

Implications of Vegetal Protein Hydrolysates for Improving Nitrogen Use Efficiency in Leafy Vegetables

Abstract
:1. Introduction
2. Protein Hydrolysates: What They Are and How They Work
3. Nitrogen Use Efficiency and Nitrate Uptake
4. Biostimulant Action of Protein Hydrolysates in Increasing NUE: A Case Study on Leafy Vegetables

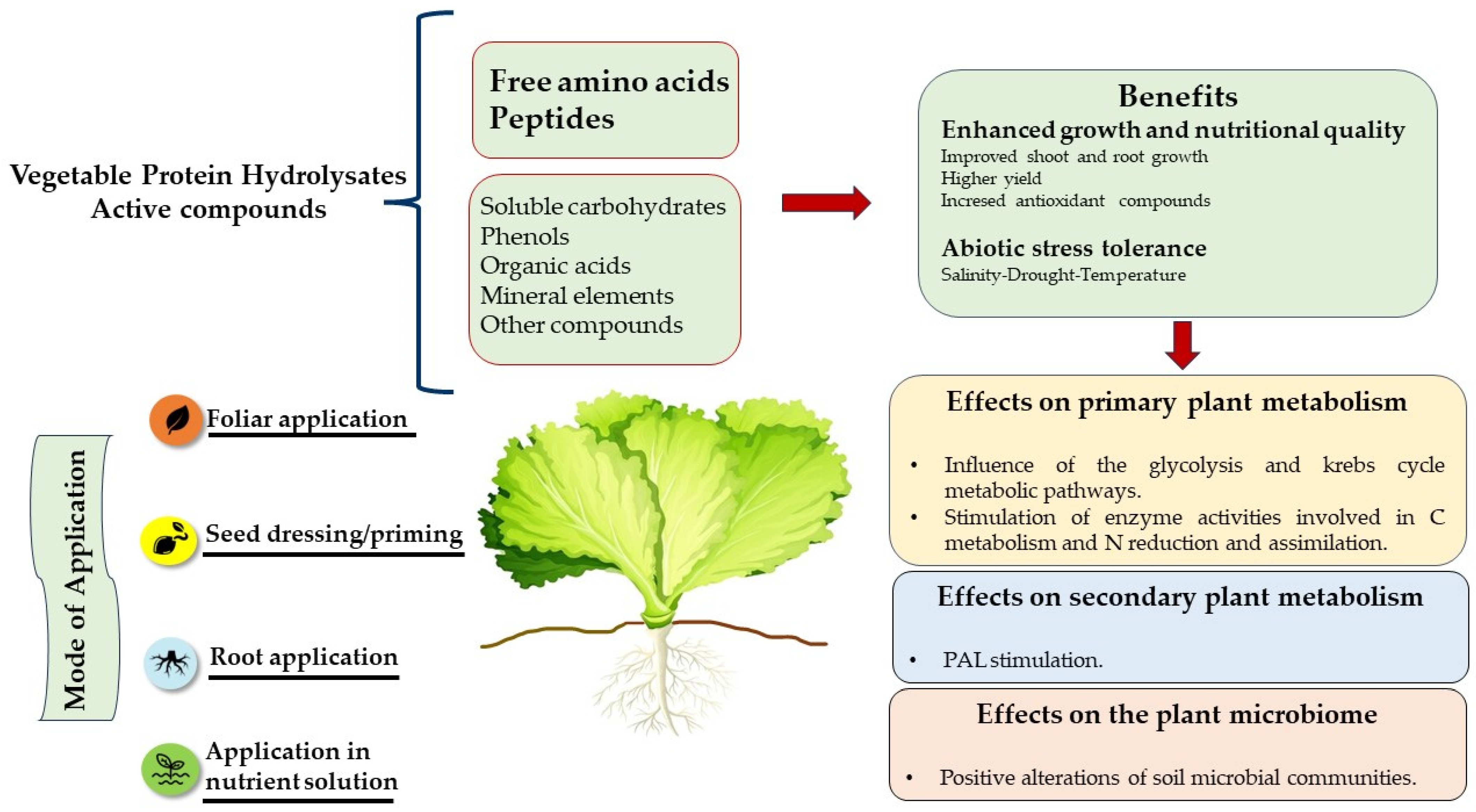{kind=link}
| Crops | V-PH Application Mode | Experimental Conditions | Nitrogen Fertilisation Management | Effects | Reference |
|---|---|---|---|---|---|
| Diplotaxis erucoides L. | Foliar | Soil culture in greenhouse | Four different nitrogen levels: 0, 60, 80, and 100 kg ha−1 | The application of V-PH increased, under N-deficient conditions (0 and 60 kg ha−1), the production of fresh rocket (+35% on average). The biostimulant treatment increased, compared to the control plants, chlorophyll and carotenoid contents, but also nitrate content. | [75] |
| Diplotaxis tenuifolia (L.) | Foliar | Soil culture in greenhouse | Optimal | V-PH improved marketable yield (+15%), colorimetric parameters, mineral composition, and antioxidant activity of the plants. | [71] |
| Lactuca sativa L. | Foliar | Soil culture in greenhouse | Optimal | A significant increase in yield (26%), nutritional and functional traits, and nitrogen indices (N use efficiency and N physiological use efficiency). | [68] |
| Lactuca sativa L. | Foliar and root | Floating raft system | Optimal | The application of V-PH directly into the nutrient solution or by foliar application increased the fresh production and mineral status of lettuce plants. | [77] |
| Lactuca sativa L. | Foliar and root | Pot culture in greenhouse | Four different levels of nitrogen in the nutrient solution: 2, 5, 10, and 15 mM | Compared to foliar application, root application of V-PH significantly increased yield, chlorophyll concentrations, and antioxidant activities, associated with increased utilization efficiency and nitrogen uptake. | [78] |
| Lactuca sativa L. | Root | Soil-less culture in growth chamber | Three different nitrogen levels: 2.4, 4.8, and 8 mmol L−1 NaNO3 | Improved N utilization and uptake efficiency as well as production and physiological performance of V-PH-treated plants. | [74] |
| Lactuca sativa L. | Foliar | Pot culture in greenhouse | Optimal | Regardless of the dose used, V-PH increased root biomass, leaf area, polyphenol content, and NUE compared to control plants. | [82] |
| Mentha × piperita | Root | Floating raft system | Optimal | Improved nutritional status and bioactive compounds (total phenols and carotenoids) with a significant reduction in nitrate content. | [80] |
| Ocimum basilicum L. | Foliar | Soil culture in greenhouse | Four different nitrogen levels: 0, 50, 100, and 150 kg ha−1 | The application of V-PH increased fresh production and NUE under N-deficient conditions without increasing the nitrate concentration. Independent of the nitrogen regime, the application of V-PH increased polyphenol content. | [73] |
| Ocimum basilicum L. | Foliar | Soil culture in greenhouse | Optimal | Compared to control conditions, the application of V-PH increased fresh production, net CO2 assimilation, and the nutritional status of basil plants (higher contents of K and N). | [72] |
