
Impact of Drought and Flooding on Alkaloid Production in Annona crassiflora Mart

 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
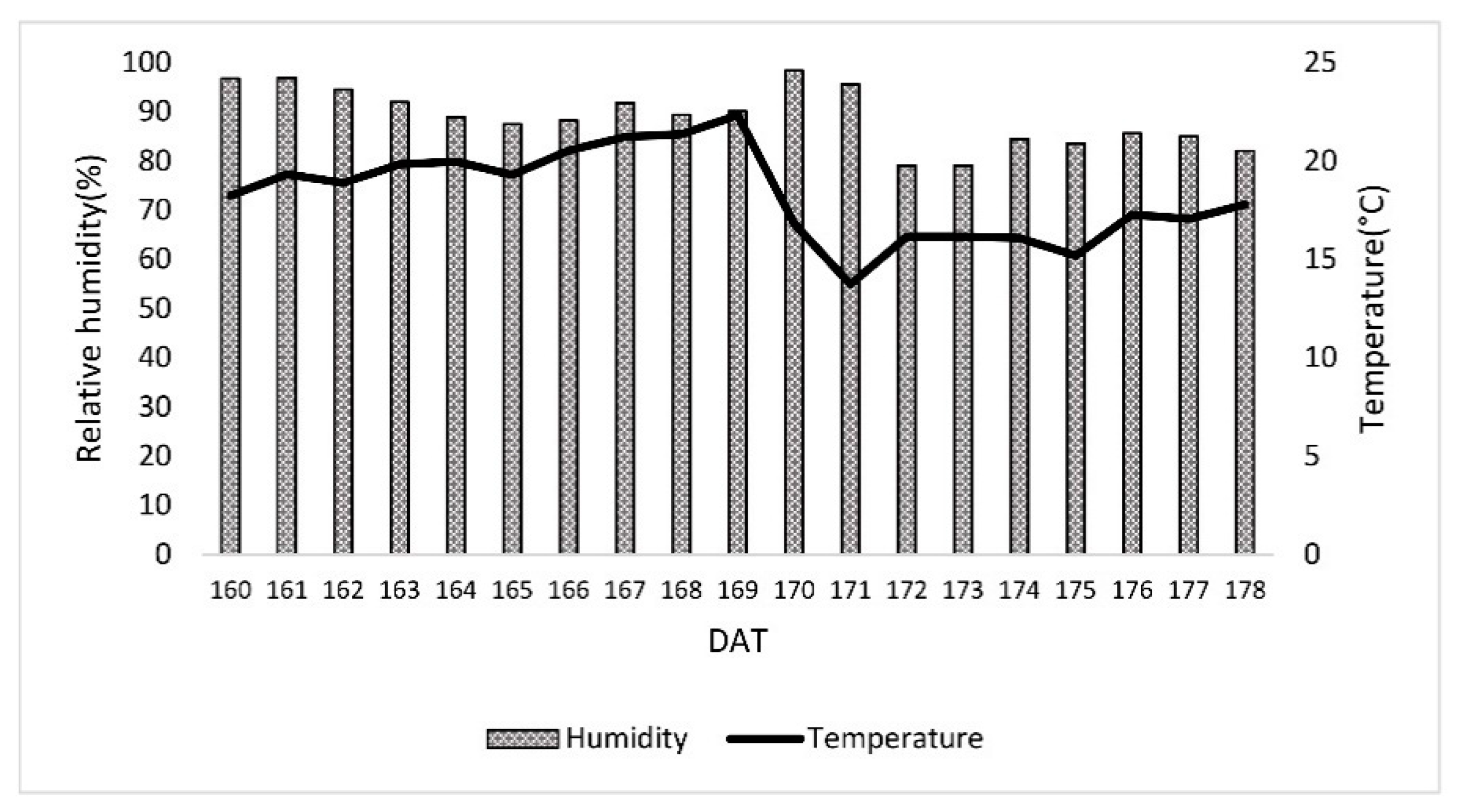
2. Materials and Methods
2.1. Plant Material
2.2. Experimental Design
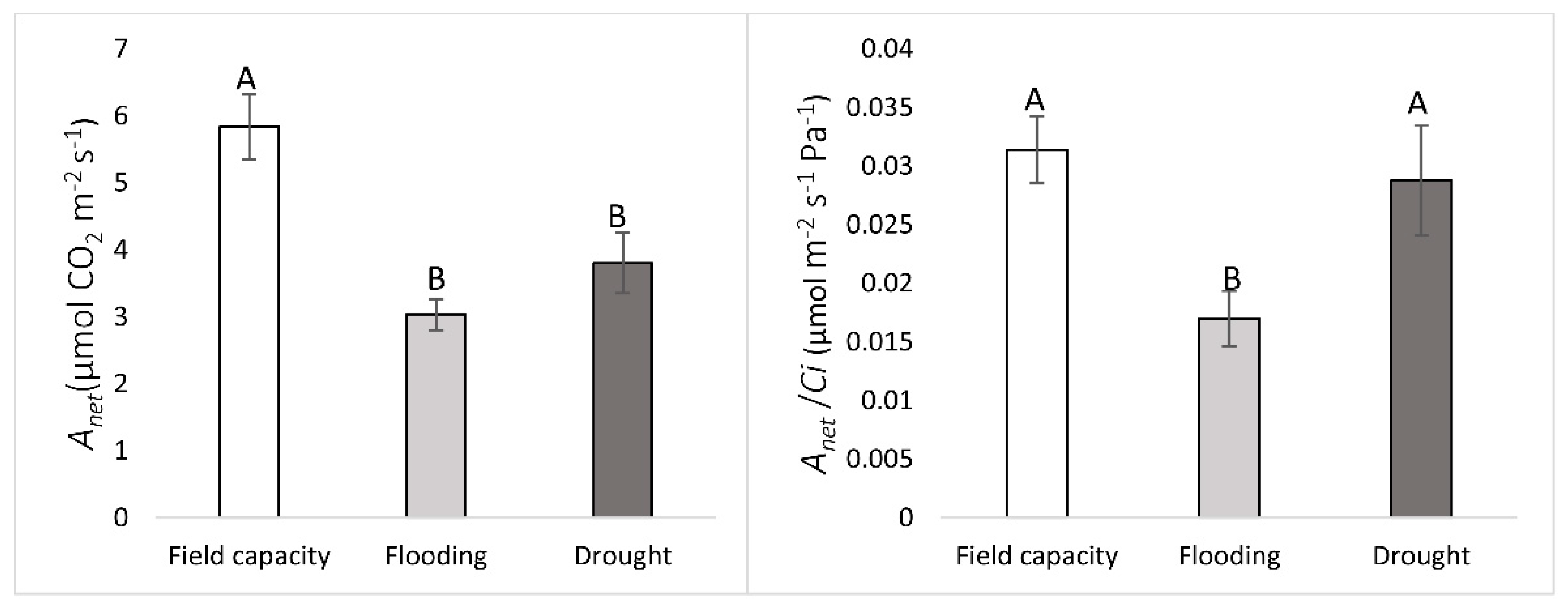
2.3. Gas Exchanges
2.4. Chlorophyll a Fluorescence
2.5. Carbohydrate Concentration
2.6. Activity of Antioxidant Enzymes, Hydrogen Peroxide and Lipid Peroxidation
2.7. Extraction of Total Alkaloids
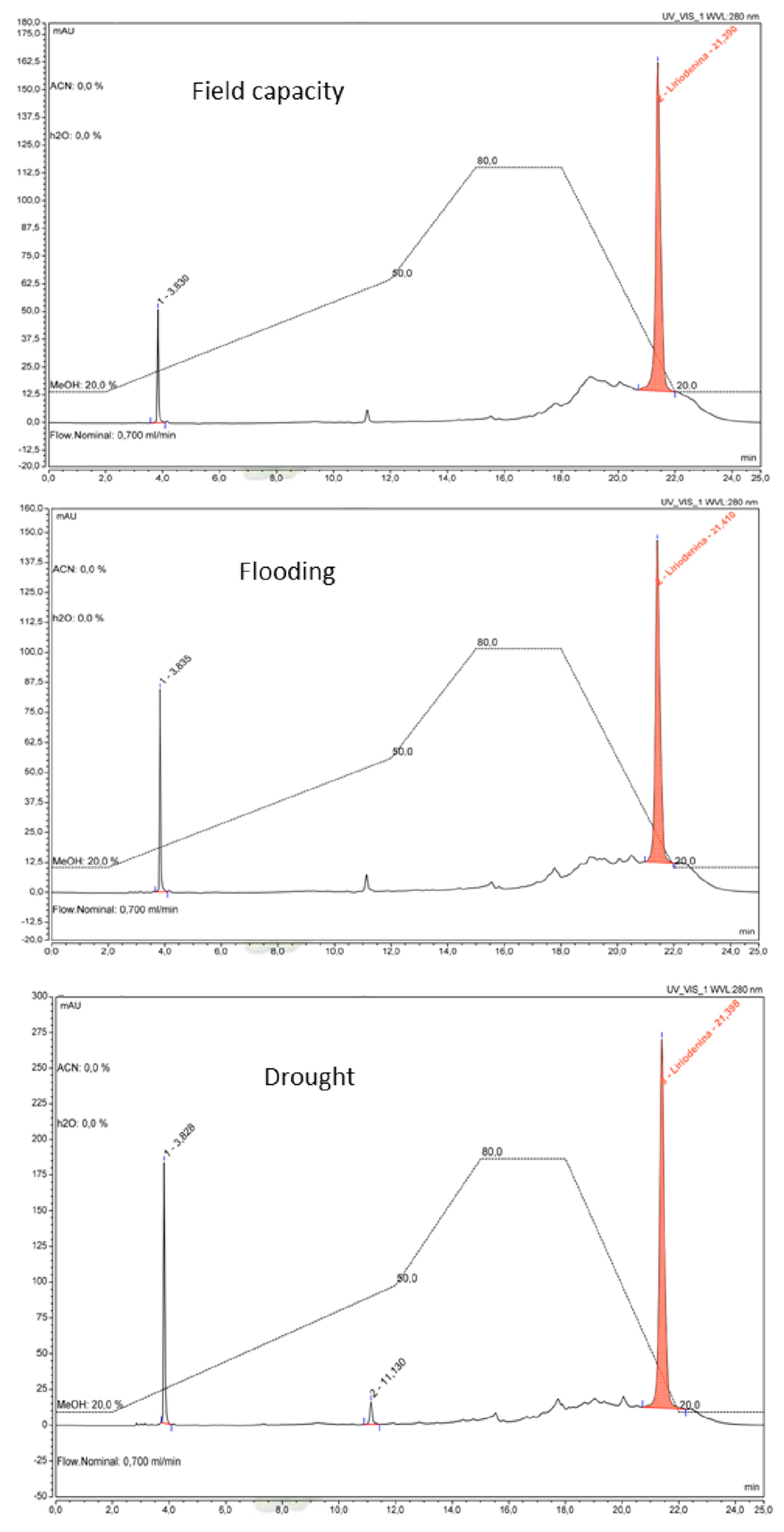
2.8. Quantification of Total Alkaloids and Liriodenine
2.9. Statistical Analysis
