
Yeast-Mediated Biomass Valorization for Biofuel Production: A Literature Review

 , , , , and
, , , , and
Abstract
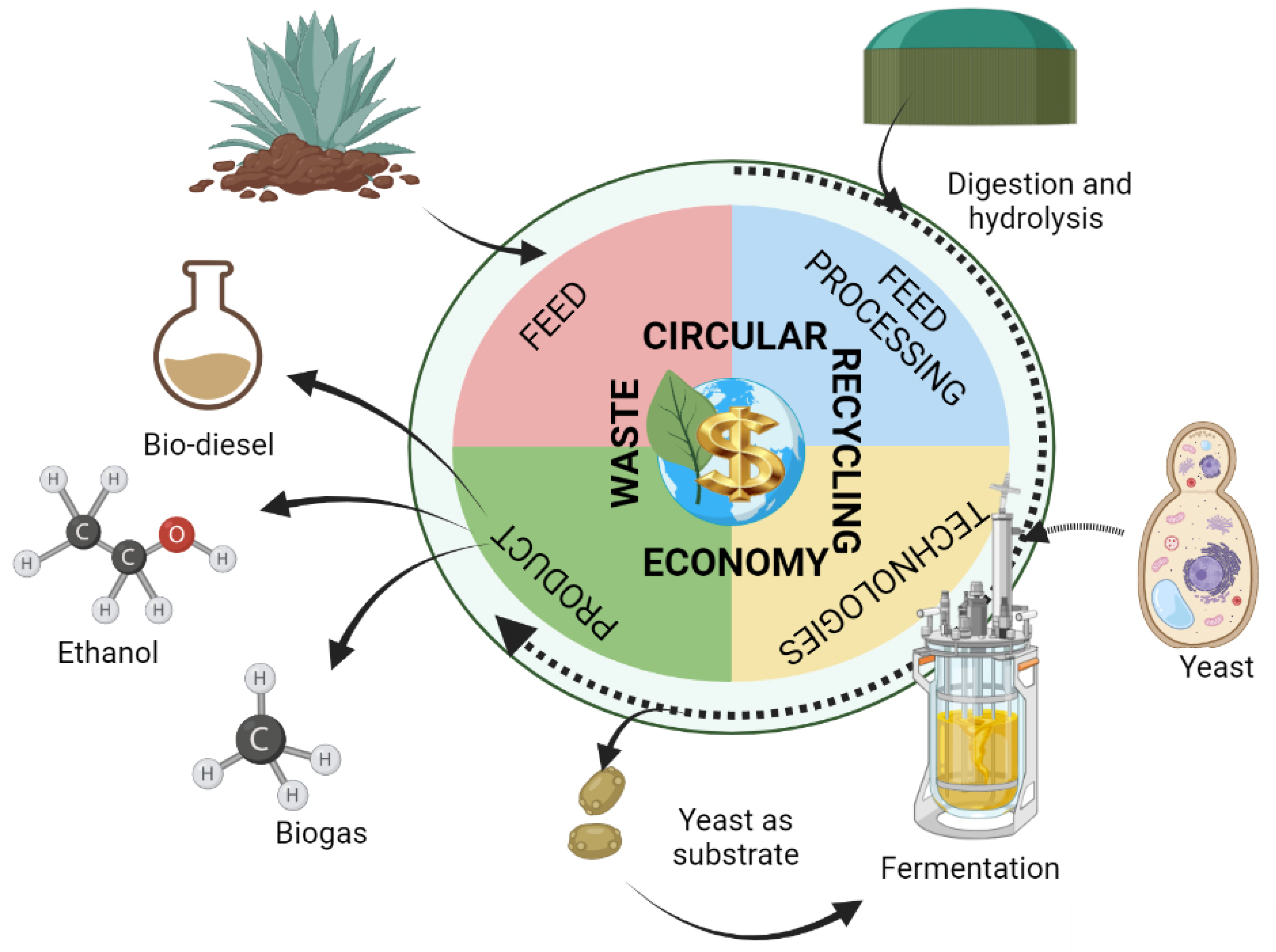
:1. Introduction
2. The Application of Yeast for Bioethanol Production
2.1. The Selection of Yeast Strains
2.2. Yeasts from Extreme Environments
2.3. Simultaneous Saccharification and Fermentation (SSF)
2.4. Consolidated Bioprocessing (CBP)
2.5. Metabolic and Evolutionary Engineering
3. The Application of Yeast for Biodiesel Production
3.1. The Selection of Yeast Strains
3.2. Cultivation of Yeast for Lipid Production and Transesterification
3.2.1. Submerged Fermentation
3.2.2. Solid-State Fermentation
3.2.3. Two-Stage Fermentation
3.2.4. Co-Culture System
3.3. Genetic Engineering of Yeast Strains
3.4. Characteristics of the Biodiesel Produced from Yeast Lipids
3.4.1. Cetane Number
3.4.2. Density
3.4.3. Viscosity
3.4.4. Chemical Composition
3.4.5. Calorific Value
4. Application of Yeast for Biogas and Bio-Methane Production
4.1. Biogas Production from Waste Biomass
4.2. Yeast Biomass as Feed and Biocatalysts
5. Challenges for Yeast-Mediated Biofuel Production from Waste Feedstock
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Y.; Cruz-Morales, P.; Zargar, A.; Belcher, M.S.; Pang, B.; Englund, E.; Dan, Q.; Yin, K.; Keasling, J.D. Biofuels for a sustainable future. Cell 2021, 184, 1636–1647. [Google Scholar] [CrossRef]
- Obobisa, E.S. Achieving 1.5 °C and net-zero emissions target: The role of renewable energy and financial development. Renew. Energy 2022, 188, 967–985. [Google Scholar] [CrossRef]
- Baptista, S.L.; Romaní, A.; Cunha, J.T.; Domingues, L. Multi-feedstock biorefinery concept: Valorization of winery wastes by engineered yeast. J. Environ. Manag. 2023, 326 Pt A, 116623. [Google Scholar] [CrossRef]
- Velvizhi, G.; Balakumar, K.; Shetti, N.P.; Ahmad, E.; Kishore, P.K.; Aminabhavi, T.M. Integrated biorefinery processes for conversion of lignocellulosic biomass to value added materials: Paving a path towards circular economy. Bioresour. Technol. 2022, 343, 126151. [Google Scholar] [CrossRef] [PubMed]
- Aditiya, H.B.; Mahlia, T.M.I.; Chong, W.T.; Nur, H.; Sebayang, A.H. Second generation bioethanol production: A critical review. Renew. Sustain. Energy Rev. 2016, 66, 631–653. [Google Scholar] [CrossRef]
- Eliodório, K.P.; Cunha, G.C.G.E.; Müller, C.; Lucaroni, A.C.; Giudici, R.; Walker, G.M.; Alves, S.L., Jr.; Basso, T.O. Advances in yeast alcoholic fermentations for the production of bioethanol, beer and wine. Adv. Appl. Microbiol. 2019, 109, 61–119. [Google Scholar] [CrossRef] [PubMed]
- The Génolevures Consortium; Souciet, J.L.; Dujon, B.; Gaillardin, C.; Johnston, M.; Baret, P.V.; Cliften, P.; Sherman, D.J.; Weissenbach, J.; Westhof, E.; et al. Comparative genomics of protoploid Saccharomycetaceae. Genome Res. 2009, 19, 1696–1709. [Google Scholar]
- Albergaria, H.; Arneborg, N. Dominance of Saccharomyces cerevisiae in alcoholic fermentation processes: Role of physiological fitness and microbial interactions. Appl. Microbiol. Biotechnol. 2016, 100, 2035–2046. [Google Scholar] [CrossRef]
- Nandal, P.; Sharma, S.; Arora, A. Bioprospecting non-conventional yeasts for ethanol production from rice straw hydrolysate and their inhibitor tolerance. Renew. Energy 2020, 147, 1694–1703. [Google Scholar] [CrossRef]
- Sharma, S.; Arora, A.; Sharma, P.; Singh, S.; Nain, L.; Paul, D. Notable mixed substrate fermentation by native Kodamaea ohmeri strains isolated from Lagenaria siceraria flowers and ethanol production on paddy straw hydrolysates. Chem. Cent. J. 2018, 12, 8. [Google Scholar] [CrossRef]
- Hawaz, E.; Tafesse, M.; Tesfaye, A.; Beyene, D.; Kiros, S.; Kebede, G.; Boekhout, T.; Theelen, B.; Groenewald, M.; Degefe, A.; et al. Isolation and characterization of bioethanol producing wild yeasts from bio-wastes and co-products of sugar factories. Ann. Microbiol. 2022, 72, 39. [Google Scholar] [CrossRef]
- Samar, W.; Arora, A.; Sharma, A.; Sharma, S.; Nandal, P. Material flow of cellulose in rice straw to ethanol and lignin recovery by NaOH pretreatment coupled with acid washing. Biomass Conv. Bioref 2021, 13, 2233–2242. [Google Scholar] [CrossRef]
- Saeed, Z.; Palamae, S.; Intharapat, P.; Khundamri, N.; Tanrattanakul, V.; Tirawanichakul, Y.; Suttinun, O. Enhanced ethanol production from lignocellulosic hydrolysate using Meyerozyma caribbica biofilm immobilized on modified epoxy foam. Biomass Bioenergy 2021, 154, 106267. [Google Scholar] [CrossRef]
