

Effect of Biofertilizers on Broccoli Yield and Soil Quality Indicators
 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Experimental Design
2.2. Soil Greenhouse Gas Measurements
2.3. Soil and Plant Sampling
2.4. Soil Chemical and Biochemical Analyses
2.5. DNA Extraction and Quantitative PCR (qPCR) Gene Analysis
2.6. Phospholipid Fatty Acid (PLFA) Analysis
2.7. Statistical Analysis
3. Results
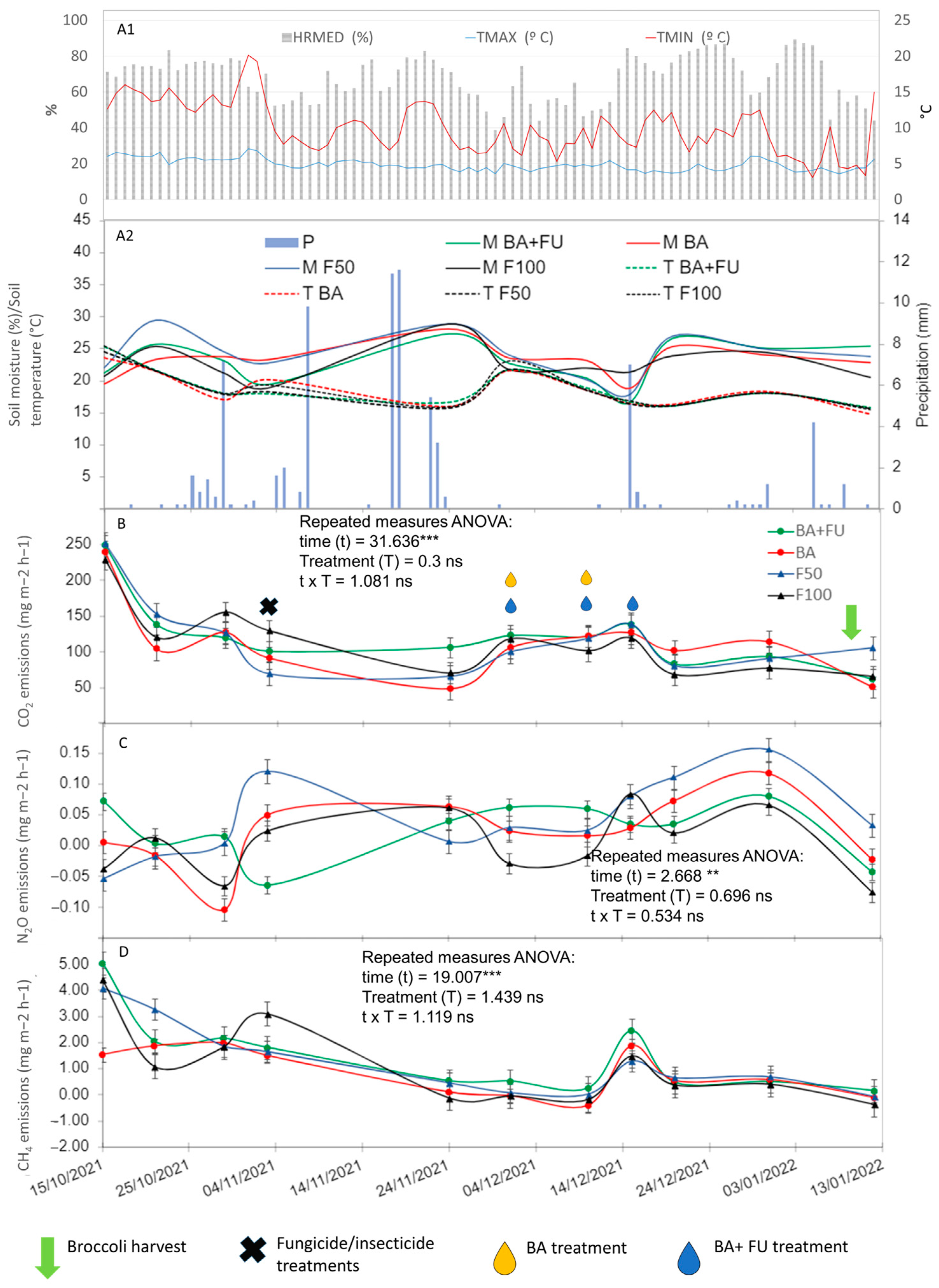
3.1. Soil Greenhouse Gas Emission Rates
3.2. Overall Cumulative Soil Emissions
3.3. Crop Yield, Soil Enzyme Activities, and Chemical Properties
3.4. PLFA Biomarkers and Functional Genes
3.5. Interrelationship between GHGs, Soil Properties and Microbial Abundance and Functioning
4. Discussion
4.1. GHG Emissions
4.2. Soil Chemical Properties, Microbial Abundance, and Potential Activity
4.3. Crop Yield
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cooper, R.N.; Houghton, J.T.; McCarthy, J.J.; Metz, B. Climate Change 2001: The Scientific Basis. Foreign Aff. 2002, 81, 208. [Google Scholar] [CrossRef]
- Oertel, C.; Matschullat, J.; Zurba, K.; Zimmermann, F.; Erasmi, S. Greenhouse Gas Emissions from Soils—A Review. Geochemistry 2016, 76, 327–352. [Google Scholar] [CrossRef]
- Prather, M.; Ehhalt, D.; Dentener, F.; Derwent, R.; Dlugokencky, E.; Holland, E.; Isaksen, I.; Katima, J.; Kirchhoff, V.; Matson, P.; et al. Atmospheric Chemistry and Greenhouse Gases. In Precise Soil Management as a Tool to Reduce CH4 and N2O Emissions from Agricultural Soils; Houghton, J.T., Ed.; Cambridge University Press: New York, NY, USA, 2001; pp. 239–287. [Google Scholar]
- Mosquera, J.; Hilhorst, M.A. Precise Soil Management as a Tool to Reduce N2O and CH4 Emissions from Agricultural Soils: Literature Review. In Proceedings of the 4th International Symposium on non-CO2 Greenhouse Gases (NCGG-4), Science, Control, Policy and Implementation, Utrecht, The Netherlands, 4–6 July 2005; Millpress Science Publishers: Rotterdam, The Netherlands, 2005; pp. 5–12. [Google Scholar]
- Ludwig, J.; Meixner, F.; Vogel, B.; Förstner, J. Soil-Air Exchange of Nitric Oxide: An Overview of Processes, Environmental Factors, and Modeling Studies. Biogeochemistry 2001, 52, 225–257. [Google Scholar] [CrossRef]
- Maraseni, T.N.; Cockfield, G. Including the Costs of Water and Greenhouse Gas Emissions in a Reassessment of the Profitability of Irrigation. Agric. Water Manag. 2012, 103, 25–32. [Google Scholar] [CrossRef]
- Iqbal, J.; Ronggui, H.; Lijun, D.; Lan, L.; Shan, L.; Tao, C.; Leilei, R. Differences in Soil CO2 Flux between Different Land Use Types in Mid-Subtropical China. Soil Biol. Biochem. 2008, 40, 2324–2333. [Google Scholar] [CrossRef]
- Sainju, U.M.; Stevens, W.B.; Caesar-TonThat, T.; Jabro, J.D. Land Use and Management Practices Impact on Plant Biomass Carbon and Soil Carbon Dioxide Emission. Soil Sci. Soc. Am. J. 2010, 74, 1613–1622. [Google Scholar] [CrossRef]
- Linquist, B.A.; Adviento-Borbe, M.A.; Pittelkow, C.M.; van Kessel, C.; van Groenigen, K.J. Fertilizer Management Practices and Greenhouse Gas Emissions from Rice Systems: A Quantitative Review and Analysis. Field Crops Res. 2012, 135, 10–21. [Google Scholar] [CrossRef]
- Piva, J.T.; Dieckow, J.; Bayer, C.; Zanatta, J.A.; de Moraes, A.; Tomazi, M.; Pauletti, V.; Barth, G.; Piccolo, M.d.C. Soil Gaseous N2O and CH4 Emissions and Carbon Pool Due to Integrated Crop-Livestock in a Subtropical Ferralsol. Agric. Ecosyst. Environ. 2014, 190, 87–93. [Google Scholar] [CrossRef]
- Sanz-Cobena, A.; García-Marco, S.; Quemada, M.; Gabriel, J.L.; Almendros, P.; Vallejo, A. Do Cover Crops Enhance N2O, CO2 or CH4 Emissions from Soil in Mediterranean Arable Systems? Sci. Total Environ. 2014, 466–467, 164–174. [Google Scholar] [CrossRef]
- Schaufler, G.; Kitzler, B.; Schindlbacher, A.; Skiba, U.; Sutton, M.A.; Zechmeister-Boltenstern, S. Greenhouse Gas Emissions from European Soils under Different Land Use: Effects of Soil Moisture and Temperature. Eur. J. Soil Sci. 2010, 61, 683–696. [Google Scholar] [CrossRef]
- Yan, H.; Xie, L.; Guo, L.; Fan, J.; Diao, T.; Lin, M.; Zhang, H.; Lin, E. Characteristics of Nitrous Oxide Emissions and the Affecting Factors from Vegetable Fields on the North China Plain. J. Environ. Manag. 2014, 144, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Zornoza, R.; Acosta, J.A.; Gabarrón, M.; Gómez-Garrido, M.; Sánchez-Navarro, V.; Terrero, A.; Martínez-Martínez, S.; Faz, Á.; Pérez-Pastor, A. Greenhouse Gas Emissions and Soil Organic Matter Dynamics in Woody Crop Orchards with Different Irrigation Regimes. Sci. Total Environ. 2018, 644, 1429–1438. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.; Song, C.; Li, Y.; Wang, J.; Song, Y.; Wang, X. Effects of Water Table Changes on Soil CO2, CH4 and N2O Fluxes during the Growing Season in Freshwater Marsh of Northeast China. Environ. Earth Sci. 2013, 69, 1963–1971. [Google Scholar] [CrossRef]
