
Mechanisms of Nitric Oxide in the Regulation of Chilling Stress Tolerance in Camellia sinensis

 and
and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Experimental Materials
2.2. Experimental Method
Experimental Design
2.3. Measurement Items and Methods
2.4. Fluorescence Quantitative PCR Analysis
2.5. Data Processing
3. Results
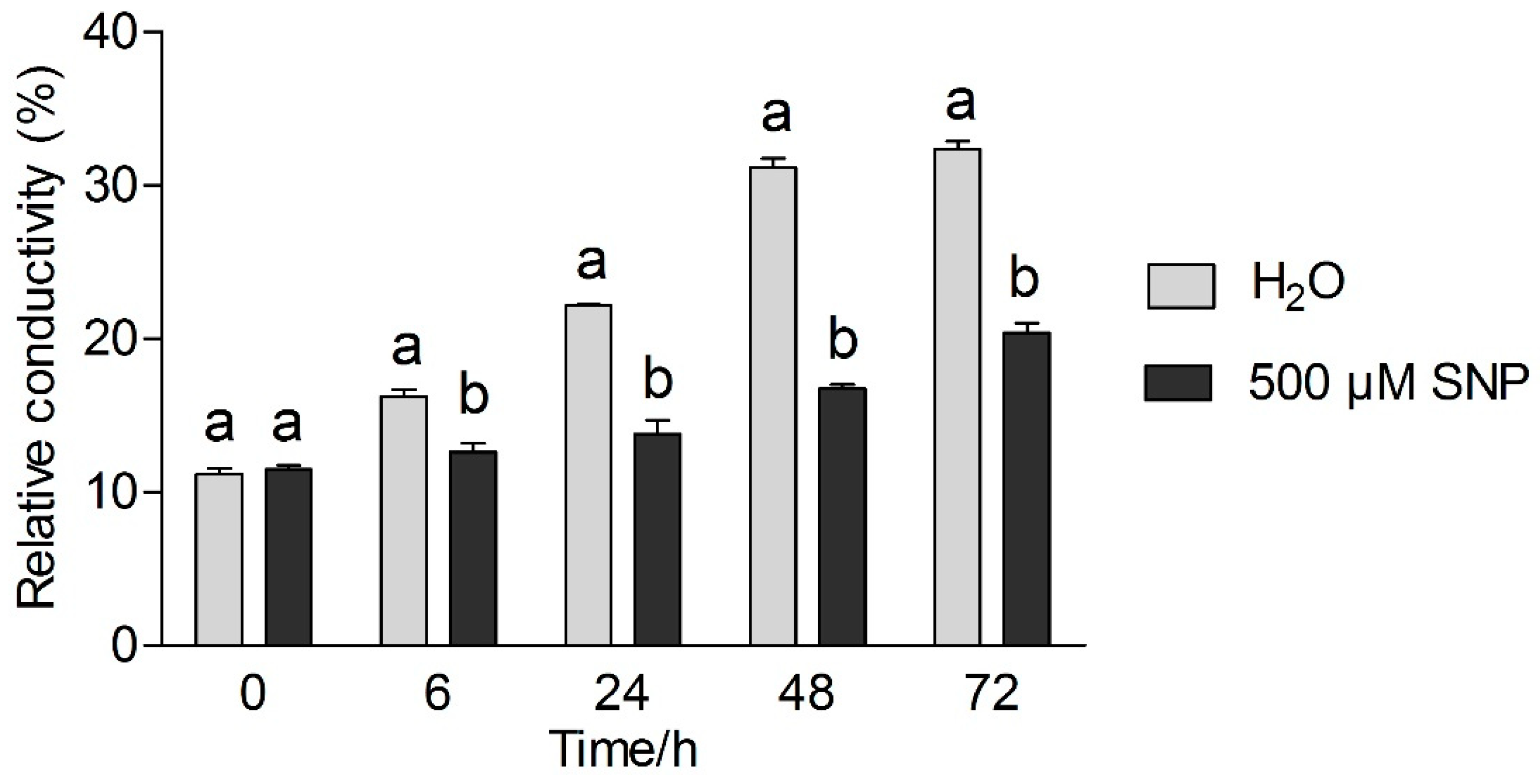
3.1. Effects of Exogenous NO and Low Temperature Stress on Relative Electrical Conductivity of Tea Leaves
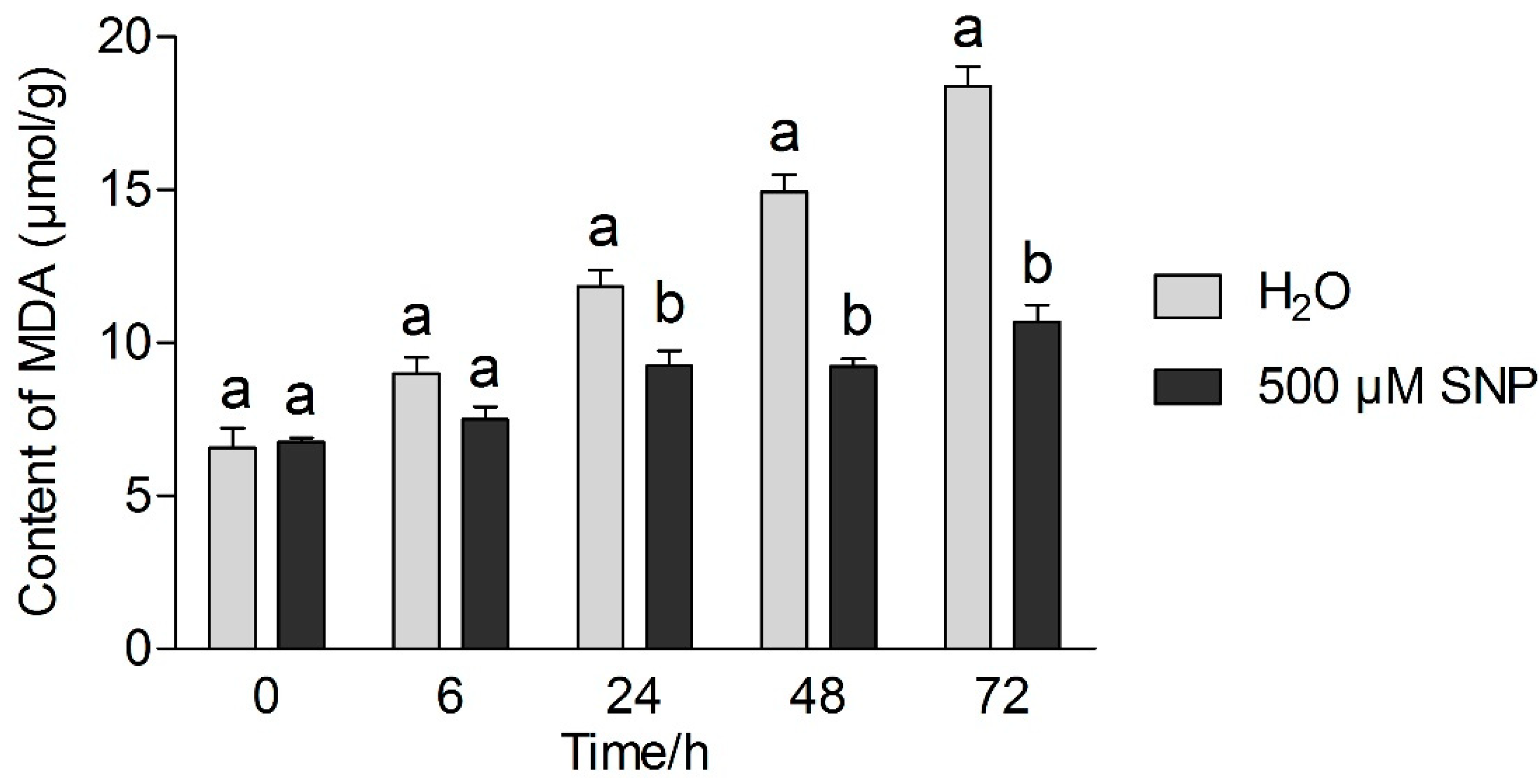
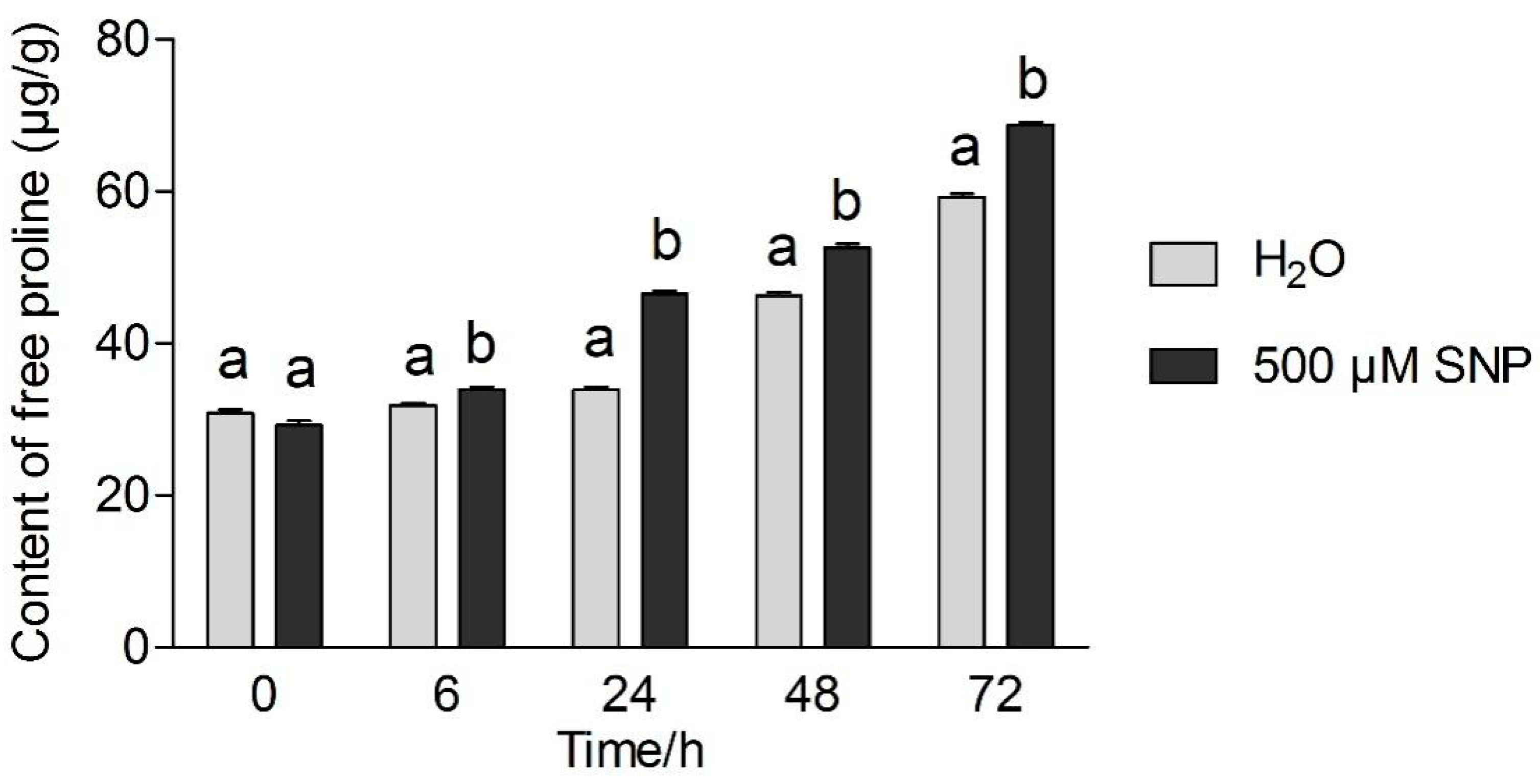
3.2. Effects of Exogenous NO and Low Temperature Stress on MDA and Proline Content in Tea Leaves
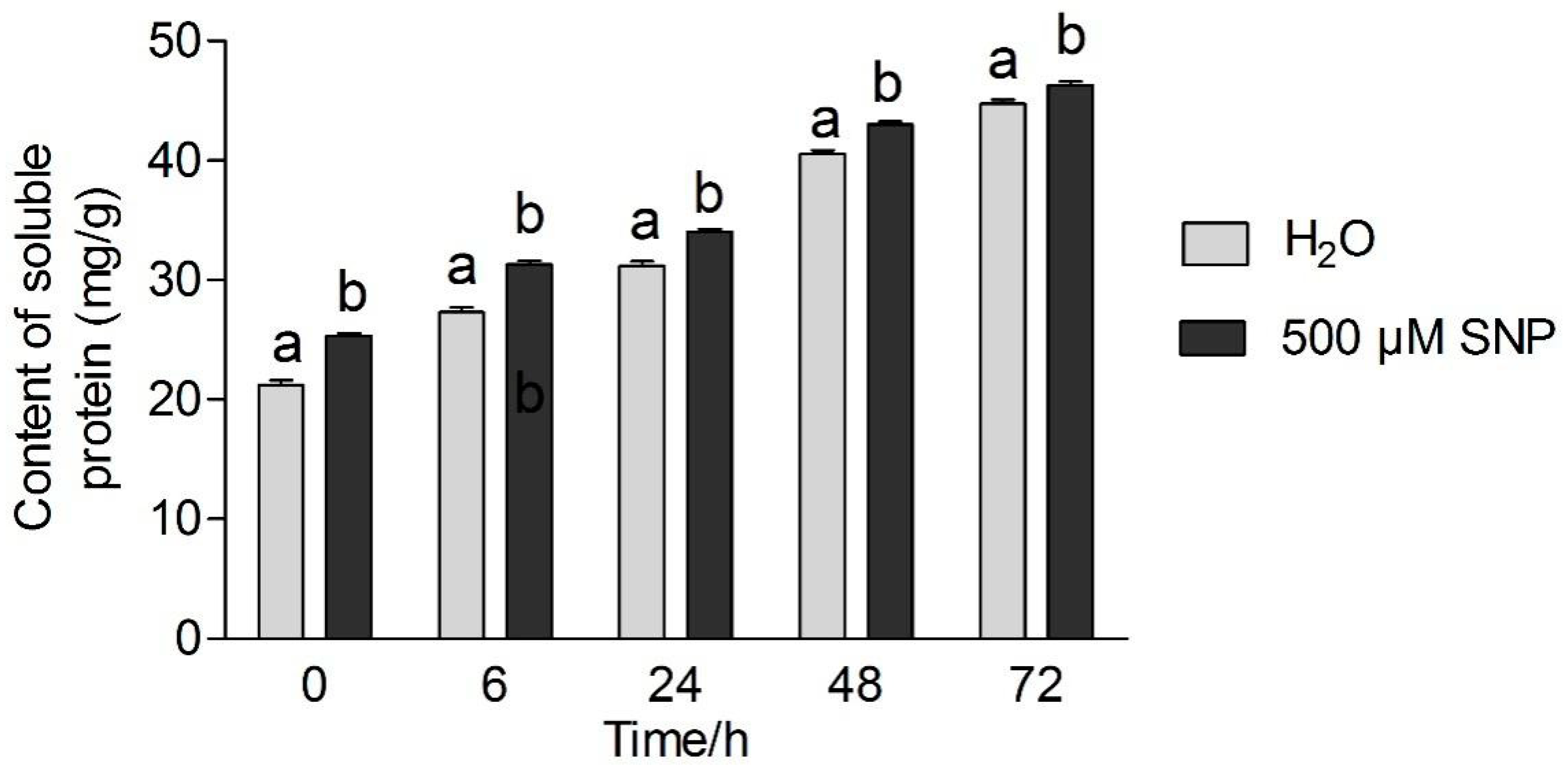
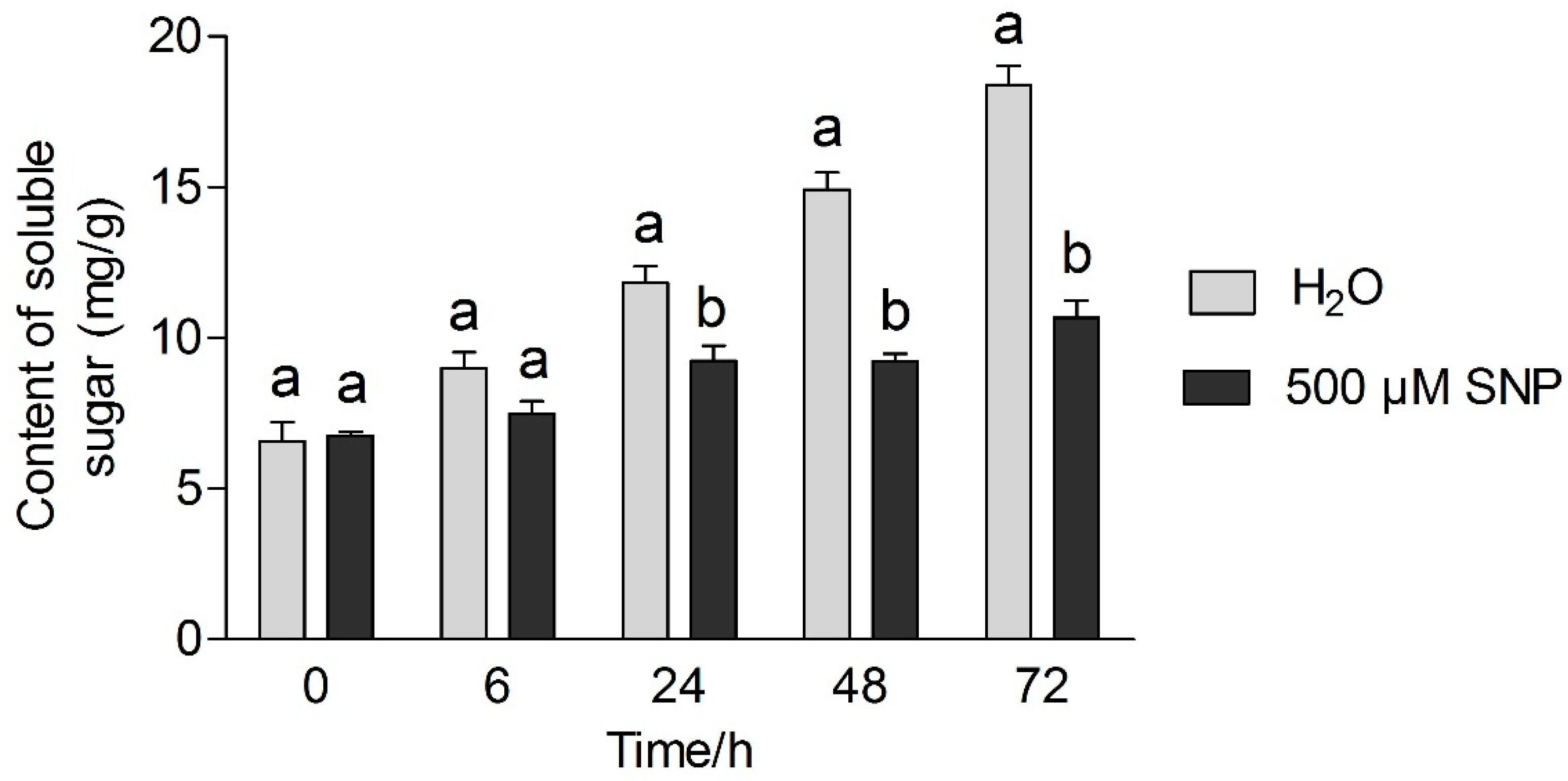
3.3. Effects of Exogenous NO and Low Temperature Stress on Soluble Protein and Sugar Content in Tea Leaves
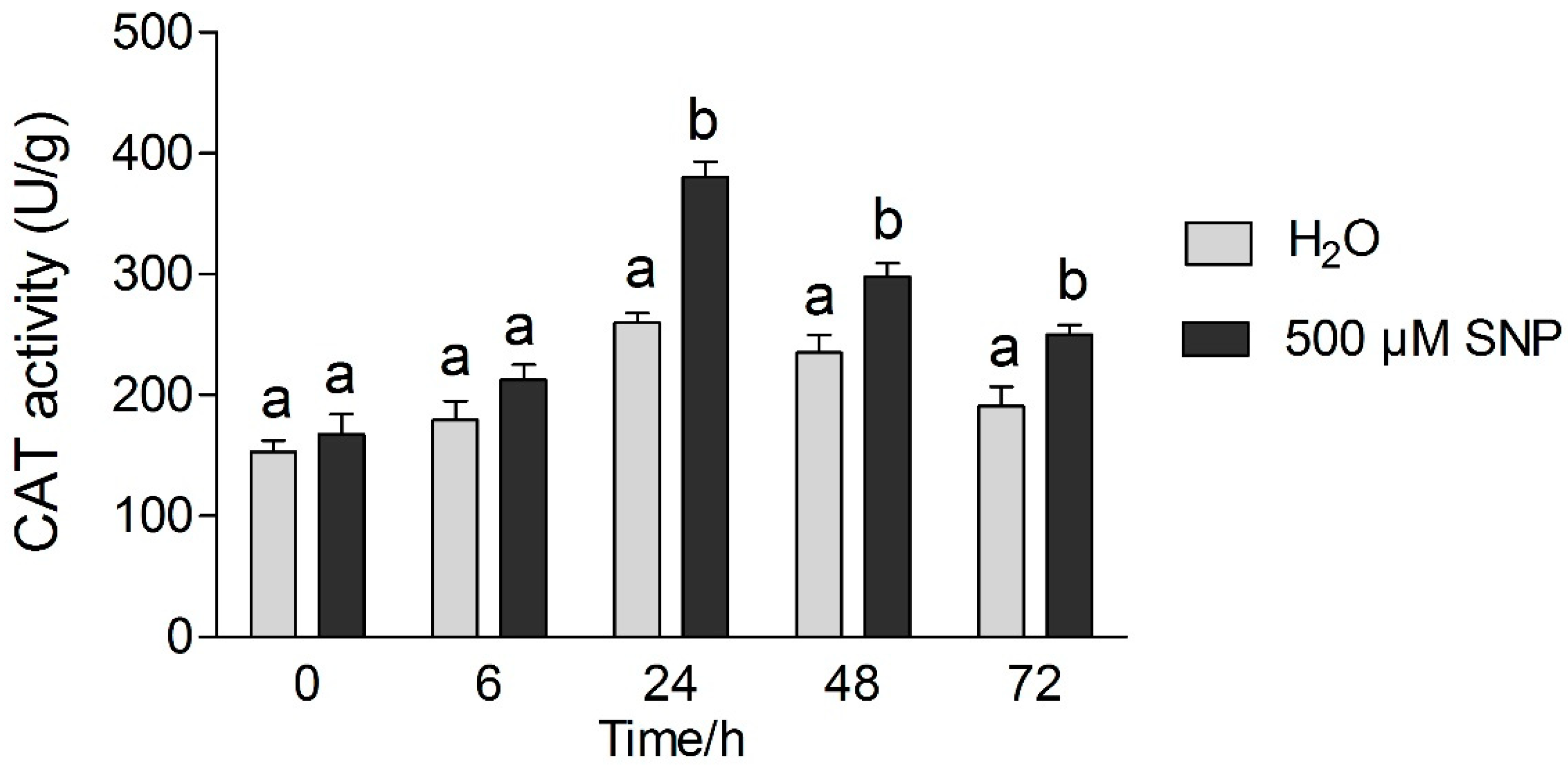
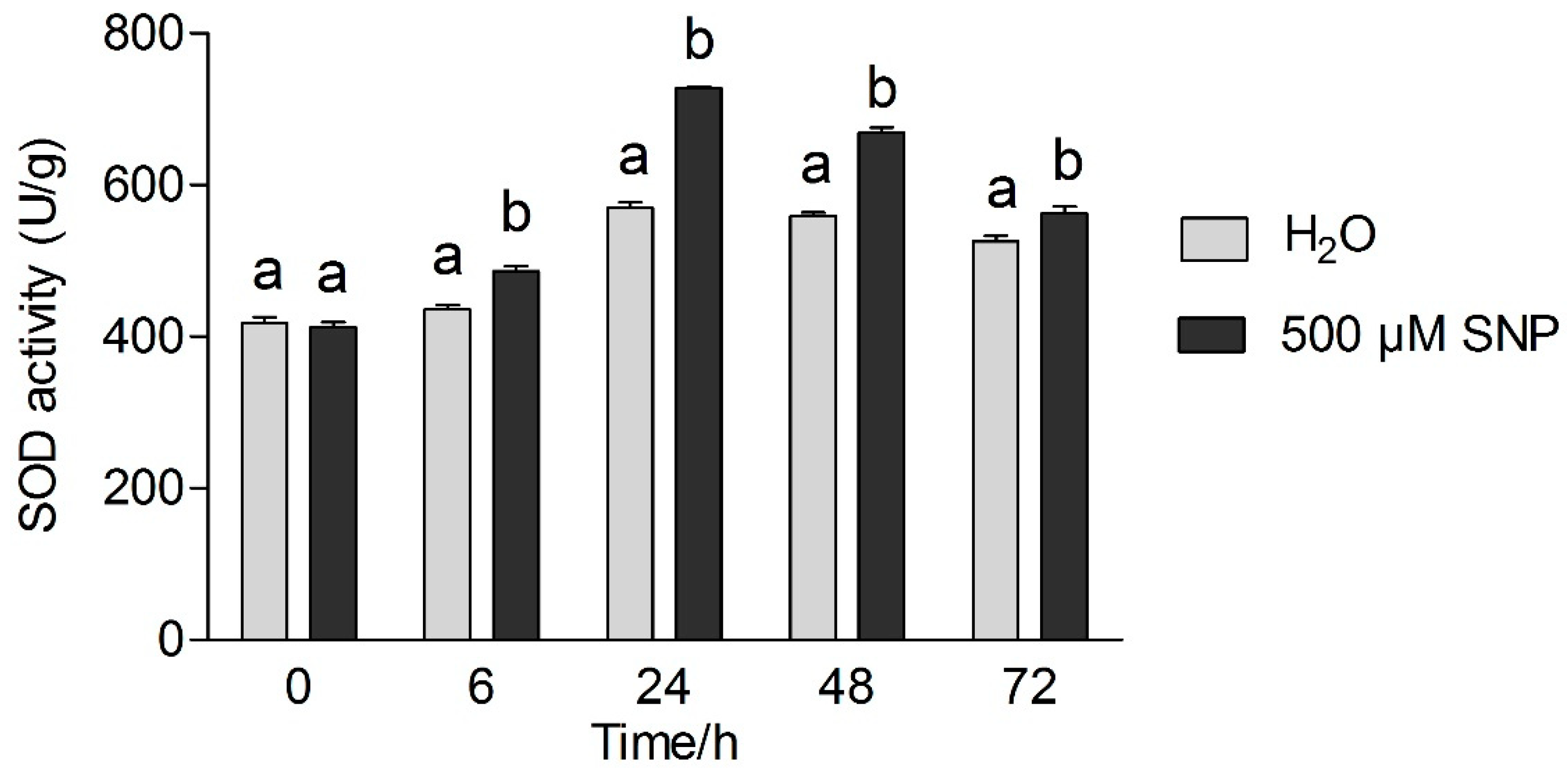
3.4. Effects of Exogenous NO and Low Temperature Stress on Antioxidant Enzymes Activity in Tea Leaves
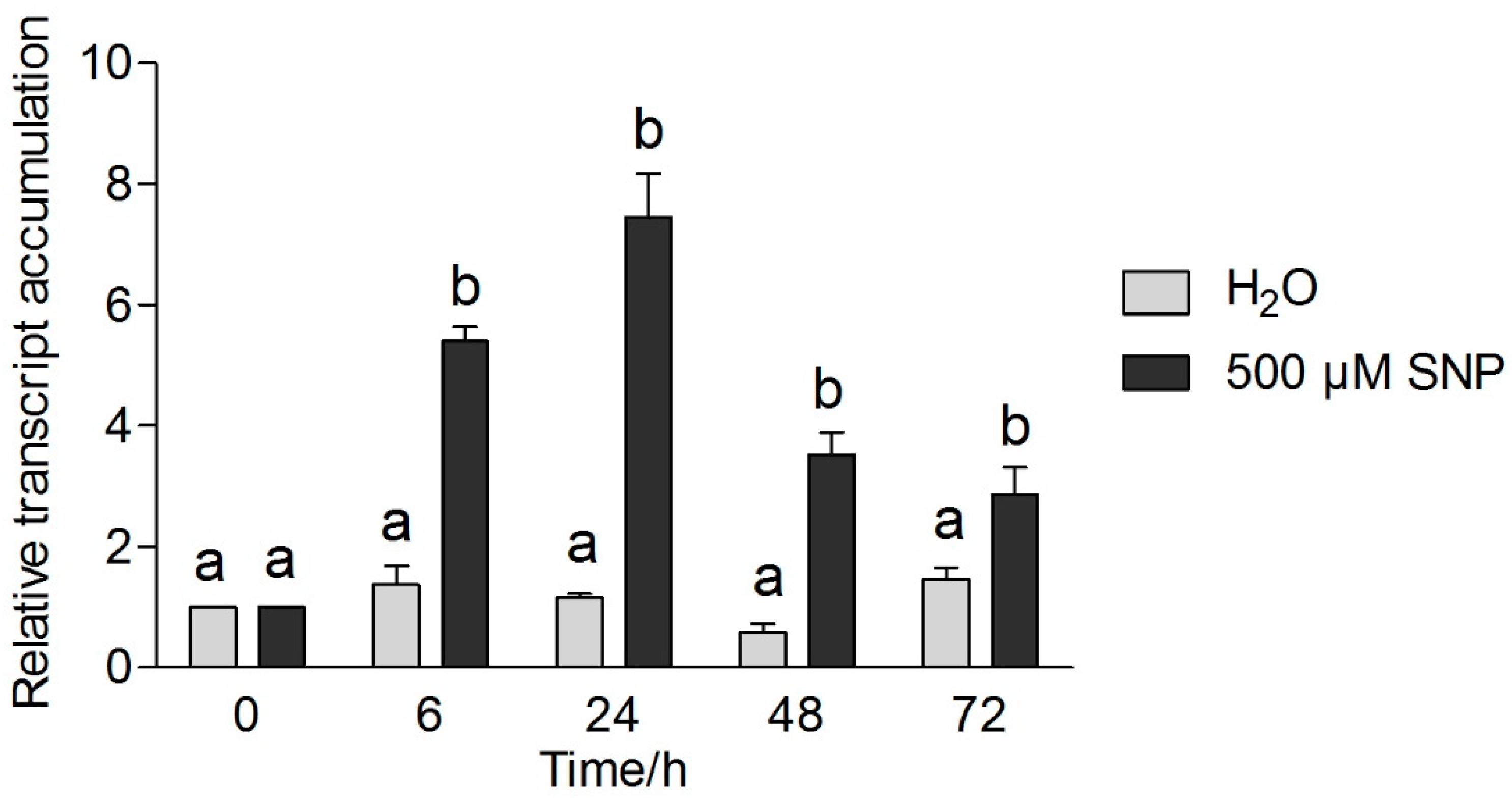
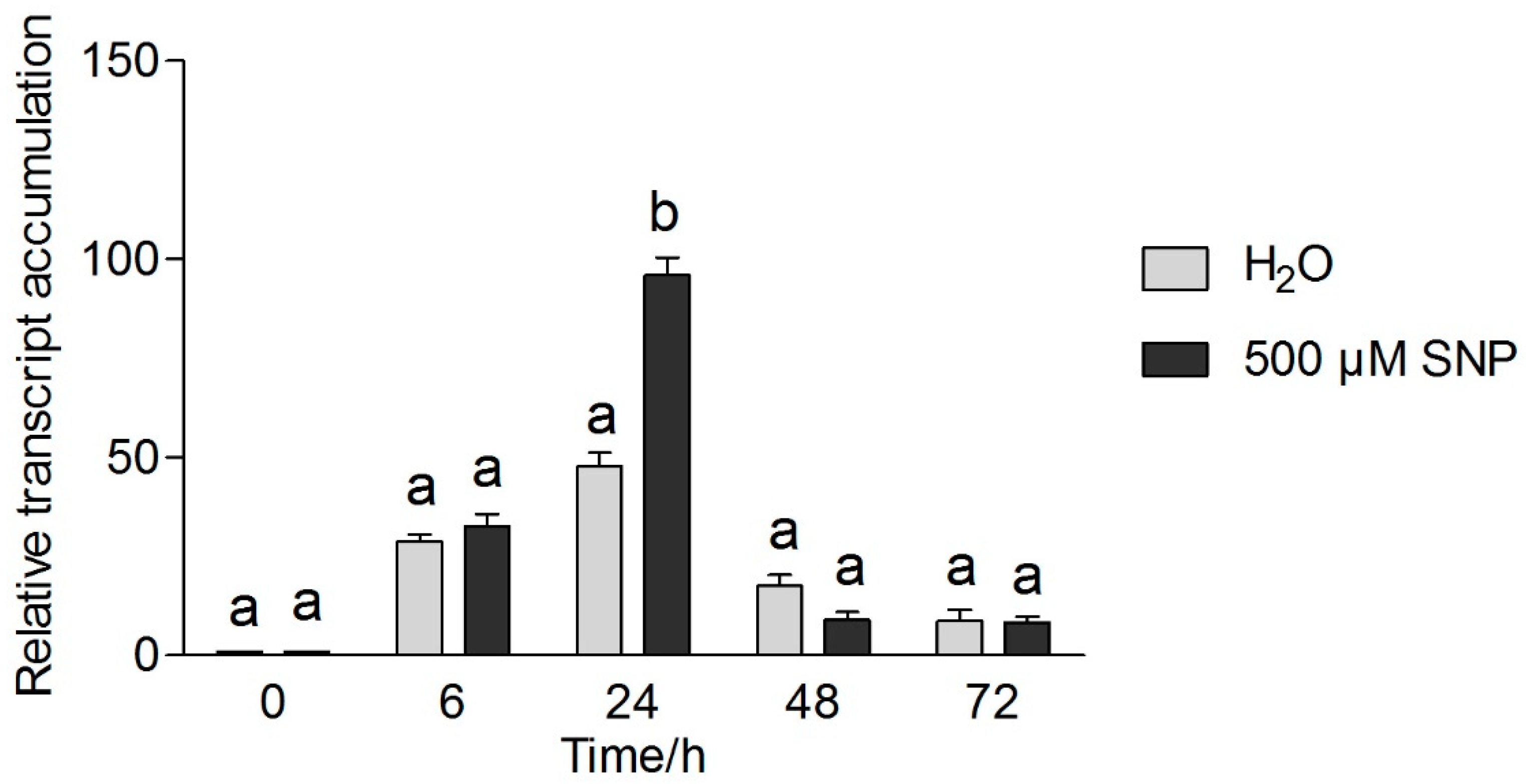
3.5. Effects of Exogenous NO and Low Temperature Stress on CsICE1 and CsCBF1 Gene Expression in Tea Leaves
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CBF | C-repeat binding factors |
| cDNA | Complementary deoxyribonucleic acid |
| cDNA-AFLP | Cdna amplified fragment length polymorphism |
