
The Effect of the Daily Light Integral and Spectrum on Mesembryanthemum crystallinum L. in an Indoor Plant Production Environment

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Growing Condition
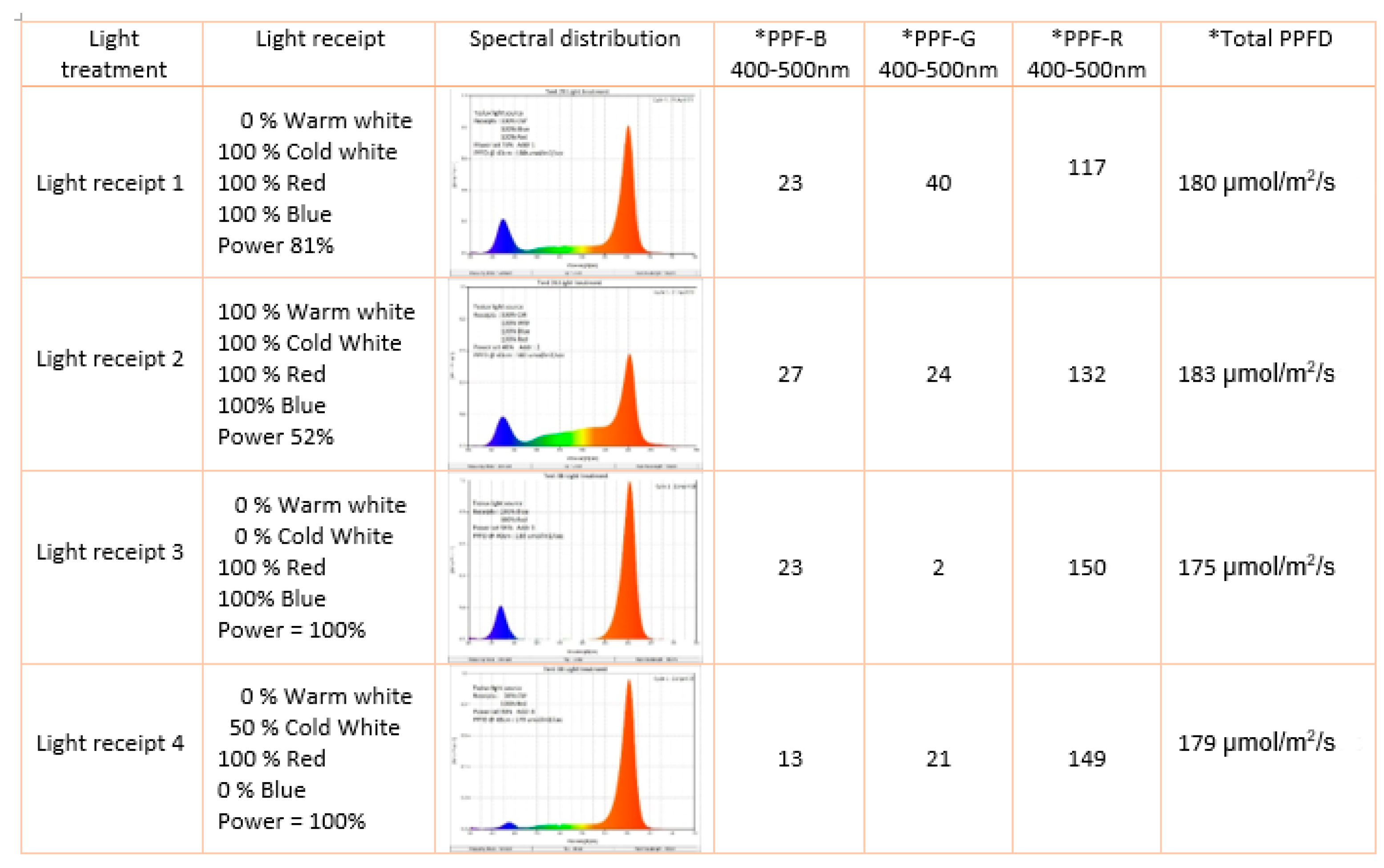
2.2. Light Treatment
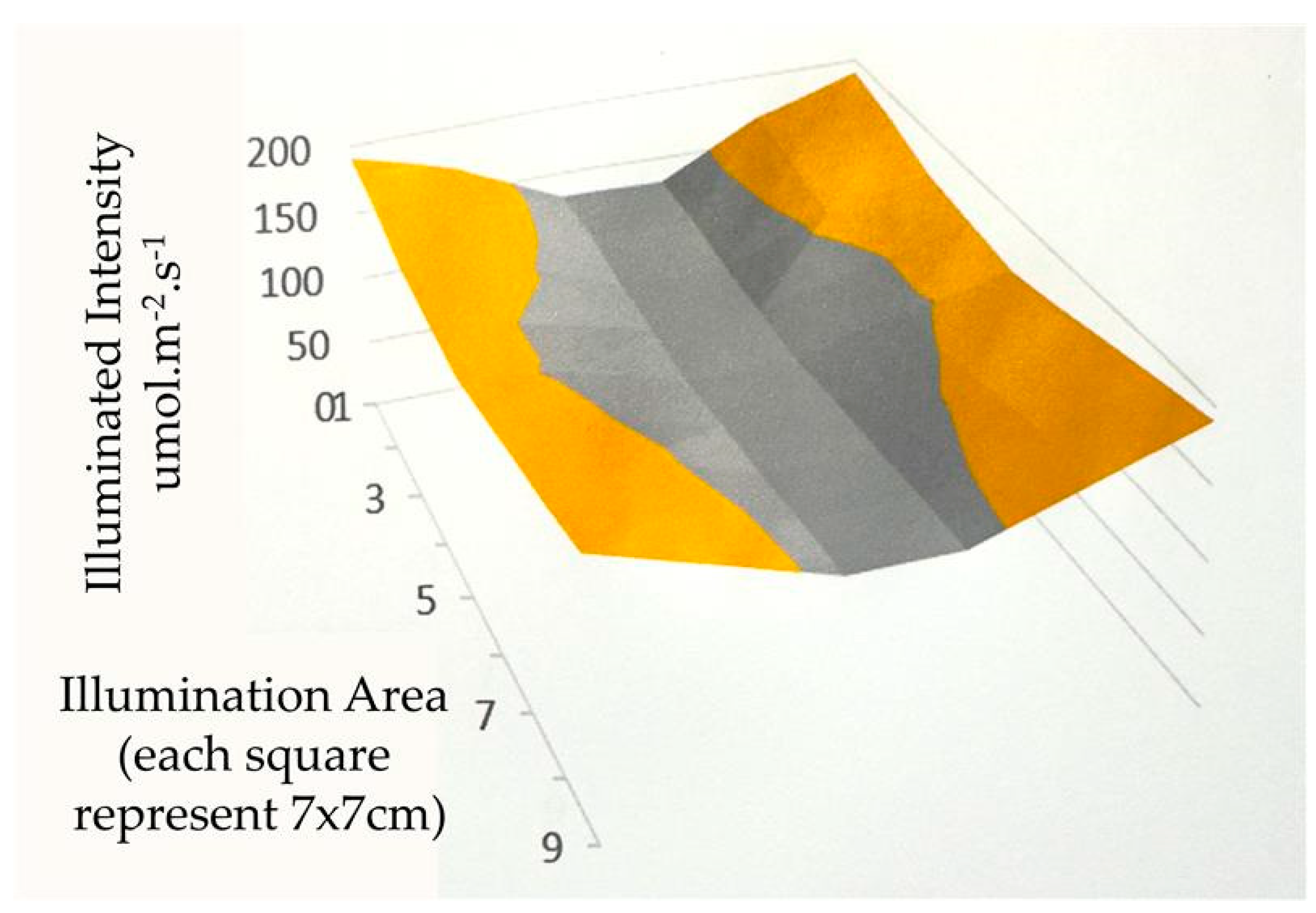
2.3. Homogeneity of the Light Distribution
2.4. Testing Parameters
2.4.1. Sampling and Statistical Analysis
2.4.2. Growth Characteristics
2.4.3. Dry Matter
2.4.4. Total Antioxidant Capacity
2.4.5. Vitamin C
2.4.6. Chlorophyll and Carotenoids
3. Results
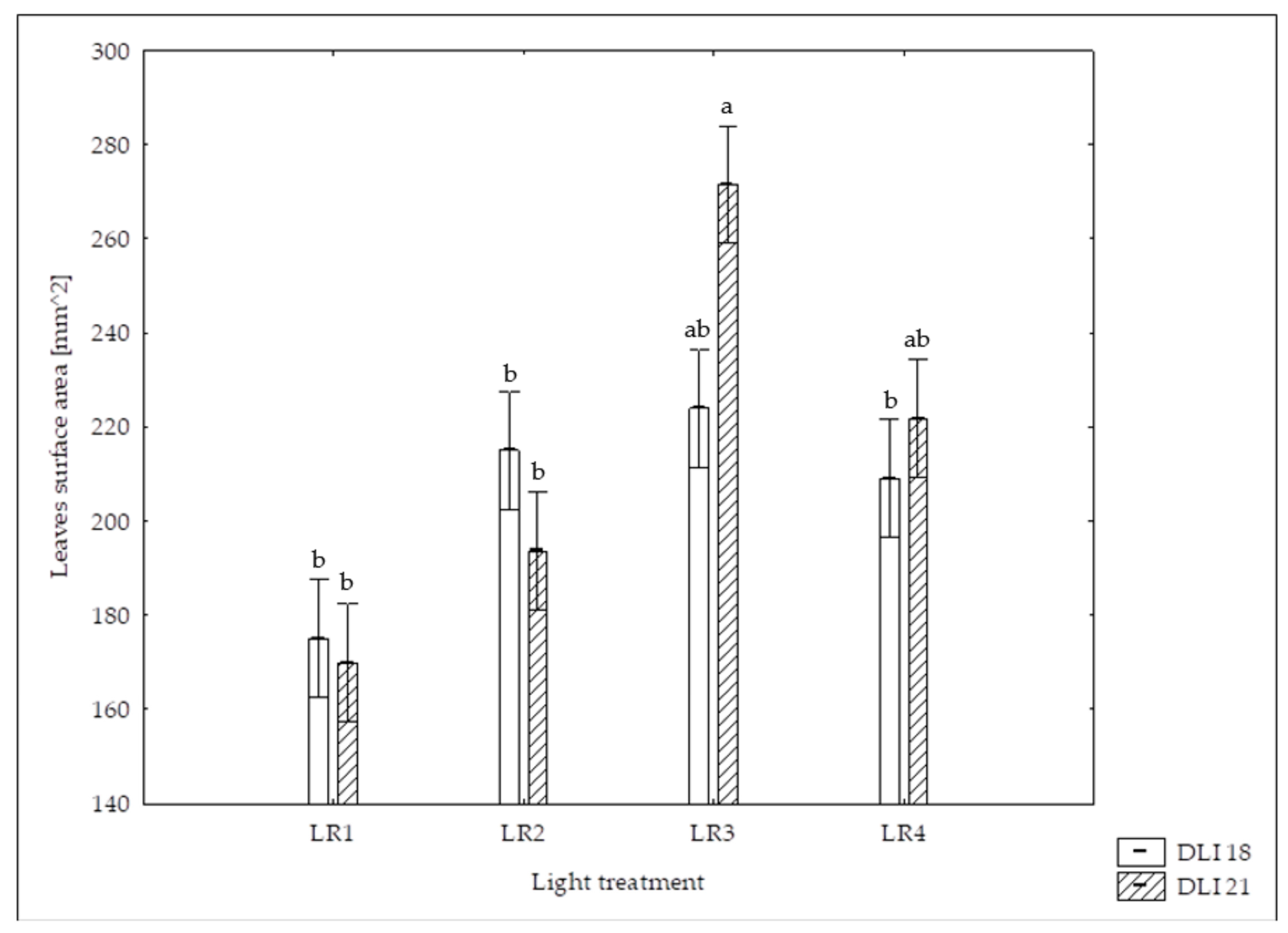
3.1. Morphological Parameters
3.2. Dry Matter
3.3. Total Antioxidant Capacity
3.4. Vitamin C
3.5. Chlorophyll a and b
3.6. Total Carotenoids
3.7. Nitrates
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, J.; Lu, W.; Tong, Y.; Yang, Q. Leaf morphology, photosynthetic performance, chlorophyll fluorescence, stomatal development of lettuce (Lactuca sativa L.) exposed to different ratios of red light to blue light. Front. Plant Sci. 2016, 7, 250. [Google Scholar] [CrossRef]
- Sale, A.I.; Uthairatanakij, A.; Laohakunjit, N.; Jitareerat, P.; Kaisangsri, N. Pre-harvest supplemental LED treatments led to improved postharvest quality of sweet basil leaves. J. Photochem. Photobiol. B Biol. 2023, 248, 112788. [Google Scholar] [CrossRef]
- Pattison, M.; Hansen, M.; Bardsley, N.; Elliott, C.; Lee, K.; Pattison, L.; Tsao, J. Lighting R&D Opportunities; US Department of Energy: Washington, DC, USA, 2020. [CrossRef]
- Larsen, D.H.; Woltering, E.J.; Nicole, C.C.S.; Marcelis, L.F.M. Response of Basil Growth and Morphology to Light Intensity and Spectrum in a Vertical Farm. Front. Plant Sci. 2020, 11, 597906. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.H.; Wheeler, R.M.; Sager, J.C.; Goins, G.D.; Norikane, J.H. Evaluation of lettuce growth using supplemental green light with red and blue light-emitting diodes in a controlled environment: A review of research at Kennedy space center. Acta Hortic. 2006, 711, 111–119. [Google Scholar] [CrossRef]