- Nwaefuna, A.E.; Rumbold, K.; Boekhout, T.; Zhou, N. Bioethanolic yeasts from dung beetles: Tapping the potential of extremophilic yeasts for improvement of lignocellulolytic feedstock fermentation. Biotechnol. Biofuels 2021, 14, 86. [Google Scholar] [CrossRef]
- Talukder, A.A.; Easmin, F.; Mahmud, S.A.; Yamada, M. Thermotolerant yeasts capable of producing bioethanol: Isolation from natural fermented sources, identification and characterization. Biotechnol. Biotechnol. Equip. 2016, 30, 1106–1114. [Google Scholar] [CrossRef]
- Nguyen, P.V.; Nguyen, K.H.V.; Nguyen, N.L.; Ho, X.T.T.; Truong, P.H.; Thi Nguyen, K.C. Lychee-Derived, Thermotolerant Yeasts for Second-Generation Bioethanol Production. Ferment 2022, 8, 515. [Google Scholar] [CrossRef]
- Antti, P. Simultaneous ethanol fermentation and recovery for packaging waste fibre. Preprint 2017. [Google Scholar] [CrossRef]
- Van Zyl, W.H.; Lynd, L.R.; den Haan, R.; McBride, J.E. Consolidated bioprocessing for bioethanol production using Saccharomyces cerevisiae. Adv. Biochem. Eng. Biotechnol. 2007, 108, 205–235. [Google Scholar]
- Olguin-Maciel, E.; Singh, A.; Chable-Villacis, R.; Tapia-Tussell, R.; Ruiz, H.A. Consolidated Bioprocessing, an Innovative Strategy towards Sustainability for Biofuels Production from Crop Residues: An Overview. Agronomy 2020, 10, 1834. [Google Scholar] [CrossRef]
- Singhania, R.R.; Patel, A.K.; Singh, A.; Haldar, D.; Soam, S.; Chen, C.W.; Tsai, M.L.; Dong, C.D. Consolidated bioprocessing of lignocellulosic biomass: Technological advances and challenges. Bioresour. Technol. 2022, 354, 127153. [Google Scholar] [CrossRef]
- Kiesenhofer, D.P.; Mach-Aigner, A.R.; Mach, R.L. Fungal Metabolic Engineering for Biofuel Production, Mycology: Current and Future Developments Fungal Biotechnology for Biofuel Production. Fungal Biotechnol. Biofuel Prod. Fungal Biotechnol. Biofuel Prod. 2015, 1, 128. [Google Scholar] [CrossRef]
- Mahmoud, Y.A.G.; Abd El-Zaher, E. Recent advancements in biofuels production with a special attention to fungi. In Applied Biotechnology Reviews, Sustainable Biofuels; Ray, R.C., Ed.; Academic Press: Cambridge, MA, USA, 2021; pp. 73–99. ISBN 9780128202975. [Google Scholar]
- Cunha, J.T.; Romaní, A.; Inokuma, K.; Johansson, B.; Hasunuma, T.; Domingues, L. Consolidated bioprocessing of corn cob-derived hemicellulose: Engineered industrial Saccharomyces cerevisiae as efficient whole cell biocatalysts. Biotechnol. Biofuels 2020, 13, 138. [Google Scholar] [CrossRef] [PubMed]
- Maleki, F.; Changizian, M.; Zolfaghari, N.; Rajaei, S.; Noghabi, K.A.; Zahiri, H.S. Consolidated bioprocessing for bioethanol production by metabolically engineered Bacillus subtilis strains. Sci. Rep. 2021, 11, 13731. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sun, T.; Wu, T.; Li, J.; Hu, D.; Liu, D.; Li, J.; Tian, C. Consolidated bioprocessing for bioethanol production by metabolically engineered cellulolytic fungus Myceliophthora thermophila. Metab. Eng. 2023, 78, 192–199. [Google Scholar] [CrossRef]
- Singh, N.; Mathur, A.S.; Tuli, D.K.; Gupta, R.P.; Barrow, C.J.; Puri, M. Cellulosic ethanol production via consolidated bioprocessing by a novel thermophilic anaerobic bacterium isolated from a Himalayan hot spring. Biotechnol. Biofuels 2023, 10, 73. [Google Scholar] [CrossRef]
- Chung, D.; Cha, M.; Guss, A.M.; Westpheling, J. Direct conversion of plant biomass to ethanol by engineered Caldicellulosiruptor Bescii. Proc. Natl. Acad. Sci. USA 2014, 111, 8931–8936. [Google Scholar] [CrossRef]
- Stephanopoulos, G. Challenges in Engineering Microbes for Biofuels Production. Science 2007, 315, 801–804. [Google Scholar] [CrossRef] [PubMed]
- Alper, H.; Stephanopoulos, G. Engineering for biofuels: Exploiting innate microbial capacity or importing biosynthetic potential? Nat. Rev. Microbiol. 2009, 7, 715–723. [Google Scholar] [CrossRef]
- Madhavan, A.; Jose, A.A.; Binod, P.; Sindhu, R.; Sukumaran, R.K.; Pandey, A.; Castro, G.E. Synthetic Biology and Metabolic Engineering Approaches and Its Impact on Non-Conventional Yeast and Biofuel Production. Front. Energy Res. 2017, 5, 8. [Google Scholar] [CrossRef]
- Baptista, S.L.; Costa, C.E.; Cunha, J.T.; Soares, P.O.; Domingues, L. Metabolic engineering of Saccharomyces cerevisiae for the production of top value chemicals from biorefinery carbohydrates. Biotechnol. Adv. 2021, 47, 107697. [Google Scholar] [CrossRef]
- Sharma, S.; Arora, A. Tracking strategic developments for conferring xylose utilization/fermentation by Saccharomyces cerevisiae. Ann. Microbiol. 2020, 70, 50. [Google Scholar] [CrossRef]