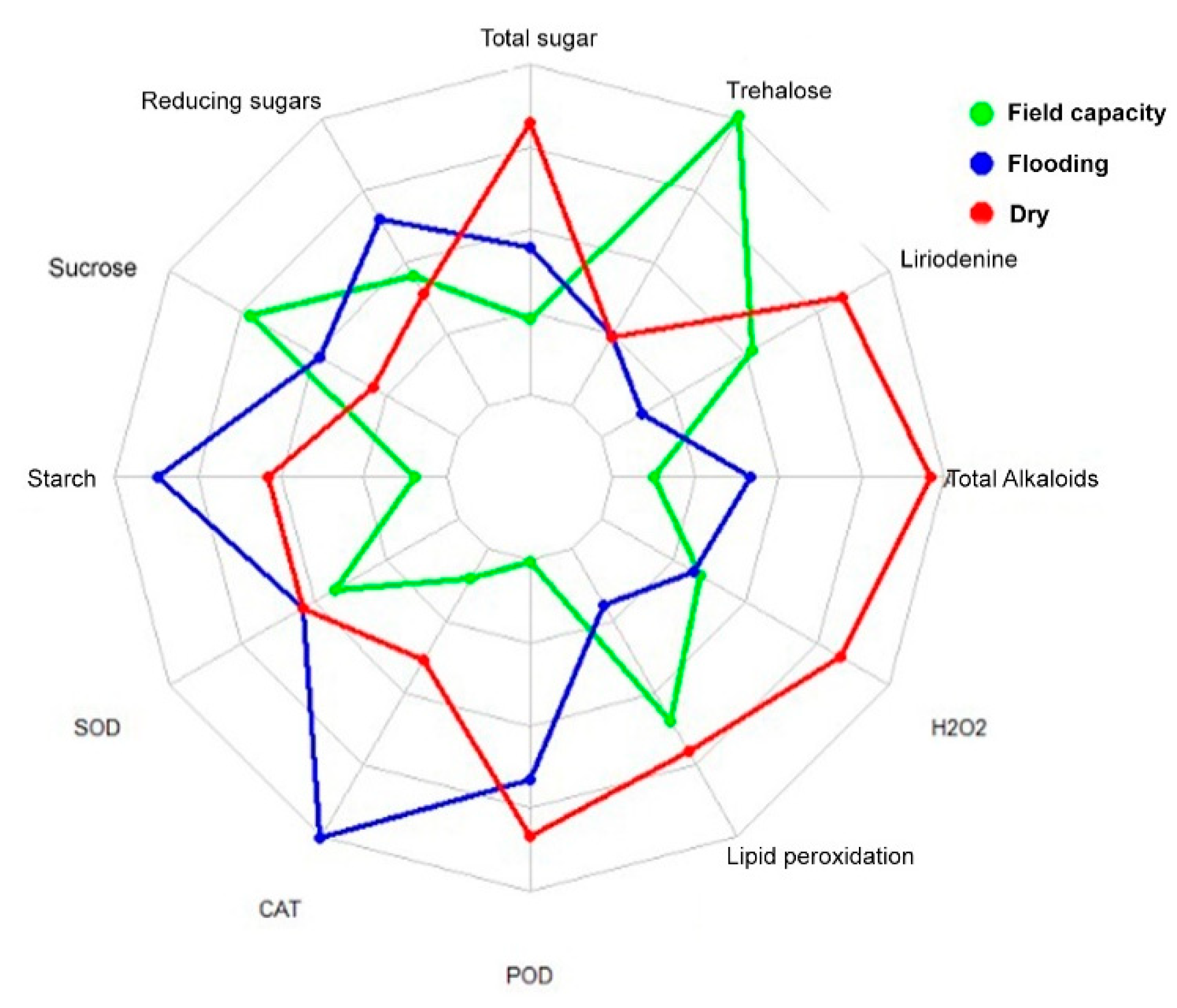
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Prado, L.G.; Arruda, H.S.; Araujo, N.M.P.; Braga, L.E.D.O.; Banzato, T.P.; Pereira, G.A.; Figueiredo, M.C.; Ruiz, A.L.T.G.; Eberlin, M.N.; de Carvalho, J.E.; et al. Antioxidant, antiproliferative and healing properties of araticum (Annona crassiflora Mart.) peel and seed. Food Res. Int. 2020, 133, 109168. [Google Scholar] [CrossRef]
- Rodrigues, J.A.M.; de Oliveira Andrade, A.C.; Viola, M.R.; Ferreira, D.D.; de Mello, C.R.; Thebaldi, M.S. Hydrological modeling in a basin of the Brazilian Cerrado biome. Rev. Ambient. Agua 2021, 16, 2–18. [Google Scholar] [CrossRef]
- Lima, J.E.F.W. Situação e perspectivass sobre as águas do Cerrado. Cienc. Cult. 2011, 63, 27–29. [Google Scholar] [CrossRef] [Green Version]
- Caldas, J.; Cerrados, E. Sustentabilidade Águas do Cerrado: Questão estratégica para o país. Manejo racional dos recursos hídricos é preocupação da pesquisa agropecuária uma vez que Cerrado é origem de grandes bacias hidrográficas. Sustentabilidade 2014, 9–11. Available online: http://www.diadecampo.com.br/zpublisher/materias/Newsletter.asp?id=28947&secao=Agrotemas (accessed on 16 October 2021).
