
Foliar Supplementation of Clove Fruit Extract and Salicylic Acid Maintains the Performance and Antioxidant Defense System of Solanum tuberosum L. under Deficient Irrigation Regimes

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of the Trial Site and Soil Analysis
2.2. Plant Material and Irrigation Regimes
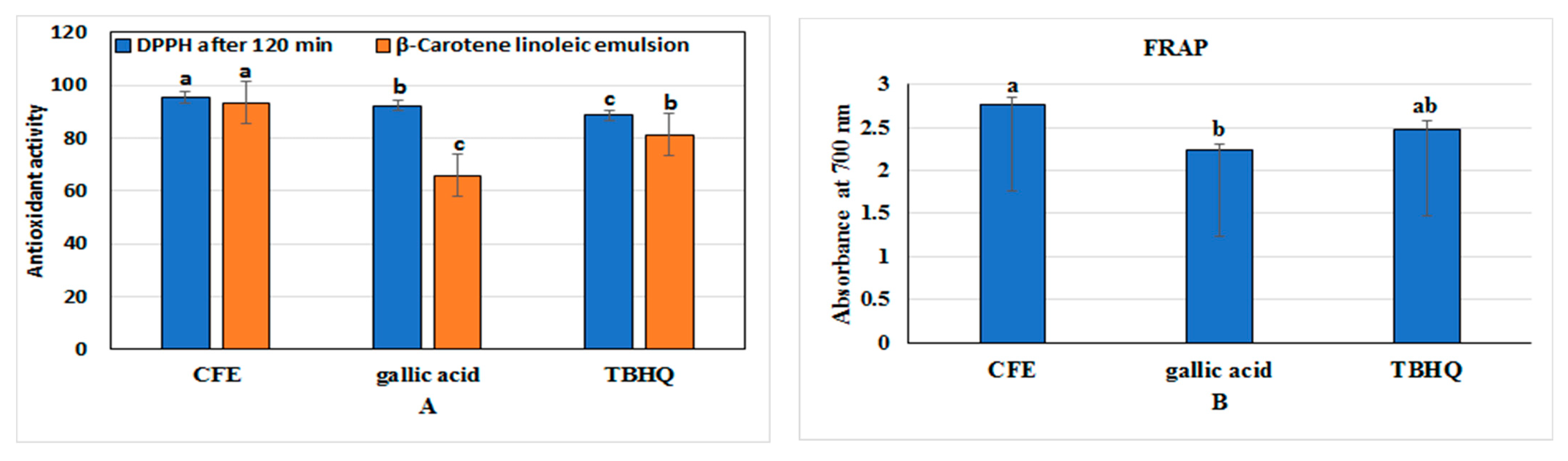
2.3. Preparation and Analysis of Clove Fruit Extract (CFE)
2.4. Applications of Clove Fruit Extract (CFE) and Salicylic Acid (SA)
2.5. Assessment of Attributes Related to Growth and Yield
2.6. Assessment of Attributes Related to Plant Physio-Biochemistry
2.7. Determination of Enzymatic and Non-Enzymatic Antioxidant Activities
2.8. Statistical Analysis
3. Results
3.1. Growth, Yield, and Photosynthetic Attributes
3.2. Cell and Membrane Integrity and Oxidative Stress Markers
3.3. Osmoprotectants and Antioxidantive Status
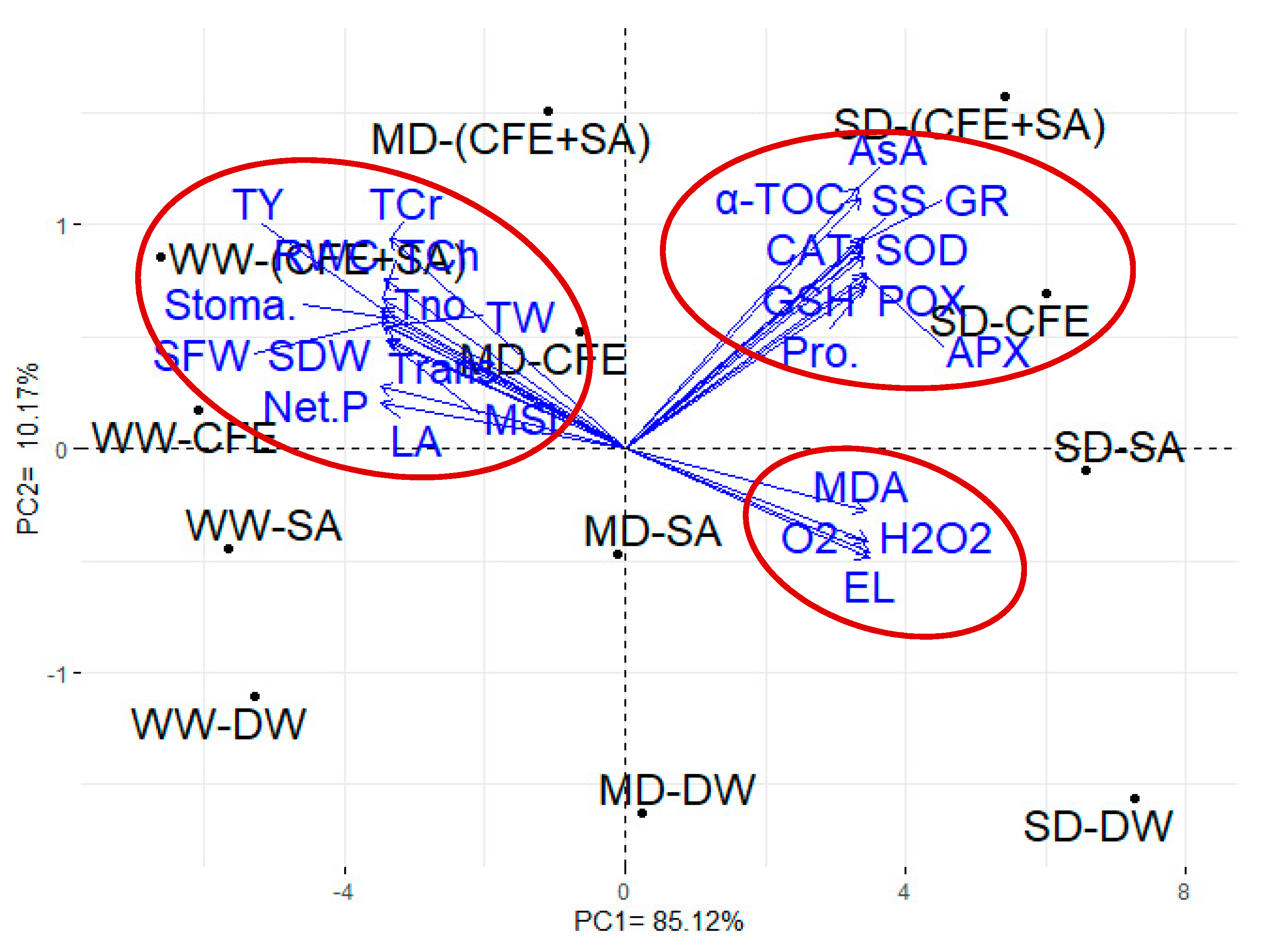
3.4. Traits Interrelationship
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lukianova, A.A.; Evseev, P.V.; Stakheev, A.A.; Kotova, I.B.; Zavriev, S.K.; Ignatov, A.N.; Miroshnikov, K.A. Quantitative Real-Time PCR Assay for the Detection of Pectobacterium parmentieri, a Causal Agent of Potato Soft Rot. Plants. Plants 2021, 10, 1880. [Google Scholar] [CrossRef] [PubMed]
- Bugarčić, Ž.; Bugarčić, R.A.; Đekić, R.; Ivan, J. A study of yields of Duch potato varieties in different agro-ecological condidtions in Serbia. J. Sci. Agric. Res. 2000, 61, 143–150. [Google Scholar]
- Chhogyel, N.; Kumar, L. Climate change and potential impacts on agriculture in Bhutan: A discussion of pertinent issues. Agric. Food Secur. 2018, 7, 1201313. [Google Scholar] [CrossRef]
- Ray, D.K.; West, P.C.; Clark, M.; Gerber, J.S.; Prishchepov, A.V.; Chatterjee, S. Climate change has likely already affected global food production. PLoS ONE 2019, 14, e0217148. [Google Scholar] [CrossRef]
- Papworth, A.; Maslin, M.; Randalls, S. Is climate change the greatest threat to global health? Geogr. J. 2015, 181, 413–422. [Google Scholar] [CrossRef]
- Li, J.; Li, X.; Yang, Q.; Luo, Y.; Gong, X.; Zhang, W.; Hu, Y.; Yang, T.; Dong, K.; Feng, B. Proteomic changes in the grains of foxtail millet (Setaria italica (L.) Beau) under drought stress. Span. J. Agric. Res. 2019, 17, 802. [Google Scholar] [CrossRef]
- Merwad, A.-R.M.; Desoky, E.-S.M.; Rady, M.M. Response of water deficit-stressed Vigna unguiculata performances to silicon, proline or methionine foliar application. Sci. Hortic. 2018, 228, 132–144. [Google Scholar] [CrossRef]
- Elrys, A.S.; Abdo, A.I.; Desoky, E.-S.M. Potato tubers contamination with nitrate under the influence of nitrogen fertilizers and spray with molybdenum and salicylic acid. Environ. Sci. Pollut. Res. 2018, 25, 7076–7089. [Google Scholar] [CrossRef] [PubMed]
- Abd Razak, A.f.; Ismail, M.R.; Karim, M.F.; Wahab, P.E.M.; Abdullah, S.N.; Kausar, H. Changes in leaf gas exchange, biochemical properties, growth and yield of chilli grown under soilless culture subjected to deficit fertigation. Aust. J. Crop Sci. 2013, 7, 1582–1589. [Google Scholar]
- Desoky, E.-S.M.; Mansour, E.; Yasin, M.A.; El Sobky, E.-S.E.; Rady, M.M. Improvement of drought tolerance in five different cultivars of Vicia faba with foliar application of ascorbic acid or silicon. Span. J. Agric. Res. 2020, 18, 16. [Google Scholar] [CrossRef]
- Mansour, E.; Desoky, E.-S.M.; Ali, M.M.; Abdul-Hamid, M.I.; Ullah, H.; Attia, A.; Datta, A. Identifying drought-tolerant genotypes of faba bean and their agro-physiological responses to different water regimes in an arid Mediterranean environment. Agric. Water Manag. 2021, 247, 106754. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Loreto, F.; Cornic, G.; Sharkey, T. Diffusive and metabolic limitations to photosynthesis under drought and salinity in C3 plants. Plant Biol. 2004, 6, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Desoky, E.-S.M.; Elrys, A.S.; Mansour, E.; Eid, R.S.; Selem, E.; Rady, M.M.; Ali, E.F.; Mersal, G.A.; Semida, W.M. Application of biostimulants promotes growth and productivity by fortifying the antioxidant machinery and suppressing oxidative stress in faba bean under various abiotic stresses. Sci. Hortic. 2021, 288, 110340. [Google Scholar] [CrossRef]
- Asada, K. The water-water cycle in chloroplasts: Scavenging of active oxygens and dissipation of excess photons. Annu. Rev. Plant Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef]
- Sitohy, M.Z.; Desoky, E.-S.M.; Osman, A.; Rady, M.M. Pumpkin seed protein hydrolysate treatment alleviates salt stress effects on Phaseolus vulgaris by elevating antioxidant capacity and recovering ion homeostasis. Sci. Hortic. 2020, 271, 109495. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef] [PubMed]
- Møller, I.M. Plant mitochondria and oxidative stress: Electron transport, NADPH turnover, and metabolism of reactive oxygen species. Annu. Rev. Plant Biol. 2001, 52, 561–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaki, S.S.; Rady, M.M. Moringa oleifera leaf extract improves growth, physiochemical attributes, antioxidant defence system and yields of salt-stressed Phaseolus vulgaris L. plants. Int. J. ChemTech Res. 2015, 8, 120–134. [Google Scholar]
- Rady, M.M.; Abd El-Mageed, T.A.; Abdurrahman, H.A.; Mahdi, A.H. Humic acid application improves field performance of cotton (Gossypium barbadense L.) under saline conditions. J. Anim. Plant Sci. 2016, 26, 487–493. [Google Scholar]
- Rady, M.M.; Mohamed, G.F. Modulation of salt stress effects on the growth, physio-chemical attributes and yields of Phaseolus vulgaris L. plants by the combined application of salicylic acid and Moringa oleifera leaf extract. Sci. Hortic. 2015, 193, 105–113. [Google Scholar] [CrossRef]