- Zhang, K.; Lu, X.; Li, Y.; Jiang, X.; Liu, L.; Wang, H. New technologies provide more metabolic engineering strategies for bioethanol production in Zymomonas mobilis. Appl. Microbiol. Biotechnol. 2019, 103, 2087–2099. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Delneri, D.; O’Keefe, R.T. Non-coding RNAs in Saccharomyces cerevisiae: What Is the Function? Biochem. Soc. Trans. 2012, 40, 907–911. [Google Scholar] [CrossRef] [PubMed]
- Robak, K.; Balcerek, M. Current state-of-the-art in ethanol production from lignocellulosic feedstocks. Microbiol. Res. 2020, 240, 126534. [Google Scholar] [CrossRef]
- Khaleghi, M.K.; Savizi, I.S.P.; Lewis, N.E.; Shojaosadati, S.A. Synergisms of Machine Learning and Constraint-Based Modeling of Metabolism for Analysis and Optimization of Fermentation Parameters. Biotechnol. J. 2021, 16, e2100212. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.F.; Reardon, K.F. Quorum-Sensing Molecules Increase Ethanol Yield from Saccharomyces cerevisiae. FEMS Yeast Res. 2021, 21, foab056. [Google Scholar] [CrossRef]
- Selim, K.A.; El-Ghwas, D.E.; Easa, S.M.; Hassan, M.I.A. Bioethanol a Microbial Biofuel Metabolite; New Insights of Yeasts Metabolic Engineering. Fermentation 2018, 4, 16. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, P.; Wu, T.; Ruan, H. Bioethanol Production Based on Saccharomyces cerevisiae: Opportunities and Challenges. Fermentation 2023, 9, 709. [Google Scholar] [CrossRef]
- Mota, M.N.; Múgica, P.; Sá-Correia, I. Exploring Yeast Diversity to Produce Lipid-Based Biofuels from Agro-Forestry and Industrial Organic Residues. J. Fungi 2022, 8, 687. [Google Scholar] [CrossRef]
- Mhlongo, S.I.; Ezeokoli, O.T.; Roopnarain, A.; Ndaba, B.; Sekoai, P.T.; Habimana, O.; Pohl, C.H. The potential of single-cell oils derived from filamentous fungi as alternative feedstock sources for biodiesel production. Front. Microbiol. 2021, 12, 637381. [Google Scholar] [CrossRef]
- Singh, S.; Pandey, D.; Saravanabhupathy, S.; Daverey, A.; Dutta, K.; Arunachalam, K. Liquid wastes as a renewable feedstock for yeast biodiesel production: Opportunities and challenges. Environ. Res. 2022, 207, 112100. [Google Scholar] [CrossRef]
- Singh, G.; Jeyaseelan, C.; Bandyopadhyay, K.K.; Paul, D. Comparative analysis of biodiesel produced by acidic transesterification of lipid extracted from oleaginous yeast Rhodosporidium toruloides. 3 Biotech 2018, 8, 434. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Loh, K.C.; Kuroki, A.; Dai, Y.; Tong, Y.W. Microbial biodiesel production from industrial organic wastes by oleaginous microorganisms: Current status and prospects. J. Hazard. Mater. 2021, 402, 123543. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Sinha, S.; Kumar, K.K.; Gaur, N.A.; Bandyopadhyay, K.K.; Paul, D. High density cultivation of oleaginous yeast isolates in ‘mandi’ waste for enhanced lipid production using sugarcane molasses as feed. Fuel 2020, 276, 118073. [Google Scholar] [CrossRef]
- Shaigani, P.; Awad, D.; Redai, V.; Fuchs, M.; Haack, M.; Mehlmer, N.; Brueck, T. Oleaginous yeasts- substrate preference and lipid productivity: A view on the performance of microbial lipid producers. Microb. Cell Fact. 2021, 20, 220. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, S.; Aggelis, G. Lipids of oleaginous yeasts. Part II: Technology and potential applications. Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073. [Google Scholar] [CrossRef]
- Martinez-Silveira, A.; Garmendia, G.; Rufo, C.; Vero, S. Production of microbial oils by the oleaginous yeast Rhodotorula graminis S1/2R in a medium based on agro-industrial by-products. World J. Microbiol. Biotechnol. 2022, 38, 46. [Google Scholar] [CrossRef]
- Kot, A.M.; Błażejak, S.; Kurcz, A.; Gientka, I.; Kieliszek, M. Rhodotorula glutinis— potential source of lipids, carotenoids, and enzymes for use in industries. Appl. Microbiol. Biotechnol. 2016, 100, 6103–6117. [Google Scholar] [CrossRef]
- Mukhtar, H.; Suliman, S.M.; Shabbir, A.; Mumtaz, M.W.; Rashid, U.; Rahimuddin, S.A. Evaluating the Potential of Oleaginous Yeasts as Feedstock for Biodiesel Production. Protein Pept. Lett. 2018, 25, 195–201. [Google Scholar] [CrossRef]
- Beopoulos, A.; Cescut, J.; Haddouche, R.; Uribelarrea, J.L.; Molina-Jouve, C.; Nicaud, J.M. Yarrowia lipolytica as a model for bio-oil production. Prog. Lipid Res. 2009, 48, 375–387. [Google Scholar] [CrossRef]
- Fakas, S. Lipid biosynthesis in yeasts: A comparison of the lipid biosynthetic pathway between the model nonoleaginous yeast Saccharomyces cerevisiae and the model oleaginous yeast Yarrowia lipolytica. Eng. Life Sci. 2017, 17, 292–302. [Google Scholar] [CrossRef]
