
Recent Advancements in Mitigating Abiotic Stresses in Crops
 ,
,  , ,
, , 
Abstract
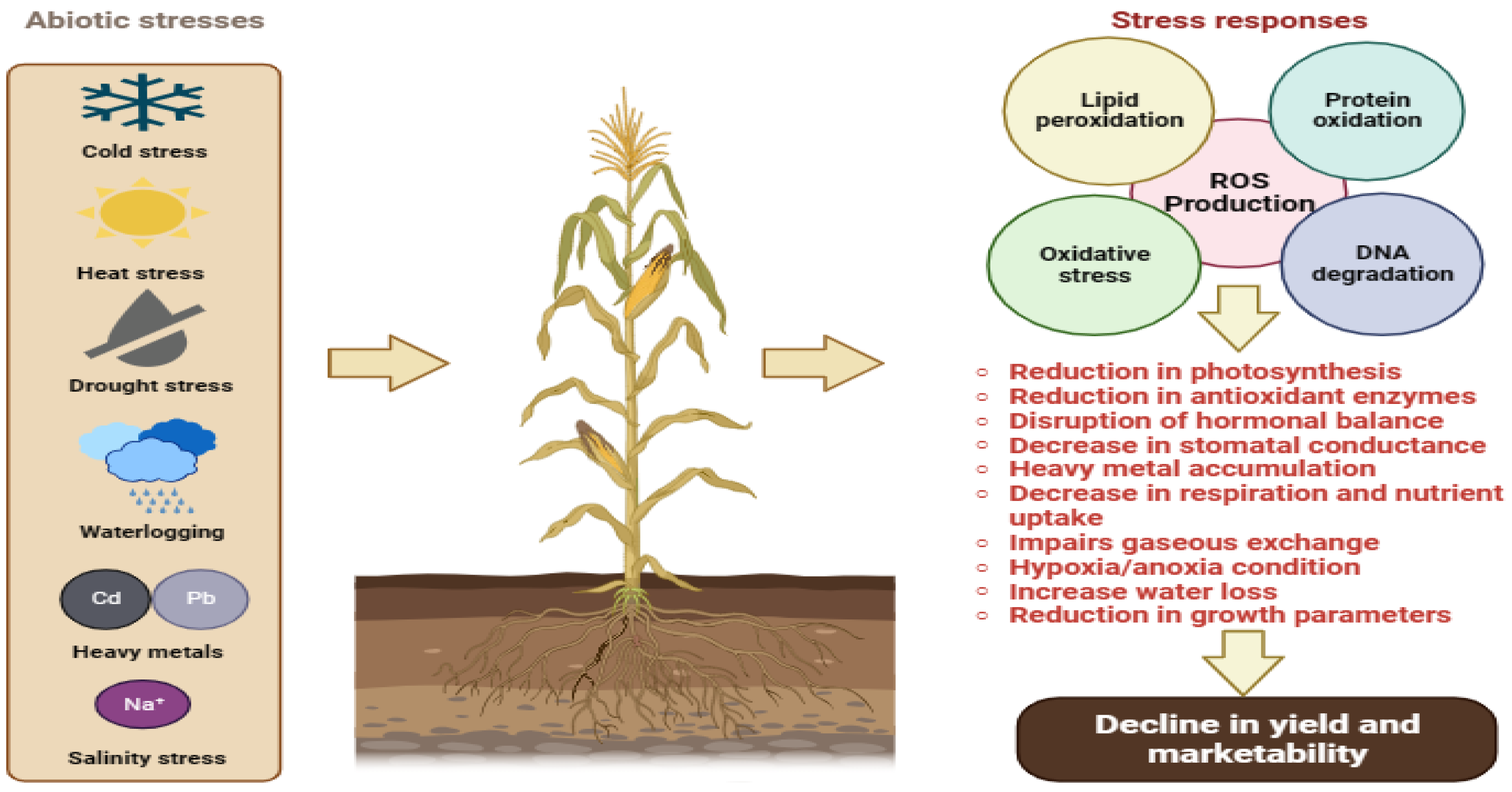
:1. Introduction
2. Management Strategies of Abiotic Stress
2.1. Breeding Approach
2.1.1. Conventional Breeding

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abiotic Stresses | Crops | Tolerant Cultivars | Selection Method | Country | References |
|---|---|---|---|---|---|
| Salinity | Tomato | Edkway | Natural selection | Egypt | [27] |
| Barley | Golden Promise | Induced mutation | Scotland | [26] | |
| Alfalfa | AZ Germ Salt 1 | Back crossing selection | USA | [27] | |
| Rice | Jiabuyu | Recurrent selection | China | [28] | |
| Rice | (CSR-1), Dasal (CSR-2), and Getu (CSR-3) | Pure line selection | India | [29] | |
| Heat stress | Tomato | L72 | Hybridization | [30] | |
| Cowpea | Pedigree backcross | [31] | |||
| Tomato | Saladette | Selection | USA | [31] | |
| Waterlogging | Soybean | NN1138-2, M8206, and ZXD | Selection | [32] | |
| Cold stress | Alfalfa | Recurrent selection | [33] | ||
| Heavy metal | Rice | Induced mutation | Japan | [34] | |
| Wheat | Strongfield | Selection | Canada | [35] | |
| Drought | Rice | Tarom Mahalli | Induced mutation | Iran | [14] |
| Cotton | BH-167 | Pedigree method | Pakistan | [36] | |
| Rice | Jiabuyu | Recurrent selection | China | [28] | |
| Rice | MR219-9 and MR219-4 | Induced mutation | Malaysia | [14] |
2.1.2. Grafting
2.1.3. QTL Mapping and Marker-Assisted Breeding
| Abiotic Stresses | Crop | Population | QTL | Traits | References |
|---|---|---|---|---|---|
| Waterlogging | Soybeans | Benning × PI 416937 | qFTS-1, qSR-5.1 | Flooding tolerance scores and survival rates | [57] |
| Rice | Indra × AC39416A | qAG3.1, qSF10.1 | Anaerobic germination | [58] | |
| Wheat | Yangmai 16 × Zhongmai 895 | QWTC.caas-4AL | Confer tolerance | [59] | |
| Drought | Maize | SNJ201126 × HKI161 | qCH1–1, qCW2–1 | Cob weight, cob height | [60] |
| Horsegram | HPKM249 × HPK4 | qRL01, qRL02, QCHL01, QPRO01 | Root length, chlorophyll, proline | [61] | |
| Chickpea | Pusa 362 × SBD 377 | qYLD7.1, qRWC1.1 | Yield, relative water content | [62] | |
| Tomato | Advanced generation | Flw1.1 and FW2.2 | Time to flower and fruit weight | [63] | |
| Bean | Portillo × Red Hawk | DF3.2, DF11.1 | Days to flowering | [64] | |
| Barley | TX9425 × Franklin | QRMO.TxFr.2H, QPC-D.TxFr.3H | Relative water content and proline | [65] | |
| Common bean | BAT 881 × G21212 | Yd4.1 | Yield components | [66] | |
| Rice | Lvhan 1 × Aixian 1 | qPH10.1, qEPN6.1, qPL9.1, qTGW2.1, qTGW6.1, qTGW8.1, qLL7.1 qLW7.1, qLA7.1 | Plant height, effective panicles number, panicle length, thousand-grain weight, leaf length, leaf width, and leaf area (LA) | [67] | |
| Groundnut | TAG 24 × ICGV 86031 | qDW-A05.2, qISC-A04.1, qNB-A07.1, qTR-A09.1 | Total dry matter, canopy conductance, number of branches, transpiration rate | [68] | |
| Salinity | Chickpea | ICCV 10 × DCP 92 | qPHC5.2, qYPPS6.1, q100SWS3.1 | Plant height and yield traits | [69] |
| Barley | TX9425 × Franklin | QPC-S.TxFr.3H, QST.TxFr.7H | Proline, salt tolerance | [65] | |
| Rice | R29 × Pokkali | qSIS1, qRSH12 | Salt injury score, height | [70] | |
| Sorghum | Shihong 137 × L-Tian | qTB6, qSFW9, qJW9, | total biomass, stem fresh weight, juice weight | [71] | |
| Tomato | Advanced generation | Flw1.1 and FW2.2 | Time to flower and fruit weight | [63] | |
| Indian mustard | CS52 × RH30 | QBYP8.1, QSP4.1, QPB4.1, QSS4.1, QSS4.2, QSYP4.1, QMI8.1 | Yield per plant, siliquae per plant, primary branches, seed per silique, seed yield per plant, and membrane injury | [72] | |
| Cucumber | CG104 × CG37 | qST6.2 | Salt tolerance | [73] | |
| Jute | J009 × GFG | qJST-1, qJST-2 | Salt tolerance | [74] | |
| Heat stress | Chickpea | DCP 92-3 × ICCV 92944 | CaDFI_LS6.1, CaDPI_LS7.2, CaCHL_NS4.3 | Days to flowering, chlorophyll, and yield traits | [75] |
| Tomato | LA1698 × LA2093 | qHII-1-1 | Heat injury index | [76] | |
| Cotton | MNH-886 × MNH-814 | qFSHa1, qNOB1 | Sympodial node height, Number of bolls | [77] | |
| Cucumber | 99,281 × 931 | qHT1.1 | Heat tolerance | [78] | |
| Bread wheat | Germplasm | QTL.16 | Thousand kernel weight | [79] | |
| Bottle gourd. | L1 (P17) × L6 (P23) | qHT2.1 | Relative electrical conductivity | [80] | |
| Cold stress | Rice | Natural population | qSR2–1, qSR3–4, qSR3–5 | Seedling survival rate | [81] |
| Alfalfa | 3010 × CW 1010 | dbr1, dft1 | Cold tolerance and biomass ratio | [82] | |
| Peanut | DF12 × Huayu 44 | qRGRB09 | Cold tolerance | [83] | |
| Heavy metal | Rice | WTR1 × Hao-an-nong | qRChlo1 | Chlorophyll content | [84] |
| Soybean | AC Hime × Westag-97 | Cda1 | Low Cd concentration | [85] | |
| Cabbage | Y177 × Y195 | Zn100SDB6 | Shoot dry biomass | [86] | |
| Maize | Huang-C × Xu178 | qLAV1 and qSAC1 | Kernel | [87] |
2.1.4. Genetic Engineering of Abiotic Stress Tolerance
2.1.5. Genome Editing
2.1.6. Speed Breeding: A Speedy Approach to Develop Crop Resilience to Abiotic Stress
2.2. Seed Priming Techniques
Seed Treatment with Microbes
2.3. Plant Growth Regulators and Osmoprotectants
Melatonin: An Emerging Plant Growth Regulator
2.4. Inoculation of Arbuscular Mycorrhiza (AMF)
2.5. Agronomical Approach
2.5.1. Cultural Practices
2.5.2. Nutrient Management
2.6. Emerging Strategies
2.6.1. Biochar Application
2.6.2. Kaolin
2.6.3. Superabsorbent: A Novel Soil Conditioner
2.6.4. Seaweed Extract
| Abiotic Stresses | Seaweed Extract | Plant under Study | Impacts | References |
|---|---|---|---|---|
| Salinity | Seaweed extract | Bell pepper | Improved leaf relative water content, leaf greenness, and membrane stability index | [207] |
| Ulva fasciata, Cystoseira compressa, Laurencia obtuse | Cowpea and Maize | Increased biomass and conferred tolerance | [210] | |
| Ascophyllum nodosum | Tomato | Increased root to shoot ratio and yield | [211] | |
| Cold stress | A. nodosum | Arabidopsis | Downregulated chlorophyll degradation genes and improved chlorophyll content | [212] |
| A. nodosum | Tobacco | Upregulated freezing tolerance genes | [213] | |
| Brown-seaweed extract | Tomato | Increased accumulation of proline, polyphenols, flavonoids, tannins, and carotenoid contents | [214] | |
| Drought | Fucus spiralis, Ulva lactuca, Laminaria ochroleuca | Fababean | Enhanced soluble sugars accumulation and relative water content and reduced malondialdehyde | [215] |
| Ascophyllum nodosum | Soybean | Expressed stress-responsive genes and antioxidant enzymes | [216] | |
| Ascophyllum nodosum | Arabidopsis | Downregulated stress-responsive negative growth regulator | [217] | |
| Ecklonia maxima | Chicory plants | Enhanced physiological traits (relative water content, water use efficiency, chlorophyll content, nutrient uptake) | [218] | |
| Heat stress | Ascophyllum nodosum | Spinach | Improved germination percentage and germination speed, and reduced hydrogen peroxide and malondialdehyde content | [219] |
| Seaweed extract | Bentgrass | Enhanced tolerance | [220] |
2.6.5. Higher Plant Extract
2.6.6. Yeast Extract
2.6.7. Chitosan
2.6.8. The Use of Nanotechnology
3. Conclusions and Future Prospect
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Rai, K.K. Integrating Speed Breeding with Artificial Intelligence for Developing Climate-smart Crops. Mol. Biol. Rep. 2022, 49, 11385–11402. [Google Scholar] [CrossRef]
- Younis, A.; Ramzan, F.; Ramzan, Y.; Zulfiqar, F.; Ahsan, M.; Lim, K.B. Molecular Markers Improve Abiotic Stress Tolerance in Crops: A Review. Plants 2020, 9, 1374. [Google Scholar] [CrossRef] [PubMed]
- El-Saadony, M.T.; Saad, A.M.; Soliman, S.M.; Salem, H.M.; Desoky, E.S.M.; Babalghith, A.O.; AbuQamar, S.F. Role of Nanoparticles in Enhancing Crop Tolerance to Abiotic Stress: A Comprehensive Review. Front. Plant Sci. 2022, 13, 946717. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop Production under Drought and Heat Stress: Plant Responses and Management Options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed]
- Alotaibi, M. Climate Change, its Impact on Crop Production, Challenges, and Possible Solutions. Not. Bot. Horti Agrobot. Cluj-Napoca 2023, 51, 13020. [Google Scholar] [CrossRef]
- Giordano, M.; Petropoulos, S.A.; Rouphael, Y. Response and Defence Mechanisms of Vegetable Crops against Drought, Heat and Salinity Stress. Agriculture 2021, 11, 463. [Google Scholar] [CrossRef]
- Waqas, M.A.; Wang, X.; Zafar, S.A.; Noor, M.A.; Hussain, H.A.; Azher Nawaz, M.; Farooq, M. Thermal Stresses in Maize: Effects and Management Strategies. Plants 2021, 10, 293. [Google Scholar] [CrossRef]
- dos Santos, T.B.; Ribas, A.F.; de Souza, S.G.H.; Budzinski, I.G.F.; Domingues, D.S. Physiological Responses to Drought, Salinity, and Heat Stress in Plants: A Review. Stresses 2022, 2, 113–135. [Google Scholar] [CrossRef]
- Rajput, V.D.; Minkina, T.; Kumari, A.; Harish; Singh, V.K.; Verma, K.K.; Mandzhieva, S.; Sushkova, S.; Srivastava, S.; Keswani, C. Coping with the Challenges of Abiotic Stress in Plants: New Dimensions in the Field Application of Nanoparticles. Plants 2021, 10, 1221. [Google Scholar] [CrossRef]
- Faiz, H.; Ayyub, C.M.; Khan, R.W.; Ahmad, R. Morphological, Physiological and Biochemical Responses of Eggplant (Solanum melongena L.) Seedling to Heat Stress. Pak. J. Agric. Sci. 2020, 57, 371–380. [Google Scholar]
- Bhat, K.A.; Mahajan, R.; Pakhtoon, M.M.; Urwat, U.; Bashir, Z.; Shah, A.A.; Agrawal, A.; Baht, A.; Sofi, P.A.; Masi, A.; et al. Low Temperature Stress Tolerance: An Insight into the Omics Approaches for Legume Crops. Front. Plant Sci. 2022, 13, 888710. [Google Scholar] [CrossRef]
- Ahmad, N.; Naeem, M.; Ali, H.; Alabbosh, K.F.; Hussain, H.; Khan, I.; Siddiqui, S.A.; Khan, A.A.; Iqbal, B. From Challenges to Solutions: The Impact of Melatonin on Abiotic Stress Synergies in Horticultural Plants Via Redox Regulation and Epigenetic Signaling. Sci. Hortic. 2023, 321, 112369. [Google Scholar] [CrossRef]
- Kompas, T.; Pham, V.H.; Che, T.N. The Effects of Climate Change on GDP by Country and the Global Economic Gains from Complying with the Paris Climate Accord. Earth’s Future 2018, 6, 1153–1173. [Google Scholar] [CrossRef]
- Oladosu, Y.; Rafii, M.Y.; Samuel, C.; Fatai, A.; Magaji, U.; Kareem, I.; Kamarudin, Z.S.; Muhammad, I.; Kolapo, K. Drought Resistance in Rice from Conventional to Molecular Breeding: A review. Int. J. Mol. Sci. 2019, 20, 3519. [Google Scholar] [CrossRef]
- Oyebamiji, Y.O.; Shamsudin, N.A.A.; Ikmal, A.M.; Rafii, M. Heat Stress in Vegetables: Impacts and Management Strategies-A Review. Sains Malays. 2023, 52, 1925–1938. [Google Scholar] [CrossRef]