| Ocimum basilicum L. | Root | Floating raft system | Two different nitrogen levels in nutrient solution: 7.5 and 14 mM | Significant increase in marketable yield due to improved photosynthetic performance of V-PH-treated plants. | [81] |
| Spinacia oleracea L. | Foliar | Soil culture in greenhouse | Four different nitrogen levels: 0, 15, 30, and 45 kg ha−1 | The application of V-PH increased the fresh and dry yield under N-deficient conditions without altering the nitrate concentration in the leaves. Independent of the nitrogen regime, V-PH increased Ca and Mg contents while reducing polyphenol content. | [69] |
| Spinacia oleracea L. | Foliar | Soil culture in greenhouse | Three different nitrogen levels: 0, 2.25, and 4.5 g m−2 | Improvement of both utilization efficiency and nitrogen uptake efficiency. For each nitrogen level, V-PH significantly increased fresh production, with the greatest effect observed under optimal nitrogen conditions. | [79] |
| Spinacia oleracea L. | Foliar | Ebb and flow soil-less cultivation system | Three different nitrogen levels in nutrient solution: 2, 8, and 14 mM | V-PH increased fresh yield under optimal nitrogen conditions (14 mM) inducing an improvement in N uptake, foliar expansion, and photosynthetic activities. | [76] |
| Valerianella locusta L. | Foliar | Soil culture in greenhouse | Three different nitrogen levels: 0, 2.25, and 4.5 g m−2 | The use of V-PH increased fresh production at sub-optimal nitrogen levels. The biostimulant increased not only the NUE but also nitrate content. | [79] |
5. Conclusions and Challenges Ahead
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hirel, B.; Le Gouis, J.; Ney, B.; Gallais, A. The challenge of improving nitrogen use efficiency in crop plants: Towards a more central role for genetic variability and quantitative genetics within integrated approaches. J. Exp. Bot. 2007, 58, 2369–2387. [Google Scholar] [CrossRef] [PubMed]
- Selvan, T.; Panmei, L.; Murasing, K.K.; Guleria, V.; Ramesh, K.R.; Bhardwaj, D.; Thakur, C.; Kumar, D.; Sharma, P.; Umedsinh, R.D. Circular economy in agriculture: Unleashing the potential of integrated organic farming for food security and sustainable development. Integr. Org. Farming Syst. Approach Effic. Food Prod. Environ. Sustain. 2023, 7, 1170380. [Google Scholar] [CrossRef]
- Petrosillo, I.; Valente, D.; Scavuzzo, C.M.; Selvan, T. Land degradation pattern and ecosystem services. Front. Environ. Sci. 2023, 11, 1137768. [Google Scholar] [CrossRef]
- Gomiero, T.; Pimentel, D.; Paoletti, M.G. Environmental impact of different agricultural management practices: Conventional vs. organic agriculture. Crit. Rev. Plant Sci. 2011, 30, 95–124. [Google Scholar] [CrossRef]
- De Corato, U. Agricultural waste recycling in horticultural intensive farming systems by on-farm composting and compost-based tea application improves soil quality and plant health: A review under the perspective of a circular economy. Sci. Total Environ. 2020, 738, 139840. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Msigwa, G.; Yang, M.; Osman, A.I.; Fawzy, S.; Rooney, D.W.; Yap, P.-S. Strategies to achieve a carbon neutral society: A review. Environ. Chem. Lett. 2022, 20, 2277–2310. [Google Scholar] [CrossRef] [PubMed]