- Li, Q.; Kubota, C. Effects of supplemental light quality on growth and phytochemicals of baby leaf lettuce. J. Environ. Exp. Bot. 2009, 67, 59–64. [Google Scholar] [CrossRef]
- Colonna, E.; Rouphael, Y.; Barbieri, G.; De Pascale, S. Nutritional quality of ten leafy vegetables harvested at two light intensities. Food Chem. 2016, 199, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Snowden, M.C.; Cope, K.R.; Bugbee, B. Sensitivity of seven diverse species to blue and green light: Interactions with photon flux. PLoS ONE 2016, 11, e0163121. [Google Scholar] [CrossRef] [PubMed]
- Cope, K.R.; Bugbee, B. Spectral effects of three types of white light emitting diodes on plant growth and development: Absolute versus relative amounts of blue light. HortScience 2013, 48, 504–509. [Google Scholar] [CrossRef]
- Zhang, H.; Tu, Y.; Kang, J.; Song, W.; Zheng, L. Blue light dosage affects photosynthesis, chlorophyll, and antioxidant properties of Mesembryanthemum crystallinum. Photosynthetica 2021, 59, 547–556. [Google Scholar] [CrossRef]
- Castilla, N. Greenhouse Technology and Management, 2nd ed.; CABI: Wallingford, UK, 2013; ISBN 1780641036. [Google Scholar]
- Durmus, D. Real-Time Sensing and Control of Integrative Horticultural Lighting Systems. J 2020, 3, 266–274. [Google Scholar] [CrossRef]
- Morrow, R.C. Led lighting in horticulture. HortScience 2008, 43, 1947–1950. [Google Scholar] [CrossRef]
- Xia, J.Q.; Mattson, N. Response of Common Ice Plant (Mesembryanthemum crystallinum L.) to Sodium Chloride Concentration in Hydroponic Nutrient Solution. HortScience 2022, 57, 750–756. [Google Scholar] [CrossRef]
- Lee, J.W.; Son, K.H.; Lee, J.H.; Kim, Y.J. Growth and biochemical responses of ice plant irradiated by various visible light spectra in plant factories. HortScience Technol. 2019, 37, 598–608. [Google Scholar] [CrossRef]