- Lee, J.-E.; Lintner, B.R.; Boyce, C.K.; Lawrence, P.J. Land use change exacerbates tropical South American drought by sea surface temperature variability. Geophys. Res. Lett. 2011, 38. [Google Scholar] [CrossRef]
- Debortoli, N.S.; Dubreuil, V.; Funatsu, B.; Delahaye, F.; De Oliveira, C.H.; Rodrigues-Filho, S.; Saito, C.; Fetter, R. Rainfall patterns in the Southern Amazon: A chronological perspective (1971–2010). Clim. Chang. 2015, 132, 251–264. [Google Scholar] [CrossRef]
- Penereiro, J.C.; Badinger, A.; Maccheri, N.A.; Meschiatti, M.C. Distribuições de Tendências Sazonais de Temperatura Média e Precipitação nos Biomas Brasileiros. Rev. Bras. Meteorol. 2018, 33, 97–113. [Google Scholar] [CrossRef]
- Aghajanlou, F.; Mirdavoudi, H.; Shojaee, M.; Mac Sweeney, E.; Mastinu, A.; Moradi, P. Rangeland Management and Ecological Adaptation Analysis Model for Astragalus curvirostris Boiss. Horticulturae 2021, 7, 67. [Google Scholar] [CrossRef]
- Palhares, D.; Franco, A.C.; Zaidan, L.P. Respostas fotossintéticas de plantas do cerrado nas estações seca e chuvosa. Rev. Bras. Biociências 2010, 8, 213–220. [Google Scholar]
- Hoffmann, W.A.; Adasme, R.; Haridasan, M.; De Carvalho, M.T.; Geiger, E.L.; Pereira, M.A.B.; Gotsch, S.G.; Franco, A.C. Tree topkill, not mortality, governs the dynamics of savanna-forest boundaries under frequent fire in central Brazil. Ecology 2009, 90, 1326–1337. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, W.A.; Franco, A.C. The importance of evolutionary history in studies of plant physiological ecology: Examples from cerrados and forests of central Brazil. Braz. J. Plant Physiol. 2008, 20, 247–256. [Google Scholar] [CrossRef] [Green Version]
- Yousefi, A.R.; Rashidi, S.; Moradi, P.; Mastinu, A. Germination and seedling growth responses of Zygophyllum fabago, Salsola kali L. and Atriplex canescens to peg-induced drought stress. Environments 2020, 7, 107. [Google Scholar] [CrossRef]
- Justino, A.B.; Pereira, M.N.; Vilela, D.D.; Peixoto, L.G.; Martins, M.M.; Teixeira, R.R.; Miranda, N.C.; da Silva, N.M.; de Sousa, R.M.F.; de Oliveira, A.; et al. Peel of araticum fruit (Annona crassiflora Mart.) as a source of antioxidant compounds with α-amylase, α-glucosidase and glycation inhibitory activities. Bioorg. Chem. 2016, 69, 167–182. [Google Scholar] [CrossRef]
- Roesler, R.; Malta, L.G.; Carrasco, L.C.; Pastore, G.C. Food Chemistry and Toxicology Evaluation of the Antioxidant Properties of the Brazilian Cerrado Fruit Annona crassiflora (Araticum). Food Chem. Toxicol. 2006, 71, 102–107. [Google Scholar]
- Joca, T.A.C.; Oliveira, D.C.; de Zotz, G.; Winkler, U.; Moreira, A.S.F.P. The velamen of epiphytic orchids: Variation in structure and correlations with nutrient absorption. Flora Morphol. Distrib. Funct. Ecol. Plants 2017, 230, 66–74. [Google Scholar] [CrossRef]
- Cardoso, L.D.M.; Oliveira, D.D.S.; Bedetti, S.D.F.; Martino, H.S.D.; Pinheiro-Sant’Ana, H.M. Araticum (Annona crassiflora Mart.) from the Brazilian Cerrado: Chemical composition and bioactive compounds. Fruits 2013, 68, 121–134. [Google Scholar] [CrossRef]
- Modesto, E.C.; Santos, G.T.; dos Vilela, D.; Gonçalvez, G.D.; Matsushita, M. Efeitos nutricionais de dietas ricas em ácidos graxos poliinsaturados para os ruminantes e alguns benefícios para o homem. Arq. Ciências Veterinárias E Zool. UNIPAR 2002, 5, 119–134. [Google Scholar]
- Luzia, D.M.M.; Jorge, N. Bioactive substance contents and antioxidant capacity of the lipid fraction of Annona crassiflora Mart. seeds. Ind. Crops Prod. 2013, 42, 231–235. [Google Scholar] [CrossRef]
- Debnath, B.; Singh, W.S.; Das, M.; Goswami, S.; Singh, M.K.; Maiti, D.; Manna, K. Role of plant alkaloids on human health: A review of biological activities. Mater. Today Chem. 2018, 9, 56–72. [Google Scholar] [CrossRef]
- Gonçalves, M.A.; Lara, T.A.; Pimenta, L.P.S. Oxaporphynic alkaloids of Annona crassiflora wood Mart. Annu. Meet. Braz. Chem. Soc. 2006, 2, 1–2. [Google Scholar]
- Silva, M.A.; Da Silva, G.A.; da Marques, M.J.; Bastos, R.G.; da Silva, A.F.; Rosa, C.P.; Espuri, P.F. Triagem fitoquímica, atividade antioxidante e leishmanicida do extrato hidroetanólico 70% (v/v) e das frações obtidas de (Annona crassiflora Mart.). Rev. Fitos 2017, 10, 505–517. [Google Scholar] [CrossRef] [Green Version]