- Rady, M.M.; Elrys, A.S.; El-Maati, M.F.A.; Desoky, E.-S.M. Interplaying roles of silicon and proline effectively improve salt and cadmium stress tolerance in Phaseolus vulgaris plant. Plant Physiol. Biochem. 2019, 139, 558–568. [Google Scholar] [CrossRef] [PubMed]
- Rady, M.; Desoky, E.-S.; Elrys, A.; Boghdady, M. Can licorice root extract be used as an effective natural biostimulant for salt-stressed common bean plants? S. Afr. J. Bot. 2019, 121, 294–305. [Google Scholar] [CrossRef]
- Desoky, E.-S.M.; Elrys, A.S.; Rady, M.M. Integrative moringa and licorice extracts application improves Capsicum annuum fruit yield and declines its contaminant contents on a heavy metals-contaminated saline soil. Ecotoxicol. Environ. Saf. 2019, 169, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Desoky, E.-S.M.; ElSayed, A.I.; Merwad, A.-R.M.; Rady, M.M. Stimulating antioxidant defenses, antioxidant gene expression, and salt tolerance in Pisum sativum seedling by pretreatment using licorice root extract (LRE) as an organic biostimulant. Plant. Physiol. Biochem. 2019, 142, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef] [Green Version]
- Desoky, E.-S.M.; El-maghraby, L.M.; Awad, A.E.; Abdo, A.I.; Rady, M.M.; Semida, W.M. Fennel and ammi seed extracts modulate antioxidant defence system and alleviate salinity stress in cowpea (Vigna unguiculata). Sci. Hortic. 2020, 272, 109576. [Google Scholar] [CrossRef]
- Horváth, E.; Szalai, G.; Janda, T. Induction of abiotic stress tolerance by salicylic acid signaling. J. Plant Growth Regul. 2007, 26, 290–300. [Google Scholar] [CrossRef]
- Simaei, M.; Khavari-Nejad, R.; Bernard, F. Exogenous application of salicylic acid and nitric oxide on the ionic contents and enzymatic activities in NaCl-stressed soybean plants. Am. J. Plant Sci. 2012, 3, 1495–1503. [Google Scholar] [CrossRef] [Green Version]
- Metwally, A.; Finkemeier, I.; Georgi, M.; Dietz, K.-J. Salicylic acid alleviates the cadmium toxicity in barley seedlings. Plant Physiol. 2003, 132, 272–281. [Google Scholar] [CrossRef] [Green Version]
- Cheng, K.L.; Bray, R.H. Determination of calcium and magnesium in soil and plant material. Soil Sci. 1951, 72, 449–458. [Google Scholar] [CrossRef]
- Black, C.A. Soil-plant relationships. Soil Sci. 1958, 85, 175. [Google Scholar] [CrossRef]
- Jackson, M.; Gillette, D.; Danielsen, E.; Blifford, I.; Bryson, R.; Syers, J. Global dustfall during the Quaternary as related to environments. Soil Sci. 1973, 116, 135–145. [Google Scholar] [CrossRef]
- Škerget, M.; Kotnik, P.; Hadolin, M.; Hraš, A.R.; Simonič, M.; Knez, Ž. Phenols, proanthocyanidins, flavones and flavonols in some plant materials and their antioxidant activities. Food Chem. 2005, 89, 191–198. [Google Scholar] [CrossRef]
- Horwitz, W.; Chichilo, P.; Reynolds, H. Official Methods of Analysis of the Association of Official Analytical Chemists; Springer: New York, NY, USA, 1970. [Google Scholar] [CrossRef]
- Ordonez, A.; Gomez, J.; Vattuone, M. Antioxidant activities of Sechium edule (Jacq.) Swartz extracts. Food Chem. 2006, 97, 452–458. [Google Scholar] [CrossRef]
- Hatano, T.; Kagawa, H.; Yasuhara, T.; Okuda, T. Two new flavonoids and other constituents in licorice root: Their relative astringency and radical scavenging effects. Chem. Pharm. Bull. 1988, 36, 2090–2097. [Google Scholar] [CrossRef] [Green Version]
- Dastmalchi, K.; Dorman, H.D.; Laakso, I.; Hiltunen, R. Chemical composition and antioxidative activity of Moldavian balm (Dracocephalum moldavica L.) extracts. LWT Food Sci. Technol. 2007, 40, 1655–1663. [Google Scholar] [CrossRef]
- Gülçin, I.; Bursal, E.; Şehitoğlu, M.H.; Bilsel, M.; Gören, A.C. Polyphenol contents and antioxidant activity of lyophilized aqueous extract of propolis from Erzurum, Turkey. Food Chem. Toxicol. 2010, 48, 2227–2238. [Google Scholar] [CrossRef] [PubMed]
- Sati, S.; Bonev, B.; Szabo, Q.; Jost, D.; Bensadoun, P.; Serra, F.; Loubiere, V.; Papadopoulos, G.L.; Rivera-Mulia, J.-C.; Fritsch, L. 4D genome rewiring during oncogene-induced and replicative senescence. Mol. Cell 2020, 78, 522–538.e529. [Google Scholar] [CrossRef] [PubMed]
- Fadeel, A. Location and properties of chloroplasts and pigment determination in roots. Physiol. Plant. 1962, 15, 130–146. [Google Scholar] [CrossRef]
- Osman, A.S.; Rady, M.M. Ameliorative effects of sulphur and humic acid on the growth, anti-oxidant levels, and yields of pea (Pisum sativum L.) plants grown in reclaimed saline soil. J. Hortic. Sci. Biotechnol. 2012, 87, 626–632. [Google Scholar] [CrossRef]
- Rady, M.M. Effect of 24-epibrassinolide on growth, yield, antioxidant system and cadmium content of bean (Phaseolus vulgaris L.) plants under salinity and cadmium stress. Sci. Hortic. 2011, 129, 232–237. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Kubiś, J. Exogenous spermidine differentially alters activities of some scavenging system enzymes, H2O2 and superoxide radical levels in water-stressed cucumber leaves. J. Plant Physiol. 2008, 165, 397–406. [Google Scholar] [CrossRef]
- Mukherjee, S.; Choudhuri, M. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant. 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Irigoyen, J.; Einerich, D.; Sánchez-Díaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativd) plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Konings, E.J.; Roomans, H.H.; Beljaars, P.R. Liquid chromatographic determination of tocopherols and tocotrienols in margarine, infant foods, and vegetables. J. AOAC Int. 1996, 79, 902–906. [Google Scholar] [CrossRef] [Green Version]
- Ching, L.S.; Mohamed, S. Alpha-tocopherol content in 62 edible tropical plants. J. Agric. Food Chem. 2001, 49, 3101–3105. [Google Scholar] [CrossRef]