- Landeta, C.; Marchant, F. Biostimulants: Emerging Trend and Opportunities. In Biostimulants: Exploring Sources and Applications; Springer: Berlin/Heidelberg, Germany, 2022; pp. 263–290. [Google Scholar]
- Ramawat, N.; Bhardwaj, V. Biostimulants: Exploring Sources and Applications; Springer: Berlin/Heidelberg, Germany, 2022. [Google Scholar]
- Malécange, M.; Sergheraert, R.; Teulat, B.; Mounier, E.; Lothier, J.; Sakr, S. Biostimulant Properties of Protein Hydrolysates: Recent Advances and Future Challenges. Int. J. Mol. Sci. 2023, 24, 9714. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Baglieri, A.; Cadili, V.; Monterumici, C.M.; Gennari, M.; Tabasso, S.; Montoneri, E.; Nardi, S.; Negre, M. Fertilization of bean plants with tomato plants hydrolysates. Effect on biomass production, chlorophyll content and N assimilation. Sci. Hortic. 2014, 176, 194–199. [Google Scholar] [CrossRef]
- Pecha, J.; Fürst, T.; Kolomazník, K.; Friebrová, V.; Svoboda, P. Protein biostimulant foliar uptake modeling: The impact of climatic conditions. AIChE J. 2012, 58, 2010–2019. [Google Scholar] [CrossRef]
- Rajarajan, G.; Irshad, A.; Raghunath, B.; Mahesh Kumar, G.; Punnagaiarasi, A. Utilization of cheese industry whey for biofuel–ethanol production. In Integrated Waste Management in India: Status and Future Prospects for Environmental Sustainability; Springer: Berlin/Heidelberg, Germany, 2016; pp. 59–64. [Google Scholar]
- Moreno-Hernández, J.M.; Benítez-García, I.; Mazorra-Manzano, M.A.; Ramírez-Suárez, J.C.; Sánchez, E. Strategies for production, characterization and application of protein-based biostimulants in agriculture: A review. Chil. J. Agric. Res. 2020, 80, 274–289. [Google Scholar] [CrossRef]
- Martínez-Alvarez, O.; Chamorro, S.; Brenes, A. Protein hydrolysates from animal processing by-products as a source of bioactive molecules with interest in animal feeding: A review. Food Res. Int. 2015, 73, 204–212. [Google Scholar] [CrossRef]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein hydrolysates as biostimulants in horticulture. Sci. Hortic. 2015, 196, 28–38. [Google Scholar] [CrossRef]
- Ashaolu, T.J.; Zarei, M.; Agrawal, H.; Kharazmi, M.S.; Jafari, S.M. A critical review on immunomodulatory peptides from plant sources; action mechanisms and recent advances. Crit. Rev. Food Sci. Nutr. 2023, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Cavani, L.; Margon, A.; Sciubba, L.; Ciavatta, C.; Marzadori, C. What we talk about when we talk about protein hydrolyzate-based biostimulants. AIMS Agric. Food 2017, 2, 221–232. [Google Scholar] [CrossRef]
- Sun, W.; Shahrajabian, M.H.; Kuang, Y.; Wang, N. Amino Acids Biostimulants and Protein Hydrolysates in Agricultural Sciences. Plants 2024, 13, 210. [Google Scholar] [CrossRef] [PubMed]
- Ertani, A.; Pizzeghello, D.; Francioso, O.; Sambo, P.; Sanchez-Cortes, S.; Nardi, S. Capsicum chinensis L. growth and nutraceutical properties are enhanced by biostimulants in a long-term period: Chemical and metabolomic approaches. Front. Plant Sci. 2014, 5, 375. [Google Scholar] [CrossRef]
- Li, J.; Van Gerrewey, T.; Geelen, D. A meta-analysis of biostimulant yield effectiveness in field trials. Front. Plant Sci. 2022, 13, 836702. [Google Scholar] [CrossRef]