- Egydio, A.P.M.; Valvassoura, T.A.; Santos, D.Y.A.C. Geographical variation of isoquinoline alkaloids of Annona crassiflora Mart. from cerrado, Brazil. Biochem. Syst. Ecol. 2013, 46, 145–151. [Google Scholar] [CrossRef]
- Ferraz, C.R.; Silva, D.B.; Prado, L.C.D.S.; Canabrava, H.A.N.; Bispo-Da-Silva, L.B. Antidiarrhoeic effect and dereplication of the aqueous extract of Annona crassiflora (Annonaceae). Nat. Prod. Res. 2019, 33, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Costa, E.V.; Da Cruz, P.E.O.; De Lourenço, C.C.; De Souza Moraes, V.R.; De Lima Nogueira, P.C.; Salvador, M.J. Antioxidant and antimicrobial activities of aporphinoids and other alkaloids from the bark of Annona salzmannii A. DC. (Annonaceae). Nat. Prod. Res. 2013, 27, 1002–1006. [Google Scholar] [CrossRef] [PubMed]
- De La Cruz Chacón, I.; González-Esquinca, A.R. Liriodenine alkaloid in Annona diversifolia during early development. Nat. Prod. Res. 2012, 26, 42–49. [Google Scholar] [CrossRef] [PubMed]
- González-Esquinca, A.R.; De-La-Cruz-Chacón, I.; Castro-Moreno, M.; Orozco-Castillo, J.A.; Riley- Saldaña, C.A. Alkaloids and acetogenins in Annonaceae development: Biological considerations. Rev. Bras. Frutic. 2014, 36, 01–16. [Google Scholar] [CrossRef]
- Sousa, M.C.; Bronzatto, A.C.; González-Esquinca, A.R.; Campos, F.G.; Dalanhol, S.J.; Boaro, C.S.F.; Martins, A.L.; da Silva Almeida, J.R.G.; Costa, E.V.; De-la-Cruz-Chacón, I.; et al. The production of alkaloids in Annona cacans seedlings is affected by the application of GA4+7 + 6-Benzyladenine. Biochem. Syst. Ecol. 2019, 84, 47–51. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Wu, H.-M.; Chao, W.-Y.; Lee, C.-H. Review on pharmacological activities of liriodenine. Afr. J. Pharm. Pharmacol. 2014, 8, 364–371. [Google Scholar] [CrossRef]
- Liu, Y.; Meng, Q.; Duan, X.; Zhang, Z.; Li, D. Effects of PEG-induced drought stress on regulation of indole alkaloid biosynthesis in Catharanthus roseus. J. Plant Interact. 2017, 12, 87–91. [Google Scholar] [CrossRef] [Green Version]
- Costa, E.V.; Pinheiro, M.L.B.; Xavier, C.M.; Silva, J.R.A.; Amaral, A.C.F.; Souza, A.D.L.; Barison, A.; Campos, F.R.; Ferreira, A.G.; Machado, G.M.C.; et al. A pyrimidine-β-carboline and other alkaloids from Annona foetida with antileishmanial activity. J. Nat. Prod. 2006, 69, 292–294. [Google Scholar] [CrossRef]
- Cota, L.G.; Vieira, F.A.; Melo Júnior, A.F.; Brandão, M.M.; Santana, K.N.O.; Guedes, M.L.; Oliveira, D.A. Genetic diversity of Annona crassiflora (Annonaceae) in northern Minas Gerais State. Genet. Mol. Res. 2011, 10, 2172–2180. [Google Scholar] [CrossRef] [PubMed]
- De Lima, J.P.S.; Pinheiro, M.L.B.; Antonio, A.M.; José, J.L.; Santos, D.M.F.; Barison, A.; Silva-Jardim, I.; Costa, E.V. In Vitro Atileishmanial and Cytotoxic Activities of Annona mucosa (Annonaceae). Rev. Virtual Quim. 2012, 4, 692–702. [Google Scholar] [CrossRef]
- Khan, T.M.; Gul, N.S.; Lu, X.; Wei, J.H.; Liu, Y.C.; Sun, H.; Liang, H.; Orvig, C.; Chen, Z.F. In vitro and in vivo anti-tumor activity of two gold(III) complexes with isoquinoline derivatives as ligands. Eur. J. Med. Chem. 2019, 163, 333–343. [Google Scholar] [CrossRef] [PubMed]
- De-la-Cruz-Chacón, I.; López-Fernández, N.Y.; Riley-Saldaña, C.A.; Castro-Moreno, M.; González-Esquinca, A.R. Antifungal activity in vitro of Sapranthus microcarpus (Annonaceae) against phytopathogens. Acta Bot. Mex. 2019, 126, 1420. [Google Scholar] [CrossRef]
- Sousa, M.A.D.L.B.; Cavalheiro, F. Planejamento paisagístico do Campus Universitário da Faculdade de Ciências Agronômicas, UNESP, Botucatu, S.P. Acta Bot. Bras. 1987, 1, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Santos, H.G.; dos Jacomine, P.K.T.; Anjos, L.H.C.; dos Oliveira, V.Á.; de Lumbreras, J.F.; Coelho, M.R.; de Almeida, J.A.; de Araujo Filho, J.C.; de Oliveira, J.B.; Cunha, T.J.F. Sistema Brasileiro de Classificação de Solos, 5th ed.; EMBRAPA: Brasília, DF, Brazil, 2018; pp. 195–212. [Google Scholar]