- He, Q.; Yang, Y.; Yang, S.; Donohoe, B.S.; Wychen, S.V.; Zhang, M.; Himmel, M.E.; Knoshaug, E.P. Oleaginicity of the yeast strain Saccharomyces cerevisiae D5A. Biotechnol. Biofuels 2018, 11, 258. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Sinha, S.; Bandyopadhyay, K.K.; Lawrence, M.; Paul, D. Triauxic growth of an oleaginous red yeast Rhodosporidium toruloides on waste ‘extract’for enhanced and concomitant lipid and β-carotene production. Microb. Cell Factories 2018, 17, 182. [Google Scholar] [CrossRef]
- Kocharin, K.; Siewers, V.; Nielsen, J. Improved polyhydroxybutyrate production by Saccharomyces cerevisiae through the use of the phosphoketolase pathway. Biotechnol. Bioeng. 2013, 110, 2216–2224. [Google Scholar] [CrossRef] [PubMed]
- Ratledge, C. The role of malic enzyme as the provider of NADPH in oleaginous microorganisms: A reappraisal and unsolved problems. Biotechnol. Lett. 2014, 36, 1557–1568. [Google Scholar] [CrossRef] [PubMed]
- Qi, F.; Zhao, X.; Kitahara, Y.; Li, T.; Ou, X.; Du, W.; Liu, D.; Huang, J. Integrative transcriptomic and proteomic analysis of the mutant lignocellulosic hydrolyzate-tolerant Rhodosporidium toruloides. Eng. Life Sci. 2016, 17, 249–261. [Google Scholar] [CrossRef]
- Ratledge, C.; Cohen, Z. Microbial and algal oils: Do they have a future for biodiesel or as commodity oils ? Lipid Technol. 2008, 20, 155–161. [Google Scholar] [CrossRef]
- Qian, X.; Gorte, O.; Chen, L.; Zhang, W.; Dong, W.; Ma, J.; Jiang, M.; Xin, F.; Ochsenreither, K. Co-production of single cell oil and gluconic acid using oleaginous Cryptococcus podzolicus DSM 27192. Biotechnol. Biofuels 2019, 12, 127. [Google Scholar] [CrossRef]
- Lizardi-Jiménez, M.A.; Hernández-Martínez, R. Solid state fermentation (SSF): Diversity of applications to valorize waste and biomass. 3 Biotech 2017, 7, 44. [Google Scholar] [CrossRef]
- Meng, X.; Yang, J.; Xu, X.; Zhang, L.; Nie, Q.; Xian, M. Biodiesel production from oleaginous microorganisms. Renew. Energy 2009, 34, 1–5. [Google Scholar] [CrossRef]
- Patel, A.; Arora, N.; Sartaj, K.; Pruthi, V.; Pruthi, P.A. Sustainable biodiesel production from oleaginous yeasts utilizing hydrolysates of various non-edible lignocellulosic biomasses. Renew. Sustain. Energy Rev. 2016, 62, 836–855. [Google Scholar] [CrossRef]
- Gientka, I.; Wirkowska-Wojdyła, M.; Ostrowska-Ligęza, E.; Janowicz, M.; Reczek, L.; Synowiec, A.; Błażejak, S. Enhancing Red Yeast Biomass Yield and Lipid Biosynthesis by Using Waste Nitrogen Source by Glucose Fed-Batch at Low Temperature. Microorganisms 2022, 10, 1253. [Google Scholar] [CrossRef] [PubMed]
- Karim, A.; Islam, M.A.; Mishra, P.; Muzahid, A.J.M.; Yousuf, A.; Khan, M.R.; Faizal, C.K.M. Yeast and bacteria co-culture-based lipid production through bioremediation of palm oil mill effluent: A statistical optimization. Biomass Conv. Bioref 2023, 13, 2947–2958. [Google Scholar] [CrossRef]
- Hu, Y.; Zhu, Z.; Nielsen, J.; Siewers, V. Engineering Saccharomyces cerevisiae cells for production of fatty acid-derived biofuels and chemicals. Open Biol. 2019, 9, 190049. [Google Scholar] [CrossRef]
- Yu, T.; Zhou, Y.J.; Huang, M.; Liu, Q.; Pereira, R.; David, F.; Nielsen, J. Reprogramming yeast metabolism from alcoholic fermentation to lipogenesis. Cell 2018, 174, 1549–1558. [Google Scholar] [CrossRef]
- van Dijken, J.P.; Scheffers, W.A. Redox balances in the metabolism of sugars by yeasts. FEMS Microbiol. Rev. 1986, 1, 199–224. [Google Scholar] [CrossRef]
- Mitrea, L.; Ranga, F.; Fetea, F.; Dulf, F.V.; Rusu, A.; Trif, M.; Vodnar, D.C. Biodiesel-derived glycerol obtained from renewable biomass-a suitable substrate for the growth of Candida zeylanoides yeast strain ATCC 20367. Microorganisms 2019, 7, 265. [Google Scholar] [CrossRef]
- Lacerda, M.P.; Oh, E.J.; Eckert, C. The model system Saccharomyces cerevisiae versus emerging non-model yeasts for the production of biofuels. Life 2020, 10, 299. [Google Scholar] [CrossRef]
- Sitepu, I.R.; Garay, L.A.; Sestric, R.; Levin, D.; Block, D.E.; German, J.B.; Boundy-Mills, K.L. Oleaginous yeasts for biodiesel: Current and future trends in biology and production. Biotechnol. Adv. 2014, 32, 1336–1360. [Google Scholar] [CrossRef]
- Martinez-Silveira, A.; Villarreal, R.; Garmendia, G.; Rufo, C.; Vero, S. Process conditions for a rapid in situ transesterification for biodiesel production from oleaginous yeasts. Electron. J. Biotechnol. 2019, 38, 1–9. [Google Scholar] [CrossRef]
