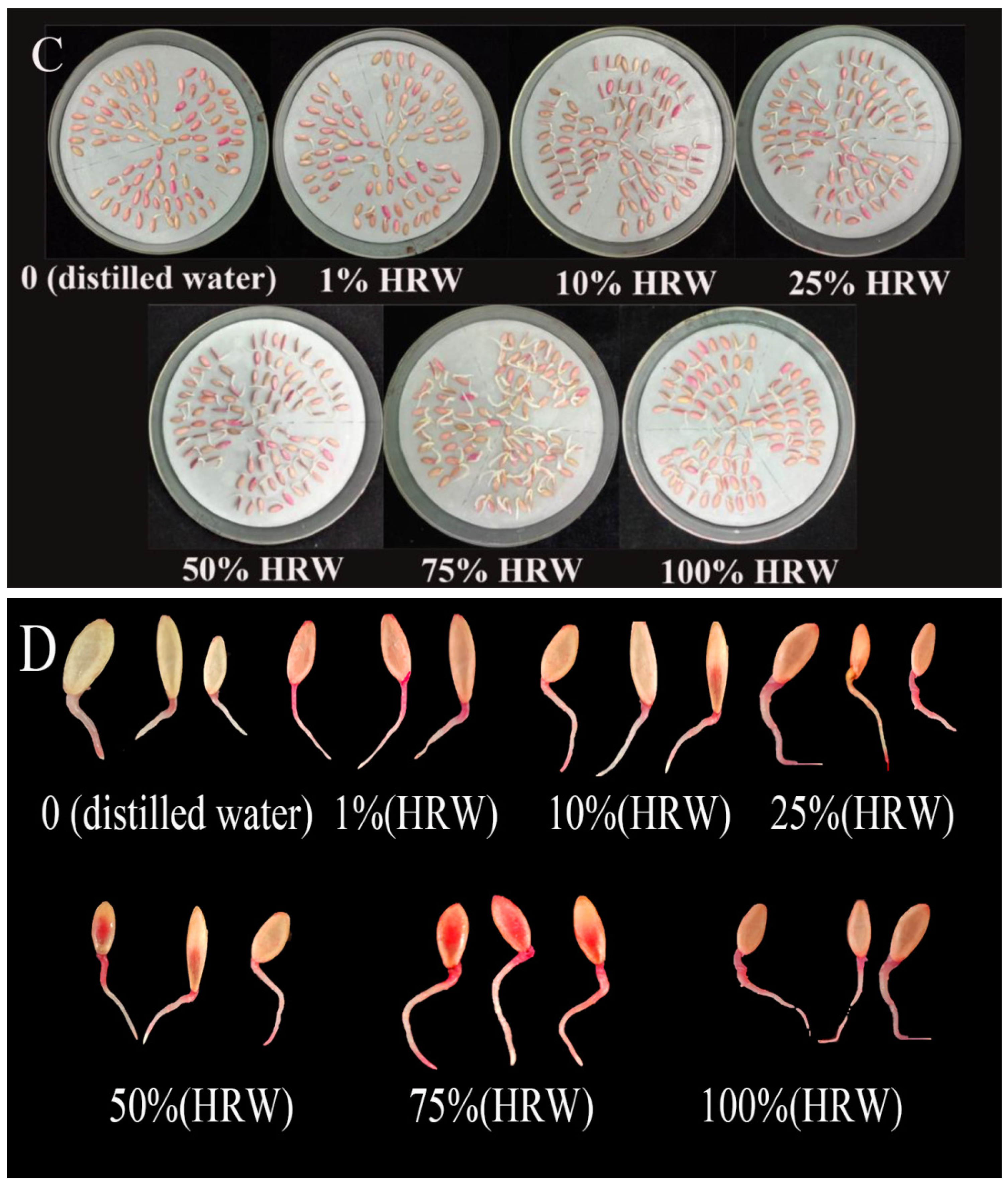
3.1. Effect of HRW on Seed Germination in a Dose-Dependent Manner
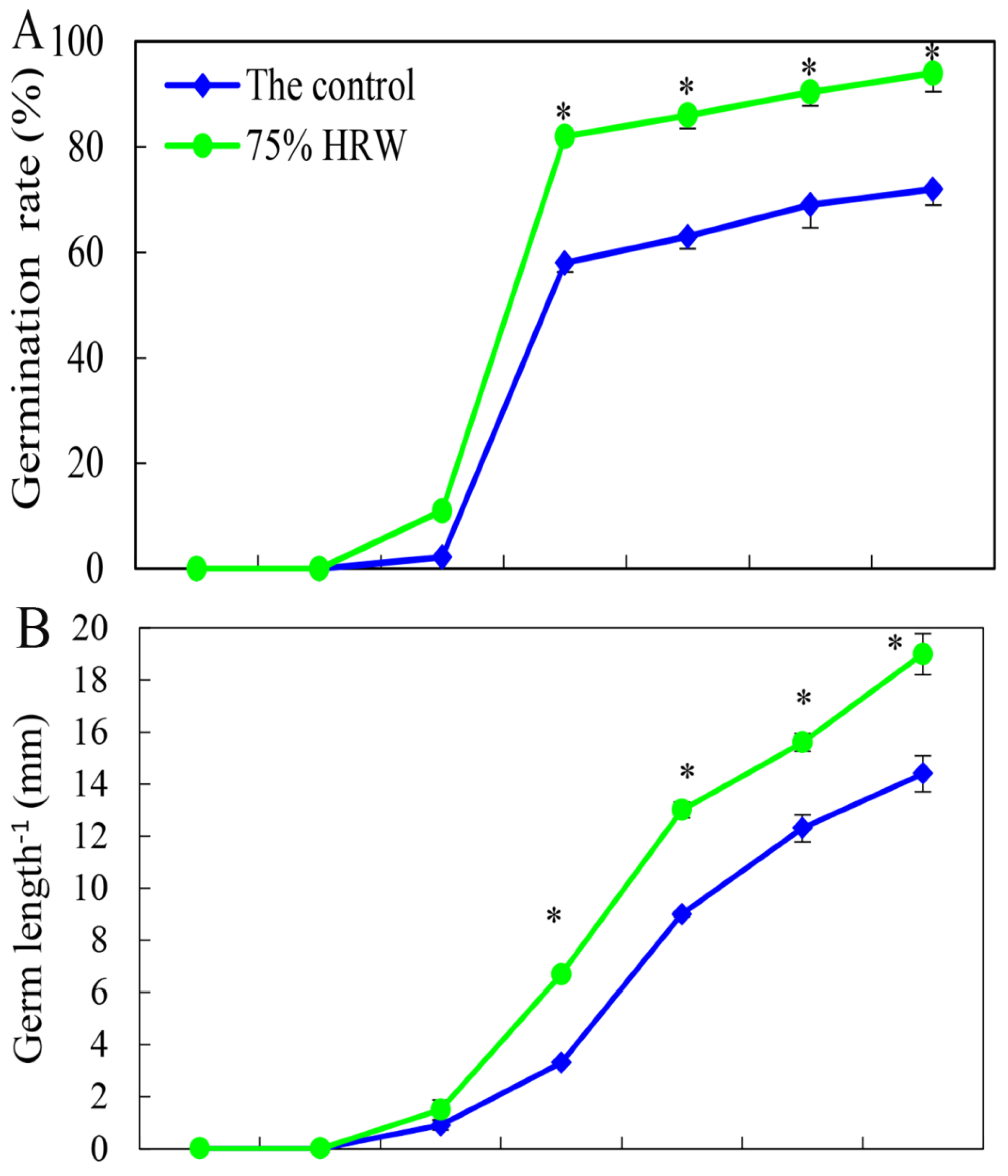
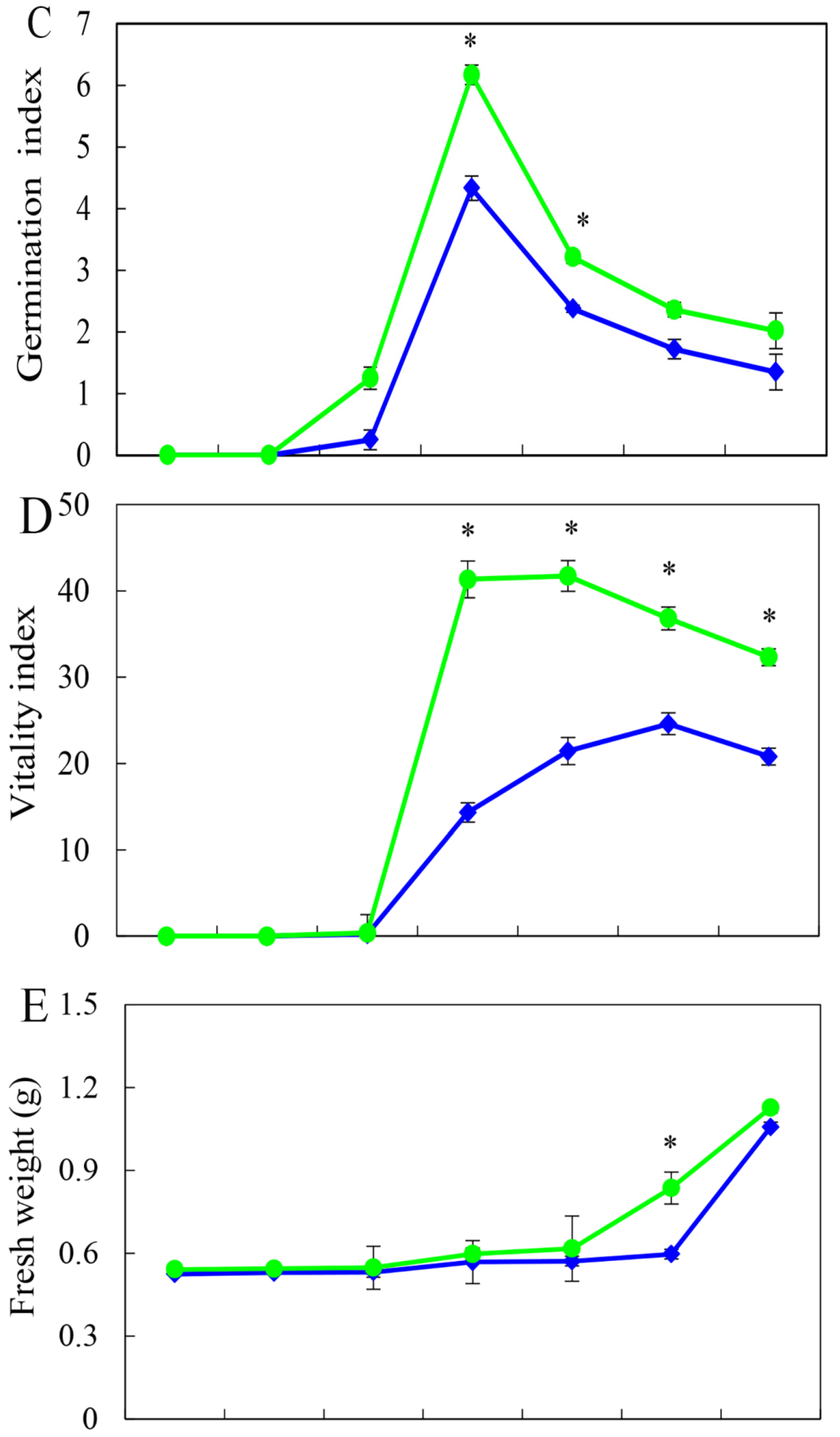
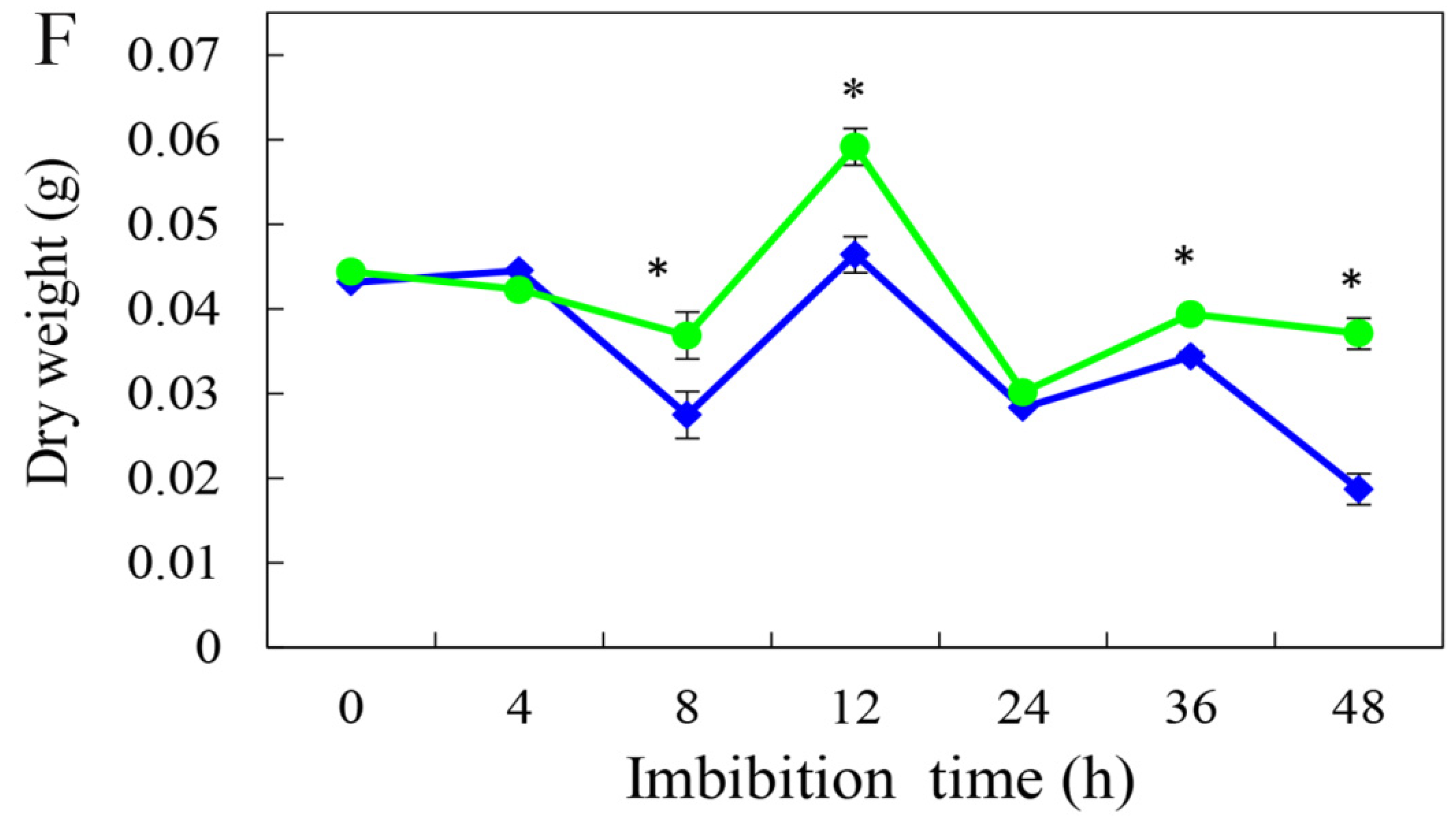
3.2. Effect of HRW on Seed Germination in a Time-Dependent Manner
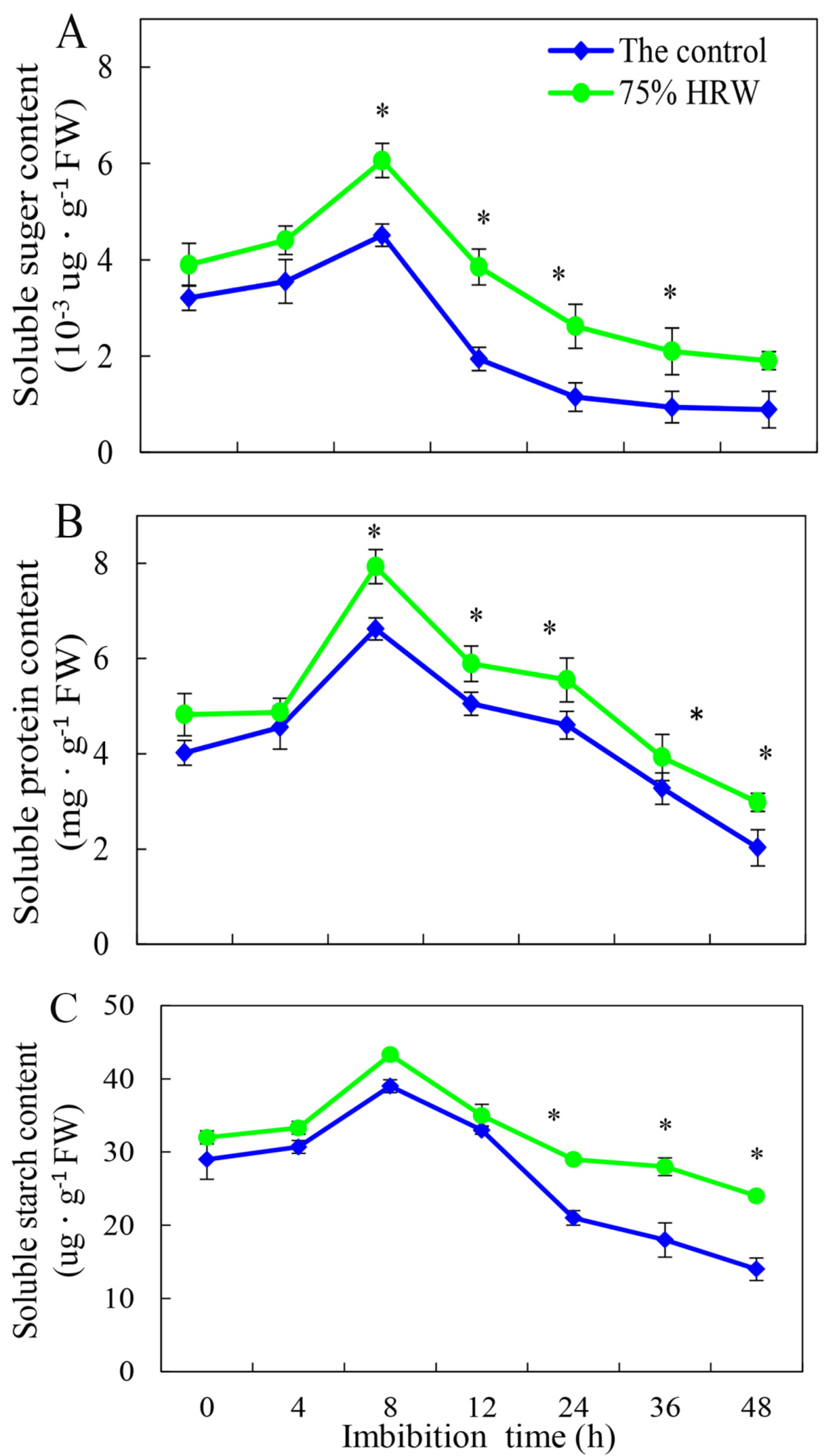
3.3. Effect of HRW on Total Soluble Sugar, Total Soluble Protein, and Total Soluble Starch Contents during Germination
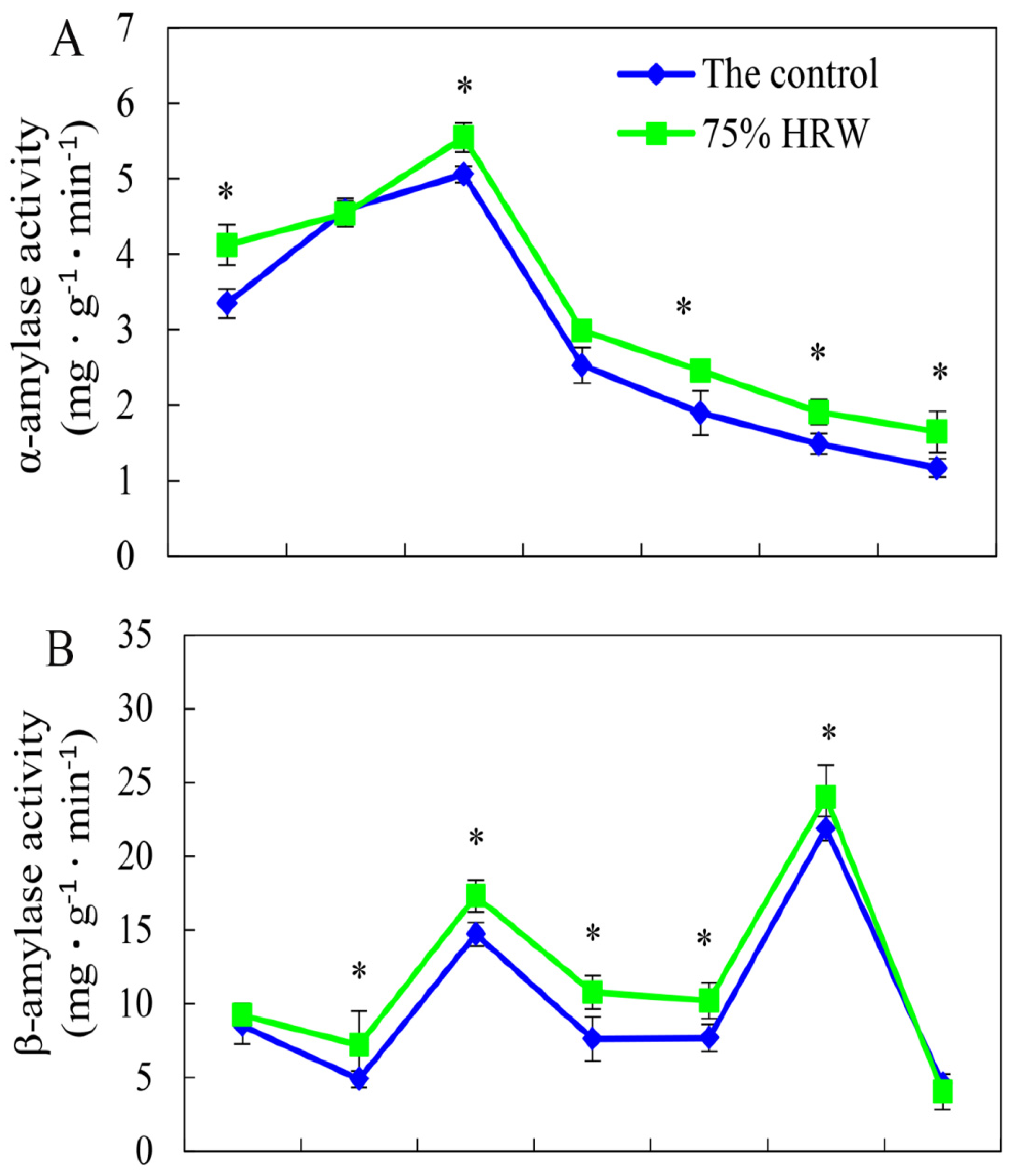
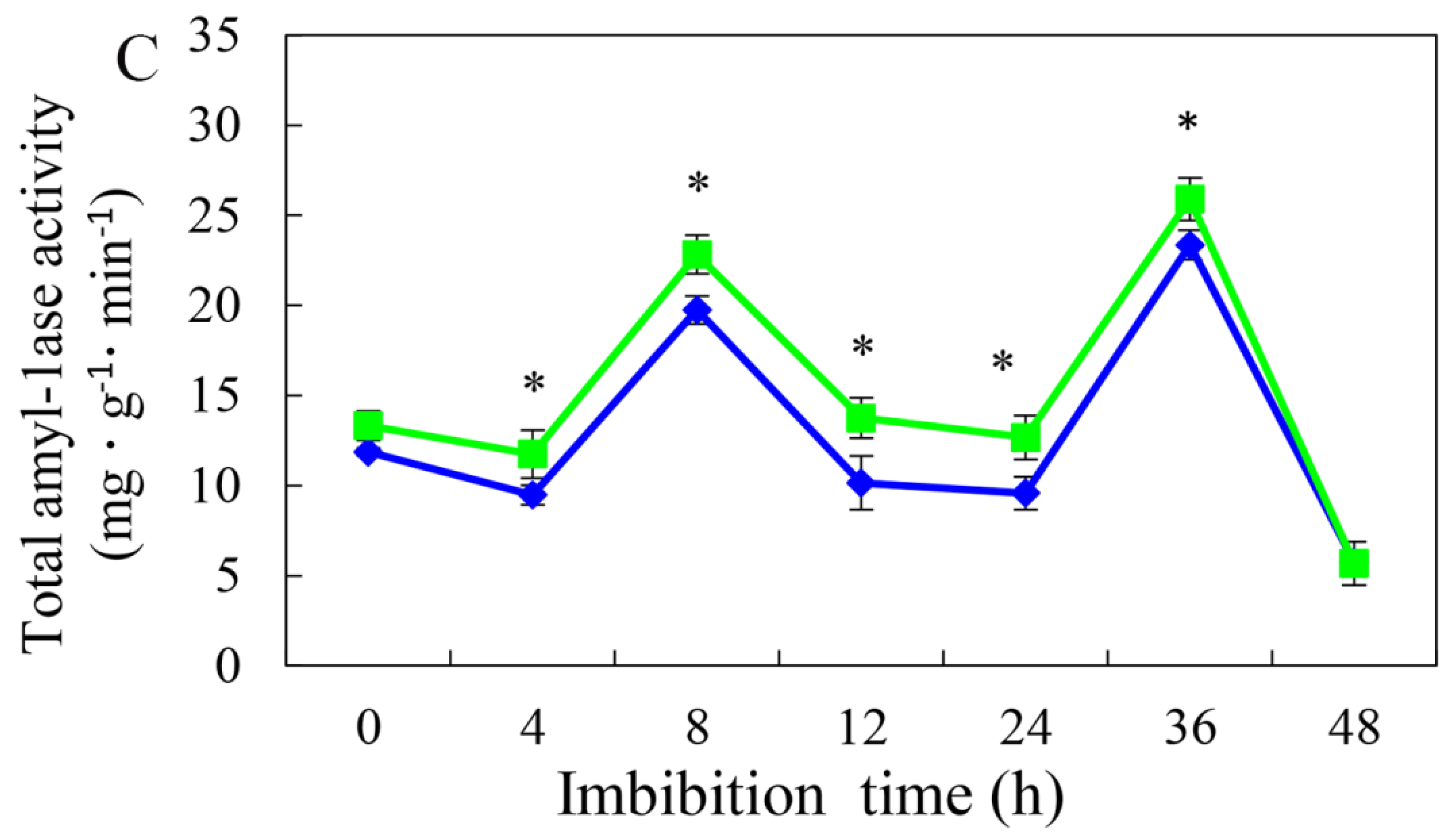
3.4. Effect of HRW on Total Amylase and α- and β-Amylase Activities during Germination
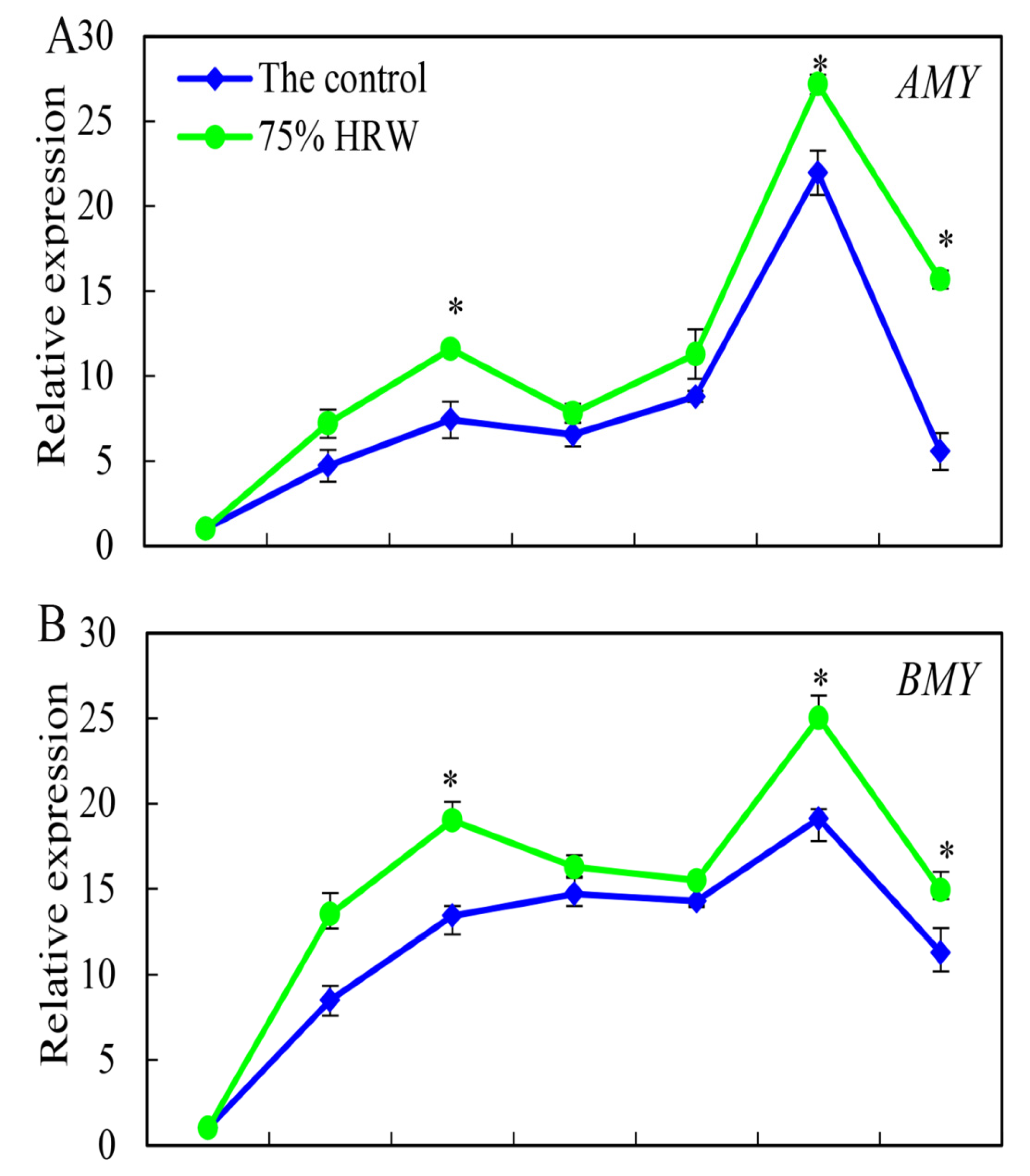
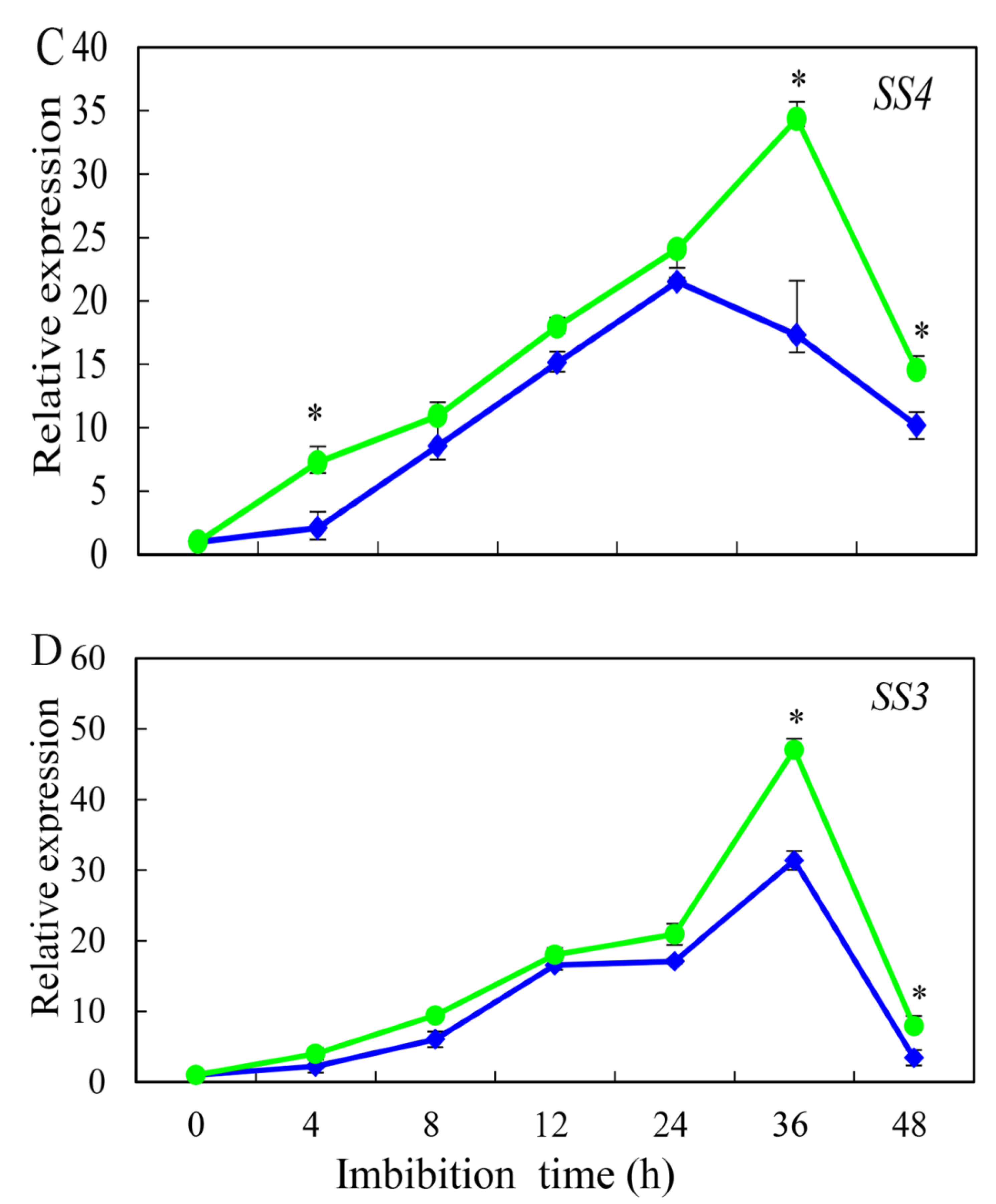
3.5. Effect of HRW on AMY, BMY, SS3, and SS4 Gene Expression during Germination
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mizuno, K.; Sasaki, A.T.; Ebisu, K.; Tajima, K.; Kajimoto, O.; Nojima, J.; Kuratsune, H.; Hori, H.; Watanabe, Y. Hydrogen-rich water for improvements of mood, anxiety, and autonomic nerve function in daily life. Med. Gas Res. 2018, 7, 247–255. [Google Scholar]
- Cui, W.T.; Gao, C.Y.; Fang, P.; Lin, G.Q.; Shen, W.B. Alleviation of cadmium toxicity in Medica gosativa by hydrogen-rich water. J. Hazard. Mater. 2013, 260, 715–724. [Google Scholar] [CrossRef] [PubMed]
- Jin, Q.J.; Zhu, K.K.; Cui, W.T.; Xie, Y.J.; Han, B.; Shen, W.B. Hydrogen gas acts as a novel bioactive molecule in enhancing plant tolerance to paraquat induced oxidative stress via the modulation of heme oxygenase-1 signalling system. Plant Cell Environ. 2013, 36, 956–969. [Google Scholar] [CrossRef]