- Kauffman, G.L.; Kneivel, D.P.; Watschke, T.L. Effects of a biostimulant on the heat tolerance associated with photosynthetic capacity, membrane thermostability, and polyphenol production of perennial ryegrass. Crop Sci. 2007, 47, 261–267. [Google Scholar] [CrossRef]
- Edward, K.; Aneta, G.; Agnieszka, S.; Renata, W. The Effect of Cultivar Type, Time of Cultivation, and Biostimulant Treatment on the Yield of Spinach (Spinacia oleracea L.); Sciendo: Warsaw, Poland, 2010; Volume 22, pp. 9–13. [Google Scholar]
- Boscaro, R.; Panozzo, A.; Piotto, S.; Moore, S.S.; Barion, G.; Wang, Y.; Vamerali, T. Effects of Foliar-Applied Mixed Mineral Fertilizers and Organic Biostimulants on the Growth and Hybrid Seed Production of a Male-Sterile Inbred Maize Line. Plants 2023, 12, 2837. [Google Scholar] [CrossRef]
- Shahrajabian, M.H.; Sun, W. Sustainable approaches to boost yield and chemical constituents of aromatic and medicinal plants by application of biostimulants. Recent Adv. Food Nutr. Agric. 2022, 13, 72–92. [Google Scholar] [CrossRef]
- Cerdán, M.; Sánchez-Sánchez, A.; Oliver, M.; Juárez, M.; Sánchez-Andreu, J. Effect of foliar and root applications of amino acids on iron uptake by tomato plants. In Proceedings of the IV Balkan Symposium on Vegetables and Potatoes 830, Plovdiv, Bulgaria, 9–12 September 2008; pp. 481–488. [Google Scholar]
- Halpern, M.; Bar-Tal, A.; Ofek, M.; Minz, D.; Muller, T.; Yermiyahu, U. The use of biostimulants for enhancing nutrient uptake. Adv. Agron. 2015, 130, 141–174. [Google Scholar]
- Ubeda-Tomás, S.; Beemster, G.T.; Bennett, M.J. Hormonal regulation of root growth: Integrating local activities into global behaviour. Trends Plant Sci. 2012, 17, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Trevisan, S.; Manoli, A.; Quaggiotti, S. A novel biostimulant, belonging to protein hydrolysates, mitigates abiotic stress effects on maize seedlings grown in hydroponics. Agronomy 2019, 9, 28. [Google Scholar] [CrossRef]
- Trouvelot, S.; Héloir, M.-C.; Poinssot, B.; Gauthier, A.; Paris, F.; Guillier, C.; Combier, M.; Trdá, L.; Daire, X.; Adrian, M. Carbohydrates in plant immunity and plant protection: Roles and potential application as foliar sprays. Front. Plant Sci. 2014, 5, 592. [Google Scholar] [CrossRef] [PubMed]
- Franklin, O.; Cambui, C.A.; Gruffman, L.; Palmroth, S.; Oren, R.; Näsholm, T. The carbon bonus of organic nitrogen enhances nitrogen use efficiency of plants. Plant Cell Environ. 2017, 40, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Ertani, A.; Schiavon, M.; Nardi, S. Transcriptome-wide identification of differentially expressed genes in Solanum lycopersicon L. in response to an alfalfa-protein hydrolysate using microarrays. Front. Plant Sci. 2017, 8, 1159. [Google Scholar] [CrossRef] [PubMed]
- Mrid, R.B.; Benmrid, B.; Hafsa, J.; Boukcim, H.; Sobeh, M.; Yasri, A. Secondary metabolites as biostimulant and bioprotectant agents: A review. Sci. Total Environ. 2021, 777, 146204. [Google Scholar] [CrossRef]
- Paul, K.; Sorrentino, M.; Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Reynaud, H.; Canaguier, R.; Trtílek, M.; Panzarová, K. Understanding the biostimulant action of vegetal-derived protein hydrolysates by high-throughput plant phenotyping and metabolomics: A case study on tomato. Front. Plant Sci. 2019, 10, 47. [Google Scholar] [CrossRef]