- Ren, C.; Wang, T.; Xu, Y.; Deng, J.; Zhao, F.; Yang, G.; Han, X.; Feng, Y.; Ren, G. Differential Soil Microbial Community Responses to the Linkage of Soil Organic Carbon Fractions with Respiration across Land-Use Changes. For. Ecol. Manag. 2018, 409, 170–178. [Google Scholar] [CrossRef]
- Sanz-Cobena, A.; Lassaletta, L.; Aguilera, E.; del Prado, A.; Garnier, J.; Billen, G.; Iglesias, A.; Sánchez, B.; Guardia, G.; Abalos, D.; et al. Strategies for Greenhouse Gas Emissions Mitigation in Mediterranean Agriculture: A Review. Agric. Ecosyst. Environ. 2017, 238, 5–24. [Google Scholar] [CrossRef]
- Hui, K.; Yuan, Y.; Xi, B.; Tan, W. A Review of the Factors Affecting the Emission of the Ozone Chemical Precursors VOCs and NOx from the Soil. Environ. Int. Anderson, I.C.; Levine, J.S. Relative Rates of Nitric Oxide and Nitrous Oxide Production by Nitrifiers, Denitrifiers, and Nitrate Respirers. Environ. Int. 2023, 172, 107799. [Google Scholar] [PubMed]
- Abbasi, M.K.; Adams, W.A. Gaseous N Emission during Simultaneous Nitrification–Denitrification Associated with Mineral N Fertilization to a Grassland Soil under Field Conditions. Soil Biol. Biochem. 2000, 32, 1251–1259. [Google Scholar] [CrossRef]
- Wang, X.; Zou, C.; Gao, X.; Guan, X.; Zhang, W.; Zhang, Y.; Shi, X.; Chen, X. Nitrous Oxide Emissions in Chinese Vegetable Systems: A Meta-Analysis. Environ. Pollut. 2018, 239, 375–383. [Google Scholar] [CrossRef]
- Senbayram, M.; Chen, R.; Mühling, K.H.; Dittert, K. Contribution of Nitrification and Denitrification to Nitrous Oxide Emissions from Soils after Application of Biogas Waste and Other Fertilizers. Rapid Commun. Mass Spectrom. 2009, 23, 2489–2498. [Google Scholar] [CrossRef]
- Hou, A.X.; Chen, G.X.; Wang, Z.P.; Van Cleemput, O.; Patrick, W.H., Jr. Methane and Nitrous Oxide Emissions from a Rice Field in Relation to Soil Redox and Microbiological Processes. Soil Sci. Soc. Am. J. 2000, 64, 2180–2186. [Google Scholar] [CrossRef]
- Foley, J.A.; DeFries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global Consequences of Land Use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Riaz, U.; Murtaza, D.G.; Anum, W.; Samreen, T.; Sarfraz, M.; Nazir, M. Plant Growth-Promoting Rhizobacteria (PGPR) as Biofertilizers and Biopesticides. In Microbiota and Biofertilizers; Springer: Cham, Switzerland, 2020; pp. 181–196. ISBN 978-3-030-48770-6. [Google Scholar]
- Hossain, M.M.; Sultana, F.; Islam, S. Plant Growth-Promoting Fungi (PGPF): Phytostimulation and Induced Systemic Resistance. In Plant-Microbe Interactions in Agro-Ecological Perspectives: Volume 2: Microbial Interactions and Agro-Ecological Impacts; Singh, D.P., Singh, H.B., Prabha, R., Eds.; Springer: Singapore, 2017; pp. 135–191. ISBN 978-981-10-6593-4. [Google Scholar]
- El-Saadony, M.T.; Saad, A.M.; Soliman, S.M.; Salem, H.M.; Ahmed, A.I.; Mahmood, M.; El-Tahan, A.M.; Ebrahim, A.A.M.; Abd El-Mageed, T.A.; Negm, S.H.; et al. Plant Growth-Promoting Microorganisms as Biocontrol Agents of Plant Diseases: Mechanisms, Challenges and Future Perspectives. Front. Plant Sci. 2022, 13, 923880. [Google Scholar] [CrossRef] [PubMed]
- Malusá, E.; Sas-Paszt, L.; Ciesielska, J. Technologies for Beneficial Microorganisms Inocula Used as Biofertilizers. Sci. World J. 2012, 2012, e491206. [Google Scholar] [CrossRef] [PubMed]
- Vessey, J.K. Plant Growth Promoting Rhizobacteria as Biofertilizers. Plant Soil 2003, 255, 571–586. [Google Scholar] [CrossRef]
- Xu, J.; Kloepper, J.W.; Huang, P.; McInroy, J.A.; Hu, C.H. Isolation and Characterization of N2-Fixing Bacteria from Giant Reed and Switchgrass for Plant Growth Promotion and Nutrient Uptake. J. Basic Microbiol. 2018, 58, 459–471. [Google Scholar] [CrossRef]
- Khan, M.S.; Zaidi, A.; Wani, P.A. Role of Phosphate-Solubilizing Microorganisms in Sustainable Agriculture—A Review. Agron. Sustain. Dev. 2007, 27, 29–43. [Google Scholar] [CrossRef]
- Ali, A.M.; Awad, M.Y.M.; Hegab, S.A.; Gawad, A.M.A.E.; Eissa, M.A. Effect of Potassium Solubilizing Bacteria (Bacillus cereus) on Growth and Yield of Potato. J. Plant Nutr. 2021, 44, 411–420. [Google Scholar] [CrossRef]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural Uses of Plant Biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef]
- Sarfraz, R.; Hussain, A.; Sabir, A.; Ben Fekih, I.; Ditta, A.; Xing, S. Role of Biochar and Plant Growth Promoting Rhizobacteria to Enhance Soil Carbon Sequestration—A Review. Env. Monit. Assess. 2019, 191, 251. [Google Scholar] [CrossRef]
- Pankievicz, V.C.S.; do Amaral, F.P.; Santos, K.F.D.N.; Agtuca, B.; Xu, Y.; Schueller, M.J.; Arisi, A.C.M.; Steffens, M.B.; de Souza, E.M.; Pedrosa, F.O.; et al. Robust Biological Nitrogen Fixation in a Model Grass–Bacterial Association. Plant J. 2015, 81, 907–919. [Google Scholar] [CrossRef]