| cGMP | Cyclic guanosine 5′-monophosphate |
| ICE1 | Inducer of CBF expression 1 |
| qRT-PCR | quantitative real-time polymerase chain reaction |
References
- Zhou, L.; Chen, X.; Wang, Y.H.; Zhu, X.J.; Li, X.H.; Fang, W.P. Effect of harpin on physiological characteristics of tea plant under cold stress. Acta Hortic. Sin. 2014, 41, 746–754. [Google Scholar]
- Lin, Z.H.; Zhong, Q.H.; Chen, C.S. Research progress of tea tree resistance breeding. Fujian Tea 2015, 37, 2–4. [Google Scholar]
- Yangk, Z.Q.; Han, D.; Wang, X.L.; Jin, Z.F. Changes in photosynthetic parameters and antioxidant enzymatic activity of four tea varieties during a cold wave. Acta Ecol. Sin. 2016, 36, 629–641. [Google Scholar]
- El-Mahdy, M.T.; Youssef, M.; Eissa, M.A. Impact of in vitro cold stress on two banana genotypes based on physio-biochemical Evaluation. S. Afr. J. Bot. 2018, 119, 219–225. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Q.; Ou, L.; Ji, D.; Liu, T.; Lan, R.; Li, X.; Jin, L. Response to the cold stress signaling of the tea plant (Camellia sinensis) elicited by chitosan oligosaccharide. Agron. J. 2020, 10, 915. [Google Scholar] [CrossRef]
- Wang, X.C.; Yang, Y.J. Research Progress on Resistance Breeding of Tea Plant. J. Tea Sci. 2003, 23, 94–98. [Google Scholar]
- Ritonga, F.N.; Chen, S. Physiological and Molecular Mechanism Involved in Cold Stress Tolerance in Plants. Plants 2020, 9, 560. [Google Scholar] [CrossRef] [PubMed]