- Hassan, M.U.; Chattha, M.U.; Khan, I.; Chattha, M.B.; Barbanti, L.; Aamer, M.; Hossain, J.; Sarkar, S.; Saha, S.; Bhadra, P.; et al. Heat Stress in Cultivated Plants: Nature, Impact, Mechanisms, and Mitigation Strategies—A Review. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2021, 155, 211–234. [Google Scholar] [CrossRef]
- Monteiro, E.; Gonçalves, B.; Cortez, I.; Castro, I. The Role of Biostimulants as Alleviators of Biotic and Abiotic Stresses in Grapevine: A Review. Plants 2022, 11, 396. [Google Scholar] [CrossRef]
- Abideen, Z.; Waqif, H.; Munir, N.; El-Keblawy, A.; Hasnain, M.; Radicetti, E.; Mancinelli, R.; Nielsen, B.L.; Haider, G. Algal-mediated nanoparticles, phycochar, and biofertilizers for mitigating abiotic stresses in plants: A review. Agronomy 2022, 12, 1788. [Google Scholar] [CrossRef]
- Ashraf, M.; Akram, N.A. Improving Salinity Tolerance of Plants through Conventional Breeding and Genetic Engineering: An Analytical Comparison. Biotechnol. Adv. 2009, 27, 744–752. [Google Scholar] [CrossRef]
- Ahmed, F.; Rafii, M.Y.; Ismail, M.R.; Juraimi, A.S.; Rahim, H.A.; Asfaliza, R.; Latif, M.A. Waterlogging Tolerance of Crops: Breeding, Mechanism of Tolerance, Molecular Approaches, and Future Prospects. BioMed Res. Int. 2013, 2013, 963525. [Google Scholar] [CrossRef]
- Guan, Y.; Xie, Z.; Guan, J.; Zhao, Q.; Rasheed, A.; Hussain, M.I.; Ali, I.; Zhang, S.; Hassan, M.U.; Hashem, M.; et al. The Intervention of Classical and Molecular Breeding Approaches to Enhance Flooding Stress Tolerance in Soybean—An review. Front. Plant Sci. 2022, 13, 1085368. [Google Scholar]
- Turan, S.; Cornish, K.; Kumar, S. Salinity Tolerance in Plants: Breeding and Genetic Engineering. Aust. J. Crop Sci. 2012, 6, 1337–1348. [Google Scholar]
- Seleiman, M.F.; Aslam, M.T.; Alhammad, B.A.; Hassan, M.U.; Maqbool, R.; Chattha, M.U.; Khan, I.; Gitari, H.I.; Uslu, S.O.; Roy, R.; et al. Salinity Stress in Wheat: Effects, Mechanisms and Management Strategies. Phyton 2022, 91, 667–694. [Google Scholar] [CrossRef]
- Srivastava, N. Salt stress responses and tolerance in wheat. In Wheat Production in Changing Environments: Responses, Adaptation and Tolerance; Springer: Berlin/Heidelberg, Germany, 2019; pp. 89–127. [Google Scholar]
- Pham, D.; Hoshikawa, K.; Fujita, S.; Fukumoto, S.; Hirai, T.; Shinozaki, Y.; Ezura, H. A Tomato Heat-tolerant Mutant Shows Improved Pollen Fertility and Fruit-setting Under Long-term Ambient High Temperature. Environ. Exp. Bot. 2020, 178, 104150. [Google Scholar] [CrossRef]
- Ashraf, M.; Athar, H.R.; Harris, P.J.C.; Kwon, T.R. Some Prospective Strategies for Improving Crop Salt Tolerance. Adv. Agron. 2008, 97, 45–110. [Google Scholar]
- Fageria, N.K.; Stone, L.F.; Santos, A.B.D. Breeding for Salinity Tolerance. In Plant Breeding for Abiotic Stress Tolerance; Fritsche-Neto, R., Borém, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 103–122. [Google Scholar] [CrossRef]
- Pang, Y.; Chen, K.; Wang, X.; Xu, J.; Ali, J.; Li, Z. Recurrent Selection Breeding by Dominant Male Sterility for Multiple Abiotic Stresses Tolerant Rice Cultivars. Euphytica 2017, 213, 268. [Google Scholar] [CrossRef]
- Meena, H.P.; Bainsla, N.K.; Yadav, D.K. Breeding for Abiotic Stress Tolerance in Crop Plants. In Recent Advances in Plant Stress Physiology; Daya Publishing House: New Delhi, India, 2016; pp. 329–378. [Google Scholar]
- Hazra, P.; Samsul, H.A.; Sikder, D.; Peter, K.V. Breeding Tomato (Lycopersicon esculentum Mill) Resistant to High Temperature Stress. Int. J. Plant Breed. 2007, 1, 31–40. [Google Scholar]
- Hall, A.E. Breeding for Heat Tolerance. Plant Breed. Rev. 2010, 10, 129–168. [Google Scholar]
- Ali, M.J.; Yu, Z.; Xing, G.; Zhao, T.; Gai, J. Establishment of Evaluation Procedure for Soybean Seed-flooding Tolerance and its Application to Screening for Tolerant Germplasm Sources. Legume Res.-Int. J. 2018, 41, 34–40. [Google Scholar] [CrossRef]
- Rémus-Borel, W.; Castonguay, Y.; Cloutier, J.; Michaud, R.; Bertrand, A.; Desgagnés, R.; Laberge, S. Dehydrin variants associated with superior freezing tolerance in alfalfa (Medicago sativa L.). Theor. Appl. Genet. 2010, 120, 1163–1174. [Google Scholar] [CrossRef]
- Satoh-Nagasawa, N.; Mori, M.; Nakazawa, N.; Kawamoto, T.; Nagato, Y.; Sakurai, K.; Takahashi, H.; Watanabe, A.; Akagi, H. Mutations in Rice (Oryza sativa) Heavy Metal ATPase 2 (OsHMA2) Restrict the Translocation of Zinc and Cadmium. Plant Cell Physiol. 2012, 53, 213–224. [Google Scholar] [CrossRef]
- Chen, Q.; Wu, F.B. Breeding for Low Cadmium Accumulation Cereals. J. Zhejiang University. Sci. B 2020, 21, 442. [Google Scholar] [CrossRef]
- Minhas, R.; Shah, S.M.; Akhtar, L.H.; Awais, S.; Shah, S. Development of a New Drought Tolerant Cotton Variety “BH-167” by Using Pedigree Method. J. Environ. Agric. Sci. 2018, 14, 54–62. [Google Scholar]
- Gaion, L.A.; Braz, L.T.; Carvalho, R.F. Grafting in Vegetable Crops: A Great Technique for Agriculture. Int. J. Veg. Sci. 2018, 24, 85–102. [Google Scholar] [CrossRef]
- Lee, J.M.; Kubota, C.; Tsao, S.J.; Bie, Z.; Echevarria, P.H.; Morra, L.; Oda, M. Current Status of Vegetable Grafting: Diffusion, Grafting Techniques, Automation. Sci. Hortic. 2010, 127, 93–105. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C.; Colla, G. Vegetable grafting: A toolbox for Securing Yield Stability Under Multiple Stress Conditions. Front. Plant Sci. 2018, 8, 2255. [Google Scholar] [CrossRef]
- Sen, A.; Chatterjee, R.; Bhaisare, P.; Subba, S. Grafting as an Alternate Tool for Biotic and Abiotic Tolerance with Improved Growth and Production of Solanaceous Vegetables: Challenges and Scopes in India. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 121–135. [Google Scholar] [CrossRef]
- Kumar, S.; Bharti, N.; Saravaiya, S.N. Vegetable Grafting: A Surgical Approach to Combat Biotic and abiotic stresses—A review. Agric. Rev. 2018, 39, 1–11. [Google Scholar] [CrossRef]
- Pradeepkumara, N.; Subhasmita, S.; Kowsalya, K.B.; Jain, S.; Hongal, D.; Meher, S.S.; Kiran, B. Grafting Technique a Novel Advancement in Vegetable Breeding. Pharma Innov. J. 2022, 11, 469–480. [Google Scholar]
- Alkharabsheh, H.M.; Seleiman, M.F.; Hewedy, O.A.; Battaglia, M.L.; Jalal, R.S.; Alhammad, B.A.; Schillaci, C.; Ali, N.; Al-Doss, A. Field Crop Responses and Management Strategies to Mitigate Soil Salinity in Modern Agriculture: A Review. Agronomy 2021, 11, 2299. [Google Scholar] [CrossRef]
- Singh, H.; Sethi, S.; Kaushik, P.; Fulford, A. Grafting Vegetables for Mitigating Environmental Stresses Under Climate Change: A Review. J. Water Clim. Change 2020, 11, 1784–1797. [Google Scholar] [CrossRef]
- Coşkun, Ö.F. The Effect of Grafting on Morphological, Physiological and Molecular Changes Induced by Drought Stress in Cucumber. Sustainability 2023, 15, 875. [Google Scholar] [CrossRef]
- Ali, S.; Rizwan, M.; Arif, M.S.; Ahmad, R.; Hasanuzzaman, M.; Ali, B.; Hussain, A. Approaches in Enhancing Thermotolerance in Plants: An Updated Review. J. Plant Growth Regul. 2020, 39, 456–480. [Google Scholar] [CrossRef]
- Muneer, S.; Ko, C.H.; Wei, H.; Chen, Y.; Jeong, B.R. Physiological and Proteomic Investigations to Study the Response of Tomato Graft Unions Under Temperature Stress. PLoS ONE 2016, 11, e0157439. [Google Scholar] [CrossRef]
- Raj, S.R.G.; Nadarajah, K. QTL and Candidate Genes: Techniques and Advancement in Abiotic Stress Resistance Breeding of Major Cereals. Int. J. Mol. Sci. 2022, 24, 6. [Google Scholar] [CrossRef]
- Ajayi, O.O.; Bregitzer, P.; Klos, K.; Hu, G.; Walling, J.G.; Mahalingam, R. QTL Mapping of Shoot and Seed Traits Impacted by Drought in Barley Using a Recombinant Inbred Line Population. BMC Plant Biol. 2023, 23, 283. [Google Scholar] [CrossRef] [PubMed]
- La Borde, N.; Rajewski, J.; Dweikat, I. Novel QTL for Chilling Tolerance at Germination and Early Seedling Stages in Sorghum. Front. Genet. 2023, 14, 1129460. [Google Scholar] [CrossRef] [PubMed]
- Muthu, V.; Abbai, R.; Nallathambi, J.; Rahman, H.; Ramasamy, S.; Kambale, R.; Thulasinathan, T.; Ayyenar, B.; Muthurajan, R. Pyramiding QTLs Controlling Tolerance against Drought, Salinity, and Submergence in Rice Through Marker Assisted Breeding. PLoS ONE 2020, 15, e0227421. [Google Scholar] [CrossRef] [PubMed]
- Abe, T.; Ito, M.; Takahashi, R.; Honma, T.; Kuramata, M.; Ishikawa, S. QTL Pyramiding and Its Use in Breeding for Increasing the Phytoextraction Efficiency of Soil Cd via High-Cd-Accumulating Rice. Plants 2022, 11, 2178. [Google Scholar] [CrossRef] [PubMed]
- Bhusal, N.; Sharma, P.; Kumar, R.R.; Sareen, S. Multiple Abiotic Stresses: Molecular, Physiological, and Genetic Responses and Adaptations in Cereals. Front. Plant Sci. 2023, 14, 1146326. [Google Scholar] [CrossRef] [PubMed]
- Gautam, T.; Amardeep; Saripalli, G.; Rakhi; Kumar, A.; Gahlaut, V.; Gadekar, D.A.; Oak, M.; Sharma, P.K.; Balyan, H.S.; et al. Introgression of a Drought Insensitive Grain Yield QTL for Improvement of Four Indian Bread Wheat Cultivars Using Marker Assisted Breeding Without Background Selection. J. Plant Biochem. Biotechnol. 2021, 30, 172–183. [Google Scholar] [CrossRef]
- Ye, C.; Ishimaru, T.; Lambio, L.; Li, L.; Long, Y.; He, Z.; Htun, T.M.; Tang, S.; Su, Z. Marker-Assisted Pyramiding of Qtls For Heat Tolerance And Escape Upgrades Heat Resilience In Rice (Oryza sativa L.). Theor. Appl. Genet. 2022, 135, 1345–1354. [Google Scholar] [CrossRef]
- Mohd Ikmal, A.; Noraziyah, A.A.S.; Wickneswari, R.; Oyebamiji, Y.O. Sub1 and qDTY 3.1 Improved Tolerance of Rice (Oryza sativa L.) Lines to Drought and Submergence Stresses. Euphytica 2023, 219, 60. [Google Scholar] [CrossRef]
- Zhang, J.; McDonald, S.C.; Wu, C.; Ingwers, M.W.; Abdel-Haleem, H.; Chen, P.; Li, Z. Quantitative Trait Loci Underlying Flooding Tolerance In Soybean (Glycine max). Plant Breed. 2022, 141, 236–245. [Google Scholar] [CrossRef]
- Rani, M.G.; Satyanarayana, P.V.; Chamundeswari, N.; Rao, P.R.; Prabhakar, M.; Ravikumar, B.N.V.S.R.; Nagakumari, P.; Kalpana, K. Mapping of QTLs for Flood Tolerance in Rice Using Recombinant Inbred Lines of Indra and A New Plant Genetic Resource AC 39416 A. Plant Genet. Resour. 2022, 20, 270–276. [Google Scholar] [CrossRef]
- Ding, F.; Tong, J.; Xu, R.; Chen, J.; Xu, X.; Nadeem, M.; Wang, S.; Zhang, Y.; Zhu, Z.; Wang, F.; et al. Identification of Stable Quantitative Trait Loci Underlying Waterlogging Tolerance Post-Anthesis in Common Wheat (Triticum aestivum). Crop J. 2023, 11, 1163–1170. [Google Scholar] [CrossRef]
