
The Effects of Mixed Foliar Nutrients of Calcium and Magnesium on the Major Bypass Respiratory Pathways in the Pulp of ‘Feizixiao’ Litchi


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Experimental Design Method
2.3. Field Treatment and Sample Collection Methods
2.4. The Assay Methods of the Indicators
2.4.1. Determination of the Pulp’s Conventional Quality
2.4.2. Measurement of Pulp Respiration Rate
2.5. Transcriptome Analysis and Fluorescence Quantitative PCR Validation
2.5.1. Differential Gene Screening Analysis
2.5.2. Primer Design and qRT-PCR Validation
2.6. Data Processing and Analysis
3. Results and Analysis
3.1. Effects of Foliar Mixed Nutrient of Calcium and Magnesium on Pericarp Colouration and the Contents of Sugar and Acid in Pulp
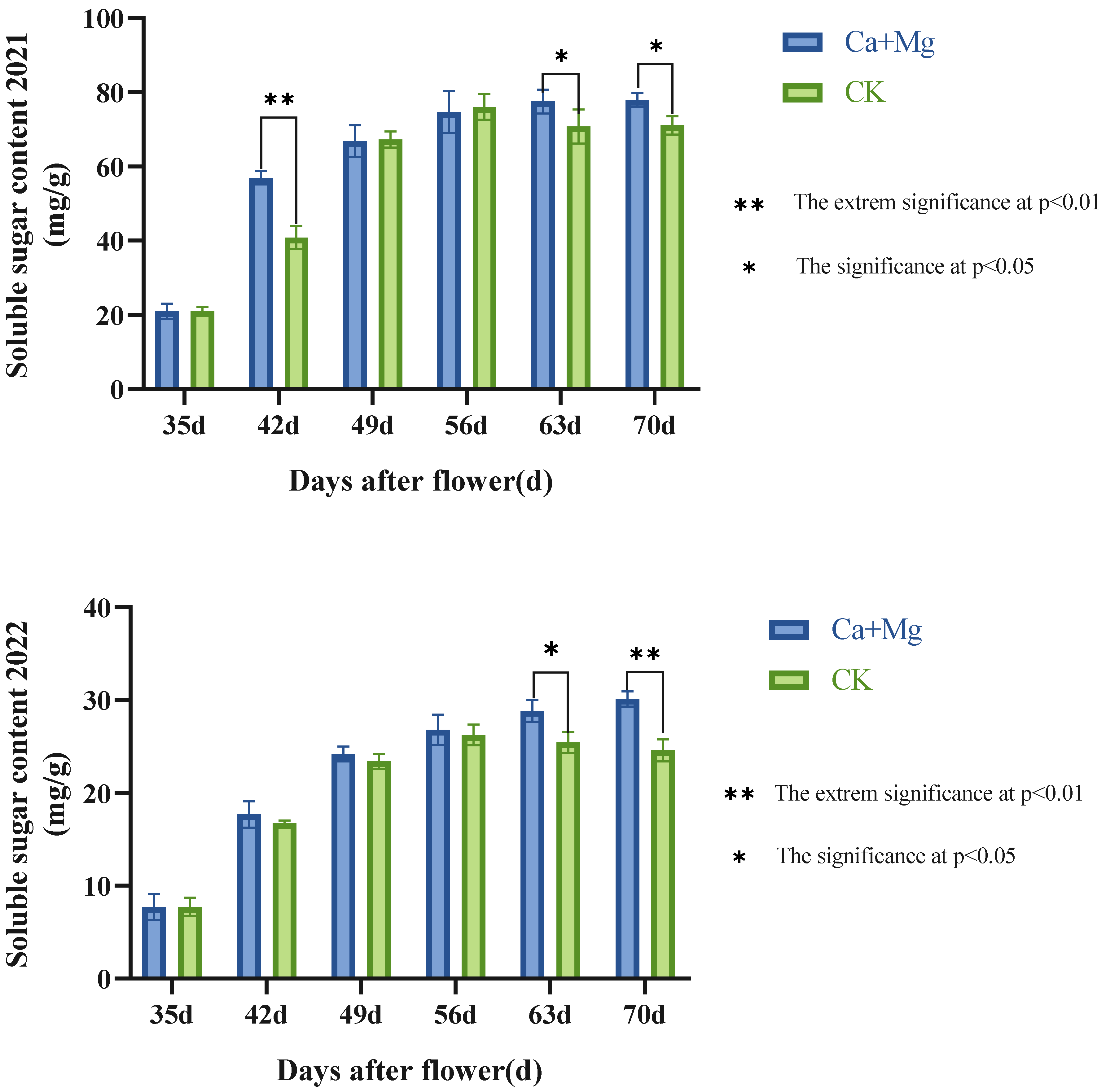
3.1.1. Change Trend of the Content of Soluble Sugar in Pulp
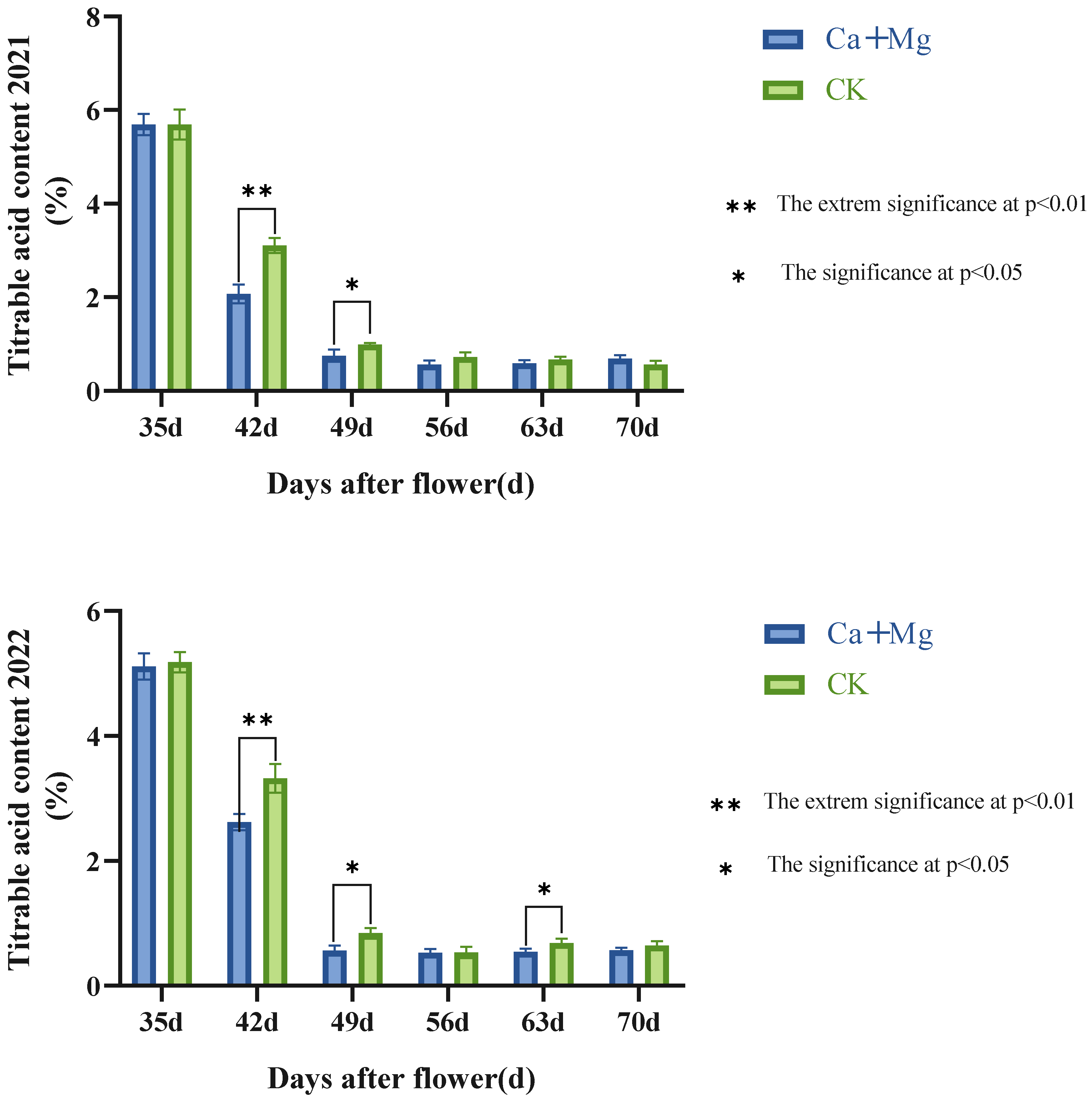
3.1.2. Chang Trends of the Content of Titratable Acid in Pulp
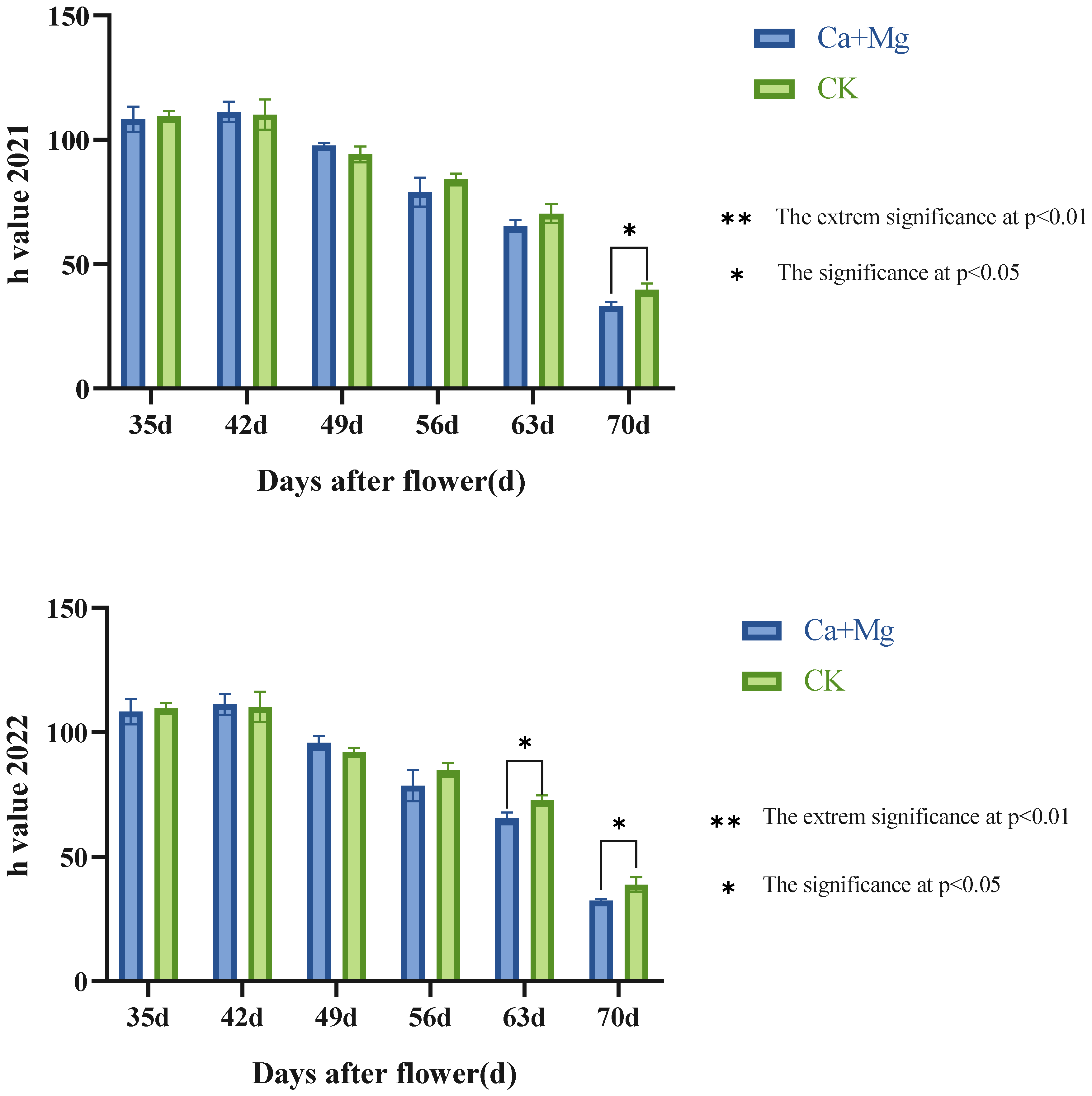
3.1.3. Effect of Foliar Mixed Nutrient of Calcium and Magnesium on the Coloration of Litchi Pericarp
3.2. Effects of Mixed Nutrient of Calcium and Magnesium on the Respiration Rate of the Bypass Pathway
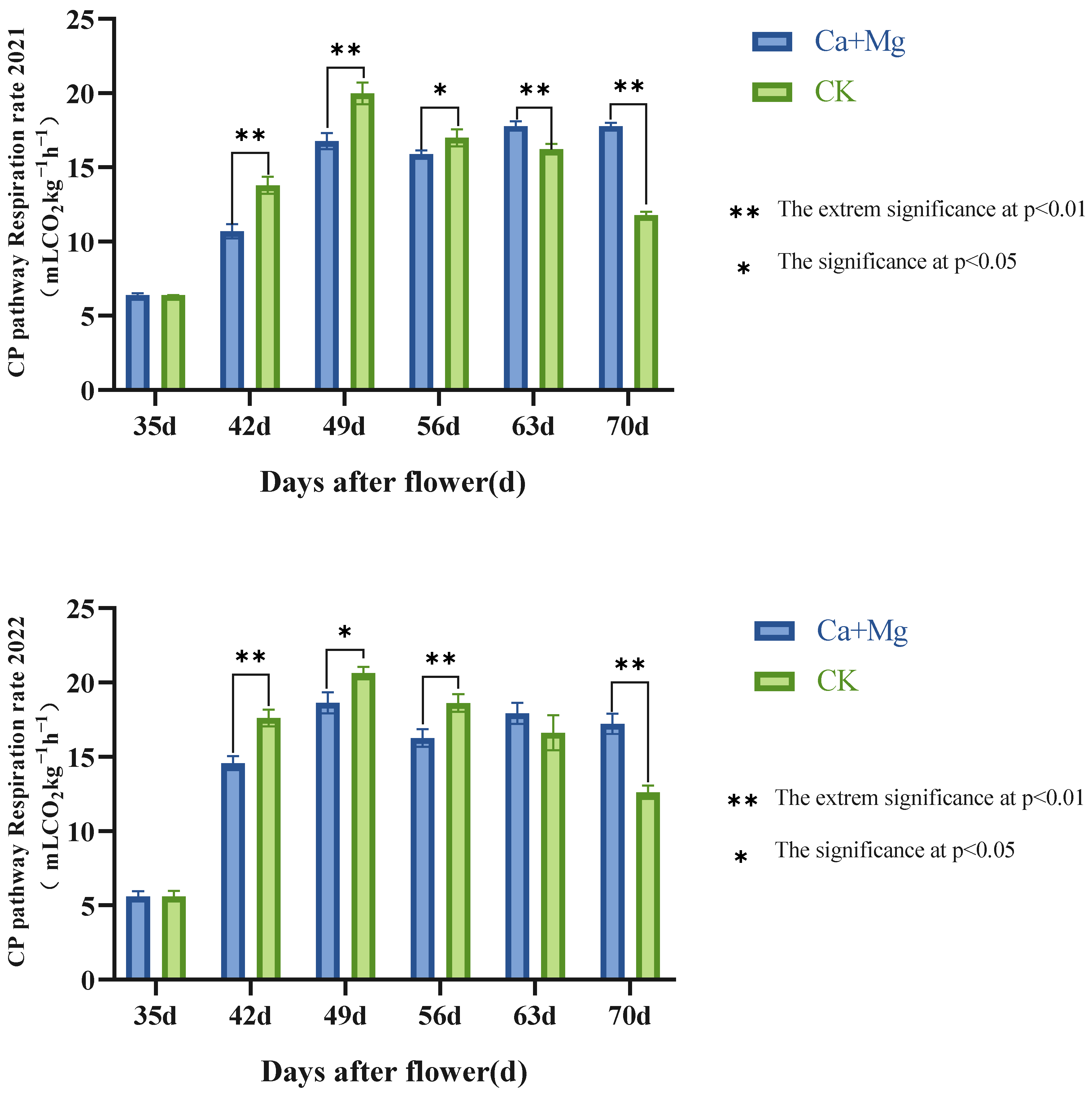
3.2.1. Trends in the Cytochrome Pathway (CP) Respiration Rates
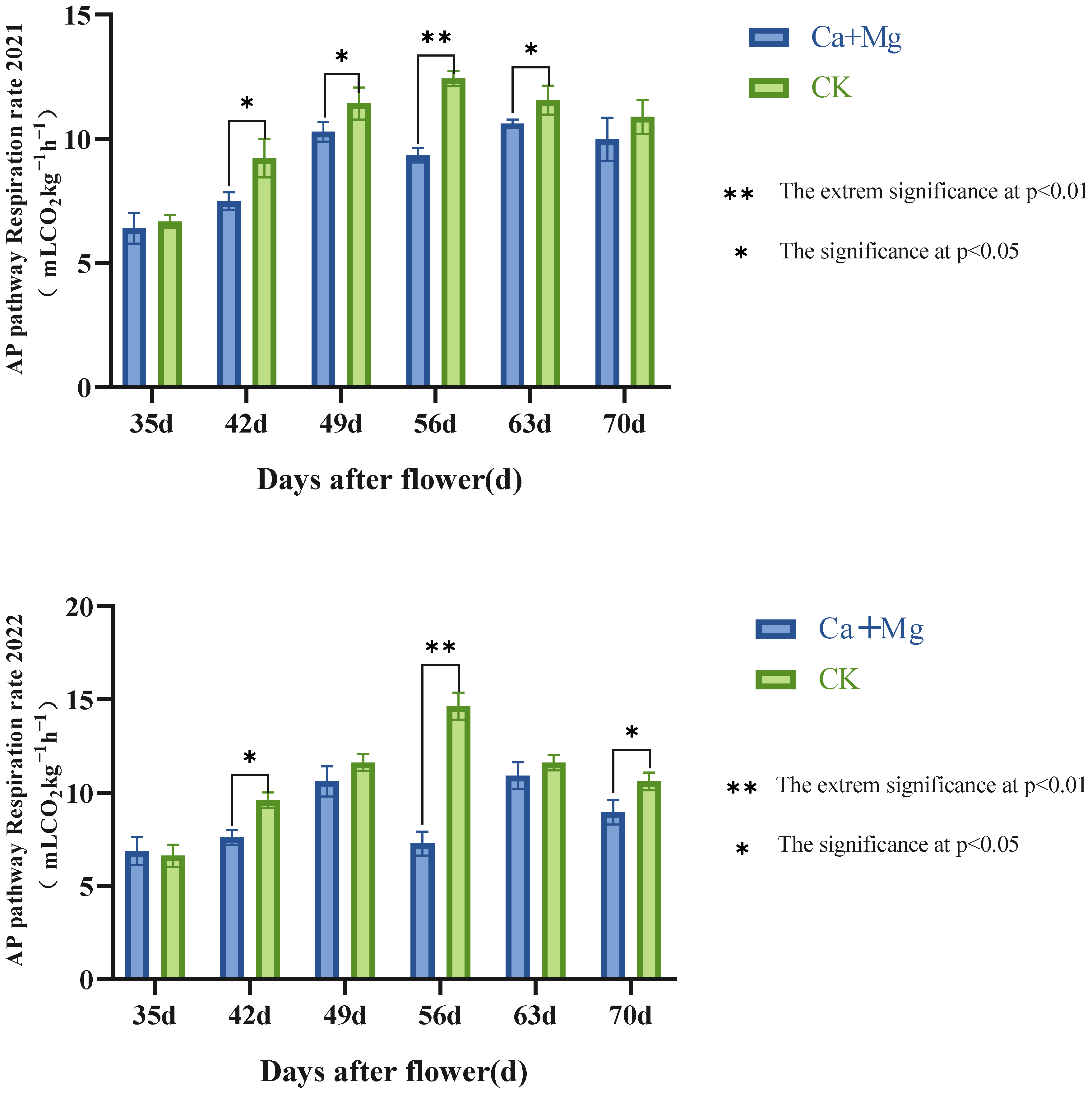
3.2.2. Trends in the Cyanide-Resistant Pathway (AP) Respiratory Rates
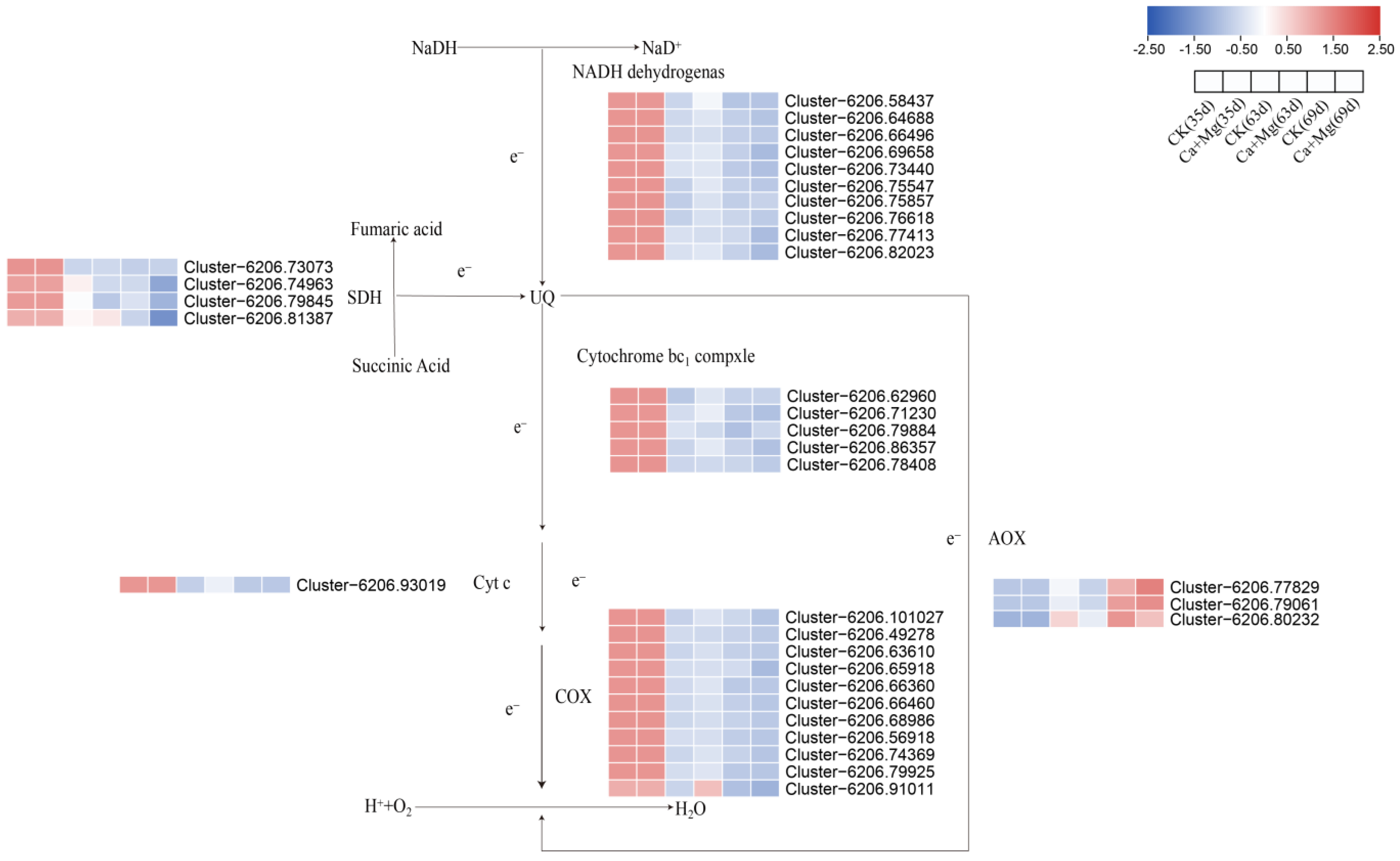
3.3. Differential Gene Screening
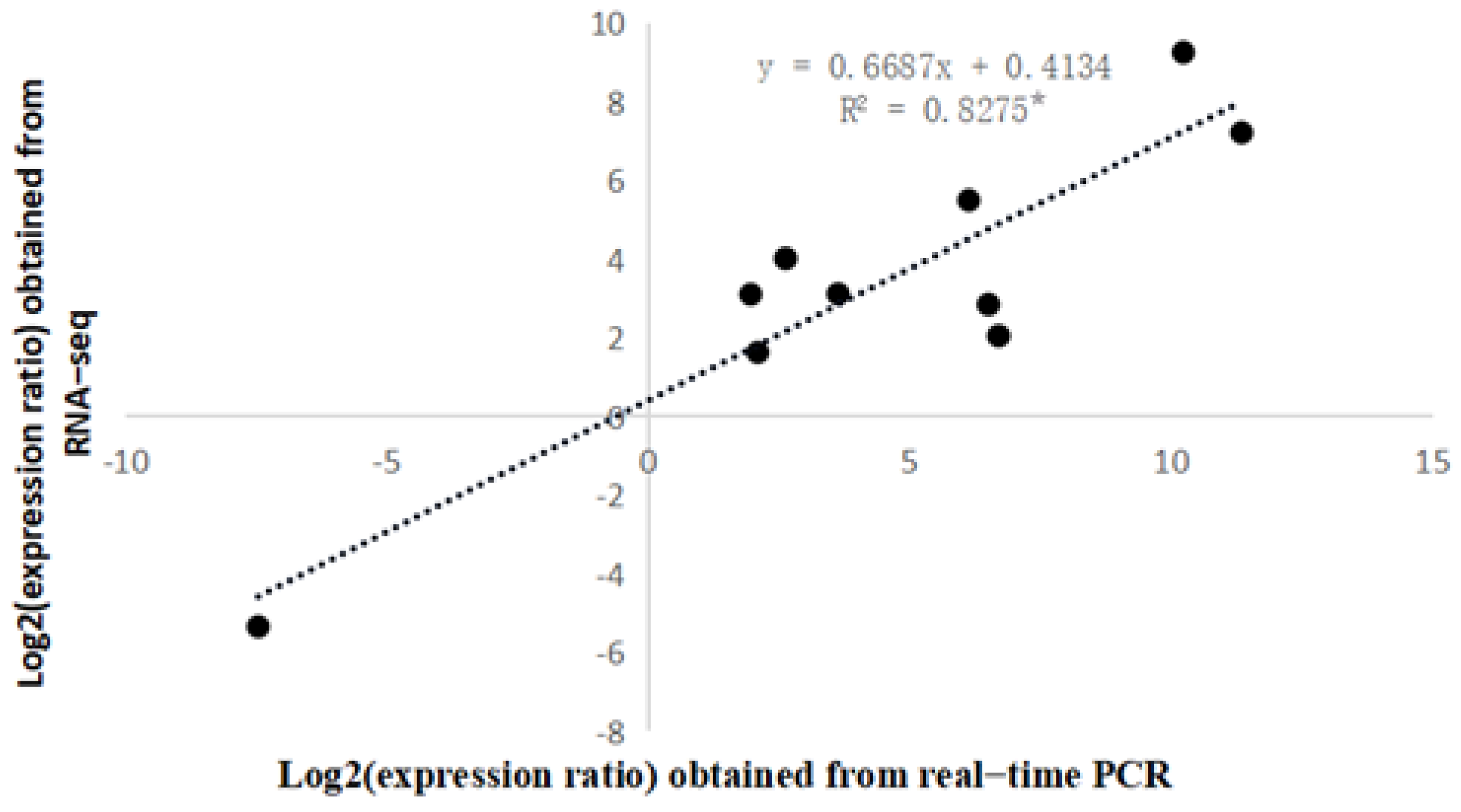
3.4. Real-Time Fluorescence Quantitative PCR Verification
4. Discussion
4.1. Effect of Different Treatments on ‘Feizixiao’ Litchi Fruit Quality