- Florio, A.; Bréfort, C.; Gervaix, J.; Bérard, A.; Le Roux, X. The Responses of NO2- and N2O-Reducing Bacteria to Maize Inoculation by the PGPR Azospirillum Lipoferum CRT1 Depend on Carbon Availability and Determine Soil Gross and Net N2O Production. Soil Biol. Biochem. 2019, 136, 107524. [Google Scholar] [CrossRef]
- Huang, M.; Zhang, Y.; Wu, J.; Wang, Y.; Xie, Y.; Geng, Y.; Zhang, N.; Michelsen, A.; Li, S.; Zhang, R.; et al. Bacillus Velezensis SQR9 Inhibition to Fungal Denitrification Responsible for Decreased N2O Emissions from Acidic Soils. Sci. Total Environ. 2023, 885, 163789. [Google Scholar] [CrossRef] [PubMed]
- Paliwoda, D.; Mikiciuk, G.; Mikiciuk, M.; Miller, T.; Kisiel, A.; Sas-Paszt, L.; Kozioł, A.; Brysiewicz, A. The Use of Plant Growth Promoting Rhizobacteria to Reduce Greenhouse Gases in Strawberry Cultivation under Different Soil Moisture Conditions. Agronomy 2023, 13, 754. [Google Scholar] [CrossRef]
- Dimkpa, C.; Weinand, T.; Asch, F. Plant–Rhizobacteria Interactions Alleviate Abiotic Stress Conditions. Plant Cell Environ. 2009, 32, 1682–1694. [Google Scholar] [CrossRef] [PubMed]
- Malgioglio, G.; Rizzo, G.F.; Nigro, S.; Lefebvre du Prey, V.; Herforth-Rahmé, J.; Catara, V.; Branca, F. Plant-Microbe Interaction in Sustainable Agriculture: The Factors That May Influence the Efficacy of PGPM Application. Sustainability 2022, 14, 2253. [Google Scholar] [CrossRef]
- Zhou, X.; Smaill, S.J.; Clinton, P.W. Methane Oxidation Needs Less Stressed Plants. Trends Plant Sci. 2013, 18, 657–659. [Google Scholar] [CrossRef]
- Shah, A.M.; Khan, I.M.; Shah, T.I.; Bangroo, S.A.; Kirmani, N.A.; Nazir, S.; Malik, A.R.; Aezum, A.M.; Mir, Y.H.; Hilal, A.; et al. Soil Microbiome: A Treasure Trove for Soil Health Sustainability under Changing Climate. Land 2022, 11, 1887. [Google Scholar] [CrossRef]
- Ortega Pérez, R.; Nieto García, J.C.; Gallegos-Cedillo, V.M.; Domene Ruiz, M.Á.; Santos Hernández, M.; Nájera, C.; Miralles Mellado, I.; Diánez Martínez, F. Biofertilizers Enriched with PGPB Improve Soil Fertility and the Productivity of an Intensive Tomato Crop. Agronomy 2023, 13, 2286. [Google Scholar] [CrossRef]
- Lori, M.; Symnaczik, S.; Mäder, P.; De Deyn, G.; Gattinger, A. Organic Farming Enhances Soil Microbial Abundance and Activity—A Meta-Analysis and Meta-Regression. PLoS ONE 2017, 12, e0180442. [Google Scholar] [CrossRef]
- Chen, X.-P.; Zhu, Y.-G.; Xia, Y.; Shen, J.-P.; He, J.-Z. Ammonia-Oxidizing Archaea: Important Players in Paddy Rhizosphere Soil? Environ. Microbiol. 2008, 10, 1978–1987. [Google Scholar] [CrossRef]
- Henry, S.; Bru, D.; Stres, B.; Hallet, S.; Philippot, L. Quantitative Detection of the nosZ Gene, Encoding Nitrous Oxide Reductase, and Comparison of the Abundances of 16S rRNA, narG, nirK, and nosZ Genes in Soils. Appl. Environ. Microbiol. 2006, 72, 5181–5189. [Google Scholar] [CrossRef]
- Poly, F.; Ranjard, L.; Nazaret, S.; Gourbière, F.; Monrozier, L.J. Comparison of nifH Gene Pools in Soils and Soil Microenvironments with Contrasting Properties. Appl. Environ. Microbiol. 2001, 67, 2255–2262. [Google Scholar] [CrossRef]
- Yousuf, B.; Keshri, J.; Mishra, A.; Jha, B. Application of Targeted Metagenomics to Explore Abundance and Diversity of CO2-Fixing Bacterial Community Using cbbL Gene from the Rhizosphere of Arachis Hypogaea. Gene 2012, 506, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Fan, F.; Li, Z.; Wakelin, S.A.; Yu, W.; Liang, Y. Mineral Fertilizer Alters Cellulolytic Community Structure and Suppresses Soil Cellobiohydrolase Activity in a Long-Term Fertilization Experiment. Soil Biol. Biochem. 2012, 55, 70–77. [Google Scholar] [CrossRef]
- Henry, H.A.L.; Juarez, J.D.; Field, C.B.; Vitousek, P.M. Interactive Effects of Elevated CO2, N Deposition and Climate Change on Extracellular Enzyme Activity and Soil Density Fractionation in a California Annual Grassland. Glob. Chang. Biol. 2005, 11, 1808–1815. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Cusack, D.; Frey, S.; Gallo, M.E.; et al. Stoichiometry of Soil Enzyme Activity at Global Scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef]
- Acosta-Martinez, V.; Cano, A.; Johnson, J. Simultaneous Determination of Multiple Soil Enzyme Activities for Soil Health-Biogeochemical Indices. Appl. Soil Ecol. 2018, 126, 121–128. [Google Scholar] [CrossRef]
- Deng, S.P.; Tabatabai, M.A. Cellulase Activity of Soils. Soil Biol. Biochem. 1994, 26, 1347–1354. [Google Scholar] [CrossRef]
- Krajewska, B.; Ureases, I. Functional, Catalytic and Kinetic Properties: A Review. J. Mol. Catal. B Enzym. 2009, 59, 9–21. [Google Scholar] [CrossRef]
- Morales, S.E.; Cosart, T.; Holben, W.E. Bacterial Gene Abundances as Indicators of Greenhouse Gas Emission in Soils. ISME J. 2010, 4, 799–808. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial Diversity and Soil Functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef]