- Wu, Q.; Su, N.N.; Cai, J.T.; Shen, Z.G.; Cui, J. Hydrogen-rich water enhances cadmium tolerance in Chinese cabbage by reducing cadmium uptake and increasing antioxidant capacities. J. Plant Physiol. 2015, 175, 174–182. [Google Scholar] [CrossRef]
- Lin, Y.T.; Zhang, W.; Qi, F.; Cui, W.T.; Xie, Y.J.; Shen, W.B. Hydrogen-rich water regulates cucumber adventitious root development in a heme oxygenase-1/carbon monoxide-dependent manner. J. Plant Physiol. 2014, 171, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Su, N.N.; Wu, Q.; Liu, Y.Y.; Cai, J.T.; Shen, W.B.; Xia, K.; Cui, J. Hydrogen-rich water re-establishes ROS homeostasis but exerts differential effects on anthocyanin synthesis in two varieties of radish sprouts under UV-A irradiation. J. Agric. Food Chem. 2014, 62, 6454–6462. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hao, H.; Chen, M.; Wang, H.; Feng, Z.; Chen, H. Hydrogen-rich water alleviates the toxicities of different stresses to my celial growth in Hypsizygus marmoreus. AMB Express 2017, 7, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Gong, T.; Bian, B.; Liao, W. Roles of hydrogen gas in plants: A review. Funct. Plant Biol. 2018, 45, 8. [Google Scholar] [CrossRef]
- Xie, Y.J.; Mao, Y.; Lai, D.W.; Zhang, W.; Shen, W.B. H2 enhances arabidopsis salt tolerance by manipulating ZAT10/12-mediated antioxidant defence and controlling sodium exclusion. PLoS ONE 2012, 7, e49800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.N.; Zhao, X.Q.; Wang, Z.Q.; Shen, W.B.; Xu, X.M. Protective effects of hydrogen-rich water on the photosynthetic apparatus of maize seedlings (Zea mays L.) as a result of an increase in antioxidant enzyme activities under high light stress. Plant Growth Regul. 2015, 77, 43–56. [Google Scholar] [CrossRef]