4.2. Effects of Different Treatments on the Respiration Rate of the Pulp Electron Transport Chain Pathway
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lv, R.; Liang, C.-h.; Zou, H.-p.; Li, W.-g.; Bai, R. Research on Fine Agricultural Climate Zoning of Feizixiao Litchi in Hainan. Trop. Agric. Sci. 2021, 12, 117–122. (In Chinese) [Google Scholar]
- Wang, H.-c.; Huang, X.-m.; Huang, H.-b. A study on the reasons for poor coloring of Feizixiao’s lychee fruit. J. Hortic. 2002, 5, 408–412. (In Chinese) [Google Scholar]
- Wang, H.-c.; Huang, X.-m.; Hu, G.-b. Huang Hui-bai Study on the Relationship between Anthocyanin Synthesis and Related Enzymes in Litchi Peel. Chin. Agric. Sci. 2004, 12, 2028–2032. (In Chinese) [Google Scholar]
- Wang, Z.; Li, S.; Yuan, M.; Zhou, K. De novo transcriptome assembly for pericarp in Litchi chinesis Sonn. cv. Feizixiao and identification of differentially expressed genes in response to Mg Foliar Nutrient. Sci. Hortic. 2017, 226, 59–67. [Google Scholar] [CrossRef]
- Du, J.-j.; Peng, J.; Chen, T.-t.; Shui, X.; Liao, H.-z.; Lin, X.-k.; Zhou, K.-b. Respiratory mechanism of calcium magnesium foliar fertilizer in alleviating the phenomenon of sugar loss in the flesh of Feizixiao lychee. South. Agric. J. 2023, 2, 356–364. (In Chinese) [Google Scholar]
- Peng, J.; Du, J.; Wuqiang, M.; Chen, T.; Shui, X.; Liao, H.; Lin, X.; Zhou, K. Transcriptomics-based analysis of the causes of sugar receding in Feizixiao litchi (Litchi chinensis Sonn.) pulp. Front. Plant Sci. 2022, 13, 1083753. [Google Scholar] [CrossRef]