- Chaudhary, D.R.; Rathore, A.P.; Sharma, S. Effect of Halotolerant Plant Growth Promoting Rhizobacteria Inoculation on Soil Microbial Community Structure and Nutrients. Appl. Soil Ecol. 2020, 150, 103461. [Google Scholar] [CrossRef]
- Siddikee, M.A.; Zereen, M.I.; Li, C.-F.; Dai, C.-C. Endophytic Fungus Phomopsis Liquidambari and Different Doses of N-Fertilizer Alter Microbial Community Structure and Function in Rhizosphere of Rice. Sci. Rep. 2016, 6, 32270. [Google Scholar] [CrossRef]
- Wang, H.-W.; Zhu, Y.-X.; Xu, M.; Cai, X.-Y.; Tian, F. Co-Application of Spent Mushroom Substrate and PGPR Alleviates Tomato Continuous Cropping Obstacle by Regulating Soil Microbial Properties. Rhizosphere 2022, 23, 100563. [Google Scholar] [CrossRef]
- Sun, B.; Gu, L.; Bao, L.; Zhang, S.; Wei, Y.; Bai, Z.; Zhuang, G.; Zhuang, X. Application of Biofertilizer Containing Bacillus Subtilis Reduced the Nitrogen Loss in Agricultural Soil. Soil Biol. Biochem. 2020, 148, 107911. [Google Scholar] [CrossRef]
- Wu, S.; Zhuang, G.; Bai, Z.; Cen, Y.; Xu, S.; Sun, H.; Han, X.; Zhuang, X. Mitigation of Nitrous Oxide Emissions from Acidic Soils by Bacillus Amyloliquefaciens, a Plant Growth-Promoting Bacterium. Glob. Chang. Biol. 2018, 24, 2352–2365. [Google Scholar] [CrossRef]
- Conversa, G.; Lazzizera, C.; Bonasia, A.; Elia, A. Growth, N Uptake and N Critical Dilution Curve in Broccoli Cultivars Grown under Mediterranean Conditions. Sci. Hortic. 2019, 244, 109–121. [Google Scholar] [CrossRef]
- IUSS, WRB World Reference Base for Soil Resources. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps, 4th ed.; International Union of Soil Sciences (IUSS): Vienna, Austria, 2022. [Google Scholar]
- CARM Normativa Reguladora, Producción Integrada, Alimentos Sanos y Garantizados [Regulatory Rules, Integrated Production, Healthy Food and Guaranteed]; Consejería de Agricultura y Agua de La Región de Murcia: Murcia, Spain, 1998.
- He, Y.; Pantigoso, H.A.; Wu, Z.; Vivanco, J.M. Co-Inoculation of Bacillus sp. and Pseudomonas Putida at Different Development Stages Acts as a Biostimulant to Promote Growth, Yield and Nutrient Uptake of Tomato. J. Appl. Microbiol. 2019, 127, 196–207. [Google Scholar] [CrossRef]
- Álvaro-Fuentes, J.; Lóczy, D.; Thiele-Bruhn, S.; Zornoza, R. Handbook of Plant and Soil Analysis for Agricultural Systems; Universidad Politécnica de Cartagena: Cartagena, Spain, 2019; ISBN 978-84-16325-86-3. [Google Scholar]
- Chen, W.; Wang, Y.; Zhao, Z.; Cui, F.; Gu, J.; Zheng, X. The Effect of Planting Density on Carbon Dioxide, Methane and Nitrous Oxide Emissions from a Cold Paddy Field in the Sanjiang Plain, Northeast China. Agric. Ecosyst. Environ. 2013, 178, 64–70. [Google Scholar] [CrossRef]
- Zornoza, R.; Landi, L.; Nannipieri, P.; Renella, G. A Protocol for the Assay of Arylesterase Activity in Soil. Soil Biol. Biochem. 2009, 41, 659–662. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An Extraction Method for Measuring Soil Microbial Biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Keeney, D.R.; Nelson, D.w. Nitrogen—Inorganic Forms. In Methods of Soil Analysis; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1983; pp. 643–698. ISBN 978-0-89118-977-0. [Google Scholar]
- Kandeler, E.; Gerber, H. Short-Term Assay of Soil Urease Activity Using Colorimetric Determination of Ammonium. Biol. Fertil. Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Ekenler, M.; Tabatabai, M.A. β-Glucosaminidase Activity as an Index of Nitrogen Mineralization in Soils. Commun. Soil Sci. Plant Anal. 2004, 35, 1081–1094. [Google Scholar] [CrossRef]
- Garcia-Alvarez, A.; Ibañez, J.J. Seasonal Fluctuations and Crop Influence on Microbiota and Enzyme Activity in Fully Developed Soils of Central Spain. Arid Soil Res. Rehabil. 1994, 8, 161–178. [Google Scholar] [CrossRef]
- Nelson, N. A Photometric Adaptation of the Somogyi Method for the Determination of Glucose. J. Biol. Chem. 1944, 153, 375–380. [Google Scholar] [CrossRef]
- Nannipieri, P.; Johnson, R.L.; Paul, E.A. Criteria for Measurement of Microbial Growth and Activity in Soil. Soil Biol. Biochem. 1978, 10, 223–229. [Google Scholar] [CrossRef]
- Rotthauwe, J.H.; Witzel, K.P.; Liesack, W. The Ammonia Monooxygenase Structural Gene amoA as a Functional Marker: Molecular Fine-Scale Analysis of Natural Ammonia-Oxidizing Populations. Appl. Environ. Microbiol. 1997, 63, 4704–4712. [Google Scholar] [CrossRef] [PubMed]
- Henry, S.; Baudoin, E.; López-Gutiérrez, J.C.; Martin-Laurent, F.; Brauman, A.; Philippot, L. Quantification of Denitrifying Bacteria in Soils by nirK Gene Targeted Real-Time PCR. J. Microbiol. Methods 2004, 59, 327–335. [Google Scholar] [CrossRef]