- Guerfali, M.; Ayadi, I.; Sassi, H.E.; Belhassen, A.; Gargouri, A.; Belghith, H. Biodiesel-derived crude glycerol as alternative feedstock for single cell oil production by the oleaginous yeast Candida viswanathii Y-E4. Ind. Crops Prod. 2020, 145, 112103. [Google Scholar] [CrossRef]
- Madani, M.; Enshaeieh, M.; Abdoli, A. Single cell oil and its application for biodiesel production. Process Saf. Environ. Prot. 2017, 111, 747–756. [Google Scholar] [CrossRef]
- Patel, A.; Arora, N.; Pruthi, V.; Pruthi, P.A. Biological treatment of pulp and paper industry effluent by oleaginous yeast integrated with production of biodiesel as sustainable transportation fuel. J. Clean. Prod. 2017, 142, 2858–2864. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, J.; Nielsen, J. Yeast synthetic biology advances biofuel production. Curr. Opin. Microbiol. 2022, 65, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Ananthi, V.; Siva Prakash, G.; Mohan Rasu, K.; Gangadevi, K.; Boobalan, T.; Raja, R.; Anand, K.; Sudhakar, M.; Chuturgoon, A.; Arun, A. Comparison of integrated sustainable biodiesel and antibacterial nano silver production by microalgal and yeast isolates. J. Photochem. Photobiol. B 2018, 186, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Chintagunta, A.D.; Zuccaro, G.; Kumar, M.; Kumar, S.P.J.; Garlapati, V.K.; Postemsky, P.D.; Kumar, N.S.S.; Chandel, A.K.; Simal-Gandara, J. Biodiesel production from lignocellulosic biomass using oleaginous microbes: Prospects for integrated biofuel production. Front. Microbiol. 2021, 12, 658284. [Google Scholar] [CrossRef] [PubMed]
- Adrio, J.L. Oleaginous yeasts: Promising platforms for the production of oleochemicals and biofuels; Oleaginous yeasts: Promising platforms for the production of oleochemicals and biofuels. Biotechnol. Bioeng. 2017, 114, 1915–1920. [Google Scholar] [CrossRef] [PubMed]
- Knothe, G.; Krahl, J.; Gerpen, J.V. (Eds.) The Biodiesel Handbook; AOCS Press: Urbana, IL, USA, 2015. [Google Scholar]
- Knothe, G.; Steidley, K.R. Kinematic viscosity of biodiesel fuel components and related compounds. Influence of compound structure and comparison to petro diesel fuel components. Fuel 2005, 84, 1059–1065. [Google Scholar] [CrossRef]
- Phukan, M.M.; Bora, P.; Gogoi, K.; Konwar, B.K. Biodiesel from Saccharomyces cerevisiae: Fuel property analysis and comparative economics. SN Appl. Sci. 2019, 1, 153. [Google Scholar] [CrossRef]
- Knothe, G. Biodiesel and renewable diesel: A comparison. Prog. Energy Combust. Sci. 2010, 36, 364–373. [Google Scholar] [CrossRef]
- Soccol, C.R.; Dalmas Neto, C.J.; Soccol, V.T.; Sydney, E.B.; da Costa, E.S.F.; Medeiros, A.B.P.; Vandenberghe, L.P.S. Pilot scale biodiesel production from microbial oil of Rhodosporidium toruloides DEBB 5533 using sugarcane juice: Performance in diesel engine and preliminary economic study. Bioresour Technol 2017, 23, 259–268. [Google Scholar] [CrossRef]
- Saha, S.; Basak, B.; Hwang, J.H.; Salama, E.S.; Chatterjee, P.K.; Jeon, B.H. Microbial Symbiosis: A Network towards Biomethanation. Trends Microbiol. 2020, 28, 968–984. [Google Scholar] [CrossRef] [PubMed]
- Kabeyi, M.J.B.; Olanrewaju, O.A. Biogas Production and Applications in the Sustainable Energy Transition. J. Energy 2022, 2022, 1–43. [Google Scholar] [CrossRef]
- Gao, M.; Zhang, S.; Ma, X.; Guan, W.; Song, N.; Wang, Q.; Wu, C. Effect of Yeast Addition on the Biogas Production Performance of a Food Waste Anaerobic Digestion System. R. Soc. Open Sci. 2020, 7, 200443. [Google Scholar] [CrossRef]
- Tshemese, Z.; Deenadayalu, N.; Linganiso, L.Z.; Chetty, M. An Overview of Biogas Production from Anaerobic Digestion and the Possibility of Using Sugarcane Wastewater and Municipal Solid Waste in a South African Context. Appl. Syst. Innov. 2023, 6, 13. [Google Scholar] [CrossRef]
- Sikarwar, V.S.; Zhao, M.; Clough, P.; Yao, J.; Zhong, X.; Memon, M.Z.; Shah, N.; Anthony, E.J.; Fennell, P.S. An Overview of Advances in Biomass Gasification. Energy Environ. Sci. 2016, 9, 2939–2977. [Google Scholar] [CrossRef]
- Li, H.; Mehmood, D.; Thorin, E.; Yu, Z. Biomethane Production Via Anaerobic Digestion and Biomass Gasification. In Proceedings of the Energy Procedia; Elsevier: Amsterdam, The Netherlands, 2017; Volume 105, pp. 1172–1177. [Google Scholar]
- Gohain, A.; Gogoi, N.; Jha, R. Biogas Production through Anaerobic Digestion of Agricultural Wastes. In Agriculture Waste Management and Bioresource; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2023; pp. 251–267. ISBN 978-1-119-80842-8. [Google Scholar]