- Vitória, A.P.; Lea, P.J.; Azevedo, R.A. Antioxidant enzymes responses to cadmium in radish tissues. Phytochemistry 2001, 57, 701–710. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A. Assay of Catalases and Peroxidases; Elsevier: Amsterdam, The Netherlands, 1955. [Google Scholar]
- Thomas, R.L.; Jen, J.J.; Morr, C.V. Changes in soluble and bound peroxidase—IAA oxidase during tomato fruit development. J. Food Sci. 1982, 47, 158–161. [Google Scholar] [CrossRef]
- Fielding, J.L.; Hall, J. A biochemical and cytochemical study of peroxidase activity in roots of Pisum sativum: II. Distribution of enzymes in relation to root development. J. Exp. Bot. 1978, 29, 983–991. [Google Scholar] [CrossRef]
- Sairam, R.K.; Rao, K.V.; Srivastava, G. Differential response of wheat genotypes to long term salinity stress in relation to oxidative stress, antioxidant activity and osmolyte concentration. Plant Sci. 2002, 163, 1037–1046. [Google Scholar] [CrossRef]
- Rao, M.V.; Paliyath, G.; Ormrod, D.P. Ultraviolet-B-and ozone-induced biochemical changes in antioxidant enzymes of Arabidopsis thaliana. Plant Physiol. 1996, 110, 125–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kampfenkel, K.; Vanmontagu, M.; Inzé, D. Extraction and determination of ascorbate and dehydroascorbate from plant tissue. Anal. Biochem. 1995, 225, 165–167. [Google Scholar] [CrossRef]
- Griffith, O.W. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef]
- Dai, A. Increasing drought under global warming in observations and models. Nat. Clim. Chang. 2013, 3, 52–58. [Google Scholar] [CrossRef]
- Ahmadi, S.H.; Andersen, M.N.; Plauborg, F.; Poulsen, R.T.; Jensen, C.R.; Sepaskhah, A.R.; Hansen, S. Effects of irrigation strategies and soils on field grown potatoes: Yield and water productivity. Agric. Water Manag. 2010, 97, 1923–1930. [Google Scholar] [CrossRef]
- Obidiegwu, J.E.; Bryan, G.J.; Jones, H.G.; Eprashar, A. Coping with drought: Stress and adaptive responses in potato and perspectives for improvement. Front. Plant Sci. 2015, 6, 542. [Google Scholar] [CrossRef] [Green Version]
- Nasir, M.W.; Toth, Z. Response of Different Potato Genotypes to Drought Stress. Agriculture 2021, 11, 763. [Google Scholar] [CrossRef]
- Earl, H.J.; Davis, R.F. Effect of drought stress on leaf and whole canopy radiation use efficiency and yield of maize. Agron. J. 2003, 95, 688–696. [Google Scholar] [CrossRef]
- Anjum, S.A.; Xie, X.-y.; Wang, L.-c.; Saleem, M.F.; Man, C.; Lei, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar]
- Kusvuran, S.; Kiran, S.; Ellialtioglu, S.S. Antioxidant enzyme activities and abiotic stress tolerance relationship in vegetable crops. In Abiotic and Biotic Stress in Plants-Recent Advances and Future Perspectives [Internet], 1st ed.; InTech: Croatia, Yugoslavia, 2016; pp. 481–503. [Google Scholar] [CrossRef] [Green Version]
- Rady, M.M.; Boriek, S.H.; El-Mageed, A.; Taia, A.; Seif El-Yazal, M.A.; Ali, E.F.; Hassan, F.A.; Abdelkhalik, A. Exogenous gibberellic acid or dilute bee honey boosts drought stress tolerance in Vicia faba by rebalancing osmoprotectants, antioxidants, nutrients, and phytohormones. Plants 2021, 10, 748. [Google Scholar] [CrossRef]
- Semida, W.M.; Abdelkhalik, A.; Mohamed, G.; El-Mageed, A.; Taia, A.; El-Mageed, A.; Shimaa, A.; Rady, M.M.; Ali, E.F. Foliar application of zinc oxide nanoparticles promotes drought stress tolerance in eggplant (Solanum melongena L.). Plants 2021, 10, 421. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, D.; Lin, P.; Guo, S. Effects of salinity on leaf characteristics and CO2/H2O exchange of Kandelia candel (L.) Druce seedlings. J. Sci. 2007, 53, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Foidl, N.; Makkar, H.; Becker, K. The potential of Moringa oleifera for agricultural and industrial uses. What Dev. Potential Moringa Prod. 2001, 20. Available online: http://moringatrees.org/moringa-doc/the_potential_of_moringa_oleifera_for_agricultural_and_industrial_uses.pdf (accessed on 1 October 2021).
- Afzal, I.; Hussain, B.; Basra, S.; Rehman, H. Priming with moringa leaf extract reduces imbibitional chilling injury in spring maize. Seed Sci. Technol. 2012, 40, 271–276. [Google Scholar] [CrossRef]
- Naikoo, M.I.; Dar, M.I.; Raghib, F.; Jaleel, H.; Ahmad, B.; Raina, A.; Khan, F.A.; Naushin, F. Role and regulation of plants phenolics in abiotic stress tolerance: An overview. Plant Signal. Mol. 2019, 157–168. [Google Scholar]
- Tanase, C.; Bujor, O.-C.; Popa, V.I. Phenolic natural compounds and their influence on physiological processes in plants. In Polyphenols in Plants; Elsevier: Amsterdam, The Netherlands, 2019; pp. 45–58. [Google Scholar]
- Bujor, O.-C.; Talmaciu, I.A.; Volf, I.; Popa, V.I. Biorefining to recover aromatic compounds with biological properties. TAPPI J 2015, 14, 187–193. [Google Scholar] [CrossRef]
- Cheynier, V.; Comte, G.; Davies, K.M.; Lattanzio, V.; Martens, S. Plant phenolics: Recent advances on their biosynthesis, genetics, and ecophysiology. Plant Physiol. Biochem. 2013, 72, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.P. Source–sink balance and carbon allocation below ground in plants exposed to ozone. N. Phytol. 2003, 157, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Ballizany, W.L.; Hofmann, R.W.; Jahufer, M.Z.; Barrett, B.A. Multivariate associations of flavonoid and biomass accumulation in white clover (Trifolium repens) under drought. Funct. Plant Biol. 2012, 39, 167–177. [Google Scholar] [CrossRef]
- Rezayian, M.; Niknam, V.; Ebrahimzadeh, H. Differential responses of phenolic compounds of Brassica napus under drought stress. Iran J. Plant Physiol. 2018, 8, 2417–2425. [Google Scholar]
- Nichols, S.N.; Hofmann, R.W.; Williams, W.M. Physiological drought resistance and accumulation of leaf phenolics in white clover interspecific hybrids. Environ. Exp. Bot. 2015, 119, 40–47. [Google Scholar] [CrossRef]