- Lee, B.H.; Lee, C.C.; Wu, S.C. Ice plant (Mesembryanthemum crystallinum) improves hyperglycaemia and memory impairments in a Wistar rat model of streptozotocin-induced diabetes. J. Sci. Food Agric. 2014, 94, 2266–2273. [Google Scholar] [CrossRef] [PubMed]
- Herppich, W.B.; Huyskens-Keil, S.; Schreiner, M. Effects of saline irrigation on growth, physiology and quality of Mesembryanthemum crystallinum L., a rare vegetable crop. J. Appl. Bot. Food Qual. 2008, 82, 47–54. [Google Scholar]
- Zbíral, J. Analysis of Plant Material; Czech National Reference Laboratory, UKZUZ: Brno, Czech Republic, 2005. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Wu, M. A quick method for the simultaneous determination of ascorbic acid and sorbic acid in fruit juices by capillary zone electrophoresis. Talanta 2005, 65, 794–798. [Google Scholar] [CrossRef] [PubMed]
- Holm-Hansen, O.; Bo, R. Chlorophyll a Determination: Improvements in Methodology. Oikos 1978, 30, 438–447. [Google Scholar] [CrossRef]
- Son, K.H.; Oh, M.M. Growth, photosynthetic and antioxidant parameters of two lettuce cultivars as affected by red, green and blue light emitting diode. Korean J. Hortic. Sci. Technol. 2015, 30, 664–672. [Google Scholar] [CrossRef]
- Young, J.K.; Hye, M.K.; Hyeon, M.K.; Hye, R.L.; Byoung, R.J.; Hyeon, J.L.; Hyun, J.K.; Seung, J.H. Growth and phytochemicals of ice plant (Mesembryanthemum crystallinum L.) as affected by various combined ratios of red and blue LEDs in a closed-type plant production system. J. Appl. Res. Med. Aromat. Plants 2021, 20, 100267. [Google Scholar]
- Nishimura, T.; Ohyama, K.; Goto, E.; Inagaki, N. Concentrations of perillaldehyde, limonene, and anthocyanin of Perilla plants as affected by light quality under controlled environments. Sci. Hortic. 2009, 122, 134–137. [Google Scholar] [CrossRef]