- Xu, S.; Jiang, Y.L.; Cui, W.T.; Jin, Q.J.; Zhang, Y.H.; Bu, D.; Fu, J.Y.; Wang, R.; Zhou, F.; Shen, W.B. Hydrogen enhances adaptation of rice seedlings to cold stress via the reestablishment of redox homeostasis mediated by miRNA expression. Plant Soil 2017, 414, 53–67. [Google Scholar] [CrossRef]
- Ren, P.; Jin, X.; Liao, W.; Wang, M.; Niu, L.; Li, X.P.; Xu, X.P.; Zhu, X.P. Effect of hydrogen-rich water on vase life and quality in cut lily and rose flowers. Hortic. Environ. Biotechnol. 2017, 58, 576–584. [Google Scholar] [CrossRef]
- Su, J.; Nie, Y.; Zhao, G.; Cheng, D.; Wang, R.; Chen, J.; Zhang, S.; Shen, W. Endogenous Hydrogen gas delays petal senescence and extends the vase life of lisianthus cut flowers. Postharvest Biol. Technol. 2019, 147, 148–155. [Google Scholar] [CrossRef]
- Wang, C.L.; Fang, H.; Gong, T.Y.; Zhang, J.; Niu, L.J.; Huang, D.J.; Huo, J.; Liao, W.B. Hydrogen gas alleviates postharvest senescence of cut rose ‘Movie star’ by antagonizing ethylene. Plant Mol. Biol. 2020, 102, 271–285. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Wang, C.L.; Wang, S.Y.; Liao, W.B. Hydrogen gas increases the vase life of cut rose ‘Movie star’ by regulating bacterial community in the stem ends. Postharvest Biol. Technol. 2021, 181, 111685. [Google Scholar] [CrossRef]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy, 3rd ed.; Seed ence Research; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Yang, L.; Liu, S.; Lin, R. The role of light in regulating seed dormancy and germination. J. Integer. Plant Biol. 2020, 62, 1310–1326. [Google Scholar] [CrossRef]
- Penfield, S.; MacGregor, D.R. Effects of environmental variation during seed production on seed dormancy and germination. J. Exp. Bot. 2017, 68, 819–825. [Google Scholar] [CrossRef] [Green Version]