- Selesi, D.; Schmid, M.; Hartmann, A. Diversity of Green-Like and Red-Like Ribulose-1,5-Bisphosphate Carboxylase/Oxygenase Large-Subunit Genes (cbbL) in Differently Managed Agricultural Soils. Appl. Environ. Microbiol. 2005, 71, 175–184. [Google Scholar] [CrossRef]
- Tian, X.; Yang, T.; He, J.; Chu, Q.; Jia, X.; Huang, J. Fungal Community and Cellulose-Degrading Genes in the Composting Process of Chinese Medicinal Herbal Residues. Bioresour. Technol. 2017, 241, 374–383. [Google Scholar] [CrossRef]
- Frostegård, Å.; Bååth, E.; Tunlio, A. Shifts in the Structure of Soil Microbial Communities in Limed Forests as Revealed by Phospholipid Fatty Acid Analysis. Soil Biol. Biochem. 1993, 25, 723–730. [Google Scholar] [CrossRef]
- Joergensen, R.G. Phospholipid Fatty Acids in Soil—Drawbacks and Future Prospects. Biol. Fertil. Soils 2022, 58, 1–6. [Google Scholar] [CrossRef]
- Tang, X.; Liu, S.; Zhou, G.; Zhang, D.; Zhou, C. Soil-Atmospheric Exchange of CO2, CH4, and N2O in Three Subtropical Forest Ecosystems in Southern China. Glob. Chang. Biol. 2006, 12, 546–560. [Google Scholar] [CrossRef]
- Iqbal, J.; Lin, S.; Hu, R.; Feng, M. Temporal Variability of Soil-Atmospheric CO2 and CH4 Fluxes from Different Land Uses in Mid-Subtropical China. Atmos. Environ. 2009, 43, 5865–5875. [Google Scholar] [CrossRef]
- Curiel Yuste, J.; Baldocchi, D.D.; Gershenson, A.; Goldstein, A.; Misson, L.; Wong, S. Microbial Soil Respiration and Its Dependency on Carbon Inputs, Soil Temperature and Moisture. Glob. Chang. Biol. 2007, 13, 2018–2035. [Google Scholar] [CrossRef]
- Arai, H.; Hadi, A.; Darung, U.; Limin, S.H.; Takahashi, H.; Hatano, R.; Inubushi, K. Land Use Change Affects Microbial Biomass and Fluxes of Carbon Dioxide and Nitrous Oxide in Tropical Peatlands. Soil Sci. Plant Nutr. 2014, 60, 423–434. [Google Scholar] [CrossRef]
- Furukawa, Y.; Inubushi, K.; Ali, M.; Itang, A.M.; Tsuruta, H. Effect of Changing Groundwater Levels Caused by Land-Use Changes on Greenhouse Gas Fluxes from Tropical Peat Lands. Nutr. Cycl. Agroecosystems 2005, 71, 81–91. [Google Scholar] [CrossRef]
- Li, Y.; Niu, W.; Wang, J.; Liu, L.; Zhang, M.; Xu, J. Effects of Artificial Soil Aeration Volume and Frequency on Soil Enzyme Activity and Microbial Abundance When Cultivating Greenhouse Tomato. Soil Sci. Soc. Am. J. 2016, 80, 1208–1221. [Google Scholar] [CrossRef]
- Zhu, Y.; Cai, H.; Song, L.; Chen, H. Aerated Irrigation Promotes Soil Respiration and Microorganism Abundance around Tomato Rhizosphere. Soil Sci. Soc. Am. J. 2019, 83, 1343–1355. [Google Scholar] [CrossRef]
- Mayer, H.P.; Conrad, R. Factors Influencing the Population of Methanogenic Bacteria and the Initiation of Methane Production upon Flooding of Paddy Soil. FEMS Microbiol. Ecol. 1990, 6, 103–111. [Google Scholar] [CrossRef]
- Fan, Y.; Hao, X.; Carswell, A.; Misselbrook, T.; Ding, R.; Li, S.; Kang, S. Inorganic Nitrogen Fertilizer and High N Application Rate Promote N2O Emission and Suppress CH4 Uptake in a Rotational Vegetable System. Soil Tillage Res. 2021, 206, 104848. [Google Scholar] [CrossRef]
- Sun, B.; Dong, Z.-X.; Zhang, X.-X.; Li, Y.; Cao, H.; Cui, Z.-L. Rice to Vegetables: Short- Versus Long-Term Impact of Land-Use Change on the Indigenous Soil Microbial Community. Microb. Ecol. 2011, 62, 474–485. [Google Scholar] [CrossRef] [PubMed]
- Stark, C.H.; Condron, L.M.; O’Callaghan, M.; Stewart, A.; Di, H.J. Differences in Soil Enzyme Activities, Microbial Community Structure and Short-Term Nitrogen Mineralisation Resulting from Farm Management History and Organic Matter Amendments. Soil Biol. Biochem. 2008, 40, 1352–1363. [Google Scholar] [CrossRef]
- Lüneberg, K.; Schneider, D.; Siebe, C.; Daniel, R. Drylands Soil Bacterial Community Is Affected by Land Use Change and Different Irrigation Practices in the Mezquital Valley, Mexico. Sci. Rep. 2018, 8, 1413. [Google Scholar] [CrossRef] [PubMed]
- Matysek, M.; Leake, J.; Banwart, S.; Johnson, I.; Page, S.; Kaduk, J.; Smalley, A.; Cumming, A.; Zona, D. Impact of Fertiliser, Water Table, and Warming on Celery Yield and CO2 and CH4 Emissions from Fenland Agricultural Peat. Sci. Total Environ. 2019, 667, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Li, C.; Li, Z.; Liu, S.; Zou, Y.; Gao, X.; Cai, Y.; Siddique, K.H.M.; Wu, P.; Zhao, X. Greenhouse Gas Emission Responses to Different Soil Amendments on the Loess Plateau, China. Agric. Ecosyst. Environ. 2023, 342, 108233. [Google Scholar] [CrossRef]
- Flessa, H.; Ruser, R.; Dörsch, P.; Kamp, T.; Jimenez, M.A.; Munch, J.C.; Beese, F. Integrated Evaluation of Greenhouse Gas Emissions (CO2, CH4, N2O) from Two Farming Systems in Southern Germany. Agric. Ecosyst. Environ. 2002, 91, 175–189. [Google Scholar] [CrossRef]
- Glatzel, S.; Stahr, K. Methane and Nitrous Oxide Exchange in Differently Fertilised Grassland in Southern Germany. Plant Soil 2001, 231, 21–35. [Google Scholar] [CrossRef]
