1. Introduction
Winter squash (
Cucurbita maxima Duch.) is a fragrant, sweet, nutritious fruit with certain medicinal and health-promoting properties and is very popular among food manufacturers and consumers [
1,
2,
3,
4]. Traditional crop breeding methods, such as interspecific hybridization and intraspecific hybridization for
C. maxima, have a long cycle and low efficiency and can no longer meet market demand. With the development of genome sequencing and genome editing technologies, the precise improvement of plant traits through genetic transformation has become a new approach for many species of the
Cucurbitaceae. According to previous reports, cucumber [
5,
6,
7,
8], melon [
9,
10,
11,
12,
13], and watermelon [
14,
15,
16,
17] have successfully undergone gene transformation and genome editing.
The genus
Cucurbita is recalcitrant to transformation, and there are few related reports on
Cucurbita moschata Duch. In 2011, Nanasato et al. first used wounding with aluminum borate whiskers to induce Agrobacterium-mediated transformation of
C. moschata, and the average transgenic efficiency was approximately 2.7% [
18,
19]. In 2022, Xin et al. utilized CRISPR/Cas9 technology to create compact structural mutants in
C. moschata by optimizing genetic transformation programs [
20]. A transient transformation system for studying gene function in
C. moschata has also been described [
21]. However, due to differences in genotype and regeneration pathways, the current transformation methods used for the genus
Cucurbita are not universally applicable. Notably, there are no reports of transformation in
C. maxima, possibly because of the lack of stable and efficient regeneration systems.
Previous studies have demonstrated that organogenesis in the genus
Cucurbita is influenced by explant type, genotype, induction media, and other factors [
22,
23,
24,
25]. Lee et al. reported the cultivation of cotyledon explants from two winter squash varieties, both of which successfully produced complete plants, and investigated several factors affecting organogenesis, such as the size of the explants and the type of culture media [
26]. The critical factor in inducing plant organogenesis is the use of hormones on explants with regenerative potential at a certain growth stage [
27]. Since the organogenesis of winter squash is also influenced by various factors, improving the regeneration rate of winter squash solely through explants is insufficient. More attention should be given to the mutual influence of these factors. A long organogenesis time is also a vital matter that must be addressed in the regeneration of the genus
Cucurbita. Additionally, a critical understanding of the timing and site of shoot formation will also be helpful for the development of genetic transformation methods. However, there is currently limited research on organogenesis in winter squash.