- Munnȇ-Bosch, S.; Alegre, L. Cross-stress tolerance and stress ‘memory’ in plants: An integrated view. Environ. Exp. Bot. 2013, 94, 1–88. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, biogeography, and evolution of dormancy and germination. Crop. Sci. 2000, 40, 334. [Google Scholar]
- Lin, P.C.; Hwang, S.G.; Endo, A.; Okamoto, M.; Koshiba, T.; Cheng, W.H. Ectopicexpression of ABSCISIC ACID 2/GLUCOSE INSENSITIVE 1 in Arabidopsis promotes seed dormancy and stress tolerance. Plant Physiol. 2007, 143, 745–758. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Tran, L.S. Effects of ethylene on seed germination of halophyte plants under salt stress. Methods Mol. Biol. 2017, 1573, 253–259. [Google Scholar] [PubMed]
- Li, J.P.; Yan, E.; Yang, X.G.; Bai, J.; Long, K.Z.; Li, H.; Xu, N. Effects of exogenous H2S on the germination of tomato seeds under nitrate stress. J. Hortic. Sci. Biotechnol. 2015, 90, 39–46. [Google Scholar] [CrossRef]
- Wang, P.; Zhu, J.K.; Lang, Z. Nitric oxide suppresses the inhibitory effect of abscisic acid on seed germination by S-nitrosylation of SnRK2 proteins. Plant Signal. Behav. 2015, 10, e1031939. [Google Scholar] [CrossRef] [Green Version]
- Feil, R.; Lunn, J.E. Quantification of soluble sugars and sugar alcohols by LC-MS/MS. Methods Mol. Biol. 2018, 87–100. [Google Scholar]
- Kaur, S.; Gupta, A.K.; Kaur, N. Gibberellic acid and kinetin partially reverse the effect of water stress on germination and seedling growth in chickpea. Plant Growth Regul. 1998, 25, 29–33. [Google Scholar] [CrossRef]