- Iqbal, J.; Hu, R.; Feng, M.; Lin, S.; Malghani, S.; Ali, I.M. Microbial Biomass, and Dissolved Organic Carbon and Nitrogen Strongly Affect Soil Respiration in Different Land Uses: A Case Study at Three Gorges Reservoir Area, South China. Agric. Ecosyst. Environ. 2010, 137, 294–307. [Google Scholar] [CrossRef]
- Zhang, S.; Fang, Y.; Luo, Y.; Li, Y.; Ge, T.; Wang, Y.; Wang, H.; Yu, B.; Song, X.; Chen, J.; et al. Linking Soil Carbon Availability, Microbial Community Composition and Enzyme Activities to Organic Carbon Mineralization of a Bamboo Forest Soil Amended with Pyrogenic and Fresh Organic Matter. Sci. Total Environ. 2021, 801, 149717. [Google Scholar] [CrossRef]
- Tang, Y.; Yu, G.; Zhang, X.; Wang, Q.; Tian, J.; Niu, S.; Tian, D.; Ge, J. Different Strategies for Regulating Free-Living N2 Fixation in Nutrient-Amended Subtropical and Temperate Forest Soils. Appl. Soil Ecol. 2019, 136, 21–29. [Google Scholar] [CrossRef]
- Duff, A.M.; Forrestal, P.; Ikoyi, I.; Brennan, F. Assessing the Long-Term Impact of Urease and Nitrification Inhibitor Use on Microbial Community Composition, Diversity and Function in Grassland Soil. Soil Biol. Biochem. 2022, 170, 108709. [Google Scholar] [CrossRef]
- Kuypers, M.M.M.; Marchant, H.K.; Kartal, B. The Microbial Nitrogen-Cycling Network. Nat. Rev. Microbiol. 2018, 16, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zheng, M.; Song, W.; Wen, S.; Wang, B.; Zhu, C.; Shen, R. Impact of 25 Years of Inorganic Fertilization on Diazotrophic Abundance and Community Structure in an Acidic Soil in Southern China. Soil Biol. Biochem. 2017, 113, 240–249. [Google Scholar] [CrossRef]
- Ouyang, Y.; Evans, S.E.; Friesen, M.L.; Tiemann, L.K. Effect of Nitrogen Fertilization on the Abundance of Nitrogen Cycling Genes in Agricultural Soils: A Meta-Analysis of Field Studies. Soil Biol. Biochem. 2018, 127, 71–78. [Google Scholar] [CrossRef]
- Dynarski, K.A.; Houlton, B.Z. Nutrient Limitation of Terrestrial Free-Living Nitrogen Fixation. New Phytol. 2018, 217, 1050–1061. [Google Scholar] [CrossRef]
- Yuan, H.; Ge, T.; Chen, C.; O’Donnell, A.G.; Wu, J. Significant Role for Microbial Autotrophy in the Sequestration of Soil Carbon. Appl. Environ. Microbiol. 2012, 78, 2328–2336. [Google Scholar] [CrossRef]
- Van Hees, P.A.W.; Jones, D.L.; Finlay, R.; Godbold, D.L.; Lundström, U.S. The Carbon We Do Not See—The Impact of Low Molecular Weight Compounds on Carbon Dynamics and Respiration in Forest Soils: A Review. Soil Biol. Biochem. 2005, 37, 1–13. [Google Scholar] [CrossRef]
- Abdel-Rahman, M.A.; Nour El-Din, M.; Refaat, B.M.; Abdel-Shakour, E.H.; Ewais, E.E.-D.; Alrefaey, H.M.A. Biotechnological Application of Thermotolerant Cellulose-Decomposing Bacteria in Composting of Rice Straw. Ann. Agric. Sci. 2016, 61, 135–143. [Google Scholar] [CrossRef]
- Ahmad, B.; Nigar, S.; Malik, N.A.; Bashir, S.; Ali, J.; Yousaf, S.; Bangash, J.A.; Jan, I. Isolation and Characterization of Cellulolytic Nitrogen Fixing Azotobacter Species from Wheat Rhizosphere of Khyber Pakhtunkhwa. World Appl. Sci. J. 2013, 27, 51–60. [Google Scholar]
- Emtiazi, G.; Etemadifar, Z.; Tavassoli, M. A Novel Nitrogen-Fixing Cellulytic Bacterium Associated with Root of Corn Is a Candidate for Production of Single Cell Protein. Biomass Bioenergy 2003, 25, 423–426. [Google Scholar] [CrossRef]
- Harindintwali, J.D.; Zhou, J.; Yu, X. Lignocellulosic Crop Residue Composting by Cellulolytic Nitrogen-Fixing Bacteria: A Novel Tool for Environmental Sustainability. Sci. Total Environ. 2020, 715, 136912. [Google Scholar] [CrossRef]
- Miao, F.; Li, Y.; Cui, S.; Jagadamma, S.; Yang, G.; Zhang, Q. Soil Extracellular Enzyme Activities under Long-Term Fertilization Management in the Croplands of China: A Meta-Analysis. Nutr. Cycl. Agroecosystems 2019, 114, 125–138. [Google Scholar] [CrossRef]
- García-Orenes, F.; Morugán-Coronado, A.; Zornoza, R.; Scow, K. Changes in Soil Microbial Community Structure Influenced by Agricultural Management Practices in a Mediterranean Agro-Ecosystem. PLoS ONE 2013, 8, e80522. [Google Scholar] [CrossRef] [PubMed]
- Siles, J.A.; Vera, A.; Díaz-López, M.; García, C.; van den Hoogen, J.; Crowther, T.W.; Eisenhauer, N.; Guerra, C.; Jones, A.; Orgiazzi, A.; et al. Land-Use- and Climate-Mediated Variations in Soil Bacterial and Fungal Biomass across Europe and Their Driving Factors. Geoderma 2023, 434, 116474. [Google Scholar] [CrossRef]
- Yang, T.; Li, X.; Hu, B.; Wei, D.; Wang, Z.; Bao, W. Soil Microbial Biomass and Community Composition along a Latitudinal Gradient in the Arid Valleys of Southwest China. Geoderma 2022, 413, 115750. [Google Scholar] [CrossRef]
- Bastida, F.; Eldridge, D.J.; García, C.; Kenny Png, G.; Bardgett, R.D.; Delgado-Baquerizo, M. Soil Microbial Diversity–Biomass Relationships Are Driven by Soil Carbon Content across Global Biomes. ISME J. 2021, 15, 2081–2091. [Google Scholar] [CrossRef]