- Hernández, I.; Alegre, L.; Van Breusegem, F.; Munné-Bosch, S. How relevant are flavonoids as antioxidants in plants? Trends Plant Sci. 2009, 14, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Ozfidan-Konakci, C.; Yildiztugay, E.; Kucukoduk, M. Upregulation of antioxidant enzymes by exogenous gallic acid contributes to the amelioration in Oryza sativa roots exposed to salt and osmotic stress. Environ. Sci. Pollut. Res. 2015, 22, 1487–1498. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.I.; Gonzalez, L.; Reigosa, M.J. Allelopathic potential of Acacia melanoxylon on the germination and root growth of native species. Weed Biol. Manag. 2011, 11, 18–28. [Google Scholar] [CrossRef]
- Sgherri, C.; Cosi, E.; Navari-Izzo, F. Phenols and antioxidative status of Raphanus sativus grown in copper excess. Physiol. Plant. 2003, 118, 21–28. [Google Scholar] [CrossRef]
- Beckman, C.H. Phenolic-storing cells: Keys to programmed cell death and periderm formation in wilt disease resistance and in general defence responses in plants? Physiol. Mol. Plant Pathol. 2000, 57, 101–110. [Google Scholar] [CrossRef]
- Fletcher, R.; Kallidumbil, V.; Steele, P. An improved bioassay for cytokinins using cucumber cotyledons. Plant Physiol. 1982, 69, 675–677. [Google Scholar] [CrossRef] [Green Version]
- Yildirim, E.; Turan, M.; Guvenc, I. Effect of foliar salicylic acid applications on growth, chlorophyll, and mineral content of cucumber grown under salt stress. J. Plant Nutr. 2008, 31, 593–612. [Google Scholar] [CrossRef]
- Aftab, T.; Masroor, M.; Khan, A.; Idrees, M.; Naeem, M. Salicylic acid acts as potent enhancer of growth, photosynthesis and artemisinin production in Artemisia annua L. J. Crop Sci. Biotechnol. 2010, 13, 183–188. [Google Scholar] [CrossRef]
- Yildirim, E.; Karlidag, H.; Turan, M. Mitigation of salt stress in strawberry by foliar K, Ca and Mg nutrient supply. Plant Soil Env. 2009, 55, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Erner, Y.; Artzi, B.; Tagari, E.; Homou, M. The use of potassium nitrate as alternative for urea foliar spray in citrus. Alon. Hanotea 2002, 56, 138–142. [Google Scholar]
- Martinez, V.; Cerda, A. Influence of N source on rate of Cl, N, Na and K uptake by cucumber seedlings grown in saline condition. J. Plant Nutr. 1989, 12, 971–983. [Google Scholar] [CrossRef]
- El-Nour, A. Growth and nutrient contents response of maize to foliar nutrition with micronutrients under irrigation with saline water. OnLine J. Biol. Sci. 2002, 2, 92–97. [Google Scholar]
- Fariduddin, Q.; Hayat, S.; Ahmad, A. Salicylic acid influences net photosynthetic rate, carboxylation efficiency, nitrate reductase activity, and seed yield in Brassica juncea. Photosynthetica 2003, 41, 281–284. [Google Scholar] [CrossRef]
- Szepesi, Á. Role of salicylic acid pre-treatment on the acclimation of tomato plants to salt-and osmotic stress. Acta Biol. Szeged. 2005, 49, 123–125. [Google Scholar]
- Gunes, A.; Inal, A.; Alpaslan, M.; Eraslan, F.; Bagci, E.G.; Cicek, N. Salicylic acid induced changes on some physiological parameters symptomatic for oxidative stress and mineral nutrition in maize (Zea mays L.) grown under salinity. J. Plant Physiol. 2007, 164, 728–736. [Google Scholar] [CrossRef] [PubMed]
- Thomas, H.; Howarth, C.J. Five ways to stay green. J. Exp. Bot. 2000, 51, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Duursma, R.A.; Medlyn, B.E.; Kelly, J.W.; Prentice, I.C. How should we model plant responses to drought? An analysis of stomatal and non-stomatal responses to water stress. Agric. For. Meteorol. 2013, 182, 204–214. [Google Scholar] [CrossRef]
- Murata, N.; Takahashi, S.; Nishiyama, Y.; Allakhverdiev, S.I. Photoinhibition of photosystem II under environmental stress. Biochim. Biophys. Acta Bioenerg. 2007, 1767, 414–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, S.; Murata, N. How do environmental stresses accelerate photoinhibition? Trends Plant Sci. 2008, 13, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Cornic, G.; Fresneau, C. Photosynthetic carbon reduction and carbon oxidation cycles are the main electron sinks for photosystem II activity during a mild drought. Ann. Bot. 2002, 89, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Nabe, H.; Funabiki, R.; Kashino, Y.; Koike, H.; Satoh, K. Responses to desiccation stress in bryophytes and an important role of dithiothreitol-insensitive non-photochemical quenching against photoinhibition in dehydrated states. Plant Cell Physiol. 2007, 48, 1548–1557. [Google Scholar] [CrossRef] [Green Version]
- Ur Rehman, H.; Alharby, H.F.; Alzahrani, Y.; Rady, M.M. Magnesium and organic biostimulant integrative application induces physiological and biochemical changes in sunflower plants and its harvested progeny on sandy soil. Plant Physiol. Biochem. 2018, 126, 97–105. [Google Scholar] [CrossRef]
- Zayed, B.; Salem, A.; El Sharkawy, H. Effect of different micronutrient treatments on rice (Oriza sativa L.) growth and yield under saline soil conditions. World J. Agric. Sci. 2011, 7, 179–184. [Google Scholar]
- Dubey, R.S. Photosynthesis in plants under stressful conditions. In Handbook of Photosynthesis; CRC Press: Boca Raton, FL, USA, 2018; pp. 629–649. [Google Scholar]
- Marschner, H. Mineral Nutrition of Higher Plants; Academic Press: San Diego, CA, USA, 1995. [Google Scholar]
- Shabala, S.; Schimanski, L.J.; Koutoulis, A. Heterogeneity in bean leaf mesophyll tissue and ion flux profiles: Leaf electrophysiological characteristics correlate with the anatomical structure. Ann. Bot. 2002, 89, 221–226. [Google Scholar] [CrossRef] [Green Version]