- Vanlerberghe, G.C.; Dahal, K.; Alber, N.A.; Chadee, A. Photosynthesis, respiration and growth: A carbon and energy balancing act for alternative oxidase. Mitochondrion 2020, 52, 197–211. [Google Scholar] [CrossRef]
- Lv, L. The Role of Mitochondrial Energy Metabolism and Reactive Oxygen Species in the Early Defense Response of Sweet Melon Fruits Induced by Powdery Mildew Infection and Silicon. Ph.D. Thesis, Gansu Agricultural University, Lanzhou, China, 2019. (In Chinese). [Google Scholar]
- Cvetkovska, M.; Vanlerberghe, G.C. Alternative oxidase modulates leaf mitochondrial concentrations of superoxide and nitric oxide. New Phytol. 2012, 195, 32–39. [Google Scholar] [CrossRef]
- Wang, C.-y. The Regulatory Effect of NO on Mitochondrial Function and Oxidative Damage in Peach Fruit. Master’s Thesis, Shandong Agricultural University, Tai’an, China, 2021. (In Chinese). [Google Scholar]
- Kadenbach, B.; Ramzan, R.; Vogt, S. Degenerative diseases, oxidative stress and cytochrome c oxidase function. Trends Mol. Med. 2009, 15, 139–147. [Google Scholar] [CrossRef]
- Lips, S.H.; Biale, J.B. Stimulations of oxygen uptake by electron transfer inhibitors. Plant Physiol. 1966, 41, 797–802. [Google Scholar] [CrossRef]
- Li, Y.-m.; Zhu, L.; Yang, J.-h.; Zhang, M.-f. Cloning and analysis of the alternating oxidase AOX2 gene in watermelon. J. Fruit Tree Sci. 2011, 28, 909–914. (In Chinese) [Google Scholar]
- Hu, W.-H.; Yan, X.-H.; He, Y.; Xi, R. 24-epibrassinolide alleviate drought-induced photoinhibition in Capsicum annuum via up-regulation of AOX pathway. Sci. Hortic. 2019, 243, 484–489. [Google Scholar] [CrossRef]