- Sarkar, B.; Varalaxmi, Y.; Vanaja, M.; RaviKumar, N.; Prabhakar, M.; Yadav, S.K.; Maheswari, M.; Singh, V.K. Mapping of QTLs for Morphophysiological and Yield Traits Under Water-Deficit Stress and Well-Watered Conditions in Maize. Front. Plant Sci. 2023, 14, 1124619. [Google Scholar] [CrossRef]
- Katoch, M.; Chahota, R.K. QTLs Identification for Biochemical and Physiological Parameters Contributing towards Drought Tolerance in Horsegram (Macrotyloma uniflorum). Appl. Biochem. Biotechnol. 2023; preprint. [Google Scholar] [CrossRef]
- Yadava, Y.K.; Chaudhary, P.; Yadav, S.; Rizvi, A.H.; Kumar, T.; Srivastava, R.; Soren, K.R.; Bharadwaj, C.; Srinivasan, R.; Singh, N.K.; et al. Genetic Mapping of Quantitative Trait Loci Associated with Drought Tolerance In Chickpea (Cicer arietinum L.). Sci. Rep. 2023, 13, 17623. [Google Scholar] [CrossRef]
- Diouf, I.A.; Derivot, L.; Bitton, F.; Pascual, L.; Causse, M. Water Deficit and Salinity Stress Reveal Many Specific QTL for Plant Growth and Fruit Quality Traits in Tomato. Front. Plant Sci. 2018, 9, 279. [Google Scholar] [CrossRef]
- Dramadri, I.O.; Nkalubo, S.T.; Kelly, J.D. Identification of QTL Associated with Drought Tolerance in Andean Common Bean. Crop Sci. 2019, 59, 1007–1020. [Google Scholar] [CrossRef]
- Fan, Y.; Shabala, S.; Ma, Y.; Xu, R.; Zhou, M. Using QTL Mapping to Investigate the Relationships between Abiotic Stress Tolerance (Drought and Salinity) and Agronomic and Physiological Traits. BMC Genom. 2015, 16, 43. [Google Scholar] [CrossRef] [PubMed]
- Diaz, L.M.; Ricaurte, J.; Tovar, E.; Cajiao, C.; Teran, H.; Grajales, M.; Polanía, J.; Rao, I.; Beebe, S.; Raatz, B. QTL Analyses for Tolerance to Abiotic Stresses in a Common Bean (Phaseolus vulgaris L.) Population. PLoS ONE 2018, 13, e0202342. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.; Hassan, M.A.; Cheng, X.; Li, Y.; Liu, H.; Fang, W.; Zhu, Q.; Wang, S. QTL mapping and analysis for drought tolerance in rice by genome-wide association study. Front. Plant Sci. 2023, 14, 1223782. [Google Scholar] [CrossRef]
- Pandey, M.K.; Gangurde, S.S.; Sharma, V.; Pattanashetti, S.K.; Naidu, G.K.; Faye, I.; Hamidou, F.; Desmae, H.; Kane, N.A.; Yuan, M.; et al. Improved Genetic Map Identified Major QTLs for Drought Tolerance- and Iron Deficiency Tolerance-Related Traits in Groundnut. Genes 2021, 12, 37. [Google Scholar] [CrossRef] [PubMed]
- Soren, K.R.; Madugula, P.; Kumar, N.; Barmukh, R.; Sengar, M.S.; Bharadwaj, C.; Sharma, P.C.; Singh, S.; Bhandari, A.; Singh, J.; et al. Genetic Dissection and Identification of Candidate Genes for Salinity Tolerance Using Axiom® Cicersnp Array in Chickpea. Int. J. Mol. Sci. 2020, 21, 5058. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Zhu, Y.; Chen, K.; Shen, C.; Zhao, X.; Shabala, S.; Shabala, L.; Meinke, H.; Venkataraman, G.; Chen, Z.-H.; et al. Identification of New QTL for Salt Tolerance from Rice Variety Pokkali. J. Agron. Crop Sci. 2020, 206, 202–213. [Google Scholar] [CrossRef]
- Wang, H.; Wang, R.; Liu, B.; Yang, Y.; Qin, L.; Chen, E.; Zhang, H.; Guan, Y. QTL Analysis of Salt Tolerance in Sorghum bicolor During Whole-Plant Growth Stages. Plant Breed. 2020, 139, 455–465. [Google Scholar] [CrossRef]
- Patel, R.; Jangra, S.; Avtar, R.; Yadav, N.R.; Yadav, R.C. Genetic Linkage Mapping and QTL Identification for Salinity Tolerance in Indian Mustard (Brassica juncea L. Czern and Coss.) using SSR markers. Oil Crop Sci. 2023, 8, 191–205. [Google Scholar] [CrossRef]
- Liu, D.; Dong, S.; Bo, K.; Miao, H.; Li, C.; Zhang, Y.; Zhang, S.; Gu, X. Identification of QTLs Controlling Salt Tolerance in Cucumber (Cucumis sativus L.) seedlings. Plants 2021, 10, 85. [Google Scholar] [CrossRef]
- Yang, Z.; Yang, Y.; Dai, Z.; Xie, D.; Tang, Q.; Cheng, C.; Xu, Y.; Liu, C.; Deng, C.; Chen, J.; et al. Construction of A High-Resolution Genetic Map and Identification of Quantitative Trait Loci for Salt Tolerance In Jute (Corchous spp.). BMC Plant Biol. 2019, 19, 391. [Google Scholar] [CrossRef]
- Jha, U.C.; Nayyar, H.; Palakurthi, R.; Jha, R.; Valluri, V.; Bajaj, P.; Chitikineni, A.; Singh, N.P.; Varshney, R.K.; Thudi, M. Major QTLs and Potential Candidate Genes for Heat Stress Tolerance Identified in Chickpea (Cicer arietinum L.). Front. Plant Sci. 2021, 12, 655103. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Jiang, F.; Weng, Y.; Sun, M.; Shi, X.; Zhou, Y.; Yu, L.; Wu, Z. Identification of Heat-Tolerance Qtls and High-Temperature Stress-Responsive Genes through Conventional QTL Mapping, QTL-seq and RNA-seq in Tomato. BMC Plant Biol. 2019, 19, 398. [Google Scholar] [CrossRef] [PubMed]
- Rani, S.; Baber, M.; Naqqash, T.; Malik, S.A. Identification and Genetic Mapping of Potential Qtls Conferring Heat Tolerance in Cotton (Gossypium hirsutum L.) By Using Micro Satellite Marker’s Approach. Agronomy 2022, 12, 1381. [Google Scholar] [CrossRef]
- Liu, Y.; Dong, S.; Wei, S.; Wang, W.; Miao, H.; Bo, K.; Gu, X.; Zhang, S. QTL Mapping of Heat Tolerance in Cucumber (Cucumis sativus L.) At Adult Stage. Plants 2021, 10, 324. [Google Scholar] [CrossRef]
- Touzy, G.; Lafarge, S.; Redondo, E.; Lievin, V.; Decoopman, X.; Le Gouis, J.; Praud, S. Identification of QTLs Affecting Post-Anthesis Heat Stress Responses in European Bread Wheat. Theor. Appl. Genet. 2022, 135, 947–964. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Huang, Y.; Gu, B. QTL-Seq Identifies Quantitative Trait Loci of Relative Electrical Conductivity Associated with Heat Tolerance in Bottle Gourd (Lagenaria siceraria). PLoS ONE 2020, 15, e0227663. [Google Scholar] [CrossRef]
- Li, C.; Liu, J.; Bian, J.; Jin, T.; Zou, B.; Liu, S.; Zhang, X.; Wang, P.; Tan, J.; Wu, G.; et al. Identification of Cold Tolerance Qtls at The Bud Burst Stage in 211 Rice Landraces by GWAS. BMC Plant Biol. 2021, 21, 542. [Google Scholar] [CrossRef]
- Adhikari, L.; Makaju, S.O.; Lindstrom, O.M.; Missaoui, A.M. Mapping Freezing Tolerance QTL in Alfalfa: Based on Indoor Phenotyping. BMC Plant Biol. 2021, 21, 403. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, X.; Wang, L.; Liu, Q.; Liang, Y.; Zhang, J.; Xue, Y.; Tian, Y.; Zhang, H.; Li, N.; et al. Fine Mapping of A QTL And Identification of Candidate Genes Associated with Cold Tolerance During Germination in Peanut (Arachis hypogaea L.) On Chromosome B09 Using Whole Genome Re-Sequencing. Front. Plant Sci. 2023, 14, 1153293. [Google Scholar] [CrossRef]
- Murugaiyan, V.; Ali, J.; Mahender, A.; Aslam, U.M.; Jewel, Z.A.; Pang, Y.; Marfori-Nazarea, C.M.; Wu, L.-B.; Frei, M.; Li, Z. Mapping of Genomic Regions Associated with Arsenic Toxicity Stress in A Backcross Breeding Populations of Rice (Oryza sativa L.). Rice 2019, 12, 61. [Google Scholar] [CrossRef]
- Jegadeesan, S.; Yu, K.; Poysa, V.; Gawalko, E.; Morrison, M.J.; Shi, C.; Cober, E. Mapping and Validation of Simple Sequence Repeat Markers Linked to A Major Gene Controlling Seed Cadmium Accumulation in Soybean [Glycine max (L.) Merr]. Theor. Appl. Genet. 2010, 121, 283–294. [Google Scholar] [CrossRef]
- Wu, J.; Yuan, Y.X.; Zhang, X.W.; Zhao, J.; Song, X.; Li, Y.; Li, X.; Sun, R.; Koornnneef, M.; Aarts, M.G.M.; et al. Mapping QTLs for Mineral Accumulation and Shoot Dry Biomass under Different Zn Nutritional Conditions in Chinese Cabbage (Brassica rapa L. ssp. pekinensis). Plant Soil 2008, 310, 25–40. [Google Scholar] [CrossRef]
- Ding, D.; Li, W.; Song, G.; Qi, H.; Liu, J.; Tang, J. Identification of QTLs for Arsenic Accumulation in Maize (Zea mays L.) Using a RIL population. PLoS ONE 2011, 6, e25646. [Google Scholar] [CrossRef]
- Hossain, A.; Skalicky, M.; Brestic, M.; Maitra, S.; Ashraful Alam, M.; Syed, M.A.; Islam, T. Consequences and mitigation strategies of abiotic stresses in wheat (Triticum aestivum L.) under the changing climate. Agronomy 2021, 11, 241. [Google Scholar] [CrossRef]
- Adhikari, L.; Baral, R.; Paudel, D.; Min, D.; Makaju, S.O.; Poudel, H.P.; Acharya, J.P.; Missaoui, A.M. Cold stress in plants: Strategies to improve cold tolerance in forage species. Plant Stress 2022, 4, 100081. [Google Scholar] [CrossRef]
- Ding, Y.; Gong, S.; Wang, Y.; Wang, F.; Bao, H.; Sun, J.; Cai, C.; Yi, K.; Chen, Z.; Zhu, C. MicroRNA166 Modulates Cadmium Tolerance and Accumulation in Rice. Plant Physiol. 2018, 177, 1691–1703. [Google Scholar] [CrossRef]
- Krishna, R.; Karkute, S.G.; Ansari, W.A.; Jaiswal, D.K.; Verma, J.P.; Singh, M. Transgenic Tomatoes for Abiotic Stress Tolerance: Status and Way Ahead. 3 Biotech 2019, 9, 143. [Google Scholar] [CrossRef]
- Key, S.; Ma, J.K.; Drake, P.M. Genetically modified plants and human health. J. R. Soc. Med. 2008, 101, 290–298. [Google Scholar] [CrossRef]
- Haider, Z.; Ahmad, I.; Zia, S.; Gan, Y. Recent Developments in Rice Molecular Breeding for Tolerance to Heavy Metal Toxicity. Agriculture 2023, 13, 944. [Google Scholar] [CrossRef]
- Kikuchi, A.; Huynh, H.D.; Endo, T.; Watanabe, K. Review of recent transgenic studies on abiotic stress tolerance and future molecular breeding in potato. Breed. Sci. 2015, 65, 85–102. [Google Scholar] [CrossRef]
- Gangadhar, B.H.; Mishra, R.K.; Kappachery, S.; Baskar, V.; Venkatesh, J.; Nookaraju, A.; Thiruvengadam, M. Enhanced Thermo-Tolerance in Transgenic Potato (Solanum tuberosum L.) Overexpressing Hydrogen Peroxide-Producing Germin-like Protein (GLP). Genomics 2021, 113, 3224–3234. [Google Scholar] [CrossRef]
- Liang, S.; Xiong, W.; Yin, C.; Xie, X.; Jin, Y.J.; Zhang, S.; Yang, B.; Ye, G.; Chyen, S.; Luan, W.J. Overexpression of OsARD1 Improves Submergence, Drought, And Salt Tolerances of Seedling through the Enhancement of Ethylene Synthesis in Rice. Front. Plant Sci. 2019, 10, 1088. [Google Scholar] [CrossRef]
- Bi, H.; Zhao, Y.; Li, H.; Liu, W. Wheat Heat Shock Factor Tahsfa6f Increases ABA Levels and Enhances Tolerance to Multiple Abiotic Stresses in Transgenic Plants. Int. J. Mol. Sci. 2020, 21, 3121. [Google Scholar] [CrossRef]
- Zhang, X.; Sun, Y.; Qiu, X.; Lu, H.; Hwang, I.; Wang, T. Tolerant Mechanism of ModelLegume Plant Medicago truncatula to Drought, Salt, and Cold Stresses. Front. Plant Sci. 2022, 13, 847166. [Google Scholar] [CrossRef]
- Kumar, V.; AlMomin, S.; Al-Shatti, A.; Al-Aqeel, H.; Al-Salameen, F.; Shajan, A.B.; Nair, S.M. Enhancement of Heavy Metal Tolerance and Accumulation Efficiency by Expressing Arabidopsis ATP Sulfurylase Gene in Alfalfa. Int. J. Phytoremediat. 2019, 21, 1112–1121. [Google Scholar] [CrossRef]