- Wendehenne, D.; Courtois, C.; Besson, A. NO-based signaling in plants. In Nitric Oxide in Plant Growth, Development and Stress Physiology; Springer: Berlin, Germany, 2007; pp. 35–51. [Google Scholar] [CrossRef]
- Tian, X.; Lei, Y. Nitric oxide treatment alleviates drought stress in wheat seedlings. Biol. Plant 2006, 50, 775–778. [Google Scholar] [CrossRef]
- Zheng, C.; Jiang, D.; Liu, F.; Dai, T.; Liu, W.; Jing, Q.; Cao, W. Exogenous nitric oxide improves seed germination in wheat against mitochondrial oxidative damage induced by high salinity. Environ. Exp. Bot. 2009, 67, 222–227. [Google Scholar] [CrossRef]
- Liu, X.; Wang, L.; Liu, L.; Guo, Y.; Ren, H. Alleviating effect of exogenous nitric oxide in cucumber seedling against chilling stress. Afr. J. Biotechnol. 2011, 10, 4380–4386. [Google Scholar]
- Chen, Y.P.; Wang, X.M.; Yang, Z.J.; Yu, F.; Si, X.P. Effects of Nitric Oxide on Seed Germination and Physiological Reaction of Maize Seedlings Under Low Temperature Stress. J. Agro-Environ. Sci. 2012, 31, 270–277. [Google Scholar]
- Huang, Y.H. Experimental Techniques of Tea Science; Agricultural Publishing Housel: Beijing, China, 1995. [Google Scholar]
- Zheng, B.S. Research Techniques of Modern Plant Physiology and Biochemistry; Meteorological Publishing House: Beijing, China, 2006. [Google Scholar]