- Wang, Y.-n. Research on the Mechanism of Ethylene and Cyanide Respiration Involved in Salicylic Acid Enhancing Postharvest Cold Resistance of Plum Fruits. Master’s Thesis, Inner Mongolia Agricultural University, Hohhot, China, 2020. (In Chinese). [Google Scholar]
- Feng, H.; Li, X.; Duan, J.; Li, H.; Liang, H. Chilling tolerance of wheat seedlings is related to an enhanced alternative respiratory pathway. Crop Sci. 2008, 48, 2381–2388. [Google Scholar] [CrossRef]
- Rizhsky, L.; Liang, H.; Mittler, R. The Combined Effect of Drought Stress and Heat Shock on Gene Expression in Tobacco. Plant Physiol. 2002, 130, 1143–1151. [Google Scholar] [CrossRef]
- Shelyakin, M.; Zakhozhiy, I.; Golovko, T. The effect of temperature on Antarctic lichen cytochrome and alternative respiratory pathway rates. Polar Biol. 2020, 43, 2003–2010. [Google Scholar] [CrossRef]
- Zenoni, S.; Savoi, S.; Busatto, N.; Tornielli, G.B.; Costa, F. Molecular regulation of apple and grape ripening: Exploring common and distinct transcriptional aspects of representative climacteric and non-climacteric fruits. J. Exp. Bot. 2023, 74, 6207–6223. [Google Scholar] [CrossRef]
- Oliveira, M.; Mazorra, L.; Souza, A.; Silva, G.; Correa, S.; Santos, W.; Saraiva, K.; Teixeira, A.; Melo, D.; Silva, M.; et al. Involvement of AOX and UCP pathways in the post-harvest ripening of papaya fruits. J. Plant Physiol. 2015, 189, 42–50. [Google Scholar] [CrossRef]
- Sluse, F.; Jarmuszkiewicz, W. Activity and functional interaction of alternative oxidase and uncoupling protein in mitochondria from tomato fruit. Braz. J. Med. Biol. Res. 2000, 33, 259–268. [Google Scholar] [CrossRef]