- Johkan, M.; Shoji, K.; Goto, F.; Hashida, S.; Yoshihara, T. Blue light-emitting diode light irradiation of seedlings improves seedling quality and growth after transplanting in red leaf lettuce. HortScience 2010, 45, 1809–1814. [Google Scholar] [CrossRef]
- Shimizu, H.; Saito, Y.; Nakashima, H.; Miyasaka, J.; Ohdoi, K. Light environment optimization for lettuce growth in plant factory. In Proceedings of the 18th IFAC World Congress, Milano, Italy, 28 August–2 September 2011; pp. 605–609. [Google Scholar]
- Xia, J.; Mattson, N. Response of Common Ice Plant (Mesembryanthemum crystallinum L.) to Photoperiod/Daily Light Integral in Vertical Hydroponic Production. Horticulturae 2022, 8, 653. [Google Scholar] [CrossRef]
- Liu, K.; Gao, M.; Jiang, H.; Ou, S.; Li, X.; He, R.; Li, Y.; Liu, H. Light Intensity and Photoperiod Affect Growth and Nutritional Quality of Brassica Microgreens. Molecules 2022, 27, 883. [Google Scholar] [CrossRef] [PubMed]
- Kurucz, E.; Antal, G.; Kincses, I.; Sipos, M.; Miklós, G.F.; Holb, I.J. Effect of Light Treatment and Maturity Stage on Biomass Production and Bioactive Compounds of Two Pepper Cultivars under a Deep Water Culture Hydroponic System. Sustainability 2023, 15, 13205. [Google Scholar] [CrossRef]
- Samuoliene, G.; Sirtautas, R.; Brazaityte, A.; Duchovskis, P. LED lighting and seasonality effects antioxidant properties of baby leaf lettuce. Food Chem. 2012, 134, 1494–1499. [Google Scholar] [CrossRef] [PubMed]
- Barber, J.; Andersson, B. Too much of a good thing: Light can be bad for photosynthesis. Trends Biochem. Sci. 1992, 17, 61–66. [Google Scholar] [CrossRef]
- Muller, P.; Li, X.P.; Niyogi, K.K. No photochemical quenching. A response to excess light energy. Plant Physiol. 2001, 125, 1558–1566. [Google Scholar] [CrossRef]
- Abdullah, A.A.; Abdulaziz, S.A.; Ibrahim, M.A.; Hail, Z.R.; Michael, P.F.; Abdullah, A.A.; Abdullah, A.I. The Effect of LED Light Spectra on the Growth, Yield and Nutritional Value of Red and Green Lettuce (Lactuca sativa). Plants 2023, 12, 463. [Google Scholar] [CrossRef]