To address the aforementioned issues, we evaluated the effects of explant type, genotype, culture media, and their interactions on the de novo organogenesis of winter squash and observed histological events related to the initiation and development of shoots. The aim of this study was to optimize the de novo organogenesis systems of five winter squash lines, determine the timing and origin site of shoots, and obtain complete plants in a short period, providing efficient and stable de novo organogenesis system support for the genetic transformation of winter squash.
4. Discussion
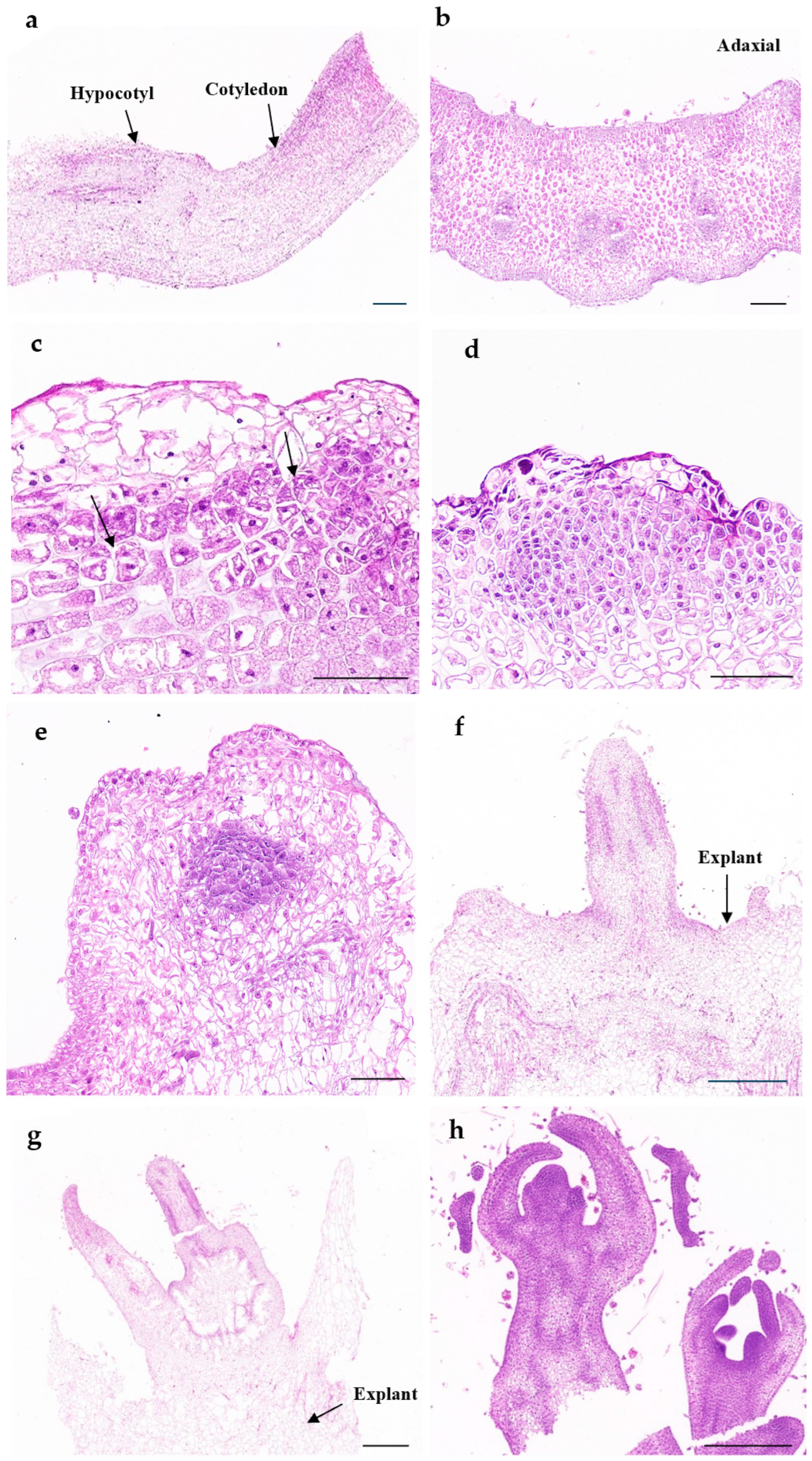
Whether or not winter squash explants will begin to regenerate will largely depend on the extent of their limitations in relation to their original tissue capacity. In this study, we designed four explants with regeneration potential, and the shoot induction rates showed that the plants could produce shoots (
Figure 2). The shoot regeneration rates observed in the cotyledons were greater than those in the half cotyledons (1/2 cotyledons). Similarly, Kim et al. reported that smaller cotyledons of figleaf gourd (
C. ficifolia Bouche) produced fewer adventitious shoots, which indicated that the shoot induction rate is greater when the area adjacent to the explant containing the cotyledon and hypocotyl is greater [
29]. We found that the shoots appeared at the junction between the cotyledon and hypocotyl, which is consistent with previous findings for
C. pepo [
30]. We achieved a regeneration rate of 52.62% for the 5 mm long hypocotyl. However, Lee et al. reported that shoots cannot be regenerated from winter squash hypocotyls [
26]. We speculate that differences in the specific location of hypocotyl sampling and genotype may account for this discrepancy in results. The shoot regeneration rate of cotyledons was significantly greater than that of the other three types of explants on ten different media (
Figure 2). Previous studies have shown that plant regeneration from cotyledons is an immensely reliable reproductive process that maintains genetic similarity. Cotyledons can serve as explants for de novo shoot regeneration in winter squash.
Genotype is an important factor affecting cell division, differentiation, and morphogenesis in the organogenesis of the family
Cucurbitaceae [
31]. Obtaining effective regenerated plants from commercial genotypes is a prerequisite for applying genetic transformation techniques for crop improvement [
32]. Differences in embryo and shoot induction rates have been observed even when cultured on the same medium in plants such as summer squash (
C. pepo) [
25] and pumpkin (
C. moschata) varieties [
33]. This experiment evaluated the effect of genotype on regeneration using three widely cultivated varieties (L-1, L-2, and L-3) in China and two inbred lines (L-4 and L-5) from Hunan Province. The results showed that the induction rate of regenerated shoots was the highest for L-3 (Jin-Li), followed by L-2 (Hong-li No. 2) and L-5 (JP) (
Table 3). In addition, the number of regeneration days and average number of shoots were closely related to the tested genotypes. Overall, under the same cultivation conditions, the shoot regeneration rate of the commercial varieties was greater than that of the inbred lines.