- Kong, X.S.; Yi, X.F. Experimental Techniques of Plant Physiol; Agricultural Publishing House: Beijing: China, 2008. [Google Scholar]
- Li, H.S. Principles and Techniques of Plant Physiological and Biochemical Experiments; Higher Education Press: Beijing, China, 2000. [Google Scholar]
- Zhu, Z. Screening of Physiological Indexes of Cold Resistance and Establishment of the Method for Identifying Cold Resistance in Camellia Sinensis; Agricultural University: Hefei, China, 2011. [Google Scholar]
- Abou-Zaid, E.A.; Eissa, M.A. Thompson seedless grapevines growth and quality as affected by glutamic acid, vitamin b, and algae. J. Soil Sci. Plant Nutr. 2019, 19, 725–733. [Google Scholar] [CrossRef]
- Ali, A.M.; Awad, M.Y.; Hegab, S.A.; Gawad, A.M.A.E.; Eissa, M.A. Effect of potassium solubilizing bacteria (Bacillus cereus) on growth and yield of potato. J. Plant Nutr. 2021, 44, 411–420. [Google Scholar] [CrossRef]
- Al-Sayed, H.; Hegab, S.A.; Youssef, M.; Khalafalla, M.; Almaroai, Y.A.; Ding, Z.; Eissa, M.A. Evaluation of quality and growth of roselle (Hibiscus sabdariffa L.) as affected by bio-fertilizers. J. Plant Nutr. 2021, 43, 1025–1035. [Google Scholar] [CrossRef]
- Ding, Z.; Zhou, Z.; Lin, X.; Zhao, F.; Wang, B.; Lin, F.; Ge, Y.; Eissa, M.A. Biochar impacts on NH3-volatilization kinetics and growth of sweet basil (Ocimum basilicum L.) under saline conditions. Ind. Crop. Prod. 2020, 157, 11290. [Google Scholar] [CrossRef]