- Xu, D.Q.; Shen, Y.K. Light stress: Photoinhibition of photosynthesis in plants under natural conditions. In Handbook of Plant and Crop Stress, 4th ed.; Mohammad, P., Ed.; Marcel Dekker, Inc.: New York, NY, USA, 1999; pp. 483–497. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Light Receipt 1 | Light Receipt 2 | Light Receipt 3 | Light Receipt 4 | |||||
|---|---|---|---|---|---|---|---|---|
| Morphological Parameters | DLI 18 h | DLI 21 h | DLI 18 h | DLI 21 h | DLI 18 h | DLI 21 h | DLI 18 h | DLI 21 h |
| Plant shoot weight [g] | 39.3 (±4.5) eE | 37.1 (±10.8) eE | 37.1 (±5.5) eE | 42.0 (±17.6) deDE | 68.0 (±5.9) aA | 64.2 (±10.0) abA | 57.2 (±6.7) bcBC | 52.0 (±9.9) cdCD |
| Leaves surface area [cm2] | 175.0 (±46.7) bB | 169.9 (±35.3) bB | 215.0 (±38.2) bAB | 193.6 (±78.1) bB | 224.0 (±41.8) abA | 271.5 (±33.8) aA | 209.0 (±60.9) bB | 221.7 (±49.5) abAB |
| Leaves width [mm] | 23.7 (±2.5) aAB | 21.9 (±2.9) cB | 25.1 (±2.5) aAB | 23.8 (±4.0) bcB | 24.6 (±2.6) aAB | 28.0 (±1.9) aA | 24.1 (±3.8) aAB | 26.9 (±1.9) abB |
| Plant Shoot Weight [g] | Leaves Surface Area [cm2] | Leaves Width [mm] | Dry Matter [%] | TAC [mg/kg] | Vitamin C [mg/kg] | Chlorophyll a [mg/kg] | Chlorophyll b [mg/kg] | Total Carotenoids [mg/kg] | Nitrates [mg/kg] | |
|---|---|---|---|---|---|---|---|---|---|---|
| Light treatment | 0.565 * | 0.338 * | 0.342 * | 0.009 | −0.562 * | −0.314 * | −0.245 | −0.142 | −0.237 | 0.251 |
| DLI | −0.052 | 0.074 | 0.122 | 0.712 * | 0.547 * | −0.086 | −0.818 * | −0.941 * | −0.797 * | 0.752 * |
| Light Receipt 1 | Light Receipt 2 | Light Receipt 3 | Light Receipt 4 | |||||
|---|---|---|---|---|---|---|---|---|
| Phytochemical Parameters | DLI 18 h | DLI 21 h | DLI 18 h | DLI 21 h | DLI 18 h | DLI 21 h | DLI 18 h | DLI 21 h |
| Dry matter [%] | 3.4 (±0.3) bcdCDE | 3.9 (±0.5) abABC | 3.1 (±0.2) cdE | 3.7 (±0.6) abcBCD | 2.9 (±0.3) dE | 4.3 (±0.3) aA | 3.2 (±0.4) bcdDE | 4.0 (±0.4) abAB |