- Wang, H.; Liu, S.; Li, H.; Tao, X.; Wang, H.; Qi, J.; Zhang, Z. Large-Scale Homogenization of Soil Bacterial Communities in Response to Agricultural Practices in Paddy Fields, China. Soil Biol. Biochem. 2022, 164, 108490. [Google Scholar] [CrossRef]
- Di Salvo, L.P.; Cellucci, G.C.; Carlino, M.E.; García de Salamone, I.E. Plant Growth-Promoting Rhizobacteria Inoculation and Nitrogen Fertilization Increase Maize (Zea mays L.) Grain Yield and Modified Rhizosphere Microbial Communities. Appl. Soil Ecol. 2018, 126, 113–120. [Google Scholar] [CrossRef]
- Sabier Saeed, K.; Abdulla Ahmed, S.; Ahmaed Hassan, I.; Hamed Ahmed, P. Effect of Bio-Fertilizer and Chemical Fertilizer on Growth and Yield in Cucumber (Cucumis sativus) in Green House Condition. Pak. J. Biol. Sci. 2015, 18, 129–134. [Google Scholar] [CrossRef]
- Angelina, E.; Papatheodorou, E.M.; Demirtzoglou, T.; Monokrousos, N. Effects of Bacillus Subtilis and Pseudomonas Fluorescens Inoculation on Attributes of the Lettuce (Lactuca sativa L.) Soil Rhizosphere Microbial Community: The Role of the Management System. Agronomy 2020, 10, 1428. [Google Scholar] [CrossRef]
- Qiu, Z.; Egidi, E.; Liu, H.; Kaur, S.; Singh, B.K. New Frontiers in Agriculture Productivity: Optimised Microbial Inoculants and in Situ Microbiome Engineering. Biotechnol. Adv. 2019, 37, 107371. [Google Scholar] [CrossRef] [PubMed]
- Fiorentino, N.; Ventorino, V.; Woo, S.L.; Pepe, O.; De Rosa, A.; Gioia, L.; Romano, I.; Lombardi, N.; Napolitano, M.; Colla, G.; et al. Trichoderma-Based Biostimulants Modulate Rhizosphere Microbial Populations and Improve N Uptake Efficiency, Yield, and Nutritional Quality of Leafy Vegetables. Front. Plant Sci. 2018, 9, 743. [Google Scholar] [CrossRef] [PubMed]
- Siebers, M.; Rohr, T.; Ventura, M.; Schütz, V.; Thies, S.; Kovacic, F.; Jaeger, K.-E.; Berg, M.; Dörmann, P.; Schulz, M. Disruption of Microbial Community Composition and Identification of Plant Growth Promoting Microorganisms after Exposure of Soil to Rapeseed-Derived Glucosinolates. PLoS ONE 2018, 13, e0200160. [Google Scholar] [CrossRef]
- Ju, W.; Liu, L.; Fang, L.; Cui, Y.; Duan, C.; Wu, H. Impact of Co-Inoculation with Plant-Growth-Promoting Rhizobacteria and Rhizobium on the Biochemical Responses of Alfalfa-Soil System in Copper Contaminated Soil. Ecotoxicol. Environ. Saf. 2019, 167, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Hidri, R.; Metoui-Ben Mahmoud, O.; Debez, A.; Abdelly, C.; Barea, J.-M.; Azcon, R. Modulation of C:N:P Stoichiometry Is Involved in the Effectiveness of a PGPR and AM Fungus in Increasing Salt Stress Tolerance of Sulla Carnosa Tunisian Provenances. Appl. Soil Ecol. 2019, 143, 161–172. [Google Scholar] [CrossRef]
- Egamberdiyeva, D. The Effect of Plant Growth Promoting Bacteria on Growth and Nutrient Uptake of Maize in Two Different Soils. Appl. Soil Ecol. 2007, 36, 184–189. [Google Scholar] [CrossRef]
- Mourão, I.; Brito, M. Effects of Direct Film Crop Cover and Top Dress Nitrogen on Earliness and Yield of Broccoli Crop (Brassica oleracea Var. Italica Plenk). In Proceedings of the International Conference on Environmental Problems Associated with Nitrogen Fertilisation of Field Grown Vegetable Crops 563, Postdam, Germany, 30 August–1 September 1999; pp. 103–109. [Google Scholar]
- Nuzzo, A.; Satpute, A.; Albrecht, U.; Strauss, S.L. Impact of Soil Microbial Amendments on Tomato Rhizosphere Microbiome and Plant Growth in Field Soil. Microb. Ecol. 2020, 80, 398–409. [Google Scholar] [CrossRef]
- Schenck zu Schweinsberg-Mickan, M.; Müller, T. Impact of Effective Microorganisms and Other Biofertilizers on Soil Microbial Characteristics, Organic-Matter Decomposition, and Plant Growth. J. Plant Nutr. Soil Sci. 2009, 172, 704–712. [Google Scholar] [CrossRef]
- Martínez-Viveros, O.; Jorquera, M.A.; Crowley, D.E.; Gajardo, G.; Mora, M.L. Mechanisms and practical considerations involved in plant growth promotion by rhizobacteria. J. Soil Sci. Plant Nutr. 2010, 10, 293–319. [Google Scholar] [CrossRef]
- Kim, S.; Kim, S.; Kiniry, J.R.; Ku, K.-M. A Hybrid Decision Tool for Optimizing Broccoli Production in a Changing Climate. Hortic. Environ. Biotechnol. 2021, 62, 299–312. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Cumulative CO2 | Cumulative N2O | Cumulative CH4 | CO2e | Crop Yield | CO2e |
|---|---|---|---|---|---|---|
| g m−2 | kg ha−1 | g kg−1 of Crop Yield | ||||
| BA + FU | 261.5 ± 19.7 | 0.0 ± 0.0 | 3.0 ± 0.3 | 351.3 ± 14.4 | 15,082.3 ± 894.3 | 225.0 ± 12.6 |
| BA | 242.8 ± 24.4 | 0.1 ± 0.1 | 2.1 ± 0.4 | 327.7 ± 36.7 | 14,928.3 ± 886.4 | 203.3 ± 18.4 |
| F50 | 233.8 ± 16.7 | 0.1 ± 0.1 | 3.0 ± 0.7 | 360.0 ± 43.6 | 15,622.7 ± 1736.2 | 262.3 ± 27.6 |