- Liu, D.; Ding, Z.; Ali, E.F.; Kheir, A.M.S.; Eissa, M.A.; Ibrahim, O.H. Biochar and compost enhance soil quality and growth of roselle (Hibiscus sabdariffa L.) under saline conditions. Sci. Rep. 2021, 11, 8739. [Google Scholar] [CrossRef] [PubMed]
- Eissa, M.A.; Abeed, A.H. Growth and biochemical changes in quail bush (Atriplex lentiformis (Torr.) S. Wats) under Cd stress. Environ. Sci. Pollut. Res. 2019, 26, 628–635. [Google Scholar] [CrossRef]
- Xu, H.L. Studies on Alleviate Effects of Nitric Oxide on Salt Stress and Low Temperature Stress of Cucumber; Agricultural University: Harbin, China, 2006. [Google Scholar]
- Ma, X.L.; Wei, X.H.; Long, R.J.; Cui, W.J.; Wan, Y.L. Studies on mechanism of enhancing the chilling resistance of annual ryegrass by exogenous nitric oxide. Acta Ecol. Sin. 2005, 25, 1270–1274. [Google Scholar]
- Fan, H.F.; Du, C.X.; Zhu, Z.J. Effects of exogenous nitric oxide on plant growth, membrane lipid peroxidation and photosynthesis in cucumber seedling leaves under low temperature. Acta Agric. Zhejiangensis 2011, 23, 538–542. [Google Scholar]
- Zhang, G. Review on Methods for Measuring Frost Hardiness in Woody Plants Abroad. World For. Res. 2005, 5, 14–20. [Google Scholar]