- Liu, S.J.; Xu, H.H.; Wang, W.Q.; Li, N.; Wang, W.P.; Lu, Z.; Møller, I.M.; Song, S.Q. Identification of embryo proteins associated with seed germination and seedling establishment in germinating rice seeds. J. Plant Physiol. 2016, 196, 79–92. [Google Scholar] [CrossRef]
- Xu, S.; Zhu, S.S.; Jiang, Y.L.; Wang, N.; Wang, R.; Shen, W.B.; Yang, J. Hydrogen-rich water alleviates salt stress in rice during seed germination. Plant Soil 2013, 370, 47–57. [Google Scholar] [CrossRef]
- Zhang, C.P.; Li, Y.C.; Yuan, F.G.; Hu, S.J.; Liu, H.Y.; He, P. Role of 5-aminolevulinic acid in the salinity stress response of the seeds and seedlings of the medicinal plant Cassia obtusifolia L. Bot. Stud. 2013, 54, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, S.; Gupta, A.K.; Kaur, N. Effect of GA3, kinetin and in dole acetic acid on carbohydrate metabolism in chickpea seedlings germinating under water stress. Plant Growth Regul. 2000, 30, 61–70. [Google Scholar] [CrossRef]
- Bu, R.F.; Xiao, X.M.; Liao, W.B.; Hu, Y.F.; Li, J.; Lv, J.; Wang, R.D.; Xie, J.M. Exogenous Si Alleviation of Autotoxicity in Cucumber (Cucumis sativus L.) Seed Germination is Correlated with Changes in Carbohydrate Metabolism. J. Plant Growth Regul. 2018, 37, 784–793. [Google Scholar] [CrossRef]
- Su, J.C.; Preiss, J. Purification and proper ties of sucrose synthase from maize kernels. Plant Physiol. 1978, 61, 389–393. [Google Scholar] [CrossRef]
- Shannon, J.C. Carbon-14 distribution in carbohydrates of immature zea mays kernels following co (2) treatment of intact plants. Plant Physiol. 1968, 43, 1215–1220. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.P.; Liu, Y.C.; Dai, H.B.; Miao, M.M. Characteristics and expression patterns of six α-galactosidases in cucumber (Cucumis sativus L.). PLoS ONE 2021, 16, e0244714. [Google Scholar]