- Holtzapffel, R.C.; Finnegan, P.M.; Millar, A.H.; Badger, M.R.; Day, D.A. Mitochondrial protein expression in tomato fruit during on-vine ripening and cold storage. Funct. Plant Biol. 2002, 29, 827–834. [Google Scholar] [CrossRef]
- Mansilla, N.; Garcia, L.; Gonzalez, D.H.; Welchen, E. AtCOX10, a protein involved in haem o synthesis during cytochrome c oxidase biogenesis, is essential for plant embryogenesis and modulates the progression of senescence. J. Exp. Bot. 2015, 66, 6761–6775. [Google Scholar] [CrossRef]
- González-Gordo, S.; Rodríguez-Ruiz, M.; Paradela, A.; Ramos-Fernández, A.; Corpas, F.J.; Palma, J.M. Mitochondrial protein expression during sweet pepper (Capsicum annuum L.) fruit ripening: iTRAQ-based proteomic analysis and role of cytochrome c oxidase. J. Plant Physiol. 2022, 274, 153734. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, Z.; Zhu, H.; Jiang, Y.; Jiang, G.; Qu, H. Energy homeostasis mediated by the LcSnRK1α–LcbZIP1/3 signaling pathway modulates litchi fruit senescence. Plant J. 2022, 111, 698–712. [Google Scholar] [CrossRef]
- Aghdam, M.S.; Alikhani-Koupaei, M. Exogenous phytosulfokine α (PSKα) applying delays senescence and relief decay in strawberry fruits during cold storage by sufficient intracellular ATP and NADPH availability. Food Chem. 2020, 336, 127685. [Google Scholar] [CrossRef]
- Lin, Y.; Lin, H.; Chen, Y.; Wang, H.; Lin, M.; Ritenour, M.A.; Lin, Y. The role of ROS-induced change of respiratory metabolism in pulp breakdown development of longan fruit during storage. Food Chem. 2020, 305, 125439. [Google Scholar] [CrossRef]
- Liu, T.; Qian, Z.-j.; Yang, E.; Wu, F.-w.; Qu, H.-x.; Jiang, Y.-m. Relationship between Respiratory Activity and Energy Metabolism and Quality Deterioration of Litchi Fruits. J. Fruit Tree Sci. 2010, 6, 946–951. (In Chinese) [Google Scholar]