- Zhao, M.G.; Chen, L.; Zhang, L.L.; Zhang, W.H. Nitric reductase-dependent nitric oxide production is involved in cold acclimation and freezing tolerance in Arabidopsis. Plant Physiol. 2009, 151, 755–767. [Google Scholar] [CrossRef] [Green Version]
- Li, X. The Effects of Nitric Oxide on Cold Response and Polyamine Synthesis of Poncirus Trifoliata; Agricultural University: Wuhan, China, 2012. [Google Scholar]
- Wu, X.H.; Lv, C.M.; Feng, J.H. Protective effect of exogenous nitric oxide against oxidative damage in pumpkin seedlings under chilling stress. Acta Prataculturae Sin. 2016, 25, 161–169. [Google Scholar]
- Guo, J.W.; Shang, S.; Mu, D.W.; Tian, L.B. Effects of Exogenous Nitric Oxide on Growth and Physiological Characteristics of Asparagus Bean Deedlings Under Cold Stress. Chin. J. Trop. Crop. 2015, 36, 2179–2183. [Google Scholar]
- Liu, X.W. Mechanisms of Nitric Oxide in the Regulation of Chilling Stress Tolerance in Cucumber; Agricultural University: Beijing, China, 2013. [Google Scholar]
- Wang, X.Y.; Shen, W.B.; Xu, L.L. Exogenous Nitric Oxide Alleviates Osmotic Stress-induced Membrane Lipid Peroxidation in Wheat Seedling Leaves. Acta Photophysiol. Sin. 2004, 30, 195–200. [Google Scholar]
- Beligni, M.V.; Fath, A.; Bethke, P.C.; Lamattina, L.; Jonesm, R.L. Nitric oxide acts as an antioxidant and delays programmed cell death in barley aleurone layer. Plant Physiol. 2002, 129, 1642–1645. [Google Scholar] [CrossRef] [Green Version]