- Wang, Z.; Hu, Y.; Wang, S.; Wu, G.; Zhan, X. A Critical Review on Dry Anaerobic Digestion of Organic Waste: Characteristics, Operational Conditions, and Improvement Strategies. Renew. Sustain. Energy Rev. 2023, 176, 113208. [Google Scholar] [CrossRef]
- Wang, D.; Hunt, K.A.; Candry, P.; Tao, X.; Wofford, N.Q.; Zhou, J.; McInerney, M.J.; Stahl, D.A.; Tanner, R.S.; Zhou, A.; et al. Cross-Feedings, Competition, and Positive and Negative Synergies in a Four-Species Synthetic Community for Anaerobic Degradation of Cellulose to Methane. mBio 2023, 14, e03189-22. [Google Scholar] [CrossRef]
- Karrabi, M.; Ranjbar, F.M.; Shahnavaz, B.; Seyedi, S. A Comprehensive Review on Biogas Production from Lignocellulosic Wastes through Anaerobic Digestion: An Insight into Performance Improvement Strategies. Fuel 2023, 340, 127239. [Google Scholar] [CrossRef]
- Pettigrew, L.; Gutbrod, A.; Domes, H.; Groß, F.; Méndez-Contreras, J.M.; Delgado, A. Modified ADM1 for High-Rate Anaerobic Co-Digestion of Thermally Pre-Treated Brewery Surplus Yeast Wastewater. Water Sci. Technol. 2017, 76, 542–554. [Google Scholar] [CrossRef] [PubMed]
- Suwannarat, J.; Ritchie, R.J. Anaerobic Digestion of Food Waste Using Yeast. Waste Manag. 2015, 42, 61–66. [Google Scholar] [CrossRef]
- Wang, Q.; Li, H.; Feng, K.; Liu, J. Oriented Fermentation of Food Waste towards High-Value Products: A Review. Energies 2020, 13, 5638. [Google Scholar] [CrossRef]
- Ma, X.; Yu, M.; Song, N.; Xu, B.; Gao, M.; Wu, C.; Wang, Q. Effect of Ethanol Pre-Fermentation on Organic Load Rate and Stability of Semi-Continuous Anaerobic Digestion of Food Waste. Bioresour. Technol. 2020, 299, 122587. [Google Scholar] [CrossRef] [PubMed]
- Anwar Saeed, M.; Ma, H.; Yue, S.; Wang, Q.; Tu, M. Concise Review on Ethanol Production from Food Waste: Development and Sustainability. Environ. Sci. Pollut. Res. 2018, 25, 28851–28863. [Google Scholar] [CrossRef]
- Yu, M.; Wu, C.; Wang, Q.; Sun, X.; Ren, Y.; Li, Y.Y. Ethanol Prefermentation of Food Waste in Sequencing Batch Methane Fermentation for Improved Buffering Capacity and Microbial Community Analysis. Bioresour. Technol. 2018, 248, 187–193. [Google Scholar] [CrossRef]
- Syaichurrozi, I.; Rusdi, R.; Hidayat, T.; Bustomi, A. Kinetics Studies Impact of Initial pH and Addition of Yeast Saccharomyces cerevisiae on Biogas Production from Tofu Wastewater in Indonesia. Int. J. Eng. 2016, 29, 1037–1046. [Google Scholar]
- Slopiecka, K.; Liberti, F.; Massoli, S.; Bartocci, P.; Fantozzi, F. Chemical and Physical Characterization of Food Waste to Improve Its Use in Anaerobic Digestion Plants. Energy Nexus 2022, 5, 100049. [Google Scholar] [CrossRef]
- Ekpeni, L.E.N.; Nkem-Ekpeni, F.F.; Benyounis, K.Y.; Aboderheeba, A.K.M.; Stokes, J.; Olabi, A.G. Yeast: A Potential Biomass Substrate for the Production of Cleaner Energy (Biogas). Energy Procedia 2014, 61, 1718–1731. [Google Scholar] [CrossRef]
- Sosa-Hernández, O.; Parameswaran, P.; Alemán-Nava, G.S.; Torres, C.I.; Parra-Saldívar, R. Evaluating Biochemical Methane Production from Brewer’s Spent Yeast. J. Ind. Microbiol. Biotechnol. 2016, 43, 1195–1204. [Google Scholar] [CrossRef]
- Zupančič, G.D.; Panjičko, M.; Zelić, B. Biogas Production from Brewer’s Yeast Using an Anaerobic Sequencing Batch Reactor. Food Technol. Biotechnol. 2017, 55, 187. [Google Scholar] [CrossRef]
- Hashemi, S.; Karimi, K.; Taherzadeh, M.J. Integrated Process for Protein, Pigments, and Biogas Production from Baker’s Yeast Wastewater Using Filamentous Fungi. Bioresour. Technol. 2021, 337, 125356. [Google Scholar] [CrossRef]
- Moeller, L.; Bauer, A.; Zehnsdorf, A.; Lee, M.Y.; Müller, R.A. Anaerobic Co-Digestion of Waste Yeast Biomass from Citric Acid Production and Waste Frying Fat. Eng. Life Sci. 2018, 18, 425–433. [Google Scholar] [CrossRef]
- Meller, R.A.; Wenner, B.A.; Ashworth, J.; Gehman, A.M.; Lakritz, J.; Firkins, J.L. Potential Roles of Nitrate and Live Yeast Culture in Suppressing Methane Emission and Influencing Ruminal Fermentation, Digestibility, and Milk Production in Lactating Jersey Cows. J. Dairy. Sci. 2019, 102, 6144–6156. [Google Scholar] [CrossRef]
- Bryant, R.W.; Burns, E.E.R.; Feidler-Cree, C.; Carlton, D.; Flythe, M.D.; Martin, L.J. spent craft brewer’s yeast reduces production of methane and ammonia by bovine rumen microbes. Front. Anim. Sci. 2021, 2, 49. [Google Scholar] [CrossRef]
- Gong, Y.L.; Liao, X.D.; Liang, J.B.; Jahromi, M.F.; Wang, H.; Cao, Z.; Wu, Y.B. Saccharomyces cerevisiae live cells decreased in vitro methane production in intestinal content of pigs. Asian-Australas. J. Anim. Sci. 2013, 26, 856. [Google Scholar] [CrossRef] [PubMed]