- Du, J.-j.; Peng, J.-j.; Chen, T.-t. Tax wise Zhou Kai-bing Analysis of the reasons for the phenomenon of “sugar loss” in “Feizixiao” lychee fruit reduced by mixed application of calcium and magnesium foliar fertilizer. Fruit Trees South. China 2023, 9, 1–10. (In Chinese) [Google Scholar]
- Shui, X.; Wang, W.; Ma, W.; Yang, C.; Zhou, K. Mechanism by Which High Foliar Calcium Contents Inhibit Sugar Accumulation in Feizixiao Lychee Pulp. Horticulturae 2022, 8, 1044. [Google Scholar] [CrossRef]
- Gao, D.; Zhou, X.; Su, Y.; Zhou, K. The effect of foliar spraying of potassium, calcium, and magnesium fertilizers on the coloring of the peel of March red lychee. J. South. Agric. 2015, 46. (In Chinese) [Google Scholar]
- Zhou, X.; Su, Y.; Zhang, R.; Zhou, K. The regulatory effect of foliar spraying potassium, calcium, and magnesium fertilizers on the color of Feizixiao lychee peel. Southwest Agric. J. 2015, 28, 1713–1718. (In Chinese) [Google Scholar]
- Wang, H.C.; Huang, H.B.; Huang, X.M.; Hu, Z.Q. Sugar and acid compositions in the arils of Litchi chinensis Sonn.: Cultivar differences and evidence for the absence of succinic acid. J. Hortic. Sci. Biotechnol. 2006, 81, 57–62. [Google Scholar] [CrossRef]
- Sivakumar, D.; Korsten, L. Fruit quality and physiological responses of litchi cultivar McLean’s Red to 1-methylcyclopropene pre-treatment and controlled atmosphere storage conditions. LWT-Food Sci. Technol. 2010, 43, 942–948. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, P.; Xia, J.; Yu, T.; Lou, B.; Wang, J.; Zheng, X.D. Effect of water activity on stress tolerance and biocontrol activity in antagonistic yeast Rhodosporidium paludigenum. Int. J. Food Microbiol. 2010, 143, 103–108. [Google Scholar] [CrossRef]