- Chinnusamy, V.; Ohta, M.; Kanrar, S.; Lee, B.H.; Hong, X.; Agarwal, M.; Zhu, J.K. ICE1: A Regulator of Cold-Induced Transcriptome and Freezing Tolerance in Arabidopsis. Genes Dev. 2003, 17, 1043–1054. Available online: http://www.genesdev.org/cgi/doi/10.1101/gad.1077503 (accessed on 10 October 2020). [CrossRef] [PubMed] [Green Version]
- Chang, X. Study on Function Analysis and Regulation Pathways of CsCBF1from Tea Plant (Camellia sinensis) under Low Temperature Stress; Agricultural University: Hefei, China, 2012. [Google Scholar]
- Xiang, D.J. Cloning of ICE1-Like Transcription Factors and Assessment of the Cold Resistance in Transgenic Rice; Agricultural University: Shenyang, China, 2012. [Google Scholar]
- Wang, Y. CsICE1 and CsCBF1: Two Transcription Factors Involved in Cold Responses in Camellia Sinensis; Agricultural University: Hefei, China, 2011. [Google Scholar]
- Zhang, S.M. The Effect of Nitric Oxide and Hydrogen Peroxide on the Growth of Rice Seedlings under Salt Stress and Its Mechanism; Yangzhou University: Yangzhou, China, 2006. [Google Scholar]
- Zou, Y.F. Roles of Reactive Oxygen Species in MAPKs of Cells Exposed to Selenite; University of Science and Technology: Wuhan, China, 2007. [Google Scholar]
- Feng, Y.; Fu, X.; Han, L.; Xu, C.; Liu, C.; Bi, H.; Ai, X. Nitric Oxide Functions as a Downstream Signal for Melatonin-Induced Cold Tolerance in Cucumber Seedlings. Front. Plant Sci. 2021, 12, 686545. [Google Scholar] [CrossRef]