- Casadesús, A.; Pérez-Llorca, M.; Munné-Bosch, S.; Polo, J. An enzymatically hydrolyzed animal protein-based biostimulant (Pepton) increases salicylic acid and promotes growth of tomato roots under temperature and nutrient stress. Front. Plant Sci. 2020, 11, 953. [Google Scholar] [CrossRef]
- Ertani, A.; Cavani, L.; Pizzeghello, D.; Brandellero, E.; Altissimo, A.; Ciavatta, C.; Nardi, S. Biostimulant activity of two protein hydrolyzates in the growth and nitrogen metabolism of maize seedlings. J. Plant Nutr. Soil Sci. 2009, 172, 237–244. [Google Scholar] [CrossRef]
- Hirel, B.; Tétu, T.; Lea, P.J.; Dubois, F. Improving nitrogen use efficiency in crops for sustainable agriculture. Sustainability 2011, 3, 1452–1485. [Google Scholar] [CrossRef]
- Tripathi, S.; Srivastava, P.; Devi, R.S.; Bhadouria, R. Influence of synthetic fertilizers and pesticides on soil health and soil microbiology. In Agrochemicals Detection, Treatment and Remediation; Elsevier: Amsterdam, The Netherlands, 2020; pp. 25–54. [Google Scholar]
- Kaur, B.; Kaur, G.; Asthir, B. Biochemical aspects of nitrogen use efficiency: An overview. J. Plant Nutr. 2017, 40, 506–523. [Google Scholar] [CrossRef]
- The, S.V.; Snyder, R.; Tegeder, M. Targeting nitrogen metabolism and transport processes to improve plant nitrogen use efficiency. Front. Plant Sci. 2021, 11, 628366. [Google Scholar] [CrossRef] [PubMed]
- Sétif, P.; Hirasawa, M.; Cassan, N.; Lagoutte, B.; Tripathy, J.N.; Knaff, D.B. New insights into the catalytic cycle of plant nitrite reductase. Electron transfer kinetics and charge storage. Biochemistry 2009, 48, 2828–2838. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Azad, N.; Behmanesh, J.; Rezaverdinejad, V.; Abbasi, F.; Navabian, M. An analysis of optimal fertigation implications in different soils on reducing environmental impacts of agricultural nitrate leaching. Sci. Rep. 2020, 10, 7797. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, M.; Muduli, M.; Ray, S. A comprehensive review on nitrate pollution and its remediation: Conventional and recent approaches. Sustain. Water Resour. Manag. 2022, 8, 113. [Google Scholar] [CrossRef]
- Abascal, E.; Gómez-Coma, L.; Ortiz, I.; Ortiz, A. Global diagnosis of nitrate pollution in groundwater and review of removal technologies. Sci. Total Environ. 2022, 810, 152233. [Google Scholar] [CrossRef]
- Giles, J. Nitrogen study fertilizes fears of pollution. Nature 2005, 433, 791–792. [Google Scholar] [CrossRef]
- Duchenne-Moutien, R.A.; Neetoo, H. Climate change and emerging food safety issues: A review. J. Food Prot. 2021, 84, 1884–1897. [Google Scholar] [CrossRef] [PubMed]
- Congreves, K.A.; Otchere, O.; Ferland, D.; Farzadfar, S.; Williams, S.; Arcand, M.M. Nitrogen use efficiency definitions of today and tomorrow. Front. Plant Sci. 2021, 12, 637108. [Google Scholar] [CrossRef] [PubMed]
- Quille, P.; Claffey, A.; Feeney, E.; Kacprzyk, J.; Ng, C.K.-Y.; O’Connell, S. The effect of an engineered biostimulant derived from Ascophyllum nodosum on grass yield under a reduced nitrogen regime in an agronomic setting. Agronomy 2022, 12, 463. [Google Scholar] [CrossRef]