- Song, C.-c. The Effect of NO Treatment on Respiration during Refrigeration of Different Fleshy Peach Fruits. Master’s Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2021. (In Chinese). [Google Scholar]
- Shi, Q.-c. Effects of Exogenous Salicylic Acid Treatment on the Respiratory Pathways of Postharvest Huangguan Pear Fruits. Master’s Thesis, Hebei Agricultural University, Baoding, China, 2009. (In Chinese). [Google Scholar]
- Zhu, Y.-c.; Cui, Z.-x.; Xu, H.; Wei, Z.-h.; Zhang, J.-w.; Bai, C.-h.; Yao, L.-x. Comparison of Fruit Quality and Flavor Characteristics of Youxin Litchi Varieties. Chin. J. Food Sci. 2023, 6, 327–338. (In Chinese) [Google Scholar]
- Liao, H.-z.; Yang, C.-k.; Lin, X.-k.; Yue, X.; Zhou, K.-b. Effect of foliar application of calcium and magnesium fertilizer on the fruit quality of Feizixiao lychee. South. Agric. J. 2021, 7, 1843–1850. (In Chinese) [Google Scholar]
- Duan, X.; Jiang, Y.; Su, X.; Liu, H.; Li, Y.; Zhang, Z.; Zheng, Y.; Jiang, W. Role of pure oxygen treatment in browning of litchi fruit after harvest. Plant Sci. 2004, 167, 665–668. [Google Scholar] [CrossRef]
- Guo, J.; Chen, Y.-l.; Wang, P.; Wang, L.; Chen, F.-p.; Luo, Z.; Yu, X. The effect of different moisture control packaging on the storage quality of lychee Food and Fermentation. Industry 2021, 47, 169–175. (In Chinese) [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Names | Left Primer Sequences (5′ to 3′) | Right Primer Sequences (5′ to 3′) |