- Kitajima, M.; Butler, W.L. Quenching of chlorophyll fluorescence and primary photochemistry in chloroplasts by dibromothymoquinone. BBA-Bioenerg. 1975, 376, 105–115. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta-Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Schreiber, U.; Schliwa, U.; Bilger, W. Continuous recording of photochemical and non-photochemical chlorophyll fluorescence quenching with a new type of modulation fluorometer. Photosynth. Res. 1986, 10, 51–62. [Google Scholar] [CrossRef]
- Bilger, W.; Björkman, O. Role of the xanthophyll cycle in photoprotection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in leaves of Hedera canariensis. Photosynth. Res. 1990, 25, 173–185. [Google Scholar] [CrossRef]
- Klughammer, C.; Schreiber, U. Complementary PS II quantum yields calculated from simple fluorescence parameters measured by PAM fluorometry and the Saturation Pulse method. PAM Appl. Notes 2008, 1, 27–35. [Google Scholar]
- Garcia, I.S.; Souza, A.; Barbedo, C.J.; Dietrich, S.M.C.; Figueiredo-Ribeiro, R.C.L. Changes in soluble carbohydrates during storage of Caesalpinia echinata LAM. (Brazilwood) seeds, an endangered leguminous tree from the Brazilian atlantic forest. Braz. J. Biol. 2006, 66, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Clegg, K.M. The application of the anthrone reagent to the estimation of starch in cereals. J. Sci. Food Agric. 1956, 7, 40–44. [Google Scholar] [CrossRef]
- Morris, D.L. Quantitative Determination of Carbohydrates with Dreywood’s Anthrone Reagent. American Association for the Advancement of Science Stable. Am. Assoc. Adv. Sci. 1948, 107, 254–255. [Google Scholar]
- Yemm, E.W.; Folkes, B.F. The Estimation of Carbhydrates in Plant Extracts by Anthrone. Int. J. Pharm. Pract. 1954, 57, 508–514. [Google Scholar] [CrossRef]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Passos, L.P. Métodos Analíticos e Laboratoriais em Fisiologia Vegetal, 1st ed.; EMBRAPA: Coronel Pacheco, MG, Brazil, 1996; p. 223. [Google Scholar]
- Kar, M.; Mishra, D. Catalase, Peroxidase and Polyphenoloxidase Activities during Rice Leaf Senescence. Plant Physiol. 1976, 57, 315–319. [Google Scholar] [CrossRef] [Green Version]
- Peixoto, P.H.P.; Cambraia, J.; Sant’Anna, R.; Mosquim, P.R.; Moreira, M.A. Aluminum effects on lipid peroxidation and on the activities of enzymes of oxidative metabolism in sorghum. Rev. Bras. Fisiol. Veg. 1999, 11, 137–143. [Google Scholar]
- Teisseire, H.; Guy, V. Copper-induced changes in antioxidant enzymes activities in fronds of duckweed (Lemna minor). Plant Sci. 2000, 153, 65–72. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizin the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 248–254. [Google Scholar] [CrossRef]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat V. Trop. Ecol. 2001, 45, 315–325. [Google Scholar] [CrossRef]
- Heatuh, R.L.; Packer, L. Photoperoxidation in Isolated Chloroplasts. Archives of Biochemistry and Biophysics 1968, 125, 850–857. [Google Scholar] [CrossRef]
- Costa, J.R. Técnicas Experimentais aplicadas às Ciências Agrárias. Embrapa Agrobiol. 2003, 7–102. [Google Scholar]
- Becker, R.A.; Chambers, J.M. Auditing of Data Analyses. SIAM J. Sci. Stat. Comput. 1988, 9, 747–760. [Google Scholar] [CrossRef]
- Nascimento, D.T.F.; Novais, G.T. Clima do Cerrado: Dinâmica atmosférica e características, variabilidades e tipologias climáticas. Élisée-Rev. De Geogr. Da UEG 2020, 9, e922021. [Google Scholar]
- Ghorbanpour, M.; Hatami, M. Role of plant growth promoting rhizobacteria on antioxidant enzyme activities and tropane alkaloid production of Hyoscyamus niger under water deficit stress Role of plant growth promoting rhizobacteria on antioxidant enzyme activities and tropane alkaloid. Turk. J. Biol. 2013, 37, 350–360. [Google Scholar] [CrossRef]
- Kleinwächter, M.; Selmar, D. New insights explain that drought stress enhances the quality of spice and medicinal plants: Potential applications. Agron. Sustain. Dev. 2015, 35, 121–131. [Google Scholar] [CrossRef] [Green Version]
- Kleinwächter, M.; Paulsen, J.; Bloem, E.; Schnug, E.; Selmar, D. Moderate drought and signal transducer induced biosynthesis of relevant secondary metabolites in thyme (Thymus vulgaris), greater celandine (Chelidonium majus) and parsley (Petroselinum crispum). Ind. Crops Prod. 2015, 64, 158–166. [Google Scholar] [CrossRef]