- Mu, J.; Fu, Y.; Liu, B.; Zhang, Y.; Wang, A.; Li, Y.; Zhu, J. SiFBA5, A Cold-Responsive Factor from Saussurea involucrata Promotes Cold Resilience and Biomass Increase in Transgenic Tomato Plants under Cold Stress. BMC Plant Biol. 2021, 21, 75. [Google Scholar] [CrossRef]
- Afzal, M.; Hindawi, S.E.S.; Alghamdi, S.S.; Migdadi, H.H.; Khan, M.A.; Hasnain, M.U.; Arslan, M.; Sohaib, M. Potential Breeding Strategies for Improving Salt Tolerance in Crop Plants. J. Plant Growth Regul. 2023, 42, 3365–3387. [Google Scholar] [CrossRef]
- Zeng, Y.; Wen, J.; Zhao, W.; Wang, Q.; Huang, W. Rational Improvement of Rice Yield and Cold Tolerance by Editing the Three Genes OsPIN5b, GS3, and OsMYB30 with the CRISPR–Cas9 system. Front. Plant Sci. 2020, 10, 1663. [Google Scholar] [CrossRef]
- Ahmar, S.; Saeed, S.; Khan, M.H.U.; Ullah Khan, S.; Mora-Poblete, F.; Kamran, M.; Faheem, A.; Maqsood, A.; Rauf, M.; Saleem, S.; et al. A Revolution toward Gene-Editing Technology and Its Application to Crop Improvement. Int. J. Mol. Sci. 2020, 21, 5665. [Google Scholar] [CrossRef]
- Kushwaha, A.K.; Shiv, A.; Samantara, K.; Chand, S.; Majhi, P.K.; Goyal, L.; Kaldate, S.; Kumar, S.; Razzaq, A.; Wani, S.H. Editing genomes to modify plant response to abiotic stress. In Plant Stress Mitigators; Academic Press: Chennai, India, 2023; pp. 1–15. [Google Scholar]
- Kumari, A.; Singh, A.P. Speed Breeding: Accelerating Crop Improvement for Global Food Security. Innov. Approaches Agric. 2022, 1, 92–103. [Google Scholar]
- Haroon, M.; Zafar, M.M.; Farooq, M.A.; Afzal, R.; Batool, M.; Idrees, F.; Babar, U.; Khan, A.S.; Mo, H.; Li, L.; et al. Conventional Breeding, Molecular Breeding and Speed Breeding; Brave Approaches to Revamp the Production of Cereal Crops. Plants 2022, 11, 1052. [Google Scholar] [CrossRef]
- Samantara, K.; Bohra, A.; Mohapatra, S.R.; Prihatini, R.; Asibe, F.; Singh, L.; Reyes, V.P.; Tiwari, A.; Maurya, A.K.; Croser, J.S.; et al. Breeding More Crops in Less Time: A Perspective on Speed Breeding. Biology 2022, 11, 275. [Google Scholar] [CrossRef]
- Watson, A.; Ghosh, S.; Williams, M.J.; Cuddy, W.S.; Simmonds, J.; Rey, M.D.; Md Hatta, M.A.; Hinchliffe, A.; Steed, A.; Reynolds, D.; et al. Speed Breeding is APowerful Tool to Accelerate Crop Research And Breeding. Nat. Plants 2018, 4, 23–29. [Google Scholar] [CrossRef]
- Zeroual, A.; Baidani, A.; Idrissi, O. Drought stress in lentil (Lens culinaris, medik) and approaches for its management. Horticulturae 2022, 9, 1. [Google Scholar] [CrossRef]
- Pandey, S.; Singh, A.; Parida, S.K.; Prasad, M. Combining speed breeding with traditional and genomics-assisted breeding for crop improvement. Plant Breed. 2022, 141, 301–313. [Google Scholar] [CrossRef]
- Wanga, M.A.; Shimelis, H.; Mashilo, J.; Laing, M.D. Opportunities and challenges of speed breeding: A review. Plant Breed. 2021, 140, 185–194. [Google Scholar] [CrossRef]
- Hickey, L.T.; Germán, S.E.; Pereyra, S.A.; Diaz, J.E.; Ziems, L.A.; Fowler, R.A.; Dieters, M.J. Speed breeding for multiple disease resistance in barley. Euphytica 2017, 213, 64. [Google Scholar] [CrossRef]
- Rana, M.M.; Takamatsu, T.; Baslam, M.; Kaneko, K.; Itoh, K.; Harada, N.; Mitsui, T. Salt tolerance improvement in rice through efficient SNP marker-assisted selection coupled with speed-breeding. Int. J. Mol. Sci. 2019, 20, 2585. [Google Scholar] [CrossRef]
- Raza, A.; Mubarik, M.S.; Sharif, R.; Habib, M.; Jabeen, W.; Zhang, C.; Chen, H.; Chen, Z.-H.; Siddique, K.H.M.; Zhuang, W.; et al. Developing drought-smart, ready-to-grow future crops. Plant Genome 2023, 16, e20279. [Google Scholar] [CrossRef]
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Hussain, M.A.; Wei, S.; He, H.; Hasanuzzaman, M. Omics: The way forward to enhance abiotic stress tolerance in Brassica napus L. GM Crops Food 2021, 12, 251–281. [Google Scholar] [CrossRef]
- Rhaman, M.S.; Imran, S.; Rauf, F.; Khatun, M.; Baskin, C.C.; Murata, Y.; Hasanuzzaman, M. Seed priming with phytohormones: An effective approach for the mitigation of abiotic stress. Plants 2020, 10, 37. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef]
- Biswas, S.; Seal, P.; Majumder, B.; Biswas, A.K. Efficacy of seed priming strategies for enhancing salinity tolerance in plants: An overview of the progress and achievements. Plant Stress 2023, 9, 100186. [Google Scholar] [CrossRef]
- Kumar, A.; Nayak, A.K.; Hanjagi, P.S.; Kumari, K.; Vijayakumar, S.; Mohanty, S.; Panneerselvam, P. Submergence stress in rice: Adaptive mechanisms, coping strategies and future research needs. Environ. Exp. Bot. 2021, 186, 104448. [Google Scholar] [CrossRef]
- Arun, M.N.; Hebbar, S.S.; Senthivel, T.; Nair, A.K.; Padmavathi, G.; Pandey, P.; Singh, A. Seed Priming: The Way Forward to Mitigate Abiotic Stress in Crops; IntechOpen: London, UK, 2022; Volume 11, p. 173. [Google Scholar]
- Rajora, N.; Vats, S.; Raturi, G.; Thakral, V.; Kaur, S.; Rachappanavar, V.; Deshmukh, R. Seed priming with melatonin: A promising approach to combat abiotic stress in plants. Plant Stress 2022, 4, 100071. [Google Scholar] [CrossRef]
- Rhaman, M.S.; Tania, S.S.; Imran, S.; Rauf, F.; Kibria, M.G.; Ye, W.; Murata, Y. Seed priming with nanoparticles: An emerging technique for improving plant growth, development, and abiotic stress tolerance. J. Soil Sci. Plant Nutr. 2022, 22, 4047–4062. [Google Scholar] [CrossRef]
- Nouri, M.; Haddioui, A. Improving seed germination and seedling growth of Lepidium sativum with different priming methods under arsenic stress. Acta Ecol. Sin. 2021, 41, 64–71. [Google Scholar] [CrossRef]
- Sytar, O.; Kumari, P.; Yadav, S.; Brestic, M.; Rastogi, A. Phytohormone priming: Regulator for heavy metal stress in plants. J. Plant Growth Regul. 2019, 38, 739–752. [Google Scholar] [CrossRef]
- Salam, A.; Khan, A.R.; Liu, L.; Yang, S.; Azhar, W.; Ulhassan, Z.; Gan, Y. Seed priming with zinc oxide nanoparticles downplayed ultrastructural damage and improved photosynthetic apparatus in maize under cobalt stress. J. Hazard. Mater. 2022, 423, 127021. [Google Scholar] [CrossRef]
- Sheteiwy, M.S.; An, J.; Yin, M.; Jia, X.; Guan, Y.; He, F.; Hu, J. Cold plasma treatment and exogenous salicylic acid priming enhances salinity tolerance of Oryza sativa seedlings. Protoplasma 2019, 256, 79–99. [Google Scholar] [CrossRef]
- Marthandan, V.; Geetha, R.; Kumutha, K.; Renganathan, V.G.; Karthikeyan, A.; Ramalingam, J. Seed priming: A feasible strategy to enhance drought tolerance in crop plants. Int. J. Mol. Sci. 2020, 21, 8258. [Google Scholar] [CrossRef] [PubMed]
- Langeroodi, A.R.S.; Noora, R. Seed priming improves the germination and field performance of soybean under drought stress. J. Anim. Plant Sci. 2017, 27, 1611–1620. [Google Scholar]
- Vwioko, E.D.; El-Esawi, M.A.; Imoni, M.E.; Al-Ghamdi, A.A.; Ali, H.M.; El-Sheekh, M.M.; Al-Dosary, M.A. Sodium azide priming enhances waterlogging stress tolerance in okra (Abelmoschus esculentus L.). Agronomy 2019, 9, 679. [Google Scholar] [CrossRef]
- Iqbal, H.; Yaning, C.; Waqas, M.; Ahmed, Z.; Raza, S.T.; Shareef, M. Improving heat stress tolerance in late planted spring maize by using different exogenous elicitors. Chil. J. Agric. Res. 2020, 80, 30–40. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, L.; Rusalepp, L.; Kaurilind, E.; Sulaiman, H.Y.; Püssa, T.; Niinemets, Ü. Heat priming improved heat tolerance of photosynthesis, enhanced terpenoid and benzenoid emission and phenolics accumulation in Achillea millefolium. Plant Cell Environ. 2021, 44, 2365–2385. [Google Scholar] [CrossRef]
- Serrano, N.; Ling, Y.; Bahieldin, A.; Mahfouz, M.M. Thermopriming reprograms metabolic homeostasis to confer heat tolerance. Sci. Rep. 2019, 9, 181. [Google Scholar] [CrossRef]
- Zhu, Z.H.; Sami, A.; Xu, Q.Q.; Wu, L.L.; Zheng, W.Y.; Chen, Z.P.; Zhou, K.J. Effects of seed priming treatments on the germination and development of two rapeseed (Brassica napus L.) varieties under the co-influence of low temperature and drought. PLoS ONE 2021, 16, e0257236. [Google Scholar] [CrossRef]
- Cao, Q.; Li, G.; Cui, Z.; Yang, F.; Jiang, X.; Diallo, L.; Kong, F. Seed priming with melatonin improves the seed germination of waxy maize under chilling stress via promoting the antioxidant system and starch metabolism. Sci. Rep. 2019, 9, 15044. [Google Scholar] [CrossRef]
- Cardarelli, M.; Woo, S.L.; Rouphael, Y.; Colla, G. Seed treatments with microorganisms can have a biostimulant effect by influencing germination and seedling growth of crops. Plants 2022, 11, 259. [Google Scholar] [CrossRef]
- Lastochkina, O.; Garshina, D.; Ivanov, S.; Yuldashev, R.; Khafizova, R.; Allagulova, C.; Bosacchi, M. Seed priming with endophytic Bacillus subtilis modulates physiological responses of two different Triticum aestivum L. cultivars under drought stress. Plants 2020, 9, 1810. [Google Scholar] [CrossRef]
- Paravar, A.; Piri, R.; Balouchi, H.; Ma, Y. Microbial seed coating: An attractive tool for sustainable agriculture. Biotechnol. Rep. 2023, 37, e00781. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Mahapatra, S.; Chakraborty, S.; Mukharjee, S. Effect of abiotic stresses and mitigation strategy associated with their tolerance in wheat. J. Cereal Res. 2021, 13, 16–37. [Google Scholar] [CrossRef]
- Alayafi, A.A.M. Exogenous ascorbic acid induces systemic heat stress tolerance in tomato seedlings: Transcriptional regulation mechanism. Environ. Sci. Pollut. Res. 2020, 27, 19186–19199. [Google Scholar] [CrossRef] [PubMed]
- Godoy, F.; Olivos-Hernández, K.; Stange, C.; Handford, M. Abiotic stress in crop species: Improving tolerance by applying plant metabolites. Plants 2021, 10, 186. [Google Scholar] [CrossRef]
- Devireddy, A.R.; Rivero, R.M.; Zandalinas, S.I. Rising stars in plant ROS/redox biology under abiotic stress conditions. Front. Plant Sci. 2023, 14, 1207275. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Waraich, E.A.; Skalicky, M.; Hussain, S.; Zulfiqar, U.; Anjum, M.Z.; El Sabagh, A. Adaptation strategies to improve the resistance of oilseed crops to heat stress under a changing climate: An overview. Front. Plant Sci. 2021, 12, 767150. [Google Scholar] [CrossRef]
- Rady, M.M.; Boriek, S.H.; Abd El-Mageed, T.A.; Seif El-Yazal, M.A.; Ali, E.F.; Hassan, F.A.; Abdelkhalik, A. Exogenous gibberellic acid or dilute bee honey boosts drought stress tolerance in Vicia faba by rebalancing osmoprotectants, antioxidants, nutrients, and phytohormones. Plants 2021, 10, 748. [Google Scholar] [CrossRef]
- Mishra, U.N.; Saha, D.; Chauhan, J.; Kumar, V.; Jatav, H.S.; Lal, D.; Chandra, K. Emerging Roles of Osmoprotectants in Response to Multiple Abiotic Stress Tolerance in Plants. In Omics Analysis of Plants under Abiotic Stress; Apple Academic Press: Waretown, NJ, USA, 2022; pp. 179–206. [Google Scholar]