|---|---|---|
| Cluster-6206.61154 | CGGACACTGCTTTACCCACT | TCACCCTTAGATGCCAAGCA |
| Cluster-6206.84300 | TTGTTCATGCGGCAACTTGG | GCCTCAGGATTGTCGACACA |
| Cluster-6206.102193 | GCAGCAGTTCCTGGTATGGT | CGCTCGTTTTCGGCTTCTTC |
| Cluster-6206.20086 | TGCTCAGCCAAGAGTAAGCC | CCAAAAGAGGCCGCTGTAGA |
| Cluster-6206.31144 | ACTTTGCCACCGAGGATCAG | ACTGAAAGCAAGCCCCTTGT |
| Cluster-6206.60735 | CGAGCTATTAACGCAGCACA | TGAAATGCCAATGACAAAGC |
| Cluster-6206.73361 | AAGTCACCATTCGAGGGCTG | GTTGAAGTGCGGGACCAAAC |
| Cluster-6206.77187 | ACTGAAAAACTTGCCGTGGC | TACAACTCCACTGCACTGGC |
| Cluster-6206.86111 | TCTGAAACGAGGCTCGAACC | ACCATATAGCAACCTGGCGG |
| Cluster-6206.80232 | GATGCTTGAAACAGTGGCGG | AGTGCTTTAATCCAGCCCCC |
| β-Actin | AGTTTGGTTGATGTGGGAGAC | TGGCTGAACCCGAGATGAT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, S.; Du, J.; Peng, J.; Zhou, K.; Ma, W. The Effects of Mixed Foliar Nutrients of Calcium and Magnesium on the Major Bypass Respiratory Pathways in the Pulp of ‘Feizixiao’ Litchi. Horticulturae 2024, 10, 248. https://doi.org/10.3390/horticulturae10030248
Shi S, Du J, Peng J, Zhou K, Ma W. The Effects of Mixed Foliar Nutrients of Calcium and Magnesium on the Major Bypass Respiratory Pathways in the Pulp of ‘Feizixiao’ Litchi. Horticulturae. 2024; 10(3):248. https://doi.org/10.3390/horticulturae10030248
Chicago/Turabian StyleShi, Shaopu, Jingjia Du, Junjie Peng, Kaibing Zhou, and Wuqiang Ma. 2024. "The Effects of Mixed Foliar Nutrients of Calcium and Magnesium on the Major Bypass Respiratory Pathways in the Pulp of ‘Feizixiao’ Litchi" Horticulturae 10, no. 3: 248. https://doi.org/10.3390/horticulturae10030248