- Castro-Moreno, M.; Tinoco-Ojangurén, C.L.; Cruz-Ortega, M.D.R.; González-Esquinca, A.R. Influence of seasonal variation on the phenology and liriodenine content of Annona lutescens (Annonaceae). J. Plant Res. 2013, 126, 529–537. [Google Scholar] [CrossRef]
- De-la-Cruz-Chacón, I.; Riley-Saldaña, C.A.; Arrollo-Gómez, S.; Sancristóbal-Domínguez, T.J.; Castro-Moreno, M.; González-Esquinca, A.R. Spatio-Temporal Variation of Alkaloids in Annona purpurea and the Associated Influence on Their Antifungal Activity. Chem. Biodivers. 2019, 16, 1–14. [Google Scholar] [CrossRef]
- Selmar, D.; Kleinwächter, M. Stress enhances the synthesis of secondary plant products: The impact of stress-related over-reduction on the accumulation of natural products. Plant Cell Physiol. 2013, 54, 817–826. [Google Scholar] [CrossRef]
- Simonneau, T.; Lebon, E.; Coupel-Ledru, A.; Marguerit, E.; Rossdeutsch, L.; Ollat, N.; Soni, A.; Kumari, P.; Dhakar, S.; Kumar, N.; et al. Physiological behavior of Annona muricata, Dipteryx odorata and Copaifera langsdorffii in response to water and light stress. Front. Plant Sci. 2017, 62, 1–8. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, A.K.M.; Gualtieri, S.C.J. Trocas gasosas e grau de tolerância ao estresse hídrico induzido em plantas jovens de Tabebuia aurea (Paratudo) submetidas a alagamento. Ciência Florest. 2017, 27, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Selmar, D.; Kleinwächter, M. Influencing the product quality by deliberately applying drought stress during the cultivation of medicinal plants. Ind. Crops Prod. 2013, 42, 558–566. [Google Scholar] [CrossRef]
- Mantoan, L.P.B.; Rolim de Almeida, L.F.; Macedo, A.C.; Ferreira, G.; Boaro, C.S.F. Photosynthetic adjustment after rehydration in Annona emarginata. Acta Physiol. Plant. 2016, 38, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef] [Green Version]
- Parolin, P.; Wittmann, F. Struggle in the flood: Tree responses to flooding stress in four tropical floodplain systems. AoB Plants 2010, 2010, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mielke, M.S.; Matos, E.M.; Couto, V.B.; Almeida, A.-A.F.; de Gomes, F.P.; Mangabeira, P.A.O. Some photosynthetic and growth responses of Annona glabra L. seedlings to soil flooding. Acta Bot. Bras. 2005, 19, 905–911. [Google Scholar] [CrossRef] [Green Version]
- Nowak, M.; Kleinwächter, M.; Manderscheid, R.; Weigel, H.J.; Selmar, D. Drought stress increases the accumulation of monoterpenes in sage (Salvia officinalis), an effect that is compensated by elevated carbon dioxide concentration. J. Appl. Bot. Food Qual. 2010, 83, 133–136. [Google Scholar]
- Junior, U.O.B.; Lima, M.D.R.; Barbosa, M.A.M.; Batista, B.L.; Lobato, A. Biochemical Responses of Two Species of Eucalyptus Exposed to Aluminium Toxicity: Oxidative Stress and Antioxidant Metabolism. Not. Bot. Horti Agrobot. Cluj-Napoca 2016, 44, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Sataloff, R.T.; Johns, M.M.; Kost, K.M.; Arndt, S.K.; Irawan, A.; Sanders, G.J.; Boanares, D.; Kozovits, A.R.; Lemos-Filho, J.P.; Isaias, R.M.S.; et al. Responses of photosynthesis, lipid peroxidation and antioxidant system in leaves of Quercus mongolica to elevated O3. J. Exp. Bot. 2014, 69, 907–912. [Google Scholar] [CrossRef]
- Silva, A.A.; da Gonçalves, R.C. Espécies reativas do oxigênio e as doenças respiratórias em grandes animais. Ciência Rural 2010, 40, 994–1002. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, M.R.; de Araújo Silva, M.M.; Willadino, L.; Ulisses, C.; Camara, T.R. Geração e desintoxicação enzimática de espécies reativas de oxigênio em plantas. Cienc. Rural 2014, 44, 453–460. [Google Scholar] [CrossRef] [Green Version]
- Lignan, C.; Suberin, L.; Stilbenes, C.; Tannin, F. Phenylpropanoids comprise a multitude of plant secondary metabolites and cell wall components. In Plant Biochemistry, 4th ed.; Elsevier Academic Press: Amsterdam, The Netherlands, 2011; pp. 435–454. [Google Scholar]
- Møller, I.M.; Jensen, P.E.; Hansson, A. Oxidative modifications to cellular components in plants. Annu. Rev. Plant Biol. 2007, 58, 459–481. [Google Scholar] [CrossRef] [Green Version]
- Hossain, Z.; López-Climent, M.F.; Arbona, V.; Pérez-Clemente, R.M.; Gómez-Cadenas, A. Modulation of the antioxidant system in citrus under waterlogging and subsequent drainage. J. Plant Physiol. 2009, 166, 1391–1404. [Google Scholar] [CrossRef]