- Asli, S.; Eid, R.; Hugerat, M. A novel pretreatment biotechnology for increasing methane yield from lipid-rich wastewater based on combination of hydrolytic enzymes with Candida rugosa fungus. Prep. Biochem. Biotechnol. 2021, 52, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Rasmeni, Z.Z.; Madyira, D.M.; Matheri, A.N. Optimum Loading Ratio for Co-Digested Wastewater Sludge and Brewery Spent Yeast. Energy Rep. 2022, 8, 1141–1149. [Google Scholar] [CrossRef]
- Unnikrishnan, G.; Ramasamy, V. Anaerobic digestion of pineapple waste for biogas production and application of slurry as liquid fertilizer carrier for phosphate solubilizers. Indian. J. Agric. Res. 2022, 56, 408–414. [Google Scholar] [CrossRef]
- Kotarska, K.; Dziemianowicz, W.; Swierczyńska, A. Study on the sequential combination of bioethanol and biogas production from corn straw. Molecules 2019, 24, 4558. [Google Scholar] [CrossRef]
- Enaboifo, M.A.; Adadu, C.A. Comparative study of biogas production from cocoa pod, maize husk, orange peels, pineapple peels and coconut fiber co-digested with yeast. Asian J. Dairy Food Res. 2020, 39, 344–348. [Google Scholar] [CrossRef]
- Xue, N.; Liu, S.; Xin, F.; Zhou, J.; Jia, H.; Xu, J.; Jiang, M.; Dong, W. Biomethane production from lignocellulose: Biomass recalcitrance and its impacts on anaerobic digestion. Front. Bioeng. Biotechnol. 2019, 7, 191. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, V.; Bhatt, A.K.; Mehta, S.; Sharma, V.; Rathour, R.K. Sheetal Xylitol production by Pseudomonas gessardii VXlt-16 from sugarcane bagasse hydrolysate and cost analysis. Bioprocess. Biosyst. Eng. 2022, 45, 1019–1031. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, D.; Ahuja, V.; Singh, R.; More, S.; Mudliar, S.; Kumar, M. Food-grade xylitol production from corncob biomass with acute oral toxicity studies. World J. Microbiol. Biotechnol. 2023, 39, 102. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; He, M.; Wu, B.; Wang, Y.; Hu, G.; Ma, K. Simultaneous concentration and detoxification of lignocellulosic hydrolysates by novel membrane filtration system for bioethanol production. J. Clean. Prod. 2019, 227, 1185–1194. [Google Scholar] [CrossRef]
- Dahadha, S.; Amin, Z.; Bazyar Lakeh, A.A.; Elbeshbishy, E. Evaluation of Different Pretreatment Processes of Lignocellulosic Biomass for Enhanced Biomethane Production. Energy Fuels 2017, 31, 10335–10347. [Google Scholar] [CrossRef]
- Ibro, M.K.; Ancha, V.R.; Lemma, D.B. Impacts of Anaerobic Co-Digestion on Different Influencing Parameters: A Critical Review. Sustainability 2022, 14, 9387. [Google Scholar] [CrossRef]
- Tomczak, W.; Gryta, M.; Grubecki, I.; Miłek, J. Biogas Production in AnMBRs via Treatment of Municipal and Domestic Wastewater: Opportunities and Fouling Mitigation Strategies. Appl. Sci. 2023, 13, 6466. [Google Scholar] [CrossRef]
- Barragán-Ocaña, A.; Merritt, H.; Sánchez-Estrada, O.E.; Méndez-Becerril, J.L.; Del Pilar Longar-Blanco, M. Biorefinery and sustainability for the production of biofuels and value-added products: A trends analysis based on network and patent analysis. PLoS ONE 2023, 18, e0279659. [Google Scholar] [CrossRef]


{kind=link}
| Organism | Strategy for Enhancing Biomass Utilization or Biofuel Production | Refs. |
|---|---|---|
| Hansenula polymorpha, Kluyveromyces lactis, Pichia pastoris, Yarrowia lipolytica | CRISPR-Cas enhancing biofuel production in non-conventional yeast | [30] |
| Saccharomyces cerevisiae | CRISPR/Cas9 for application in CBP | [31] |
| Saccharomyces cerevisiae | Engineering yeast for xylose utilization and ethanol production | [32] |
| Zymomonas mobilis | Metabolic engineering to broaden substrate range, remove competing pathways, and enhance tolerance to ethanol and lignocellulosic hydrolysate inhibitors | [33] |
| S. cerevisiae | Mining and identification of regulatory elements of bioethanol synthesis pathways, such as non-coding RNAs | [34] |
| S. cerevisiae | Genetic engineering, heterologous expression of cellulase genes, xylose transporters, knock-out, and the overexpression of key genes and promoters may be closely related to bioethanol yield | [35] |
| S. cerevisiae | Direction design optimization based on machine learning can effectively regulate the pretreatment parameters | [36] |
| S. cerevisiae | Improving ethanol yield via the addition of quorum-sensing molecules that deter growth of S. cerevisiae cells | [37] |