- Pagnussat, G.C.; Lanteri, M.L.; Lombardo, M.C.; Lamattina, L. Nitrie oxide mediates the indole acetic acid induction activation of amitogen-activated protein kinase caseade involved inadventitious root development. Plant Physiol. 2004, 135, 279. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| CsICE1 | ATGTTTTGTAGCCGCAGAC | GCTTTGATTTGGTCAGGATG |
| CsCBF1 | AGAAATCGGATGGCTTGTGT | TTGTCGTCTCAGTCGCAGTT |
| GAPDH | TTGGCATCGTTGAGGGTCT | CAGTGGGAACACGGAAAGC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Yu, Q.; Li, Y.; Li, J.; Chen, J.; Liu, Z.; Huang, J.; Al-Harbi, M.S.; Ali, E.F.; Eissa, M.A. Mechanisms of Nitric Oxide in the Regulation of Chilling Stress Tolerance in Camellia sinensis. Horticulturae 2021, 7, 410. https://doi.org/10.3390/horticulturae7100410
Wang Y, Yu Q, Li Y, Li J, Chen J, Liu Z, Huang J, Al-Harbi MS, Ali EF, Eissa MA. Mechanisms of Nitric Oxide in the Regulation of Chilling Stress Tolerance in Camellia sinensis. Horticulturae. 2021; 7(10):410. https://doi.org/10.3390/horticulturae7100410
Chicago/Turabian StyleWang, Yingzi, Qin Yu, Yinhua Li, Juan Li, Jinhua Chen, Zhonghua Liu, Jian’an Huang, Mohammad S. Al-Harbi, Esmat F. Ali, and Mamdouh A. Eissa. 2021. "Mechanisms of Nitric Oxide in the Regulation of Chilling Stress Tolerance in Camellia sinensis" Horticulturae 7, no. 10: 410. https://doi.org/10.3390/horticulturae7100410