- Cortina, C.; Culiáñez-Macià, F.A. Tomato abiotic stress enhanced tolerance by trehalose biosynthesis. Plant Sci. 2005, 169, 75–82. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Manivannan, P.; Kishorekumar, A.; Sankar, B.; Gopi, R.; Somasundaram, R.; Panneerselvam, R. Alterations in osmoregulation, antioxidant enzymes and indole alkaloid levels in Catharanthus roseus exposed to water deficit. Colloids Surf. B Biointerfaces 2007, 59, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2015, 63, 1593–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Liang, J.; Tang, G.; Wang, X.; Liu, F.; Zhao, D. Drought stress affects on growth, water use efficiency, gas exchange and chlorophyll fluorescence of Juglans rootstocks. Sci. Hortic. 2019, 250, 230–235. [Google Scholar] [CrossRef]
- Thalmann, M.; Santelia, D. Starch as a determinant of plant fitness under abiotic stress. New Phytol. 2017, 214, 943–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, F.; Ra, M.; Ismail, M.R.; Juraimi, A.S.; Rahim, H.A.; Asfaliza, R.; Latif, M.A. Waterlogging Tolerance of Crops: Breeding Mechanism of Tolerance Molecular Approaches and Future Prospects. BioMed Res. Int. 2013, 2013. [Google Scholar] [CrossRef]
- Malik, A.I.; Colmer, T.D.; Lambers, H.; Setter, T.L.; Schortemeyer, M. Short-term waterlogging has long-term effects on the growth and physiology of wheat. New Phytol. 2002, 4, 225–236. [Google Scholar] [CrossRef]
- Huang, B.; Johnson, J.W. Root respiration and carbohydrate status of two wheat genotypes in response to hypoxia. Annals of Botany 1995, 75, 427–432. [Google Scholar] [CrossRef]
- Malik, A.I.; Colmer, T.D. Changes in physiological and morphological traits of roots and shoots of wheat in response to different depths of waterlogging. Funct. Plant Biol. 2001, 28, 1121–1131. [Google Scholar] [CrossRef]
- Setter, T.L.; Waters, I.; Sharma, S.K.; Singh, K.N.; Kulshreshtha, N.; Yaduvanshi, N.P.S.; Ram, P.C. Review of wheat improvement for waterlogging tolerance in Australia and India: The importance of anaerobiosis and element toxicities associated with different soils. Ann. Bot. 2008, 103, 221–235. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, O.; Béthencourt, L.; Quero, A.; Sangwan, R.S.; Clément Christophe, C. Trehalose and plant stress responses: Friend or foe? Trends Plant Sci. 2010, 15, 409–417. [Google Scholar] [CrossRef]
- Paul, M.J.; Primavesi, L.F.; Jhurreea, D.; Zhang, Y. Trehalose metabolism and signaling. Annu. Rev. Plant Biol. 2008, 59, 417–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tension | Humidity (%) |
|---|---|
| Flooding (>0.01 MPa) | 37 |
| Field capacity (−0.01 MPa) | 16 |
| Drought (−1.5 MPa) | 8 |
| Water Condition | Total Alk | Liriodenine | CAT | POD | Trehalose | Starch |
|---|---|---|---|---|---|---|
| Field capacity | 54.26 B | 10.8770 AB | 0.0101 B | 0.2681 B | 83.90 A | 69.713 B |
| Flooding | 63.49 AB | 8.4098 B | 0.1034 A | 0.6655 A | 12.46 B | 104.089 A |
| Drought | 80.87 A | 13.4374 A | 0.0391 B | 0.7656 A | 13.16 B | 89.381 AB |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Honório, A.B.M.; De-la-Cruz-Chacón, I.; Martínez-Vázquez, M.; da Silva, M.R.; Campos, F.G.; Martin, B.C.; da Silva, G.C.; Fernandes Boaro, C.S.; Ferreira, G. Impact of Drought and Flooding on Alkaloid Production in Annona crassiflora Mart. Horticulturae 2021, 7, 414. https://doi.org/10.3390/horticulturae7100414
Honório ABM, De-la-Cruz-Chacón I, Martínez-Vázquez M, da Silva MR, Campos FG, Martin BC, da Silva GC, Fernandes Boaro CS, Ferreira G. Impact of Drought and Flooding on Alkaloid Production in Annona crassiflora Mart. Horticulturae. 2021; 7(10):414. https://doi.org/10.3390/horticulturae7100414
Chicago/Turabian StyleHonório, Ana Beatriz Marques, Iván De-la-Cruz-Chacón, Mariano Martínez-Vázquez, Magali Ribeiro da Silva, Felipe Girotto Campos, Bruna Cavinatti Martin, Gustavo Cabral da Silva, Carmen Sílvia Fernandes Boaro, and Gisela Ferreira. 2021. "Impact of Drought and Flooding on Alkaloid Production in Annona crassiflora Mart" Horticulturae 7, no. 10: 414. https://doi.org/10.3390/horticulturae7100414