- Brugnoli, E.; Björkman, O. Chloroplast movements in leaves: Influence on chlorophyll fluorescence and measurements of light-induced absorbance changes related to ΔpH and zeaxanthin formation. Photosynth. Res. 1992, 32, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Rady, M.M.; Hemida, K.A. Sequenced application of ascorbate-proline-glutathione improves salt tolerance in maize seedlings. Ecotoxicol. Environ. Saf. 2016, 133, 252–259. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Huang, B. Drought and heat stress injury to two cool-season turfgrasses in relation to antioxidant metabolism and lipid peroxidation. Crop Sci. 2001, 41, 436–442. [Google Scholar] [CrossRef]
- Yadavi, A.; Aboueshaghi, R.; Dehnavi, M.; Balouchi, H. Effect of micronutrients foliar application on grain qualitative characteristics and some physiological traits of bean (Phaseolus vulgaris L.) under drought stress. Indian J. Fundam. Appl. Life Sci. 2014, 4, 124–131. [Google Scholar]
- Stevens, J.; Senaratna, T.; Sivasithamparam, K. Salicylic acid induces salinity tolerance in tomato (Lycopersicon esculentum cv. Roma): Associated changes in gas exchange, water relations and membrane stabilisation. Plant Growth Regul. 2006, 49, 77–83. [Google Scholar]
- González, L.; González-Vilar, M. Determination of relative water content. In Handbook of Plant Ecophysiology Techniques; Springer: Berlin/Heidelberg, Germany, 2001; pp. 207–212. [Google Scholar]
- Stępień, P.; Kłbus, G. Water relations and photosynthesis in Cucumis sativus L. leaves under salt stress. Biol. Plant. 2006, 50, 610–616. [Google Scholar] [CrossRef]
- Prisco, J. Alguns aspectos da fisiologia do “stress” salino. Rev. Bras. De Botânica 1980, 3, 85–94. [Google Scholar]
- Trapp, S.; Feificova, D.; Rasmussen, N.F.; Bauer-Gottwein, P. Plant uptake of NaCl in relation to enzyme kinetics and toxic effects. Environ. Exp. Bot. 2008, 64, 1–7. [Google Scholar] [CrossRef]
- Tari, I. Acclimation of tomato plants to salinity stress after a salicylic acid pre-treatment. Acta Biol. Szeged. 2002, 46, 55–56. [Google Scholar]
- Sakhabutdinova, A.; Fatkhutdinova, D.; Bezrukova, M.; Shakirova, F. Salicylic acid prevents the damaging action of stress factors on wheat plants. Bulg. J. Plant Physiol. 2003, 21, 314–319. [Google Scholar]
- Allen, D.; McKee, I.; Farage, P.; Baker, N. Analysis of limitations to CO2 assimilation on exposure of leaves of two Brassica napus cultivars to UV-B. Plant Cell Environ. 1997, 20, 633–640. [Google Scholar] [CrossRef]
- Hernández, J.A.; Almansa, M.S. Short-term effects of salt stress on antioxidant systems and leaf water relations of pea leaves. Physiol. Plant. 2002, 115, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Ruppenthal, V.; Zoz, T.; Steiner, F.; do Carmo, L.M.; Castagnara, D.D. Silicon does not alleviate the adverse effects of drought stress in soybean plants. Semin. Ciências Agrárias 2016, 37, 3941–3954. [Google Scholar] [CrossRef] [Green Version]
- Moharekar, S.; Lokhande, S.; Hara, T.; Tanaka, R.; Tanaka, A.; Chavan, P. Effect of salicylic acid on chlorophyll and carotenoid contents of wheat and moong seedlings. Photosynthetica 2003, 41, 315–317. [Google Scholar] [CrossRef]
- Hsu, S.-Y.; Kao, C.H. Differential effect of sorbitol and polyethylene glycol on antioxidant enzymes in rice leaves. Plant Growth Regul. 2003, 39, 83–90. [Google Scholar] [CrossRef]
- Serrano, R.; Mulet, J.M.; Rios, G.; Marquez, J.A.; De Larrinoa, I.F.; Leube, M.P.; Mendizabal, I.; Pascual-Ahuir, A.; Proft, M.; Ros, R. A glimpse of the mechanisms of ion homeostasis during salt stress. J. Exp. Bot. 1999, 1023–1036. [Google Scholar] [CrossRef]
- Michalak, A. Phenolic compounds and their antioxidant activity in plants growing under heavy metal stress. Pol. J. Environ. Stud. 2006, 15. [Google Scholar]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef]
- Duh, P.-D. Antioxidant activity of water extract of four Harng Jyur (Chrysanthemum morifolium Ramat) varieties in soybean oil emulsion. Food Chem. 1999, 66, 471–476. [Google Scholar] [CrossRef]
- Ghaderi, N.; Normohammadi, S.; Javadi, T. Morpho-physiological responses of strawberry (Fragaria × ananassa) to exogenous salicylic acid application under drought stress. J. Agric. Sci. Technol. 2015, 17, 167–178. [Google Scholar]
- Zhu, J.-K. Plant salt tolerance. Trends Plant Sci. 2001, 6, 66–71. [Google Scholar] [CrossRef]
- Howladar, S.M. A novel Moringa oleifera leaf extract can mitigate the stress effects of salinity and cadmium in bean (Phaseolus vulgaris L.) plants. Ecotoxicol. Environ. Saf. 2014, 100, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Farhangi-Abriz, S.; Torabian, S. Antioxidant enzyme and osmotic adjustment changes in bean seedlings as affected by biochar under salt stress. Ecotoxicol. Environ. Saf. 2017, 137, 64–70. [Google Scholar] [CrossRef]
- Taha, R. Improving salt tolerance of Helianthus annuus (L.) plants by Moringa oleifera leaf extract. Egypt J. Agron. 2016, 38, 117–140. [Google Scholar]
- Bano, S.; Ashraf, M.; Akram, N.A. Salt stress regulates enzymatic and nonenzymatic antioxidative defense system in the edible part of carrot (Daucus carota L.). J. Plant Interact. 2014, 9, 324–329. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [Green Version]
- Semida, W.M.; Rady, M.M. Presoaking application of propolis and maize grain extracts alleviates salinity stress in common bean (Phaseolus vulgaris L.). Sci. Hortic. 2014, 168, 210–217. [Google Scholar] [CrossRef]
- Zhou, Y.; Wen, Z.; Zhang, J.; Chen, X.; Cui, J.; Xu, W.; Liu, H.-y. Exogenous glutathione alleviates salt-induced oxidative stress in tomato seedlings by regulating glutathione metabolism, redox status, and the antioxidant system. Sci. Hortic. 2017, 220, 90–101. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Anee, T.I.; Fujita, M. Exogenous silicon attenuates cadmium-induced oxidative stress in Brassica napus L. by modulating AsA-GSH pathway and glyoxalase system. Front. Plant Sci. 2017, 8, 1061. [Google Scholar] [CrossRef] [PubMed]