- Sacher, J.A.; Hatch, M.D.; Glasziou, K.T. Sugar accumulation cycle in sugar cane. iii. physical & metabolic aspects of cycle in immature storage tissues. Plant Physiol. 1963, 38, 348–354. [Google Scholar]
- Pharr, D.M.; Sox, H.N.; Smart, E.L.; Lower, R.L.; Fleming, H.P. Identification and distribution of soluble saccharides in pickling cucumber plants and their fate in fermentation. J. Am. Soc. Hortic. Sci. 1977, 102, 406409. [Google Scholar]
- Penfield, S.; Li, Y.; Gilday, A.D.; Graham, I.A. Arabidopsis ABA INSENSITIVE4 regulates lipid mobilization in the embryo and reveals repression of seed germination by the endosperm. Plant Cell. 2006, 18, 1887–1899. [Google Scholar] [CrossRef] [Green Version]
- Bailly, C.; El-Maarouf Bouteau, H.; Corbineau, F. From intracellular signaling networks to cell death: The dual role of reactive oxygen species in seed physiology. C. R. Biol. 2008, 331, 806–814. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, B.; Xu, Z.; Shi, Z.; Chen, S.; Huang, X.; Chen, S.; Wang, X. Involvement of reactive oxygen species in endosperm cap weakening and embryo elongation growth during lettuce seed germination. J. Exp. Bot. 2014, 65, 3189–3200. [Google Scholar] [CrossRef] [Green Version]
- Kittock, D.L.; Law, A.G. Relationship of seedling vitality to respiration and tetrazolium chloride reduction by germinating wheat seeds. Agron. J. 1968, 60, 286–288. [Google Scholar] [CrossRef]
- Vasquez-Tello, A.Y.; Zuily-Fodil, Y.; Thi, A.P.; Da Silva, J.V. Electrolyte and Pi leakages and soluble sugar content as physiological tests for screening resistance to water stress in Phaseolus and Vigna species. J. Exp. Bot. 1990, 41, 827–832. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Li, C.X.; Bian, B.T.; Gong, T.Y.; Liao, W.B. Comparative proteomic analysis of key proteins during abscisic acid-hydrogen peroxide-induced adventitious rooting in cucumber (Cucumis sativus L.) under drought stress. J. Plant Physiol. 2018, 229, 185–194. [Google Scholar] [CrossRef]
- Dan, Y.Y.; Niu, Y.; Wang, C.L.; Yan, M.; Liao, W.B. Genome-wide identification and expression analysis of the trehalose-6-phosphate synthase (TPS) gene family in cucumber (Cucumis sativus L.). Peer J. 2021, 9, e11398. [Google Scholar] [CrossRef]
- Rajjou, L.; Rajjou, L.; Gallardo, K.; Debeaujon, I.; Vandekerckhove, J.; Job, C.; Job, D. The effect of alpha-amanitin on the Arabidopsis seed proteome highlights the distinct roles of stored and neosyn the sized mRNAs during germination. Plant Physiol. 2004, 134, 1598–1613. [Google Scholar] [CrossRef] [Green Version]
- Jacobsen, J.V.; Hanson, A.D.; Chandler, P.C. Water stress enhances expression of an α-amylase gene in barley leaves. Plant Physiol. 1986, 80, 350–359. [Google Scholar] [CrossRef] [Green Version]
- Todaka, D.; Matsushima, H.; Morohashi, Y. Water stress enhances β-amylase activity in cucumber cotyledons. J. Exp. Bot. 2000, 51, 739–745. [Google Scholar] [CrossRef]
- Xu, X.; Liao, G.; Huang, C.; Zhong, M.; Jia, D.; Qu, X.; Liu, Q.; He, Y.; Li, Y. Differences of sucrose accumulation concentration and related genes expression between two sucrose accumulation types of Actinidia eriantha. Sci. Rep. 2020, 10, 20474. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagent Name (μL) | Determination of α-Amylase Activities | Determination of Total Amylase Activities | ||
|---|---|---|---|---|
| The Control Tube | The Test Tube | The Control Tube | The Test Tube | |