- Islam, M.R.; Rahman, M.M.; Mohi-Ud-Din, M.; Akter, M.; Zaman, E.; Keya, S.S.; Hasanuzzaman, M. Cytokinin and gibberellic acid-mediated waterlogging tolerance of mungbean (Vigna radiata L. Wilczek). PeerJ 2022, 10, e12862. [Google Scholar] [CrossRef]
- Ren, B.; Hu, J.; Zhang, J.; Dong, S.; Liu, P.; Zhao, B. Spraying exogenous synthetic cytokinin 6-benzyladenine following the waterlogging improves grain growth of waterlogged maize in the field. J. Agron. Crop Sci. 2019, 205, 616–624. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Ahmed, N.; Saha, T.; Rahman, M.; Rahman, K.; Alam, M.M.; Nahar, K. Exogenous salicylic acid and kinetin modulate reactive oxygen species metabolism and glyoxalase system to confer waterlogging stress tolerance in soybean (Glycine max L.). Plant Stress 2022, 3, 100057. [Google Scholar] [CrossRef]
- Lakshmi, G.; Beena, R.; Soni, K.B.; Viji, M.M.; Jha, U.C. Exogenously applied plant growth regulator protects rice from heat-induced damage by modulating plant defense mechanism. J. Crop Sci. Biotechnol. 2023, 26, 63–75. [Google Scholar] [CrossRef]
- Hussain, R.; Ayyub, C.M.; Shaheen, M.R.; Rashid, S.; Nafees, M.; Ali, S.; Mora-Poblete, F. Regulation of osmotic balance and increased antioxidant activities under heat stress in Abelmoschus esculentus L. triggered by exogenous proline application. Agronomy 2021, 11, 685. [Google Scholar] [CrossRef]
- Saini, S.; Kaur, N.; Pati, P.K. Phytohormones: Key players in the modulation of heavy metal stress tolerance in plants. Ecotoxicol. Environ. Saf. 2021, 223, 112578. [Google Scholar] [CrossRef]
- Zaid, A.; Mohammad, F.; Fariduddin, Q. Plant growth regulators improve growth, photosynthesis, mineral nutrient and antioxidant system under cadmium stress in menthol mint (Mentha arvensis L.). Physiol. Mol. Biol. Plants 2020, 26, 25–39. [Google Scholar] [CrossRef]
- Raza, A.; Charagh, S.; Najafi-Kakavand, S.; Abbas, S.; Shoaib, Y.; Anwar, S.; Siddique, K.H. Role of phytohormones in regulating cold stress tolerance: Physiological and molecular approaches for developing cold-smart crop plants. Plant Stress 2023, 8, 100152. [Google Scholar] [CrossRef]
- Jankovska-Bortkevič, E.; Katerova, Z.; Todorova, D.; Jankauskienė, J.; Mockevičiūtė, R.; Sergiev, I.; Jurkonienė, S. Effects of Auxin-Type Plant Growth Regulators and Cold Stress on the Endogenous Polyamines in Pea Plants. Horticulturae 2023, 9, 244. [Google Scholar] [CrossRef]
- Hmmam, I.; Raza, A.; Djalovic, I.; Khedr, N.; Abdellatif, A. An in Vitro Approach to Investigate the Role of Abscisic Acid in Alleviating the Negative Effects of Chilling Stress on Banana Shoots. Phyton-Int. J. Exp. Bot. 2023, 92, 1–17. [Google Scholar] [CrossRef]
- Omoarelojie, L.O.; Kulkarni, M.G.; Finnie, J.F.; Van Staden, J. Strigolactone analog (rac-GR24) enhances chilling tolerance in mung bean seedlings. S. Afr. J. Bot. 2021, 140, 173–181. [Google Scholar] [CrossRef]
- Zeng, W.; Mostafa, S.; Lu, Z.; Jin, B. Melatonin-mediated Abiotic Stress Tolerance in Plants. Front. Plant Sci. 2022, 13, 847175. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Ali, S.; Manghwar, H.; Saqib, S.; Ullah, F.; Ayaz, A.; Zaman, W. Melatonin function and crosstalk with other phytohormones under normal and stressful conditions. Genes 2022, 13, 1699. [Google Scholar] [CrossRef] [PubMed]
- Colombage, R.; Singh, M.B.; Bhalla, P.L. Melatonin and Abiotic Stress Tolerance in Crop Plants. Int. J. Mol. Sci. 2023, 24, 7447. [Google Scholar] [CrossRef] [PubMed]
- Rehaman, A.; Mishra, A.K.; Ferdose, A.; Per, T.S.; Hanief, M.; Jan, A.T.; Asgher, M. Melatonin in plant defense against abiotic stress. Forests 2021, 12, 1404. [Google Scholar] [CrossRef]
- Hoque, M.N.; Tahjib-Ul-Arif, M.; Hannan, A.; Sultana, N.; Akhter, S.; Hasanuzzaman, M.; Akter, F.; Hossain, M.S.; Sayed, M.A.; Hasan, M.T.; et al. Melatonin modulates plant tolerance to heavy metal stress: Morphological responses to molecular mechanisms. Int. J. Mol. Sci. 2021, 22, 11445. [Google Scholar] [CrossRef] [PubMed]
- Pardo-Hernández, M.; López-Delacalle, M.; Rivero, R.M. ROS and NO regulation by melatonin under abiotic stress in plants. Antioxidants 2020, 9, 1078. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.U.; Mahmood, A.; Awan, M.I.; Maqbool, R.; Aamer, M.; Alhaithloul, H.A.S.; Huang, G.; Skalicky, M.; Brestic, M.; Pandey, S.; et al. Melatonin-Induced Protection against Plant Abiotic Stress: Mechanisms and Prospects. Front. Plant Sci. 2022, 13, 902694. [Google Scholar] [CrossRef]
- Ali, S.; Moon, Y.S.; Hamayun, M.; Khan, M.A.; Bibi, K.; Lee, I.J. Pragmatic role of microbial plant biostimulants in abiotic stress relief in crop plants. J. Plant Interact. 2022, 17, 705–718. [Google Scholar] [CrossRef]
- Behrooz, A.; Vahdati, K.; Rejali, F.; Lotfi, M.; Sarikhani, S.; Leslie, C. Arbuscular mycorrhiza and plant growth-promoting bacteria alleviate drought stress in walnut. HortScience 2019, 54, 1087–1092. [Google Scholar] [CrossRef]
- Ruiz-Sánchez, M.; Aroca, R.; Muñoz, Y.; Polón, R.; Ruiz-Lozano, J.M. The arbuscular mycorrhizal symbiosis enhances the photosynthetic efficiency and the antioxidative response of rice plants subjected to drought stress. J. Plant Physiol. 2010, 167, 862–869. [Google Scholar] [CrossRef]
- Sun, Z.; Song, J.; Xin, X.A.; Xie, X.; Zhao, B. Arbuscular mycorrhizal fungal 14-3-3 proteins are involved in arbuscule formation and responses to abiotic stresses during AM symbiosis. Front. Microbiol. 2018, 9, 91. [Google Scholar] [CrossRef]
- Langeroodi, A.R.S.; Osipitan, O.A.; Radicetti, E.; Mancinelli, R. To what extent Arbuscular mycorrhiza can protect chicory (Cichorium intybus L.) against Drought Stress. Sci. Hortic. 2020, 263, 109109. [Google Scholar] [CrossRef]
- Hassan, Z.U.; Ali, S.; Rizwan, M.; Ibrahim, M.; Nafees, M.; Waseem, M. Role of bioremediation agents (bacteria, fungi, and algae) in alleviating heavy metal toxicity. In Probiotics in Agroecosystems; Springer: Singapore, 2017; pp. 517–537. [Google Scholar]
- Jumrani, K.; Bhatia, V.S.; Kataria, S.; Alamri, S.A.; Siddiqui, M.H.; Rastogi, A. Inoculation with arbuscular mycorrhizal fungi alleviates the adverse effects of high temperature in soybean. Plants 2022, 11, 2210. [Google Scholar] [CrossRef]
- Tang, C.; Zhang, Z.; Yu, L.; Li, Y. Research Progress of Arbuscular Mycorrhizal Fungi Promoting Citrus Growth. Horticulturae 2023, 9, 1162. [Google Scholar] [CrossRef]
- Xu, J.; Liu, S.; Song, S.; Guo, H.; Tang, J.; Yong, J.W.; Chen, X. Arbuscular mycorrhizal fungi influence decomposition and the associated soil microbial community under different soil phosphorus availability. Soil Biol. Biochem. 2018, 120, 181–190. [Google Scholar] [CrossRef]
- Khatun, M.; Sarkar, S.; Era, F.M.; Islam, A.M.; Anwar, M.P.; Fahad, S.; Islam, A.A. Drought stress in grain legumes: Effects, tolerance mechanisms and management. Agronomy 2021, 11, 2374. [Google Scholar] [CrossRef]
- Formisano, L.; Ciriello, M.; Cirillo, V.; Pannico, A.; El-Nakhel, C.; Cristofano, F.; De Pascale, S. Divergent Leaf Morpho-Physiological and Anatomical Adaptations of Four Lettuce Cultivars in Response to Different Greenhouse Irradiance Levels in Early Summer Season. Plants 2021, 10, 1179. [Google Scholar] [CrossRef]
- Ida Wenefrida, I.; Utomo, H.; Mohd Ikmal, A.; Gireesh, C.; Mallikarjuna Swamy, B.P. Reducing Cadmium Content in Zinc Biofortified Rice through Genetic Manipulation. In Genetic Engineering and Genome Editing for Zinc Biofortification of Rice; Elsevier Inc.: Amsterdam, The Netherlands, 2023; pp. 115–130. [Google Scholar]
- Kumari, V.V.; Banerjee, P.; Verma, V.C.; Sukumaran, S.; Chandran, M.A.S.; Gopinath, K.A.; Awasthi, N.K. Plant Nutrition: An Effective Way to Alleviate Abiotic Stress in Agricultural Crops. Int. J. Mol. Sci. 2022, 23, 8519. [Google Scholar] [CrossRef] [PubMed]
- Shalaby, T.A.; Abd-Alkarim, E.; El-Aidy, F.; Hamed, E.S.; Sharaf-Eldin, M.; Taha, N.; Dos Reis, A.R. Nano-Selenium, Silicon and H2O2 Boost Growth and Productivity of Cucumber Under Combined Salinity and Heat Stress. Ecotoxicol. Environ. Saf. 2021, 212, 111962. [Google Scholar] [CrossRef]
- Bhat, J.A.; Shivaraj, S.M.; Singh, P.; Navadagi, D.B.; Tripathi, D.K.; Dash, P.K.; Deshmukh, R. Role of Silicon in Mitigation of Heavy Metal Stresses in Crop Plants. Plants 2019, 8, 71. [Google Scholar] [CrossRef]
- Tavanti, T.R.; de Melo, A.A.R.; Moreira, L.D.K.; Sanchez, D.E.J.; dos Santos Silva, R.; da Silva, R.M.; Dos Reis, A.R. Micronutrient Fertilization Enhances ROS Scavenging System for Alleviation of Abiotic Stresses in Plants. Plant Physiol. Biochem. 2021, 160, 386–396. [Google Scholar] [CrossRef]
- Parkash, V.; Singh, S. Potential of Biochar Application to Mitigate Salinity Stress in Eggplant. HortScience 2020, 55, 1946–1955. [Google Scholar] [CrossRef]
- Tang, H.; Wang, S.; Liu, Y.; Mu, H.; Song, Y.; Huang, G.; Hashem, M.; Alamri, S.; Mostafa, Y.S. Biochar: A Promising Soil Amendment to Mitigate Heavy Metals Toxicity in Plants. Not. Bot. Horti Agrobot. Cluj-Napoca 2022, 50, 12778. [Google Scholar] [CrossRef]
- Wani, O.A.; Parthiban, M.; Bhat, M.A.; Mahdi, S.S.; Jan, R.; Bhat, M.A.; Kumar, S.S. Biochar: A New Emerging Tool to Mitigate Abiotic Stresses and Its Effect on Soil Properties. In Secondary Agriculture: Sustainability and Livelihood in India; Springer International Publishing: Cham, Switzerland, 2022; pp. 97–108. [Google Scholar]
- Wu, Y.; Wang, X.; Zhang, L.; Zheng, Y.; Liu, X.; Zhang, Y. The Critical Role of Biochar to Mitigate the Adverse Impacts of Drought and Salinity Stress in Plants. Front. Plant Sci. 2023, 14, 1163451. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wei, J.; Guo, L.; Fang, H.; Liu, X.; Liang, K.; Siddique, K.H. Effects of Two Biochar Types on Mitigating Drought and Salt Stress in Tomato Seedlings. Agronomy 2023, 13, 1039. [Google Scholar] [CrossRef]
- Reyes-Herrera, D.F.; Sánchez-Reinoso, A.D.; Lombardini, L.; Restrepo-Díaz, H. Physiological Responses of Coffee (Coffea arabica L.) Plants to Biochar Application under Water Deficit Conditions. Not. Bot. Horti Agrobot. Cluj-Napoca 2023, 51, 12873. [Google Scholar] [CrossRef]
- Ibrahim, E.A.; El-Sherbini, M.A.; Selim, E.M.M. Effects of Biochar on Soil Properties, Heavy Metal Availability and Uptake, and Growth of Summer Squash Grown in Metal-Contaminated Soil. Sci. Hortic. 2022, 301, 111097. [Google Scholar] [CrossRef]