- Salem, N.; Msaada, K.; Dhifi, W.; Limam, F.; Marzouk, B. Effect of salinity on plant growth and biological activities of Carthamus tinctorius L. extracts at two flowering stages. Acta Physiol. Plant. 2014, 36, 433–445. [Google Scholar] [CrossRef]
- Xu, Q.; Xu, X.; Zhao, Y.; Jiao, K.; Herbert, S.J.; Hao, L. Salicylic acid, hydrogen peroxide and calcium-induced saline tolerance associated with endogenous hydrogen peroxide homeostasis in naked oat seedlings. Plant Growth Regul. 2008, 54, 249–259. [Google Scholar] [CrossRef]
- Abogadallah, G.M. Insights into the significance of antioxidative defense under salt stress. Plant Signal. Behav. 2010, 5, 369–374. [Google Scholar] [CrossRef]
- Ma, H.; Zhu, Q.; Zhao, W. Soil water response to precipitation in different micro-topographies on the semi-arid Loess Plateau, China. J. For. Res. 2020, 31, 245–256. [Google Scholar] [CrossRef]
- Chen, Z.; Klessig, D.F. Identification of a soluble salicylic acid-binding protein that may function in signal transduction in the plant disease-resistance response. Proc. Natl. Acad. Sci. USA 1991, 88, 8179–8183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saruhan, N.; Saglam, A.; Kadioglu, A. Salicylic acid pretreatment induces drought tolerance and delays leaf rolling by inducing antioxidant systems in maize genotypes. Acta Physiol. Plant. 2012, 34, 97–106. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Soil Characteristics | Unit | Values | |
|---|---|---|---|
| 2020 Season | 2021 Season | ||
| Sand | % | 90.50 ± 1.5 | 90.4 ± 1.3 |
| Silt | 4.78 ± 0.1 | 4.83 ± 0.13 | |
| Clay | 3.72 ± 0.12 | 3.97 ± 0.16 | |
| Texture class | Sandy | ||
| Organic matter | % | 4.45 ± 0.05 | 4.57 ± 0.05 |
| pH (in 1: 1 of soil: water suspension) | 7.97 ± 0.04 | 7.99 ± 0.04 | |
| EC (in soil paste extract) | dS m−1 | 1.69 ± 0.01 | 1.72 ±0.01 |
| N-NH4+ | mg kg−1 soil | 14.8 ± 0.5 | 15.01 ± 0.4 |
| N-NO3− | 6.65 ± 0.1 | 6.71 ± 0.08 | |
| Total N | 23.7 ± 0.05 | 27.5 ± 0.04 | |
| Total P | 6.81 ± 0.1 | 6.37 ± 0.05 | |
| Total K | 59.3 ± 0.3 | 56.6 ± 0.4 | |
| Component | Unit | Value |
|---|---|---|
| Total phenolic compounds (TPC) | mg GAE g−1 CFE | 323.79 |
| Total flavonoids (TF) | mg QE g−1 CFE | 34.65 |
| Phenolic compounds | ||
| 3,4-Dihydroxybenzoic-acid | mg g−1 CFE | 0.74 |
| Ellagic-acid | 0.62 | |
| Eugenol | 104.7 | |
| Eugenyl-acetate | 86.39 | |
| Gallic acid | 18.33 | |
| Naphthalene | 0.21 | |
| Tannic acid | 0.78 | |
| Vanillin | 1.49 | |
| Antioxidants and osmoprotectants | ||
| Total free amino acid | g Kg−1 DW | 70.2 |
| Free Proline | 0.19 | |
| Soluble sugars | 0.56 | |
| Mineral nutrients | ||
| Mg | g Kg−1 DW | 3.2 |
| Ca | 12.3 | |
| Fe | 1.3 | |
| P | 11.8 | |
| K | 16.5 | |
| N | 16.9 | |
| Vitamins | ||
| Vitamin A | mg Kg−1 DW | 25.6 |
| Vitamin E | 55.2 | |
| Vitamin D | 32.4 | |
| Vitamin C | 36.9 | |
| Treatments | Shoot Fresh Weight (g) | Shoot Dry Weight (g) | Leaf Area Plant−1 (cm2) | Tuber Number Plant−1 | Average Tuber Weight (g) | Total Yield (Ton Fad−1) | |
|---|---|---|---|---|---|---|---|
| Stress | Foliar Spray | ||||||
| 2020 Season | |||||||
| Well-watered | Control | 311.9 ± 3.2 d | 33.5 ± 1.1 d | 4158 ± 4.5 d | 3.79 ± 0.09 c | 191.3 ± 3.2 d | 20.3 ± 1.1 d |
| SA | 322.2 ± 3.6 c | 34.6 ± 1.3 c | 4277 ± 4.6 c | 3.89 ± 0.07 b | 196.3 ± 2.5 c | 21.1 ± 1.2 c | |
| CFE | 327.1 ± 3.8 b | 35.1 ± 1.2 b | 4425 ± 4.8 b | 3.97 ± 0.06 ab | 203.1 ± 2.8 b | 21.3 ± 1.3 b | |
| CFE+SA | 331.7 ± 3.2 a | 35.6 ± 1.5 a | 4510 ± 4.4 a | 4.02 ± 0.11 a | 210.4 ± 2.7 a | 21.7 ± 1.4 a | |
| Moderate drought | Control | 243.5 ± 3.6 h | 26.1 ± 0.9 h | 3577 ± 3.9 h | 3.17 ± 0.07 g | 171.6 ± 2.6 h | 17.5 ± 1.2 h |
| SA | 262.6 ±3.8 g | 28.2 ± 0.8 g | 3709 ± 4.2 g | 3.36 ± 0.08 f | 174.2 ± 1.9 g | 18.2 ± 1.4 g | |
| CFE | 280.3 ± 4.1 f | 30.1 ± 1.1 f | 3713 ± 3.7 f | 3.47 ± 0.08 e | 177.5 ± 2.2 f | 19.1 ± 1.0 f | |
| CFE+SA | 292.2 ± 3.9 e | 31.4 ± 1.3 e | 3936 ± 3.8 e | 3.64 ± 0.06 d | 181.1 ± 1.8 e | 19.5 ± 1.0 e | |
| Severe drought | Control | 152.6 ± 1.5 l | 16.4 ± 1.4 l | 2048 ± 3.5 l | 2.60 ± 0.07 k | 154.8 ± 1.6 l | 13.3 ± 0.9 l |
| SA | 178.5 ± 1.5 k | 19.1 ± 0.9 k | 2185 ± 4.2 k | 2.73 ± 0.08 j | 159.9 ± 2.2 k | 14.3 ± 0.8 k | |
| CFE | 189.6 ± 1.8 j | 20.3 ± 0.8 j | 2371 ± 4.9 j | 2.85 ± 0.06 i | 162.1 ± 2.5 j | 15.1 ± 0.9 j | |
| CFE+SA | 208.9 ± 2.3 i | 22.4 ± 1.2 i | 2549 ± 3.8 i | 2.97 ± 0.05 h | 165.2 ± 2.3 i | 16.1 ± 0.8 i | |
| 2021 season | |||||||
| Well-watered | Control | 309.3 ± 4.2 d | 32.7 ± 1.4 c | 4155 ± 5.3 d | 3.75 ± 0.07 bc | 190.4 ± 3.4 d | 20.3 ± 1.0 c |
| SA | 319.4 ± 4.5 c | 33.7 ± 1.7 b | 4274 ± 5.6 c | 3.84 ± 0.08 ab | 192.3 ± 3.5 c | 20.9 ± 1.0 b | |
| CFE | 324.4 ± 3.8 b | 34.4 ± 1.5 a | 4423 ± 5.4 b | 3.93 ± 0.09 a | 202.7 ± 3.9 b | 21.2 ± 0.9 b | |
| CFE+SA | 329.1 ± 3.5 a | 34.7 ± 1.3 a | 4507 ± 4.8 a | 3.96 ± 0.06 a | 209.6 ± 2.8 a | 21.7 ± 0.9 a | |
| Moderate drought | Control | 241.2 ± 3.8 h | 25.4 ± 1.2 g | 3574 ± 4.9 h | 3.15 ± 0.07 e | 171.0 ± 2.7 h | 17.3 ± 0.9 g |
| SA | 260.3 ± 4.2 g | 27.5 ± 1.3 f | 3706 ± 5.6 g | 3.43 ± 0.05 d | 173.5 ± 3.5 g | 18.1 ± 0.8 f | |
| CFE | 278.0 ± 2.9 f | 29.7 ± 1.4 e | 3811 ± 4.7 f | 3.45 ± 0.07 d | 176.9 ± 1.6 f | 18.9 ± 0.8 e | |
| CFE+SA | 289.7 ± 2.3 e | 30.9 ± 1.1 d | 3935 ± 3.9 e | 3.61 ± 0.08 c | 180.4 ± 1.8 e | 19.3 ± 0.9 d | |
| Severe drought | Control | 150.7 ± 2.7 l | 16.1 ± 1.3 k | 2045 ± 4.3 l | 2.57 ± 0.09 h | 154.3 ± 2.5 l | 13.2 ± 0.8 k |
| SA | 176.6 ± 3.6 k | 18.7 ± 0.9 j | 2183 ± 3.6 k | 2.70 ± 0.06g h | 159.3 ± 2.7 k | 14.2 ± 0.9 j | |
| CFE | 187.6 ± 3.4 j | 19.6 ± 0.9 i | 2369 ± 5.2 j | 2.82 ± 0.07 fg | 161.5 ± 1.9 j | 14.9 ± 0.7 i | |
| CFE+SA | 207.0 ± 1.9 i | 22.1 ± 1.0 h | 2546 ± 5.3 i | 2.95 ± 0.06 f | 164.5 ± 1.8 i | 16.1 ± 0.8 h | |