- Cocetta, G.; Ertani, A.; Bulgari, R.; Franzoni, G.; Nicola, S.; Ferrante, A. Biostimulants and Plant Response Under Adverse Environmental Conditions: A Functional Interplay. In Plant Performance Under Environmental Stress: Hormones, Biostimulants and Sustainable Plant Growth Management; Springer: Berlin/Heidelberg, Germany, 2021; pp. 417–436. [Google Scholar]
- Petropoulos, S.A. Practical applications of plant biostimulants in greenhouse vegetable crop production. Agronomy 2020, 10, 1569. [Google Scholar] [CrossRef]
- Parks, S.E.; Irving, D.E.; Milham, P.J. A critical evaluation of on-farm rapid tests for measuring nitrate in leafy vegetables. Sci. Hortic. 2012, 134, 1–6. [Google Scholar] [CrossRef]
- Swanson, E.O.; Carlson, J.L.; Perkus, L.A.; Grossman, J.; Rogers, M.A.; Erwin, J.E.; Slavin, J.L.; Rosen, C.J. Nutrient and nitrate composition of greenhouse-grown leafy greens: A trial comparison between conventional and organic fertility treatments. Front. Sustain. Food Syst. 2022, 6, 811995. [Google Scholar] [CrossRef]
- Rocha, B.S. The nitrate-nitrite-nitric oxide pathway on healthy ageing: A review of pre-clinical and clinical data on the impact of dietary nitrate in the elderly. Front. Aging 2021, 2, 778467. [Google Scholar] [CrossRef] [PubMed]
- Sundqvist, M.L.; Larsen, F.J.; Carlström, M.; Bottai, M.; Pernow, J.; Hellénius, M.-L.; Weitzberg, E.; Lundberg, J.O. A randomized clinical trial of the effects of leafy green vegetables and inorganic nitrate on blood pressure. Am. J. Clin. Nutr. 2020, 111, 749–756. [Google Scholar] [CrossRef]
- Ahmed, M.; Rauf, M.; Akhtar, M.; Mukhtar, Z.; Saeed, N.A. Hazards of nitrogen fertilizers and ways to reduce nitrate accumulation in crop plants. Environ. Sci. Pollut. Res. 2020, 27, 17661–17670. [Google Scholar] [CrossRef]
- Sindelar, J.J.; Milkowski, A.L. Human safety controversies surrounding nitrate and nitrite in the diet. Nitric Oxide 2012, 26, 259–266. [Google Scholar] [CrossRef]
- Luetic, S.; Knezovic, Z.; Jurcic, K.; Majic, Z.; Tripkovic, K.; Sutlovic, D. Leafy Vegetable Nitrite and Nitrate Content: Potential Health Effects. Foods 2023, 12, 1655. [Google Scholar] [CrossRef] [PubMed]
- Salehzadeh, H.; Maleki, A.; Rezaee, R.; Shahmoradi, B.; Ponnet, K. The nitrate content of fresh and cooked vegetables and their health-related risks. PLoS ONE 2020, 15, e0227551. [Google Scholar] [CrossRef] [PubMed]
- Luo, F.; Yan, X.-J.; Hu, X.-F.; Yan, L.-J.; Cao, M.-Y.; Zhang, W.-J. Nitrate Quantification in Fresh Vegetables in Shanghai: Its Dietary Risks and Preventive Measures. Int. J. Environ. Res. Public Health 2022, 19, 14487. [Google Scholar] [CrossRef] [PubMed]
- Nájera, C.; Urrestarazu, M. Effect of the intensity and spectral quality of LED light on yield and nitrate accumulation in vegetables. Hortscience 2019, 54, 1745–1750. [Google Scholar] [CrossRef]
- Agostini, F.; Tei, F.; Silgram, M.; Farneselli, M.; Benincasa, P.; Aller, M. Decreasing nitrate leaching in vegetable crops with better N management. In Genetic Engineering, Biofertilisation, Soil Quality and Organic Farming; Springer: Berlin/Heidelberg, Germany, 2010; pp. 147–200. [Google Scholar]
- Bindraban, P.S.; Dimkpa, C.; Nagarajan, L.; Roy, A.; Rabbinge, R. Revisiting fertilisers and fertilisation strategies for improved nutrient uptake by plants. Biol. Fertil. Soils 2015, 51, 897–911. [Google Scholar] [CrossRef]