| F100 | 250.3 ± 8.1 | 0.0 ± 0.0 | 3.1 ± 0.4 | 347.2 ± 22.3 | 16,797.3 ± 650.3 | 204.3 ± 24.9 |
| F-ANOVA | 0.744 ns | 0.336 ns | 0.518 ns | 0.903 ns | 0.649 ns | 0.260 ns |
| BA + FU | BA | F50 | F100 | F-ANOVA | ||
|---|---|---|---|---|---|---|
| Cls | μmol gearbox sugars g−1 h−1 | 36.3 ± 6.0 | 20.3 ± 11.1 | 30.1 ± 10.3 | 31.0 ± 3.45 | 0.649 ns |
| Urs | μmol NH4+ g−1 h−1 | 4.9 ± 1.4 | 4.7 ± 0.8 | 4.1 ± 0.7 | 7.2 ± 2.1 | 1.049 ns |
| Aryl | μmol PNP g−1 h−1 | 13.3 ± 1.8 | 15.4 ± 1.9 | 14.4 ± 1.2 | 15.6 ± 0.9 | 0.490 ns |
| Glu | μmol PNP g−1 h−1 | 0.3 ± 0.0 | 0.3 ± 0.0 | 0.3 ± 0.1 | 0.3 ± 0.0 | 0.505 ns |
| M | % | 14.4 ± 0.6 | 15.5 ± 0.9 | 15.4 ± 0.8 | 13.6 ± 0.5 | 0.254 ns |
| SOC | % | 1.0 ± 0.0 | 1.0 ± 0.1 | 1.1 ± 0.0 | 1.0 ± 0.1 | 0.656 ns |
| Nt | % | 0.1 ± 0.0 | 0.1 ± 0.0 | 0.1 ± 0.0 | 0.1 ± 0.0 | 0.411 ns |
| Csol | mg kg−1 | 1405.8 ± 71.4 | 1255.5 ± 136.4 | 1080.5 ± 52.3 | 1129.5 ± 56.6 | 0.082 ns |
| Nsol | mg kg−1 | 94.6 ± 8.3 | 78.2 ± 7.2 | 98.1 ± 16.8 | 95.0 ± 4.4 | 0.534 ns |
| NO3− | mg kg−1 | 47.2 ± 13.6 | 33.2 ± 5.5 | 40.4 ± 6.1 | 79.8 ± 19.0 | 0.088 ns |
| NH4+ | mg kg−1 | 1.7 ± 0.0 | 1.8 ± 0.1 | 1.7 ± 0.0 | 1.8 ± 0.0 | 0.293 ns |
| Microbial Groups | BA + FU | BA | F50 | F100 | F-ANOVA |
|---|---|---|---|---|---|
| nmol g−1 | |||||
| Total PLFA | 15.6 ± 1.0 | 15.8 ± 1.1 | 15.6± 1.1 | 14.6 ± 0.7 | 0.834 ns |
| Firmicutes | 3.6 ± 0.2 | 3.5 ± 0.3 | 3.4 ± 0.3 | 3.1 ± 0.2 | 0.523 ns |
| Actinobacteria | 1.8 ± 0.1 | 1.7 ± 0.1 | 1.4 ± 0.1 | 1.5 ± 0.1 | 0.097 ns |
| Gram-positive | 5.4 ± 0.3 | 5.2 ± 0.4 | 4.7 ± 0.2 | 4.6 ± 0.3 | 0.265 ns |
| Gram-negative | 2.5 ± 0.2 | 2.7 ± 0.2 | 2.9 ± 0.2 | 2.6 ± 0.2 | 0.729 ns |
| Bacteria | 7.9 ± 0.5 | 7.8 ± 0.6 | 7.6 ± 0.4 | 7.2 ± 0.5 | 0.763 ns |
| AMF | bdl | bdl | bdl | bdl | |
| Zygomycota | 2.0 ± 0.2 | 2.1 ± 0.1 | 2.2 ± 0.2 | 1.9 ± 0.1 | 0.558 ns |
| Ascomycota and Basidiomycota | 0.5 ± 0.0 | 0.6 ± 0.0 | 0.6 ± 0.0 | 0.6 ± 0.0 | 0.194 ns |
| Total fungi | 2.6 ± 0.2 | 2.7 ± 0.1 | 2.8 ± 0.3 | 2.5 ± 0.1 | 0.530 ns |
| Unspecific microbial PLFA | 5.1 ± 0.4 | 5.3 ± 0.4 | 5.1 ± 0.4 | 4.9 ± 0.2 | 0.922 ns |
| Y | X | m | Partial Correlation | β | R2 | R2 adj | F Value |
|---|---|---|---|---|---|---|---|
| Cumulative CO2 (mg m−2) | Constant | 422.86 | 0.70 | 0.63 | 10.34 (p < 0.01) | ||
| nifH (Log copies g−1 dry weight) | −57.51 | −0.70 | −0.56 | ||||
| Crop yield (kg ha−1) | 0.01 | 0.68 | 0.52 |
| Variance Explained | PC1 (33.98%) | PC2 (11.50%) | PC3 (10.01%) | PC4 (8.70%) |
|---|---|---|---|---|
| Total PLFA | 0.98 | 0.06 | 0.01 | −0.01 |
| Bacteria | 0.95 | 0.02 | 0.15 | 0.03 |
| Unspecific microbial PLFA | 0.95 | 0.16 | −0.08 | 0.05 |
| Firmicutes | 0.91 | −0.04 | 0.05 | −0.05 |
| Zygomycota | 0.90 | 0.01 | −0.20 | −0.23 |
| Gram-negative | 0.90 | 0.21 | 0.15 | 0.07 |
| Fungi | 0.88 | −0.01 | −0.19 | −0.22 |
| Gram-positive | 0.85 | −0.10 | 0.13 | 0.00 |
| Soil organic C | 0.78 | 0.04 | 0.41 | 0.16 |
| amoA gene | 0.69 | 0.11 | −0.12 | 0.60 |
| β-glucosidase | 0.68 | −0.05 | 0.05 | 0.34 |
| Soluble C | −0.57 | 0.01 | −0.21 | −0.22 |
| CO2e | 0.16 | 0.89 | 0.02 | −0.17 |
| N2O | 0.27 | 0.73 | 0.07 | 0.05 |
| nirK gene | 0.04 | −0.71 | 0.38 | 0.00 |
| CH4 | 0.26 | 0.59 | 0.25 | −0.08 |
| GH7 gene | 0.24 | 0.08 | 0.89 | −0.15 |
| cbbL gene | 0.29 | −0.24 | 0.61 | 0.15 |
| NO3− | 0.07 | −0.12 | 0.60 | 0.06 |
| Urease | −0.42 | 0.44 | 0.50 | 0.15 |
| NH4+ | 0.05 | −0.08 | −0.12 | 0.89 |
| nifH gene | 0.45 | −0.18 | 0.21 | 0.68 |
| Actinobacteria | 0.43 | −0.15 | 0.23 | 0.09 |
| Ascomycota and Basidiomycota | 0.41 | −0.11 | −0.14 | −0.16 |
| Moisture | 0.35 | 0.09 | −0.01 | 0.11 |
| Total N | 0.50 | 0.04 | 0.38 | 0.33 |
| Soluble N | 0.02 | 0.12 | −0.18 | −0.07 |
| CO2 | −0.19 | 0.39 | −0.23 | −0.25 |
| Cellulase | 0.23 | −0.03 | −0.18 | −0.54 |
| Arylesterase | 0.02 | 0.18 | 0.05 | 0.08 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ollio, I.; Santás-Miguel, V.; Gómez, D.S.; Lloret, E.; Sánchez-Navarro, V.; Martínez-Martínez, S.; Egea-Gilabert, C.; Fernández, J.A.; Calviño, D.F.; Zornoza, R. Effect of Biofertilizers on Broccoli Yield and Soil Quality Indicators. Horticulturae 2024, 10, 42. https://doi.org/10.3390/horticulturae10010042
Ollio I, Santás-Miguel V, Gómez DS, Lloret E, Sánchez-Navarro V, Martínez-Martínez S, Egea-Gilabert C, Fernández JA, Calviño DF, Zornoza R. Effect of Biofertilizers on Broccoli Yield and Soil Quality Indicators. Horticulturae. 2024; 10(1):42. https://doi.org/10.3390/horticulturae10010042
Chicago/Turabian StyleOllio, Irene, Vanesa Santás-Miguel, Diego Soto Gómez, Eva Lloret, Virginia Sánchez-Navarro, Silvia Martínez-Martínez, Catalina Egea-Gilabert, Juan A. Fernández, David Fernández Calviño, and Raúl Zornoza. 2024. "Effect of Biofertilizers on Broccoli Yield and Soil Quality Indicators" Horticulturae 10, no. 1: 42. https://doi.org/10.3390/horticulturae10010042