| Treatments | TChC (mg g−1 FW) | TCarC (mg g−1 FW) | Pn (µmol CO2 m−2 s−1) | Tr (mmol H2O m−2 s−1) | gs (mmol H2O m−2 s−1) | |
|---|---|---|---|---|---|---|
| Stress | Foliar Spray | |||||
| 2020 Season | ||||||
| Well-watered | Control | 2.09 ± 0.09 bc | 0.74 ± 0.03 d | 12.4 ± 1.1 d | 6.34 ± 0.44 c | 0.466 ± 0.02 d |
| SA | 2.12 ± 0.11 b | 0.76 ± 0.04 c | 12.6 ± 1.3 c | 6.51 ± 0.42 bc | 0.480 ± 0.03 c | |
| CFE | 2.17 ± 0.12 ab | 0.79 ± 0.04 b | 12.8 ± 1.4 b | 6.72 ± 0.45 b | 0.493 ± 0.02 b | |
| CFE+SA | 2.22 ± 0.10 a | 0.84 ± 0.05 a | 13.1 ± 1.5 a | 7.17 ± 0.52 a | 0.513 ± 0.04 a | |
| Moderate drought | Control | 1.65 ± 0.07 f | 0.70 ± 0.03 e | 8.33 ± 1.96 h | 4.40 ± 0.41 fg | 0.386 ± 0.01 h |
| SA | 1.77 ± 0.08 e | 0.71 ± 0.04 e | 8.84 ± 0.085 g | 4.54 ± 0.46 f | 0.413 ± 0.02 g | |
| CFE | 1.93 ± 0.07 d | 0.73 ± 0.03 d | 9.78 ± 0.94 f | 4.83 ± 0.38 e | 0.430 ± 0.02 f | |
| CFE+SA | 2.02 ± 0.10 cd | 0.74 ± 0.03 d | 10.3 ± 1.1 e | 5.17 ± 0.37 d | 0.453 ± 0.03 e | |
| Severe drought | Control | 1.33 ± 0.06 i | 0.60 ± 0.011 | 6.18 ± 0.74 l | 3.38 ± 0.33 j | 0.260 ± 0.01 l |
| SA | 1.40 ± 0.08 h | 0.63 ± 0.02 h | 6.55 ± 0.73 k | 3.67 ± 0.21 i | 0.286 ± 0.01 k | |
| CFE | 1.44 ± 0.07 h | 0.65 ± 0.01 g | 6.79 ± 0.62 j | 4.07 ± 0.12 h | 0.313 ± 0.02 j | |
| CFE+SA | 1.54 ± 0.08 g | 0.67 ± 0.02 f | 7.10 ± 0.53 i | 4.19 ± 0.32 gh | 0.326 ± 0.02 i | |
| 2021 season | ||||||
| Well-watered | Control | 2.07 ± 0.12 bc | 0.73 ± 0.05 cd | 11.9 ± 0.95 b | 6.06 ± 0.39 c | 0.463 ± 0.03 b |
| SA | 2.11 ± 0.10 b | 0.75 ± 0.06 c | 12.1 ± 1.3 ab | 6.32 ± 0.38 bc | 0.466 ± 0.03 b | |
| CFE | 2.16 ± 0.11 ab | 0.78 ± 0.07 b | 12.2 ± 1.4 ab | 6.50 ± 0.28 ab | 0.476 ± 0.02 ab | |
| CFE+SA | 2.21 ± 0.13 a | 0.83 ± 0.06 a | 12.4 ± 1.5 a | 6.88 ± 0.15 a | 0.4933 ± 0.03 a | |
| Moderate drought | Control | 1.55 ± 0.06 f | 0.67 ± 0.03 f | 7.92 ± 0.66 f | 4.14 ± 0.34 f | 0.373 ± 0.01 e |
| SA | 1.76 ± 0.07 e | 0.70 ± 0.05 e | 8.29 ± 0.74 e | 4.28 ± 0.32 f | 0.400 ± 0.01 d | |
| CFE | 1.83 ± 0.08 d | 0.70 ± 0.03 e | 9.22 ± 0.85 d | 4.67 ± 0.36 e | 0.420 ± 0.02 cd | |
| CFE+SA | 2.00 ± 0.13 cd | 0.72 ± 0.04 de | 9.89 ± 0.89 c | 5.13 ± 0.37 d | 0.436 ± 0.03 c | |
| Severe drought | Control | 1.22 ± 0.05 i | 0.56 ± 0.01 h | 6.14 ± 0.42 i | 3.27 ± 0.19 g | 0.243 ± 0.01 h |
| SA | 1.29 ± 0.03 h | 0.60 ± 0.02 h | 6.40 ± 0.43 hi | 3.56 ± 0.12 g | 0.276 ± 0.01 g | |
| CFE | 1.33 ± 0.04 h | 0.63 ± 0.03 g | 6.53 ± 0.33 h | 3.94 ± 0.13 f | 0.300 ± 0.01 f | |
| CFE+SA | 1.43 ± 0.06 g | 0.59 ± 0.01 f | 6.90 ± 0.36 g | 4.05 ± 0.18 f | 0.316 ± 0.02 f | |
| Treatments | RWC (%) | MSI (%) | EL (%) | MDA (µmol g−1 FW) | O2•− (A580 g−1 FW) | H2O2 (µmol g−1 FW) | |
|---|---|---|---|---|---|---|---|
| Stress | Foliar Spray | ||||||
| 2020 season | |||||||
| Well- watered | Control | 85.7 ± 2.3 d | 71.5 ± 2.2 c | 7.07 ± 0.11 h | 13.0 ± 0.69 i | 0.37 ± 0.01 i | 5.53 ± 0.21 i |
| SA | 87.3 ± 3.6 c | 72.6 ± 2.5 b | 6.86 ± 0.14 h | 12.6 ± 0.88 i | 0.36 ± 0.01 j | 5.41 ± 0.26 i | |
| CFE | 89.1 ± 3.5 b | 74.5 ± 2.6 a | 6.55 ± 0.19 j | 12.5 ± 0.89 i | 0.34 ± 0.02 k | 5.21 ± 0.24 j | |
| CFE+SA | 90.4 ± 2.8 a | 75.4 ± 2.3 a | 6.11 ± 0.18 j | 12.4 ± 0.79 i | 0.32 ± 0.01 l | 5.05 ± 0.22 j | |
| Moderate drought | Control | 80.6 ± 3.6 h | 65.7 ± 2.1 g | 11.4 ± 0.32 d | 20.5 ± 1.2 e | 0.58 ± 0.03 e | 10.3 ± 0.56 e |
| SA | 81.7 ± 3.9 g | 67.0 ± 2.8 f | 10.6 ± 0.35 e | 18.6 ± 1.5 f | 0.55 ± 0.04 f | 9.54 ± 0.49 f | |
| CFE | 82.8 ± 3.4 f | 68.5 ± 2.9 e | 9.92 ± 0.25 f | 17.1 ± 1.4 g | 0.52 ± 0.03 g | 8.96 ± 0.39 g | |
| CFE+SA | 84.8 ± 2.9 e | 69.5 ± 3.1 d | 9.42 ± 0.24 g | 15.2 ± 1.1 h | 0.49 ± 0.02 h | 8.21 ± 0.48 h | |
| Severe drought | Control | 75.6 ± 2.5 l | 42.2 ± 2.4 k | 14.6 ± 0.29 a | 36.6 ± 2.3 a | 0.71 ± 0.04 a | 16.1 ± 0.78 a |
| SA | 76.9 ± 2.6 k | 48.8 ± 1.2 j | 13.3 ± 0.28 b | 33.5 ± 2.4 b | 0.67 ± 0.03 b | 13.9 ± 0.88 b | |
| CFE | 77.9 ± 3.1 j | 51.2 ± 1.7 i | 13.1 ± 0.32 b | 31.6 ± 2.4 c | 0.64 ± 0.04 c | 12.7 ± 0.79 c | |
| CFE+SA | 79.3 ± 3.3 i | 54.8 ± 1.6 h | 12.1 ± 0.41 c | 30.3 ± 1.5 d | 0.62 ± 0.05 d | 12.4 ± 0.95 d | |
| 2021 season | |||||||
| Well- watered | Control | 85.1 ± 4.1 b | 70.8 ± 1.9 d | 7.09 ± 0.32 h | 13.7 ± 0.75 i | 0.38 ± 0.02 i | 5.68 ± 0.32 i |
| SA | 85.3 ± 4.5 b | 71.9 ± 2.5 c | 6.74 ± 0.36 hi | 12.7 ± 0.74 i | 0.37 ± 0.01 i | 5.60 ± 0.33 ij | |
| CFE | 86.6 ± 4.3 ab | 73.5 ± 2.9 b | 6.40 ± 0.25 ij | 12.6 ± 0.86 i | 0.35 ± 0.01 j | 5.27 ± 0.34 jk | |
| CFE+SA | 88.1 ± 4.8 a | 74.8 ± 3.1 a | 6.04 ± 0.29 j | 12.6 ± 1.2 i | 0.33 ± 0.01 k | 5.16 ± 0.52 k | |
| Moderate drought | Control | 79.8 ± 3.5 d | 65.2 ± 3.2 f | 11.7 ± 0.45 d | 21.4 ± 1.3 e | 0.60 ± 0.03 e | 10.5 ± 0.68 e |
| SA | 81.1 ± 3.2 cd | 66.0 ± 1.9 f | 10.8 ± 0.44 e | 18.7 ± 1.5 f | 0.55 ± 0.03 f | 9.69 ± 0.78 f | |
| CFE | 81.9 ± 3.9 c | 67.3 ± 2.8 e | 10.2 ± 0.52 f | 17.4 ± 1.4 g | 0.53 ± 0.04 g | 9.25 ± 0.68 g | |
| CFE+SA | 83.9 ± 3.3 b | 68.4 ± 2.4 e | 9.57 ± 0.56 g | 15.5 ± 1.6 h | 0.50 ± 0.03 h | 8.47 ± 0.65 h | |
| Severe drought | Control | 74.4 ± 2.5 g | 41.1 ± 1.3 j | 15.1 ± 0.66 a | 37.6 ± 2.3 a | 0.72 ± 0.04 a | 16.7 ± 0.79 a |
| SA | 75.9 ± 2.8 fg | 47.6 ± 1.4 i | 13.5 ± 0.69 b | 34.2 ± 3.2 b | 0.68 ± 0.03 b | 14.2 ± 0.99 b | |
| CFE | 76.7 ± 3.5 ef | 50.5 ± 1.5 h | 13.0 ± 0.66 c | 32.1 ± 3.1 c | 0.66 ± 0.02 c | 13.1 ± 0.96 c | |
| CFE+SA | 78.2 ± 2.8 de | 53.9 ± 1.6 g | 12.3 ± 0.68 d | 30.7 ± 3.3 d | 0.63 ± 0.01 d | 12.5 ± 0.81 d | |
| Treatments | Free Proline (µmol g−1 DW) | S.Sugar (mg g−1 DW) | α-TOC (µmol g−1 DW) | AsA (µmol g−1 FW) | GSH (µmol g−1 FW) | |
|---|---|---|---|---|---|---|
| Stress | Foliar Spray | |||||
| 2020 Season | ||||||
| Well-watered | Control | 2.52 ± 0.11 j | 10.7 ± 0.42 j | 1.55 ± 0.07 k | 1.20 ± 0.06 k | 0.830 ± 0.02 i |