- Bian, Z.; Wang, Y.; Zhang, X.; Li, T.; Grundy, S.; Yang, Q.; Cheng, R. A review of environment effects on nitrate accumulation in leafy vegetables grown in controlled environments. Foods 2020, 9, 732. [Google Scholar] [CrossRef] [PubMed]
- Bulgari, R.; Cocetta, G.; Trivellini, A.; Vernieri, P.; Ferrante, A. Biostimulants and crop responses: A review. Biol. Agric. Hortic. 2015, 31, 1–17. [Google Scholar] [CrossRef]
- du Jardin, P.; Xu, L.; Geelen, D. Agricultural functions and action mechanisms of plant biostimulants (pbs) an introduction. In The Chemical Biology of Plant Biostimulants; Wiley: Hoboken, NJ, USA, 2020; pp. 1–30. [Google Scholar]
- Nardi, S.; Pizzeghello, D.; Schiavon, M.; Ertani, A. Plant biostimulants: Physiological responses induced by protein hydrolyzed-based products and humic substances in plant metabolism. Sci. Agric. 2016, 73, 18–23. [Google Scholar] [CrossRef]
- Sabatino, L.; Consentino, B.B.; Rouphael, Y.; De Pasquale, C.; Iapichino, G.; D’Anna, F.; La Bella, S. Protein hydrolysates and mo-biofortification interactively modulate plant performance and quality of ‘canasta’ lettuce grown in a protected environment. Agronomy 2021, 11, 1023. [Google Scholar] [CrossRef]
- Consentino, B.B.; Vultaggio, L.; Sabatino, L.; Ntatsi, G.; Rouphael, Y.; Bondì, C.; De Pasquale, C.; Guarino, V.; Iacuzzi, N.; Capodici, G. Combined effects of biostimulants, N level and drought stress on yield, quality and physiology of greenhouse-grown basil. Plant Stress 2023, 10, 100268. [Google Scholar] [CrossRef]
- Tsouvaltzis, P.; Koukounaras, A.; Siomos, A.S. Application of amino acids improves lettuce crop uniformity and inhibits nitrate accumulation induced by the supplemental inorganic nitrogen fertilization. Int. J. Agric. Biol. 2014, 16, 951. [Google Scholar]
- Caruso, G.; El-Nakhel, C.; Rouphael, Y.; Comite, E.; Lombardi, N.; Cuciniello, A.; Woo, S.L. Diplotaxis tenuifolia (L.) DC. yield and quality as influenced by cropping season, protein hydrolysates, and Trichoderma applications. Plants 2020, 9, 697. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Carillo, P.; Cristofano, F.; Cardarelli, M.; Colla, G. Effects of vegetal-versus animal-derived protein hydrolysate on sweet basil morpho-physiological and metabolic traits. Sci. Hortic. 2021, 284, 110123. [Google Scholar] [CrossRef]
- Carillo, P.; Colla, G.; Fusco, G.M.; Dell’Aversana, E.; El-Nakhel, C.; Giordano, M.; Pannico, A.; Cozzolino, E.; Mori, M.; Reynaud, H. Morphological and physiological responses induced by protein hydrolysate-based biostimulant and nitrogen rates in greenhouse spinach. Agronomy 2019, 9, 450. [Google Scholar] [CrossRef]
- Navarro-León, E.; López-Moreno, F.J.; Borda, E.; Marín, C.; Sierras, N.; Blasco, B.; Ruiz, J.M. Effect of l-amino acid-based biostimulants on nitrogen use efficiency (NUE) in lettuce plants. J. Sci. Food Agric. 2022, 102, 7098–7106. [Google Scholar] [CrossRef] [PubMed]
- Di Mola, I.; Ottaiano, L.; Cozzolino, E.; Senatore, M.; Giordano, M.; El-Nakhel, C.; Sacco, A.; Rouphael, Y.; Colla, G.; Mori, M. Plant-based biostimulants influence the agronomical, physiological, and qualitative responses of baby rocket leaves under diverse nitrogen conditions. Plants 2019, 8, 522. [Google Scholar] [CrossRef] [PubMed]