- Huang, M.; Yin, X.; Chen, J.; Cao, F. Biochar Application Mitigates the Effect of Heat Stress on Rice (Oryza sativa L.) by Regulating the Root-Zone Environment. Front. Plant Sci. 2021, 12, 711725. [Google Scholar] [CrossRef] [PubMed]
- Fahad, S.; Hussain, S.; Saud, S.; Tanveer, M.; Bajwa, A.A.; Hassan, S.; Huang, J. A Biochar Application Protects Rice Pollen from High-Temperature Stress. Plant Physiol. Biochem. 2015, 96, 281–287. [Google Scholar] [CrossRef]
- Alharbi, K.; Khan, A.A.; Alhaithloul, H.A.S.; Al-Harbi, N.A.; Al-Qahtani, S.M.; Aloufi, S.S.; Soliman, M.H. Synergistic Effect Of Β-Sitosterol and Biochar Application for Improving Plant Growth of Thymus vulgaris Under Heat Stress. Chemosphere 2023, 340, 139832. [Google Scholar] [CrossRef]
- Kanwal, S.; Ilyas, N.; Shabir, S.; Saeed, M.; Gul, R.; Zahoor, M.; Mazhar, R. Application of biochar in mitigation of negative effects of salinity stress in wheat (Triticum aestivum L.). J. Plant Nutr. 2018, 41, 526–538. [Google Scholar] [CrossRef]
- Kang, S.M.; Adhikari, A.; Khan, M.A.; Kwon, E.H.; Park, Y.S.; Lee, I.J. Influence of the rhizobacterium Rhodobacter sphaeroides KE149 and biochar on waterlogging stress tolerance in Glycine max L. Environments 2021, 8, 94. [Google Scholar] [CrossRef]
- Li, X.; Cao, Y.; Xiao, J.; Salam, M.M.A.; Chen, G. Bamboo Biochar Greater Enhanced Cd/Zn Accumulation in Salix psammophila under Non-Flooded Soil Compared with Flooded. Biochar 2022, 4, 7. [Google Scholar] [CrossRef]
- Yuan, J.; Meng, J.; Liang, X.E.Y.; Yang, X.; Chen, W. Organic Molecules from Biochar Leacheates Have a Positive Effect on Rice Seedling Cold Tolerance. Front. Plant Sci. 2017, 8, 1624. [Google Scholar] [CrossRef] [PubMed]
- Terán, F.; Vives-Peris, V.; López-Climent, M.F.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Palliative Effects of Kaolin on Citrus Plants under Controlled Stress Conditions of High Temperature and High Light Intensity. J. Plant Growth Regul. 2023, 1–14. [Google Scholar] [CrossRef]
- Hamdy, A.E.; Abdel-Aziz, H.F.; El-khamissi, H.; AlJwaizea, N.I.; El-Yazied, A.A.; Selim, S.; Elkelish, A. Kaolin Improves Photosynthetic Pigments, and Antioxidant Content, and Decreases Sunburn of Mangoes: Field study. Agronomy 2022, 12, 1535. [Google Scholar] [CrossRef]
- Mahmoudian, M.; Rahemi, M.; Karimi, S.; Yazdani, N.; Tajdini, Z.; Sarikhani, S.; Vahdati, K. Role of Kaolin on Drought Tolerance and Nut Quality of Persian Walnut. J. Saudi Soc. Agric. Sci. 2021, 20, 409–416. [Google Scholar] [CrossRef]
- Brito, C.; Dinis, L.T.; Moutinho-Pereira, J.; Correia, C. Kaolin, an emerging tool to alleviate the effects of abiotic stresses on crop performance. Sci. Hortic. 2019, 250, 310–316. [Google Scholar] [CrossRef]
- Oladosu, Y.; Rafii, M.Y.; Arolu, F.; Chukwu, S.C.; Salisu, M.A.; Fagbohun, I.K.; Haliru, B.S. Superabsorbent Polymer Hydrogels for Sustainable Agriculture: A Review. Horticulturae 2022, 8, 605. [Google Scholar] [CrossRef]
- Du, F.; Liu, L.; Pan, Y.; Wu, C.; Wang, R.; Zhao, Z.; Wang, J.A. Novel Biochar-Based Composite Hydrogel for Removing Heavy Metals in Water and Alleviating Cadmium Stress in Tobacco Seedlings. Sci. Rep. 2023, 13, 15656. [Google Scholar] [CrossRef]
- Mnyika, A.W. Effect of Irrigation Regime, Super-Absorbent Polymer and Rabbit Manure on Growth and Yield of Eggplant (Solanum melongena L.) in Kilifi County. Master’s Thesis, Pwani University, Kilifi, Kenya, 2020; pp. 1–88, Unpublished. [Google Scholar]
- Jamal, A.; Hussain, S.; Hussain, S.; Matloob, A.; Awan, T.H.; Irshad, F.; Waraich, E. Super Absorbent Polymer Application under Suboptimal Environments: Implications and Challenges for Marginal Lands and Abiotic Stresses. Turk. J. Agric. For. 2022, 46, 662–676. [Google Scholar] [CrossRef]
- Ostrand, M.S.; DeSutter, T.M.; Daigh, A.L.; Limb, R.F.; Steele, D.D. Superabsorbent Polymer Characteristics, Properties, and Applications. Agrosyst. Geosci. Environ. 2020, 3, e20074. [Google Scholar] [CrossRef]
- Deolu-Ajayi, A.O.; van der Meer, I.M.; Van der Werf, A.; Karlova, R. The Power of Seaweeds as Plant Biostimulants to Boost Crop Production under Abiotic Stress. Plant Cell Environ. 2022, 45, 2537–2553. [Google Scholar] [CrossRef]
- Soualiou, S.; Duan, F.; Li, X.; Zhou, W. Crop Production under Cold Stress: An Understanding of Plant Responses, Acclimation Processes, and Management Strategies. Plant Physiol. Biochem. 2022, 190, 47–61. [Google Scholar] [CrossRef]
- Jacomassi, L.M.; Viveiros, J.D.O.; Oliveira, M.P.; Momesso, L.; de Siqueira, G.F.; Crusciol, C.A.C. A Seaweed Extract-Based Biostimulant Mitigates Drought Stress in Sugarcane. Front. Plant Sci. 2022, 13, 865291. [Google Scholar] [CrossRef]
- Hernández-Herrera, R.M.; Sánchez-Hernández, C.V.; Palmeros-Suárez, P.A.; Ocampo-Alvarez, H.; Santacruz-Ruvalcaba, F.; Meza-Canales, I.D.; Becerril-Espinosa, A. Seaweed Extract Improves Growth and Productivity of Tomato Plants under Salinity Stress. Agronomy 2022, 12, 2495. [Google Scholar] [CrossRef]
- Goyal, V.; Kumari, A.; Avtar, R.; Baliyan, V.; Mehrotra, S. Orthosilicic Acid and Seaweed Extract Alleviate the Deteriorative Effects of High Temperature Stress in Brassica juncea (L.) Czern & Coss. Silicon 2023, 15, 4909–4919. [Google Scholar]
- Pal, S.C.; Hossain, M.B.; Mallick, D.; Bushra, F.; Abdullah, S.R.; Dash, P.K.; Das, D. Combined Use of Seaweed Extract and Arbuscular Mycorrhizal Fungi for Alleviating Salt Stress in Bell Pepper (Capsicum annuum L.). Sci. Hortic. 2024, 325, 112597. [Google Scholar] [CrossRef]
- Lotfi, A.; Kottb, M.; Elsayed, A.; Shafik, H. Antifungal activity of some Mediterranean seaweed against Macrophomina phaseolina and Fusarium oxysporum in Vitro. Alfarama J. Basic Appl. Sci. 2021, 2, 81–96. [Google Scholar] [CrossRef]
- Pourakbar, L.; Moghaddam, S.S.; Enshasy, H.A.E.; Sayyed, R.Z. Antifungal activity of the extract of a macroalgae, Gracilariopsis persica, against four plant pathogenic fungi. Plants 2021, 10, 1781. [Google Scholar] [CrossRef]
- Hussein, M.H.; Eltanahy, E.; Al Bakry, A.F.; Elsafty, N.; Elshamy, M.M. Seaweed Extracts as Prospective Plant Growth Bio-Stimulant and Salinity Stress Alleviator for Vigna sinensis and Zea mays. J. Appl. Phycol. 2021, 33, 1273–1291. [Google Scholar] [CrossRef]
- Di Stasio, E.; Cirillo, V.; Raimondi, G.; Giordano, M.; Esposito, M.; Maggio, A. Osmo-Priming with Seaweed Extracts Enhances Yield of Salt-Stressed Tomato Plants. Agronomy 2020, 10, 1559. [Google Scholar] [CrossRef]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The Role of Biostimulants and Bioeffectors as Alleviators of Abiotic Stress in Crop Plants. Chem. Biol. Technol. Agric. 2017, 4, 5. [Google Scholar] [CrossRef]
- Ali, O.; Ramsubhag, A.; Jayaraman, J. Biostimulant Properties of Seaweed Extracts in Plants: Implications towards Sustainable Crop Production. Plants 2021, 10, 531. [Google Scholar] [CrossRef]
- Borella, M.; Baghdadi, A.; Bertoldo, G.; Della Lucia, M.C.; Chiodi, C.; Celletti, S.; Deb, S.; Baglieria, A.; Nardi, S. Transcriptomic and Physiological Approaches to Decipher Cold Stress Mitigation Exerted by Brown-Seaweed Extract (BSE) Application in Tomato. Front. Plant Sci. 2023, 14, 1232421. [Google Scholar] [CrossRef]
- El Boukhari, M.E.M.; Barakate, M.; Drissi, B.; Bouhia, Y.; Lyamlouli, K. Seaweed Extract Biostimulants Differentially act in Mitigating Drought Stress on Faba Bean (Vicia faba L.). J. Plant Growth Regul. 2023, 42, 5642–5652. [Google Scholar] [CrossRef]
- Shukla, P.S.; Shotton, K.; Norman, E.; Neily, W.; Critchley, A.T.; Prithiviraj, B. Seaweed Extract Improve Drought Tolerance of Soybean by Regulating Stress-Response Genes. AoB Plants 2018, 10, plx051. [Google Scholar] [CrossRef] [PubMed]
- Rasul, F.; Gupta, S.; Olas, J.J.; Gechev, T.; Sujeeth, N.; Mueller-Roeber, B. Priming with a Seaweed Extract Strongly Improves Drought Tolerance in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 1469. [Google Scholar] [CrossRef] [PubMed]
- Sabatino, L.; Consentino, B.B.; Rouphael, Y.; Baldassano, S.; De Pasquale, C.; Ntatsi, G. Ecklonia Maxima-Derivate Seaweed Extract Supply as Mitigation Strategy to Alleviate Drought Stress in Chicory Plants. Sci. Hortic. 2023, 312, 111856. [Google Scholar] [CrossRef]
- Anjos Neto, A.P.D.; Oliveira, G.R.F.; Mello, S.D.C.; Silva, M.S.D.; Gomes-Junior, F.G.; Novembre, A.D.D.L.C.; Azevedo, R.A. Seed Priming with Seaweed Extract Mitigate Heat Stress in Spinach: Effect on Germination, Seedling Growth and Antioxidant Capacity. Bragantia 2020, 79, 502–511. [Google Scholar] [CrossRef]
- Zhang, X.; Ervin, E.H. Impact of Seaweed Extract-Based Cytokinins and Zeatin Riboside on Creeping Bentgrass Heat Tolerance. Crop Sci. 2008, 48, 364–370. [Google Scholar] [CrossRef]
- Lakhdar, A.; Trigui, M.; Montemurro, F. An Overview of Biostimulants’ Effects in Saline Soils. Agronomy 2023, 13, 2092. [Google Scholar] [CrossRef]
- Abir, M.U.; Nupur, J.A.; Hunter, C.T.; Al, A.M.S.; Sagar, A.; Hossain, M.S.; Dawood, M.F.A.; Lateef, A.A.H.A.; Brestyic, M.; Tahjib-UI-Arif, M. Crop Improvement and Abiotic Stress Tolerance Promoted by Moringa Leaf Extract. Phyton 2022, 91, 1557–1583. [Google Scholar] [CrossRef]
- Ahmad, A.; Blasco, B.; Martos, V. Combating Salinity Through Natural Plant Extracts Based Biostimulants: A Review. Front. Plant Sci. 2022, 13, 862034. [Google Scholar] [CrossRef]
- Bibi, A.; Ullah, F.; Mehmood, S.; Bibi, K.; Khan, S.U.; Khattak, A.; Ullah Khan, R. Moringa oleifera Lam. Leaf Extract as Bioregulator for Improving Growth of Maize Under Mercuric Chloride Stress. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2016, 66, 469–475. [Google Scholar]
- ElSayed, A.I.; Rafudeen, M.S.; Ganie, S.A.; Hossain, M.S.; Gomaa, A.M. Seed Priming with Cypress Leaf Extract Enhances Photosynthesis and Antioxidative Defense in Zucchini Seedlings Under Salt Stress. Sci. Hortic. 2022, 293, 110707. [Google Scholar] [CrossRef]
- Air, T.K. The Effect of Moringa oleifera Lam. Leaf Aqueous Extract on Seed Yield and Fibre Quality of Linseed under Water Deficit Stress. Sains Malays. 2022, 51, 1027–1044. [Google Scholar]
- Besrukow, P.; Will, F.; Dussling, S.; Berkelmann-Löhnertz, B.; Schweiggert, R. Additive and Synergistic Antifungal Effects of Copper and Phenolic Extracts from Grape Cane and Apples. Pest Manag. Sci. 2023, 79, 3334–3341. [Google Scholar] [CrossRef]
- Huang, P.; He, L.; Abbas, A.; Hussain, S.; Hussain, S.; Du, D.; Hafeez, M.B.; Balooch, S.; Zahra, N.; Ren, X.; et al. Seed Priming with Sorghum Water Extract Improves the Performance of Camelina (Camelina sativa (L.) Crantz.) Under Salt Stress. Plants 2021, 10, 749. [Google Scholar] [CrossRef]