| Amylase stock solution | 250 | 250 | ||
| In 70 °C water bath for 15 min and cooled rapidly | ||||
| Amylase diluent | 250 | 250 | ||
| Distilled water | 250 | 250 | ||
| Reagent 2 | 250 | 250 | ||
| At 40 °C (±0.5 °C) constant temperature water bath for 5 min | ||||
| Reagent 1 | 500 | 500 | 500 | 500 |
| Gene | Full Gene Name | Accession No. a | Primer Pair (5′–3′) | Schematic Diagram |
|---|---|---|---|---|
| AMY | The alpha amylase | XM_004151148 | F:CACGGTTATTACACCCAGGACT |  |
| R:TAAATCATCTTCGTTGCCCAT | ||||
| BMY | The beta amylase | XM_004138543 | F:GGTGTCAAGTGGTAGCAACAATAAC |  |
| R:TGTCCTCTCTTTCTCTTCTAATGGTCT | ||||
| SS4 | The sucros synthase 4 | LOC101205508 | F: CCTGAACTTCTGCCATCTGCTATC |  |
| R: ACTGGGTGTGGCTTTGGTGAATG | ||||
| SS3 | The sucros synthase 4 | LOC101213925 | F: ATGGGAGCGTTCAATGACTGGAAG |  |
| R: ATGAATCTGACACGACCACCAATCC | ||||
| ACT | Actin | AB010922 | F: TTCTGGTGATGGTGTGAGTC |  |
| R: GGCAGTGGTGGTGAACATG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, P.; Li, C.; Liu, H.; Zhao, Z.; Liao, W. Hydrogen Gas Improves Seed Germination in Cucumber by Regulating Sugar and Starch Metabolisms. Horticulturae 2021, 7, 456. https://doi.org/10.3390/horticulturae7110456
Huang P, Li C, Liu H, Zhao Z, Liao W. Hydrogen Gas Improves Seed Germination in Cucumber by Regulating Sugar and Starch Metabolisms. Horticulturae. 2021; 7(11):456. https://doi.org/10.3390/horticulturae7110456
Chicago/Turabian StyleHuang, Panpan, Changxia Li, Huwei Liu, Zongxi Zhao, and Weibiao Liao. 2021. "Hydrogen Gas Improves Seed Germination in Cucumber by Regulating Sugar and Starch Metabolisms" Horticulturae 7, no. 11: 456. https://doi.org/10.3390/horticulturae7110456