| SA | 2.58 ± 0.18 ij | 11.4 ± 0.54 i | 1.64 ± 0.09 j | 1.25 ± 0.04 j | 0.850 ± 0.04 i | |
| CFE | 2.59 ± 0.15 ij | 11.8 ± 0.49 h | 1.71 ± 0.08 i | 1.28 ± 0.06 ij | 0.853 ± 0.06 i | |
| CFE+SA | 2.61 ± 0.12 i | 12.0 ± 0.63 h | 1.75 ± 0.06 i | 1.31 ± 0.03 i | 0.906 ± 0.04 h | |
| Moderate drought | Control | 4.31 ± 0.19 h | 16.4 ± 0.98 g | 2.14 ± 0.11 h | 1.54 ± 0.05 h | 1.10 ± 0.07 g |
| SA | 4.55 ± 0.21 g | 17.1 ± 0.87 f | 2.25 ± 0.13 g | 1.67 ± 0.06 g | 1.66 ± 0.06 f | |
| CFE | 4.79 ± 0.23 f | 18.5 ± 1.1 e | 2.41 ± 0.14 f | 1.84 ± 0.07 f | 1.39 ± 0.03 e | |
| CFE+SA | 5.02 ± 0.24 e | 19.6 ± 1.2 d | 2.56 ± 0.13 e | 1.96 ± 0.09 e | 1.56 ± 0.05 d | |
| Severe drought | Control | 6.43 ± 0.29 d | 23.3 ± 1.6 c | 2.97 ± 0.15 d | 2.36 ± 0.11 d | 2.07 ± 0.13 c |
| SA | 7.72 ± 0.32 c | 24.5 ± 1.4 b | 3.18 ± 0.17 c | 2.49 ± 0.14 c | 2.15 ± 0.14 b | |
| CFE | 6.88 ± 0.35 b | 24.8 ± 1.5 b | 3.28 ± 0.19 b | 2.66 ± 0.16 b | 2.19 ± 0.18 ab | |
| CFE+SA | 7.08 ± 0.34 a | 25.4 ± 1.5 a | 3.39 ± 0.16 a | 2.73 ± 0.18 a | 2.23 ± 0.19 a | |
| 2021 season | ||||||
| Well-watered | Control | 2.49 ± 0.11 g | 10.5 ± 0.55 i | 1.54 ± 0.05 l | 1.14 ± 0.05 k | 0.813 ± 0.03 h |
| SA | 2.53 ± 0.14 g | 11.3 ± 0.64 h | 1.62 ± 0.06 k | 1.20 ± 0.08 j | 0.823 ± 0.04 gh | |
| CFE | 2.54 ± 0.13 g | 11.6 ± 0.58 h | 1.67 ± 0.08 j | 1.24 ± 0.07 ij | 0.833 ± 0.03 gh | |
| CFE+SA | 2.57 ± 0.12 g | 11.7 ± 0.72 h | 1.73 ± 0.07 i | 1.29 ± 0.07 i | 0.883 ± 0.04 g | |
| Moderate drought | Control | 4.20 ± 0.22 f | 16.3 ± 0.81 g | 2.11 ± 0.12 h | 1.52 ± 0.06 h | 1.07 ± 0.08 f |
| SA | 4.51 ± 0.24 e | 16.7 ± 1.2 g | 2.20 ± 0.13 g | 1.65 ± 0.07 g | 1.21 ± 0.06 e | |
| CFE | 4.76 ± 0.26 d | 18.1 ± 1.4 f | 2.38 ± 0.16 f | 1.80 ± 0.08 f | 1.33 ± 0.05 d | |
| CFE+SA | 4.94 ± 0.33 d | 19.1 ± 1.7 e | 2.54 ± 0.11 e | 1.91 ± 0.09 e | 1.51 ± 0.08 c | |
| Severe drought | Control | 6.32 ± 0.36 c | 22.8 ± 1.5 b | 2.90 ± 0.13 d | 2.33 ± 0.14 d | 2.04 ± 0.16 b |
| SA | 6.50 ± 0.35 bc | 24.1 ± 1.6 c | 3.14 ± 0.14 c | 2.46 ± 0.17 c | 2.11 ± 0.18 a | |
| CFE | 6.61 ± 0.41 b | 24.5 ± 1.3 b | 3.23 ± 0.16 b | 2.64 ± 0.13 b | 2.13 ± 0.14 a | |
| CFE+SA | 7.05 ± 0.43 a | 25.1 ± 1.6 a | 3.34 ± 0.19 a | 2.71 ± 0.18 a | 2.15 ± 0.11 a | |
| Treatments | POX | CAT | APX | SOD | GR | |
|---|---|---|---|---|---|---|
| Stress | Foliar Spray | A564 min−1 mg−1 Protein | ||||
| 2020 season | ||||||
| Well-watered | Control | 1.56 ± 0.06 k | 39.8 ± 2.1 j | 30.5 ± 1.1 k | 4.14 ± 0.13 k | 26.5 ± 1.2 k |
| SA | 1.63 ± 0.08 j | 40.7 ± 1.9 i | 32.0 ± 1.3 j | 4.32 ± 0.21 j | 27.3 ± 1.4 j | |
| CFE | 1.67 ± 0.05 i | 42.0 ± 1.6 h | 32.9 ± 1.5 i | 4.43 ± 0.15 i | 27.7 ± 1.5 ij | |
| CFE+SA | 1.70 ± 0.09 i | 42.5 ± 1.7 h | 33.3 ± 1.2 i | 4.52 ± 0.23 i | 28.4 ± 1.4 i | |
| Moderate drought | Control | 2.35 ± 0.11 h | 48.1 ± 2.2 g | 46.2 ± 1.6 h | 6.63 ± 0.31 h | 37.3 ± 2.1 h |
| SA | 2.46 ± 0.12 g | 49.0 ± 2.5 f | 47.9 ± 1.8 g | 6.81 ± 0.35 g | 39.1 ± 2.2 g | |
| CFE | 2.57 ± 0.14 f | 49.9 ± 2.8 e | 49.8 ± 1.9 f | 7.31 ± 0.36 f | 43.0 ± 2.5 f | |
| CFE+SA | 2.63 ± 0.12 e | 50.5 ± 2.6 e | 51.0 ± 1.8 e | 7.51 ± 0.42 e | 45.7 ± 2.3 e | |
| Severe drought | Control | 3.15 ± 0.16 d | 62.7 ± 3.2 d | 63.9 ± 2.1 d | 8.77 ± 0.35 d | 57.7 ± 3.1 d |
| SA | 3.20 ± 0.17 c | 65.3 ± 3.6 c | 65.7 ± 2.2 c | 9.18 ± 0.39 c | 59.6 ± 3.6 c | |
| CFE | 3.25 ± 0.19 b | 68.2 ± 3.5 b | 66.9 ± 2.5 b | 9.40 ± 0.38 b | 61.2 ± 3.5 b | |
| CFE+SA | 3.30 ± 0.21 a | 69.7 ± 3.8 a | 67.8 ± 2.6 a | 9.52 ± 0.46 a | 63.0 ± 3.8 a | |
| 2021 season | ||||||
| Well-watered | Control | 1.51 ± 0.07 j | 38.9 ± 1.1 j | 30.1 ± 1.4 h | 4.11 ± 0.11 k | 26.0 ± 1.5 j |
| SA | 1.59 ± 0.06 i | 40.0 ± 1.5 i | 31.1 ± 1.5 gh | 4.23 ± 0.16 j | 26.5 ± 1.6 j | |
| CFE | 1.63 ± 0.05 hi | 41.2 ± 1.4 h | 31.9 ± 1.4 g | 4.39 ± 0.18 i | 26.7 ± 1.4 ij | |
| CFE+SA | 1.67 ± 0.08 h | 41.6 ± 1.9 h | 32.1 ± 1.2 g | 4.45 ± 0.15 i | 27.5 ± 1.3 i | |
| Moderate drought | Control | 2.32 ± 0.13 g | 47.2 ± 1.5 g | 45.6 ± 1.5 f | 6.60 ± 0.21 h | 36.4 ± 2.1 h |
| SA | 2.44 ± 0.15 f | 48.3 ± 1.9 f | 47.0 ± 1.3 e | 6.78 ± 0.32 g | 38.4 ± 2.5 g | |
| CFE | 2.54 ± 0.14 e | 48.9 ± 2.1 ef | 49.1 ± 1.4 d | 7.20 ± 0.36 f | 42.2 ± 2.6 f | |
| CFE+SA | 2.59 ± 0.12 d | 49.8 ± 2.3 e | 50.3 ± 1.7 d | 7.44 ± 0.35 e | 44.8 ± 2.8 e | |
| Severe drought | Control | 3.11 ± 0.15 c | 61.9 ± 2.6 d | 63.0 ± 2.3 c | 8.71 ± 0.39 d | 56.5 ± 3.4 d |
| SA | 3.14 ± 0.14 bc | 64.4 ± 3.1 c | 64.8 ± 2.5 b | 9.14 ± 0.32 c | 58.3 ± 3.5 c | |
| CFE | 3.16 ± 0.18 ab | 67.2 ± 3.2 b | 65.6 ± 3.1 b | 9.31 ± 0.41 b | 60.5 ± 3.9 b | |
| CFE+SA | 3.21 ± 0.16 a | 68.2 ± 35 a | 67.8 ± 3.1 a | 9.43 ± 0.42 a | 62.8 ± 3.7 a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Desoky, E.-S.M.; Selem, E.; Abo El-Maati, M.F.; Hassn, A.A.S.A.; Belal, H.E.E.; Rady, M.M.; AL-Harbi, M.S.; Ali, E.F. Foliar Supplementation of Clove Fruit Extract and Salicylic Acid Maintains the Performance and Antioxidant Defense System of Solanum tuberosum L. under Deficient Irrigation Regimes. Horticulturae 2021, 7, 435. https://doi.org/10.3390/horticulturae7110435
Desoky E-SM, Selem E, Abo El-Maati MF, Hassn AASA, Belal HEE, Rady MM, AL-Harbi MS, Ali EF. Foliar Supplementation of Clove Fruit Extract and Salicylic Acid Maintains the Performance and Antioxidant Defense System of Solanum tuberosum L. under Deficient Irrigation Regimes. Horticulturae. 2021; 7(11):435. https://doi.org/10.3390/horticulturae7110435
Chicago/Turabian StyleDesoky, El-Sayed M., Eman Selem, Mohamed F. Abo El-Maati, Asem A. S. A. Hassn, Hussein E. E. Belal, Mostafa M. Rady, Mohammad S. AL-Harbi, and Esmat F. Ali. 2021. "Foliar Supplementation of Clove Fruit Extract and Salicylic Acid Maintains the Performance and Antioxidant Defense System of Solanum tuberosum L. under Deficient Irrigation Regimes" Horticulturae 7, no. 11: 435. https://doi.org/10.3390/horticulturae7110435