- El-Yazal, M.A.S.; Salama, M.M.; Rady, M.M. Impact of Presoaking and Foliar Spray Application by Maize Grain Extract in Alleviates Salinity Stress in Common Bean (Phaseolus vulgaris L.) Plants Grown under Salt Stress. J. Agric. Res. Pestic. Biofertil. 2021, 2, 1–10. [Google Scholar]
- Desoky, E.S.M.; El-maghraby, L.M.; Awad, A.E.; Abdo, A.I.; Rady, M.M.; Semida, W.M. Fennel and Ammi Seed Extracts Modulate Antioxidant Defence System and Alleviate Salinity Stress in Cowpea (Vigna unguiculata). Sci. Hortic. 2020, 272, 109576. [Google Scholar] [CrossRef]
- Suryaman, M.; Sunarya, Y.; Istarimila, I.; Fudholi, A. Effect of Salinity Stress on the Growth and Yield of Mungbean (Vigna radiata (L.) R. Wilczek) Treated with Mangosteen Pericarp Extract. Biocatal. Agric. Biotechnol. 2021, 36, 102132. [Google Scholar] [CrossRef]
- Kasim, W.A.E.A.; Nessem, A.A.; Gaber, A. Alleviation of Drought Stress in Vicia faba by Seed Priming with Ascorbic Acid or Extracts of Garlic and Carrot. Egypt. J. Bot. 2017, 57, 45–59. [Google Scholar] [CrossRef]
- Toscano, S.; Gómez-Bellot, M.J.; Romano, D.; Sánchez-Blanco, M.J. Physiological and Biochemical Changes in Response to Moringa oleifera Biostimulant in Petunia Plants under Water Deficit. Sci. Hortic. 2023, 319, 112187. [Google Scholar] [CrossRef]
- Buthelezi, D.; Gololo, D.S.S.; Mugivhisa, D.L.L. An Assessment of Moringa (Moringa oleifera) Seed Extract on Water Use Efficiency and Physico-Biochemical Properties of Cancer Bush (Sutherlandia frutescens) under Deficit Irrigation. Horticulturae 2022, 8, 938. [Google Scholar] [CrossRef]
- Elsayed, S.I.; Mazhar, A.A.; El-Sayed, S.M.; Said Mohamed, A. Improvement the Drought Tolerance of Eucalyptus Citriodora Seedling by Spraying Basil Leaves Extract and Its Influence on Growth, Volatile Oil Components and Some Enzymatic Activity. Egypt. J. Chem. 2022, 65, 619–635. [Google Scholar]
- Afzal, I.; Akram, M.W.; Rehman, H.U.; Rashid, S.; Basra, S.M.A. Moringa Leaf and Sorghum Water Extracts and Salicylic Acid to Alleviate Impacts of Heat Stress in Wheat. S. Afr. J. Bot. 2020, 129, 169–174. [Google Scholar] [CrossRef]
- Rashid, N.; Basra, S.M.; Shahbaz, M.; Iqbal, S.; Hafeez, M.B. Foliar Applied Moringa Leaf Extract Induces Terminal Heat Tolerance in Quinoa. Int. J. Agric. Biol. 2018, 20, 157–164. [Google Scholar]
- Batool, A.; Wahid, A.; Abbas, G.; Shah, S.H.; Akhtar, M.N.; Perveen, N.; Hassnain, Z. Application of Moringa oleifera plant extracts for enhancing the concentration of photosynthetic pigments leading to stable photosynthesis under heat stress in maize (Zea mays L.). Pak. J. Bot. 2019, 51, 3021–3026. [Google Scholar] [CrossRef]
- Ezeonu, C.S.; Tatah, S.V.; Imo, C.; Yakubu, O.E.; Garba, Q.H.; Arowora, K.; Umaru, I.J.; Abah, M.A.; Abu, M.S.; Yohanna, E.R.; et al. Antioxidant Potential of Ginger Extract on Metals (Lead, Cadmium, and Boron) Induced Oxidative Stress in Maize Plant. Asian J. Trop. Biotechnol. 2022, 19, 45–51. [Google Scholar]
- Nessem, A.A.; Mohsen, A.A.; Atlam, A.; Ragab, G.A. Effect of Priming with Natural Plant Extract on the Growth, Hormonal Status, and Yield Value of Triticum asetivum (L.) Grown under Lead and Nickel Stress. Egypt. J. Bot. 2023, 63, 159–174. [Google Scholar] [CrossRef]
- Baroniya, M.B.; Vyas, H.; Titov, A.; Vyas, A.; Baroniya, S.S. Protection of Soybean from Ambient UV Radiation THROUGH Foliar Spray with Lawsonia inermis Leaves Extract. Plant Arch. 2020, 20, 4293–4300. [Google Scholar]
- Batool, S.; Khan, S.; Basra, S.M.A. Foliar Application of Moringa Leaf Extract Improves the Growth of Moringa Seedlings in Winter. S. Afr. J. Bot. 2020, 129, 347–353. [Google Scholar] [CrossRef]
- Abdelaal, K.; Attia, K.A.; Niedbała, G.; Wojciechowski, T.; Hafez, Y.; Alamery, S.; Alateeq, T.K.; Arafa, S.A. Mitigation of Drought Damages by Exogenous Chitosan and Yeast Extract with Modulating the Photosynthetic Pigments, Antioxidant Defense System and Improving the Productivity of Garlic Plants. Horticulturae 2021, 7, 510. [Google Scholar] [CrossRef]
- Sadak, M.S.; Dawood, M.G. Biofertilizer Role in Alleviating the Deleterious Effects of Salinity on Wheat Growth and Productivity. Gesunde Pflanz. 2023, 75, 1207–1219. [Google Scholar] [CrossRef]
- Liu, W.; Wang, B.; Wang, Q.; Hou, J.; Wu, L.; Wood, J.L.; Luo, Y.; Franks, A.E. Characteristics of Metal-Tolerant Plant Growth-Promoting Yeast (Cryptococcus sp. NSE1) and its Influence on Cd Hyperaccumulator Sedum Plumbizincicola. Environ. Sci. Pollut. Res. 2016, 23, 18621–18629. [Google Scholar] [CrossRef] [PubMed]
- Taha, S.R.; Seleiman, M.F.; Alhammad, B.A.; Alkahtani, J.; Alwahibi, M.S.; Mahdi, A.H. Activated Yeast extract enhances growth, anatomical structure, and productivity of Lupinus termis L. plants under actual salinity conditions. Agronomy 2020, 11, 74. [Google Scholar] [CrossRef]
- Babaousmail, M.; Nili, M.S.; Brik, R.; Saadouni, M.; Yousif, S.K.; Omer, R.M.; Osman, N.A.; Alsahli, A.A.; Ashour, H.; El-Taher, A.M. Improving the Tolerance to Salinity Stress in Lettuce Plants (Lactuca sativa L.) using exogenous application of salicylic acid, yeast, and zeolite. Life 2022, 12, 1538. [Google Scholar] [CrossRef] [PubMed]
- Alzandi, A.A.; Naguib, D.M. Effect of Yeast Application on Soil Health and Root Metabolic Status of Corn Seedlings under Drought Stress. Arch. Microbiol. 2022, 204, 233. [Google Scholar] [CrossRef]
- Yousef, E.A.A.; Ali, M.A.M. Alleviation of Cold Stress on Tomato during Winter Season by Application of Yeast Extract and Glycinebetaine. Egypt. J. Hortic. 2019, 46, 117–131. [Google Scholar] [CrossRef]
- Sadak, M.S. Physiological Role of Yeast Extract and Nicotinamide on Pisum sativum L. Plants under Heat Stress. Int. J. PharmTech Res. 2016, 9, 170–178. [Google Scholar]
- Aazami, M.A.; Maleki, M.; Rasouli, F.; Gohari, G. Protective Effects Of Chitosan Based Salicylic Acid Nanocomposite (CS-SA NCs) in Grape (Vitis vinifera cv.‘Sultana’) Under Salinity Stress. Sci. Rep. 2023, 13, 883. [Google Scholar] [CrossRef]
- Ali, M.; Ayyub, C.M.; Silverman, E.; Rehman, M.A.; Iqbal, S.; Hussain, Z.; Nawaz, M.A.; Bazmi, M.S.A. Evaluation of Physiological Traits and Flowering in Cucumis sativus L. by Foliar Application of Chitosan at Three Sowing Dates Grown under Hot Environment. JPAA 2021, 6, 62–75. [Google Scholar]
- Tan, C.; Li, N.; Wang, Y.; Yu, X.; Yang, L.; Cao, R.; Ye, X. Integrated Physiological and Transcriptomic Analyses Revealed Improved Cold Tolerance in Cucumber (Cucumis sativus L.) by exogenous chitosan oligosaccharide. Int. J. Mol. Sci. 2023, 24, 6202. [Google Scholar] [CrossRef]
- Hafez, Y.; Attia, K.; Alamery, S.; Ghazy, A.; Al-Doss, A.; Ibrahim, E.; Rashwan, E.; El-Maghraby, L.; Awad, A.; Abdelaal, K. Beneficial Effects of Biochar and Chitosan on antioxidative Capacity, Osmolytes Accumulation, and Anatomical Characters of Water-Stressed Barley Plants. Agronomy 2020, 10, 630. [Google Scholar] [CrossRef]
- Rasheed, R.; Ashraf, M.A.; Arshad, A.; Iqbal, M.; Hussain, I. Interactive Effects of Chitosan and Cadmium on Growth, Secondary Metabolism, Oxidative Defense, And Element Uptake in Pea (Pisum sativum L.). Arab. J. Geosci. 2020, 13, 847. [Google Scholar] [CrossRef]
- Kocięcka, J.; Liberacki, D. The Potential of Using Chitosan on Cereal Crops in the Face of Climate Change. Plants 2021, 10, 1160. [Google Scholar] [CrossRef]
- Wang, A.; Li, J.; Al-Huqail, A.A.; Al-Harbi, M.S.; Ali, E.F.; Wang, J.; Ding, Z.; Rekaby, S.A.; Ghoneim, A.M.; Eissa, M.A. Mechanisms of Chitosan Nanoparticles in the Regulation of Cold Stress Resistance in Banana Plants. Nanomaterials 2021, 11, 2670. [Google Scholar] [CrossRef] [PubMed]
- Hidangmayum, A.; Dwivedi, P.; Katiyar, D.; Hemantaranjan, A. Application of Chitosan on Plant Responses with Special Reference to Abiotic Stress. Physiol. Mol. Biol. Plants 2019, 25, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Manzoor, N.; Ali, L.; Ahmed, T.; Noman, M.; Adrees, M.; Shahid, M.S.; Ogunyemi, S.O.; Radwan, K.S.A.; Wang, G.; Zaki, H.E. Recent Advancements and Development in Nano-Enabled Agriculture for Improving Abiotic Stress Tolerance in Plants. Front. Plant Sci. 2022, 13, 951752. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, S.; Waheed, Z.; Naseem, A. Nanotechnology and Abiotic Stresses. In Nanoagronomy; Springer: Cham, Switzerland, 2020; pp. 37–52. [Google Scholar]
- Al-Khayri, J.M.; Rashmi, R.; Surya Ulhas, R.; Sudheer, W.N.; Banadka, A.; Nagella, P.; Aldaej, M.I.; Rezk, A.A.-S.; Shehata, W.F.; Almaghasla, M.I. The Role of Nanoparticles in Response of Plants to Abiotic Stress at Physiological, Biochemical, and Molecular Levels. Plants 2023, 12, 292. [Google Scholar] [CrossRef] [PubMed]





| Transgenic Crop | Abiotic Stresses | Gene and Its Source | Traits Improved | References |
|---|---|---|---|---|
| Tobacco | Salinity | bet A from Escherichia coli | Enhanced biomass | [26] |
| Arabidopsis and tobacco | Salinity, drought, and cold stress | GmbZIP1 from soybean | Enhanced tolerance to multiple stresses | [22] |
| Rice | Heavy metal | MTH1745 from thermophilic archaea | Improved photosynthesis and conferred mercury tolerance | [93] |
| Potato | Salinity | pyrroline-5-carboxylate synthetase (P5CS) gene from Arabidopsis | Increased proline content and tuber production | [94] |
| Tomato | Heat stress | StGLP from yeast | Improved tolerance to gradual heat stress | [95] |
| Tall fescue | Salinity | AtNHX1 from Arabidopsis | Increased shoot and root dry weight | [19] |
| Arabidopsis | Salinity and drought | TaMYB2A from wheat | Enhanced tolerance to drought and salinity | [22] |
| Rice | Waterlogging, salinity, and drought | OsARD1 from rice | Increased ethylene synthesis and conferred tolerance to submergence, drought, salt, and osmotic stresses | [96] |
| Potato | Cold stress | Δ12-desaturase gene from cyanobacteria | Improved tolerance by reducing malondialdehyde content | [94] |
| Tobacco | Cold stress | TaMYC2 from caucasian clover | Increased production of antioxidant enzymes | [89] |
| Arabidopsis | Heat stress | TaHsfA6f from wheat | Enhanced heat tolerance and abscisic acid (ABA) accumulation | [97] |
| Medicago truncatula | Cold stress | MtDREB1C/MtCBF3 from Medicago truncatula | Inhibited growth of the shoot and enhanced cold tolerance | [98] |
| Alfalfa | Heavy metal | ATP sulfurylase gene from Arabidopsis | Improved tolerance to heavy metal | [99] |
| Tomato | Cold stress | SiFBA5 from Snow Lotus | Enhanced photosynthetic efficiency and tolerance to cold stress | [100] |
| Potato | Drought | BADH transgene from spinach | Enhanced fresh weight | [94] |