The induction medium components had a strong impact on the regeneration of winter squash. It is thought that cytokinin activity in explants can be effectively triggered by lower doses of auxin to produce more shoots [
34]. However, Zhang et al. concluded that 6-BA is necessary for
C. moschata to induce shoot formation in cotyledons and that there was no difference in the concentration of 6-BA between 0.50~2.00 mg/L [
35]. Similarly, Samuel Aworunse et al. reported a lack of shoot response in
C. pepo explants cultured on media supplemented with both 6-BA and 2,4-D, whereas lower concentrations of 6-BA alone achieved better outcomes [
36]. In this paper, shoot induction was assessed using four media supplemented with 6-BA and six combinations of media, and the results indicated that on culture media supplemented with only 6-BA (1.0 mg/L-4 mg/L), the shoot induction rate tended to increase and then decrease with increasing 6-BA concentration (
Figure 2). Although 6-BA influences cotyledon regeneration, the addition of 2.0 mg/L6-BA and 0.2 mg/L IAA media resulted in a greater shoot induction rate than did the addition of media supplemented with only 2.0 mg/L6-BA (
Table 4). Previous studies have also reported similar phenomena [
37,
38,
39]. A further finding was the production of fragile white embryogenic callus tissue at the contact point between the explant and culture medium, which increased with increasing hormone content. This phenomenon also occurs in
C. moschata [
35]. Ananthakrishnan et al. suggested that rapid callus growth in
C. pepo reduces shoot regeneration [
30]. However, our results showed that this fact did not affect the regeneration of shoots. This difference may be due to the difference in the ratio of auxin to cytokinin in the media.
Several factors can affect the induction of
Cucurbitaceae shoot growth, the two most important of which are genotype and medium composition [
40,
41]. The mutual influence between these factors leads to increased complexity of this process. To date, many studies have focused on single-factor effects, but evaluations of the interactions between factors in the genus
Cucurbita have not yet been performed. However, we can still draw some insights from previous research. Lee et al. induced shoot formation from two genotypes of winter squash on 6-BA media supplemented with four different concentrations of salt [
26]. The results showed that the different genotypes had distinct regeneration rates on the same medium and that the same genotype had varying regeneration rates on different media, indicating an interaction between these two factors. A similar conclusion was reached in another study on shoot regeneration in
C. pepo [
25]. In this study, we determined the impact of genotype and induction medium interactions on shoot induction through two-way ANOVA and found that their interactions had a greater impact on shoot induction than did their individual effects. This result indicates that mutual interactions among factors represent the main stimuli for inducing shoot growth and should receive increased attention.
MS, 1/2 MS, or medium supplemented with the hormones IAA and NAA are commonly used to induce root growth in
Cucurbita. Kurtar et al. rooted
C. maxima and
C. moschata plants on MS media supplemented with 0.01 mg/l IAA [
42]. Kabir et al. also used this medium to induce root growth in sweet gourd (
C. moschata) [
33]. We evaluated whether the rooting duration, rooting rate, and average rooting number of the winter squash plants were variable in the five different culture media (
Table 6). The highest rooting rate was observed for R3 (97.77%), followed by R2 (95.55%), and the lowest rooting rate was observed for R5 (66.87%). From the perspective of plant growth status, R2 (1/2 MS) is the most suitable as a root induction medium for winter squash. We also found that the rooting time of shoots in culture medium with 0.1–0.3 mg/L NAA added was reduced compared to that in MS medium.
The development of plants is largely determined by their ability to form new organs [
43]. Somatic cell regeneration in the family
Cucurbitaceae can occur from cotyledons or through calli. Although genetic transformation plants through cotyledon regeneration may exhibit chimerism, de novo organogenesis is more suitable for the genus
Cucurbita due to its minimal potential for somaclonal variation and capacity for a shorter regeneration time [
20]. Histological observations revealed that cell division occurred in the subepidermal region before the appearance of meristem tissue (
Figure 4). These cells, which have perpetual proliferative abilities, continuously proliferate through mitosis and differentiate into shoots. However, during the normal growth process of plants, cells with potential meristematic ability usually have little or no meristematic function [
27]. The anatomical structures of the de novo regenerated shoots and the vegetative buds produced on intact plants are similar. We believe that these proliferating cells under the epidermis may be targets for direct gene transfer in winter squash cotyledon explants. If the intensity of the Agrobacterium infection can be controlled at these sites, the probability of obtaining transformed plants will greatly increase.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}