| Saccharomyces cerevisiae, Pichia stipitis | Metabolic engineering for enhanced bioethanol production | [38] |
| S. cerevisiae extract | Constructing a cell-free system using Synthetic Biology tools | [39] |
| Yarrowia lipolytica | Overexpression of Diacylglycerol acyltransferases the I (DGA) gene(s) to promote lipid accumulation | [40] |
| Rhodosporidium toruloides | Overexpressing malic enzyme and acetyl-CoA carboxylase to redirect central C metabolism to enhance the availability of precursors toward lipogenic activity | [40] |
| Y. lipolytica | Co-overexpressing the glyceraldehyde-3-phosphate dehydrogenase and malate dehydrogenase for increasing lipid accumulation | [40] |
| Y. lipolytica | The overexpression of aldehyde dehydrogenase endogenous genes to enhance the conversion of furfural to furoic acid | [40] |
| R. toruloides L1–1 | Breeding strategy to improve lipid accumulation along with stress tolerance | [40] |
| Y. lipolytica | Elimination of lipid catabolism by deleting lipid-assimilating genes, e.g., the acyl-CoA oxidases (POX) or peroxisomal biogenesis (PEX) genes | [40] |
| Role | Yeast/Source | Operating Conditions | Effect/Yield | Ref. |
|---|---|---|---|---|
| Biocatalyst | Saccharomyces cerevisiae (Yea-Sacc 1026, Alltech Inc., Nicholasville, KY, USA) | In vivo model of Jersey cows (Addition of NO3− (electron acceptor) and live yeast culture) | Methane production reduced from 22.6 mol/day to 18.8 mol/days | [107] |
| Co-substrate | Bakers’ yeast and craft yeast Saccharomyces cerevisiae (Brewery in Asheville, NC, USA) | In vitro fermentation; 10 mL rumen fluid mixture with cell suspension buffer was mixed with culture media (cornmeal and silage feed) inoculated with yeast at 39 °C in a CO2 environment | Baker’s yeast: 4760 ppm Craft yeast: 3530 ppm Bakers’ + craft yeast: 3880 ppm | [108] |
| Biocatalysts | Yeast isolate 1 (Commercial probiotic; Angela yeast Co., Guangzhou, China) S. cerevisiae YST2 (Bakery; Kuala Lumpur, Malaysia) | In vitro fermentation Large intestinal content as inoculum with freeze-dried yeast powder is inoculated in fermentation medium at 39 °C for 24 h | Methane production reduction potential >25% | [109] |
| Biocatalyst | S. cerevisiae | Food waste anaerobic digestion. Food waste, sludge, and yeast at 37 °C, 60 rpm | Biogas production increased by 33.2% | [86] |
| Source of enzyme for waste pretreatment | Enzyme mixture from Candida rogusa | Simultaneous hydrolysis and the anaerobic digestion of lipid-rich waste water from the dairy industry Enzymatic hydrolysis with lipase + Fungal enzymatic mixture 0.05 w/v | Methane yield increased by 140% | [110] |
| Co-biocatalyst | Saccharomyces cerevisiae (Ragi) | Tofu wastewater; pH 8; 1 atm pressure; rumen fluid as inoculum | 421 mL | [100] |
| Co-substrate | Brewer’s yeast | Wastewater Biosolids + brewer’s yeast; sewage sludge inoculum; period of 21 days at 37 °C | 338.2 NmL CH4/g volatile solids (VS) | [110] |
| 110Substrate | Yeast (local brewery) | Expanded granular sludge as inoculum. Temp 35 °C; pH 6.5 | Specific biogas productivity 0.430 m3/kg | [103] |
| Co-biocatalyst | Meyerozyma gullerimondi (rhizosphere) | Providencia rettgeri and Meyerozyma gullerimondi (6 × 104 colonies/mL) | Methane content 63.81% | [112] |
| Co-catalyst | S. cerevisiae D2 | Simultaneous saccharification and fermentation Step 1: alcoholic fermentation for 72 h Step 2: fermentation substrate + methane bacteria at 37 °C | Ethanol 16.98 ± 0.00 g/L Biogas 330 L/kg dry organic matter | [113] |
| Catalyst | Yeast | Case 1: 2 g yeast + 4 g coconut fibre Case 2: 2 g yeast + 4 g cocoa pods Case 3: 2 g yeast + 4 g maize husk Case 4: 2 g yeast + 4 g orange peels Case 5: 2 g yeast + 4 g pineapple peels Period: 12 days | Maize + yeast: 23.33 mL | [114] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahuja, V.; Arora, A.; Chauhan, S.; Thakur, S.; Jeyaseelan, C.; Paul, D. Yeast-Mediated Biomass Valorization for Biofuel Production: A Literature Review. Fermentation 2023, 9, 784. https://doi.org/10.3390/fermentation9090784
Ahuja V, Arora A, Chauhan S, Thakur S, Jeyaseelan C, Paul D. Yeast-Mediated Biomass Valorization for Biofuel Production: A Literature Review. Fermentation. 2023; 9(9):784. https://doi.org/10.3390/fermentation9090784
Chicago/Turabian StyleAhuja, Vishal, Anju Arora, Shikha Chauhan, Sheetal Thakur, Christine Jeyaseelan, and Debarati Paul. 2023. "Yeast-Mediated Biomass Valorization for Biofuel Production: A Literature Review" Fermentation 9, no. 9: 784. https://doi.org/10.3390/fermentation9090784