| TAC [mg/kg] | 73.4 (±24.7) aC | 151.0 (±10.8) aA | 68.2 (±10.9) aC | 114.3 (±30.3) abB | 57.9 (±13.7) aC | 72.8 (±10.8) bC | 53.4 (±25.7) aC | 69.8 (±13.9) bC |
| Vitamin C [mg/kg] | 83.4 (±19.2) aBC | 105.8 (±16.3) aA | 72.2 (±11.6) aC | 79.1 (±10.3) abC | 102.2 (±20.0) aAB | 64.0 (±9.7) bC | 76.3 (±19.4) aC | 71.7 (±8.0) abC |
| Chlorophyll a [mg/kg] | 160.1 (±21.2) aA | 120.3 (±9.1) bcB | 144.5 (±18.9) abA | 111.4 (±17.2) cBC | 152.5 (±9.9) aA | 98.9 (±10.0) cC | 147.0 (±14.6) abA | 95.3 (±13.7) cC |
| Chlorophyll b [mg/kg] | 63.3 (±8.3) aA | 29.9 (±4.0) aB | 62.5 (±9.6) aA | 26.3 (±8.1) aB | 59.9 (±2.4) aA | 23.9 (±2.2) aB | 58.0 (±7.7) aA | 20.8 (±4.9) aB |
| Total carotenoids [mg/kg] | 49.7 (±5.2) aA | 38.7 (±2.8) bcB | 43.7 (±7.1) abAB | 30.5 (±5.1) cdC | 45.0 (±3.8) abA | 28.5 (±2.9) dC | 45.1 (±3.3) abA | 31.6 (±4.2) cdC |
| Nitrates [mg/kg] | 325.0 (±224.4) aE | 994.3 (±406.4) aCD | 607.5 (±358.9) aDE | 1161.0 (±391.7) aBC | 250.0 (±157.0) aE | 1579.4 (±359) aAB | 334.4 (±82.4) aE | 2020.0 (±782.9) aA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.W.; Patloková, K.; Pokluda, R. The Effect of the Daily Light Integral and Spectrum on Mesembryanthemum crystallinum L. in an Indoor Plant Production Environment. Horticulturae 2024, 10, 266. https://doi.org/10.3390/horticulturae10030266
Chen JW, Patloková K, Pokluda R. The Effect of the Daily Light Integral and Spectrum on Mesembryanthemum crystallinum L. in an Indoor Plant Production Environment. Horticulturae. 2024; 10(3):266. https://doi.org/10.3390/horticulturae10030266
Chicago/Turabian StyleChen, Jun Wei, Kateřina Patloková, and Robert Pokluda. 2024. "The Effect of the Daily Light Integral and Spectrum on Mesembryanthemum crystallinum L. in an Indoor Plant Production Environment" Horticulturae 10, no. 3: 266. https://doi.org/10.3390/horticulturae10030266