| Medicago truncatula | Drought | MtP5CS from Medicago truncatula | Increased proline production and conferred tolerance | [98] |
| Abiotic Stress | Crop | Priming Technique | Impacts | References |
|---|---|---|---|---|
| Heavy metal | Pigeon pea | Phytohormones priming | Increased germination speed index, germination percentage, and tolerance to cadmium | [116] |
| Garden cress | Phytohormone priming | Improved seed germination, seedling emergence and tolerance index and reduced phytotoxicity. | [123] | |
| Mustard | Phytohormone priming | Improved shoot emergence and decreased accumulation of heavy metal | [124] | |
| Maize | Nanoparticle priming | Reduced uptake of toxins and improved photosynthesis pigments and plant ultra-cellular structures | [125] | |
| Salinity | Maize and wheat | Biopriming | Improved physiological and morphological traits | [120] |
| Rice | Phytohormone priming | Reduced sodium ions accumulation and increased potassium concentration | [126] | |
| Capsicum | Nanoparticle priming | Increased enlargement of roots and promoted salt tolerance | [122] | |
| Rapeseed | Hydropriming | Increased germination percentage | [118] | |
| Sweet pepper | Osmopriming | Conferred tolerance due to enhanced biosynthesis of plastidial pigments | [118] | |
| Drought | Plants | Molecular priming | Induced tolerance against oxidative stress | [117] |
| Rye seed | Hydropriming | Stimulated antioxidant enzymes production | [127] | |
| Soybean | Hydropriming | Enhanced germination and seed yield | [128] | |
| Caraway | Osmopriming | Increased germination and fresh and dry weights of the plumule in the seeds | [127] | |
| Waterlogging | Rice | Phytohormone priming | Increased α-Amylase activity, sucrose, glucose, and fructose | [116] |
| Rice | Chemical and hormonal priming | Improved tolerance to waterlogging | [119] | |
| Okra | Chemical priming | Induced aerenchyma development resulting in okra survival | [129] | |
| Heat stress | Tall fescue | Phytohormone priming | Induced heat-responsive genes | [121] |
| Maize | Phytohormone priming | Increased photosynthetic pigment, membrane stability index, relative water content, growth rate, grain yield, harvest index, and grain protein | [130] | |
| Yarrow | Heat priming | Improved photosynthesis and stomatal conductance | [131] | |
| Arabidopsis | Thermopriming | Modified energy pathway and enhanced branched-chain amino acids production | [132] | |
| Cold stress | Canola | Thermopriming | Improved germination | [26] |
| Rapeseed | Phytohormones and hydropriming | Improved photosynthesis, stomatal conductance, transpiration rate, and antioxidant enzymes | [133] | |
| Chilling stress | Maize | Phytohormone priming | Enhanced germination, antioxidant enzyme, starch metabolism, and malondialdehyde content | [134] |
| Salinity and drought | Chickpea | Osmopriming | Suppressed stress effect | [116] |
| Abiotic Stresses | Plant Growth Regulators and Osmoprotectants | Crop | Mechanism | References |
|---|---|---|---|---|
| Salinity | 5-Aminolevulinic acid | Cotton | Enhanced seedling germination | [26] |
| Proline | Wheat | Enhanced hormones accumulation and reduced malondialdehyde | [23] | |
| Thiamin | Sunflower | Enhanced chlorophyll content, biomass, and leaf relative water content | [26] | |
| Indole acetic acid | Wheat | Reduced oxidative stress | [88] | |
| Brassinosteroid | Wheat | Increased yield | [23] | |
| Drought | Melatonin | Wheat | Enhanced seedling vigour and tolerance | [138] |
| Abscisic Acid | Rice | Increased antioxidant enzymes, proline, and soluble sugar content | [117] | |
| Gibberellic acid | Faba bean | Improved relative water content, cell membrane stability, and nutrient status | [143] | |
| Salicylic acid | Sunflower | Enhanced yield traits and quality | [144] | |
| Waterlogging | Gibberellic acid | Soybean | Improved flooding tolerance | [21] |
| Cytokinin and gibberellic acid | Mungbean | Enhanced growth and biomass, photosynthesis pigments and proline, and total soluble sugars | [145] | |
| 6-Benzyladenine | Maize | Suppressed the effect of stress on grain yield | [146] | |
| Salicylic acid and kinetin | Soybean | Increased antioxidant enzymes activities and the glyoxalase system | [147] | |
| Heat stress | 24- epibrassinolide | Mustard | Induced heat tolerance | [16] |
| Trehalose | Maize | Reduced the accumulation of malondialdehyde and enhanced cell integrity | [144] | |
| Indole-3-acetic acid | Rice | Improved yield | [148] | |
| Proline | Okra | Enhanced antioxidant enzymes activities and growth traits | [149] | |
| Heavy metal | Gibberellic acid | Lupin plants | Improved amylase and catalase activities | [150] |
| Salicylic acid gibberellic acid and triacontanol | Menthol mint | Reduced cadmium accumulation in the leaves and upregulated proline and antioxidant enzymes | [151] | |
| 24-epibrassinolide | Mustard | Reduced heavy metal accumulation and enhanced growth rate | [150] | |
| Cold stress | Brassinosteroids | Tomato | Improved growth by reducing oxidative stress | [152] |
| Auxin | Pea | Reduced conjugated polyamines and increased proline content | [153] | |
| Abscisic acid | Banana | Maintained cell membrane stability by reducing ion leakage and lipid peroxidation | [154] | |
| Strigolactones | Mung bean | Reduced phenolic contents and enhanced water status and proline content | [155] |
| Abiotic Stresses | Crop | Biochar Dosage | Impact | References |
|---|---|---|---|---|
| Drought | Barley | 1% | Enhanced growth of root and shoot | [181] |
| Cabbage | 10% | Improved rate of photosynthesis, nutrient uptake, and growth traits | [182] | |
| Tomato | 5% | Promoted seedling growth | [183] | |
| Coffee | 8 t/ha | Improved photosynthesis | [184] | |
| Heavy metal | Wheat | 5% | Increased growth and plant dry weight | [180] |
| Squash | 4% | Reduced heavy uptake by roots and shoot | [185] | |
| Sunflower | 50% | Enhanced biomass and decreased heavy metal accumulation | [180] | |
| Heat stress | Rice | 40 g/kg | Improved uptake and assimilation of nitrogen and transport of proteins and enhanced root traits | [186] |
| Rice | 300 g | Enhanced pollen germination rate, anther dehiscence, and pollen fertility | [187] | |
| Thymus | 5% | Improved growth traits, leaf chlorophyll, and photosynthetic rates | [188] | |
| Salinity | Beans | 10% and 20% | Increased growth of root and shoot | [181] |
| Eggplant | 5% | Improved photosynthesis activities and reduced leaf temperature and electrolyte leakage. | [179] | |
| Maize | 5% | Increased proline content and decreased sodium content | [182] | |
| Wheat | 2% | Improved root and shoot length, leaf water potential, and osmotic potential | [189] | |
| Waterlogging | Soybean | 0.1% | Enhanced growth traits | [190] |
| Salix psammophila | 3% | Promoted cadmium and zinc accumulation in the plant | [191] | |
| Cold stress | Rice | 10% | Enhanced tolerance | [192] |
| Abiotic Stresses | Plant Extract | Extract Type | Method of Application | Plant under Study | Impacts | References |
|---|---|---|---|---|---|---|
| Salinity | Sorghum | Whole plant | Seed priming | Camelina | Increased growth traits (emergence percentage, root length, shoot length, biomass, α-amylase activity, chlorophyll content, antioxidant enzymes activity) | [228] |
| Maize | Grain | Soaking and foliar | Bean | Improved relative water content, proline, photosynthesis, ascorbic acid, and mineral nutrients | [229] | |
| Rosmarinus officinalis and Artemisia herba-alba | Leaf | Seed priming | Maize | Enhanced germination percentage, germination indexes, and photosynthesis pigments | [223] | |
| Foeniculum vulgare and Ammi visnaga | Seed | Foliar spray | Cowpea | Improved osmoprotectants content and antioxidant enzymes | [230] | |
| Mangosteen | Pericarp | Soaking | Mungbean | Enhanced growth and yield traits and conferred tolerance | [231] | |
| Drought | Carrot | Root | Seed priming | Bean | Enhanced chlorophyll content and photosynthesis | [232] |
| Moringa oleifera | Leaf | Foliar | Soybean and Maize | Increased growth traits (leaf area, plant height, and biomass production) | [222] | |
| Moringa oleifera | Leaf | Foliar | Petunia plants | Scavenged ROS and increased the accumulation of phenol compounds and total soluble sugars | [233] | |
| Moringa oleifera | Seed | Foliar | Cancer bush | Enhanced growth and yield traits, water use efficiency, leaf photosynthetic pigments, and antioxidant enzymes | [234] | |
| Basil | Leaf | Foliar | Eucalyptus citriodora | Improved the accumulation of essential oil fresh leaves | [235] | |
| Heat stress | Sorghum and Moringa oleifera | Whole plant and leaf | Foliar | Wheat | Enhanced yield traits (spike length, number of grains per spike, and 1000-grain weight) | [236] |
| Moringa oleifera | Leaf | Foliar | Quinoa | Enhanced leaf chlorophyll and antioxidants enzymes | [237] | |
| Moringa oleifera | Leaf | Foliar | Maize | Enhanced chlorophyll content and reduced chlorophyll-to-carotenoids ratio | [238] | |
| Heavy metal | Ginger | Whole plant | Mixed with soil | Maize | Increased accumulation of antioxidant enzymes | [239] |
| Moringa oleifera | Leaf | Foliar | Bean | Enhanced photosynthesis, proline, and relative water content | [222] | |
| Sonchus oleraceus | Grain | Priming | Wheat | Increased growth traits, photosynthetic efficiency, and yield | [240] | |
| UV-radiation | Lawsonia inermis L. | Leaf | Foliar | Soybean | Enhance crop performance | [241] |
| Cold stress | Moringa oleifera | Leaf | Foliar | Moringa | Improved morpho-physiological traits (number of branches, leaves, chlorophyll, and phenolic contents) | [242] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oyebamiji, Y.O.; Adigun, B.A.; Shamsudin, N.A.A.; Ikmal, A.M.; Salisu, M.A.; Malike, F.A.; Lateef, A.A. Recent Advancements in Mitigating Abiotic Stresses in Crops. Horticulturae 2024, 10, 156. https://doi.org/10.3390/horticulturae10020156
Oyebamiji YO, Adigun BA, Shamsudin NAA, Ikmal AM, Salisu MA, Malike FA, Lateef AA. Recent Advancements in Mitigating Abiotic Stresses in Crops. Horticulturae. 2024; 10(2):156. https://doi.org/10.3390/horticulturae10020156
Chicago/Turabian StyleOyebamiji, Yusuf Opeyemi, Basit Akolade Adigun, Noraziyah Abd Aziz Shamsudin, Asmuni Mohd Ikmal, Monsuru Adekunle Salisu, Fadila Ahmad Malike, and Adebola Azeez Lateef. 2024. "Recent Advancements in Mitigating Abiotic Stresses in Crops" Horticulturae 10, no. 2: 156. https://doi.org/10.3390/horticulturae10020156