- Bonasia, A.; Conversa, G.; Lazzizera, C.; Elia, A. Foliar application of protein hydrolysates on baby-leaf spinach grown at different n levels. Agronomy 2021, 12, 36. [Google Scholar] [CrossRef]
- Cristofano, F.; El-Nakhel, C.; Pannico, A.; Giordano, M.; Colla, G.; Rouphael, Y. Foliar and root applications of vegetal-derived protein hydrolysates differentially enhance the yield and qualitative attributes of two lettuce cultivars grown in floating system. Agronomy 2021, 11, 1194. [Google Scholar] [CrossRef]
- Choi, S.; Colla, G.; Cardarelli, M.; Kim, H.-J. Effects of plant-derived protein hydrolysates on yield, quality, and nitrogen use efficiency of greenhouse grown lettuce and tomato. Agronomy 2022, 12, 1018. [Google Scholar] [CrossRef]
- Di Mola, I.; Cozzolino, E.; Ottaiano, L.; Nocerino, S.; Rouphael, Y.; Colla, G.; El-Nakhel, C.; Mori, M. Nitrogen use and uptake efficiency and crop performance of baby spinach (Spinacia oleracea L.) and Lamb’s Lettuce (Valerianella locusta L.) grown under variable sub-optimal N regimes combined with plant-based biostimulant application. Agronomy 2020, 10, 278. [Google Scholar] [CrossRef]
- Aktsoglou, D.-C.; Kasampalis, D.S.; Sarrou, E.; Tsouvaltzis, P.; Chatzopoulou, P.; Martens, S.; Siomos, A.S. Protein hydrolysates supplement in the nutrient solution of soilless grown fresh peppermint and spearmint as a tool for improving product quality. Agronomy 2021, 11, 317. [Google Scholar] [CrossRef]
- Ciriello, M.; Formisano, L.; El-Nakhel, C.; Corrado, G.; Rouphael, Y. Biostimulatory action of a plant-derived protein hydrolysate on morphological traits, photosynthetic parameters, and mineral composition of two basil cultivars grown hydroponically under variable electrical conductivity. Horticulturae 2022, 8, 409. [Google Scholar] [CrossRef]
- Carillo, P.; De Micco, V.; Ciriello, M.; Formisano, L.; El-Nakhel, C.; Giordano, M.; Colla, G.; Rouphael, Y. Morpho-anatomical, physiological, and mineral composition responses induced by a vegetal-based biostimulant at three rates of foliar application in greenhouse lettuce. Plants 2022, 11, 2030. [Google Scholar] [CrossRef]
- He, G.; Liu, X.; Cui, Z. Achieving global food security by focusing on nitrogen efficiency potentials and local production. Glob. Food Secur. 2021, 29, 100536. [Google Scholar] [CrossRef]
- Ren, C.; Zhang, X.; Reis, S.; Gu, B. Socioeconomic barriers of nitrogen management for agricultural and environmental sustainability. Agric. Ecosyst. Environ. 2022, 333, 107950. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciriello, M.; Campana, E.; De Pascale, S.; Rouphael, Y. Implications of Vegetal Protein Hydrolysates for Improving Nitrogen Use Efficiency in Leafy Vegetables. Horticulturae 2024, 10, 132. https://doi.org/10.3390/horticulturae10020132
Ciriello M, Campana E, De Pascale S, Rouphael Y. Implications of Vegetal Protein Hydrolysates for Improving Nitrogen Use Efficiency in Leafy Vegetables. Horticulturae. 2024; 10(2):132. https://doi.org/10.3390/horticulturae10020132
Chicago/Turabian StyleCiriello, Michele, Emanuela Campana, Stefania De Pascale, and Youssef Rouphael. 2024. "Implications of Vegetal Protein Hydrolysates for Improving Nitrogen Use Efficiency in Leafy Vegetables" Horticulturae 10, no. 2: 132. https://doi.org/10.3390/horticulturae10020132