
Photobioreactor Design for Polyhydroxyalkanoate Production Using Anoxygenic Photoheterotrophs: A Review

Abstract
:1. Introduction
2. Purple Non-Sulfur Bacteria (PNSB)
3. Polyhydroxyalkanoates (PHAs)
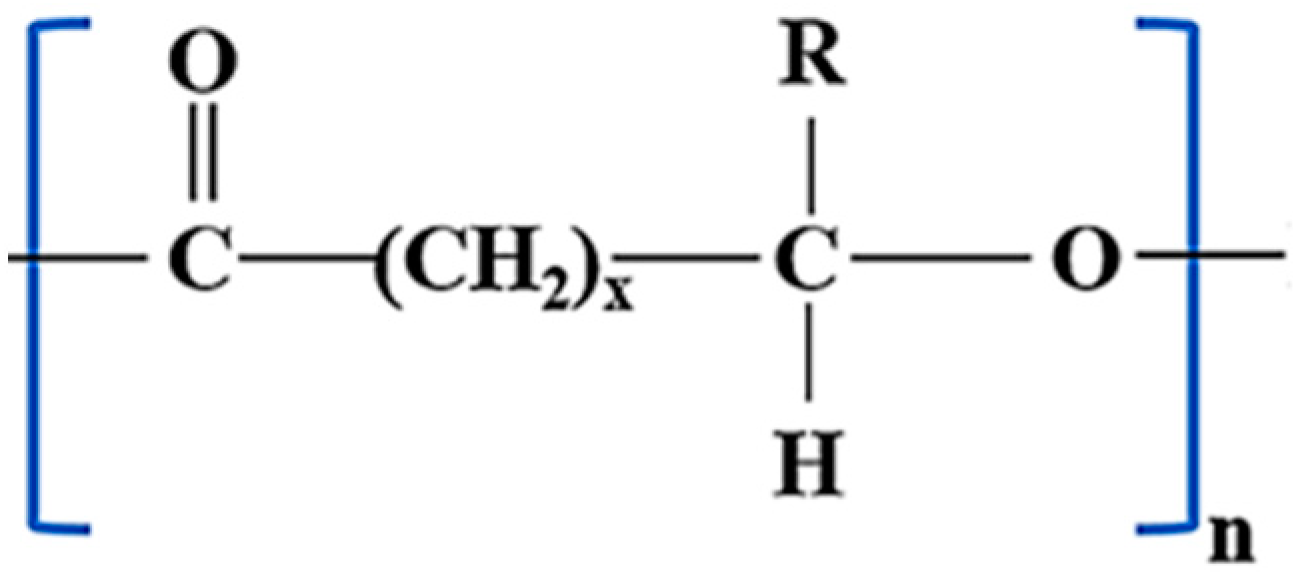
3.1. Chemical Structure, Classification, and Properties of PHAs
3.2. Commercial Development of PHAs
3.3. Applications of PHAs
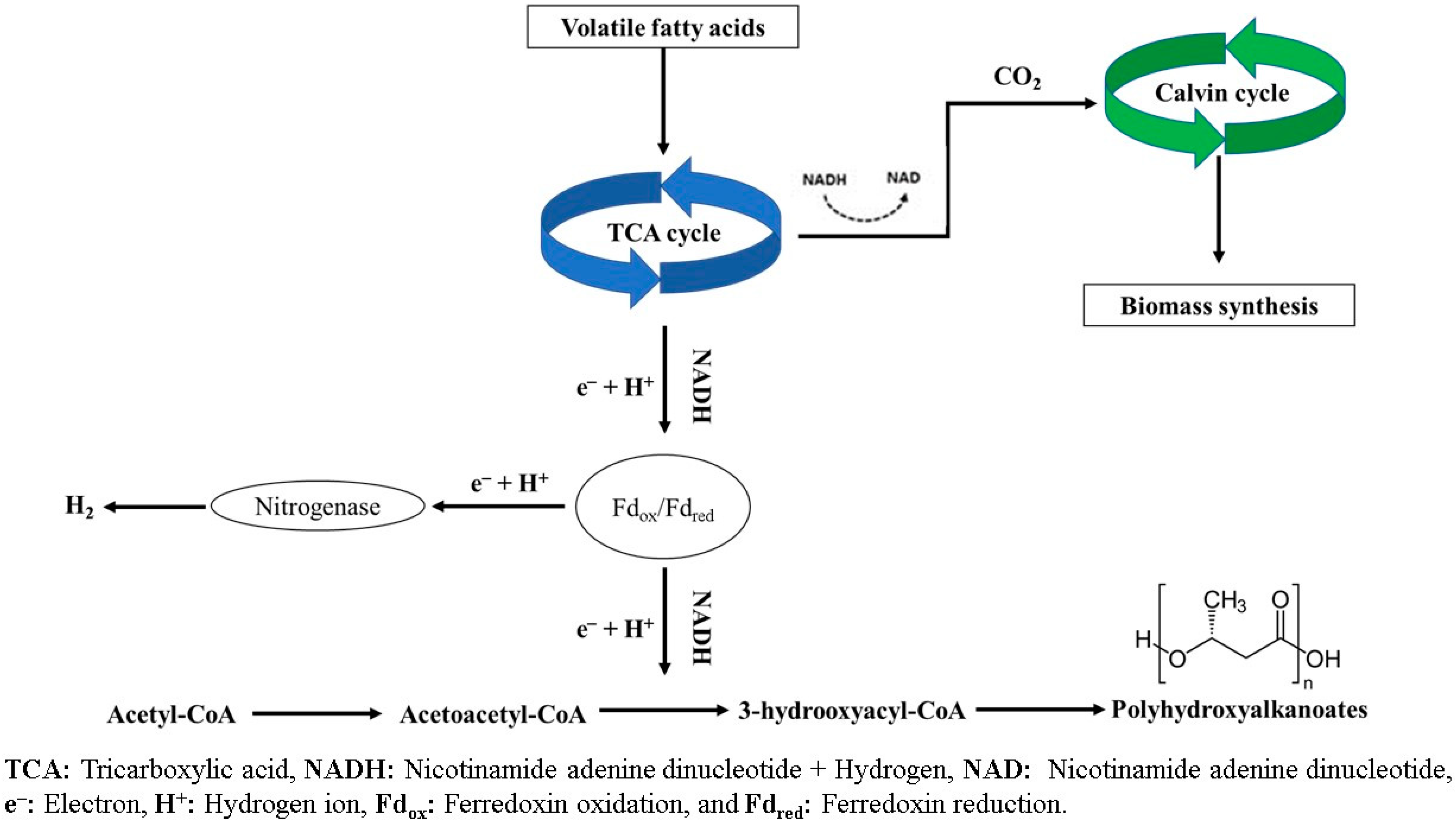
4. PHA Biosynthesis Using PNSB
4.1. Influence of Carbon and Nutrients
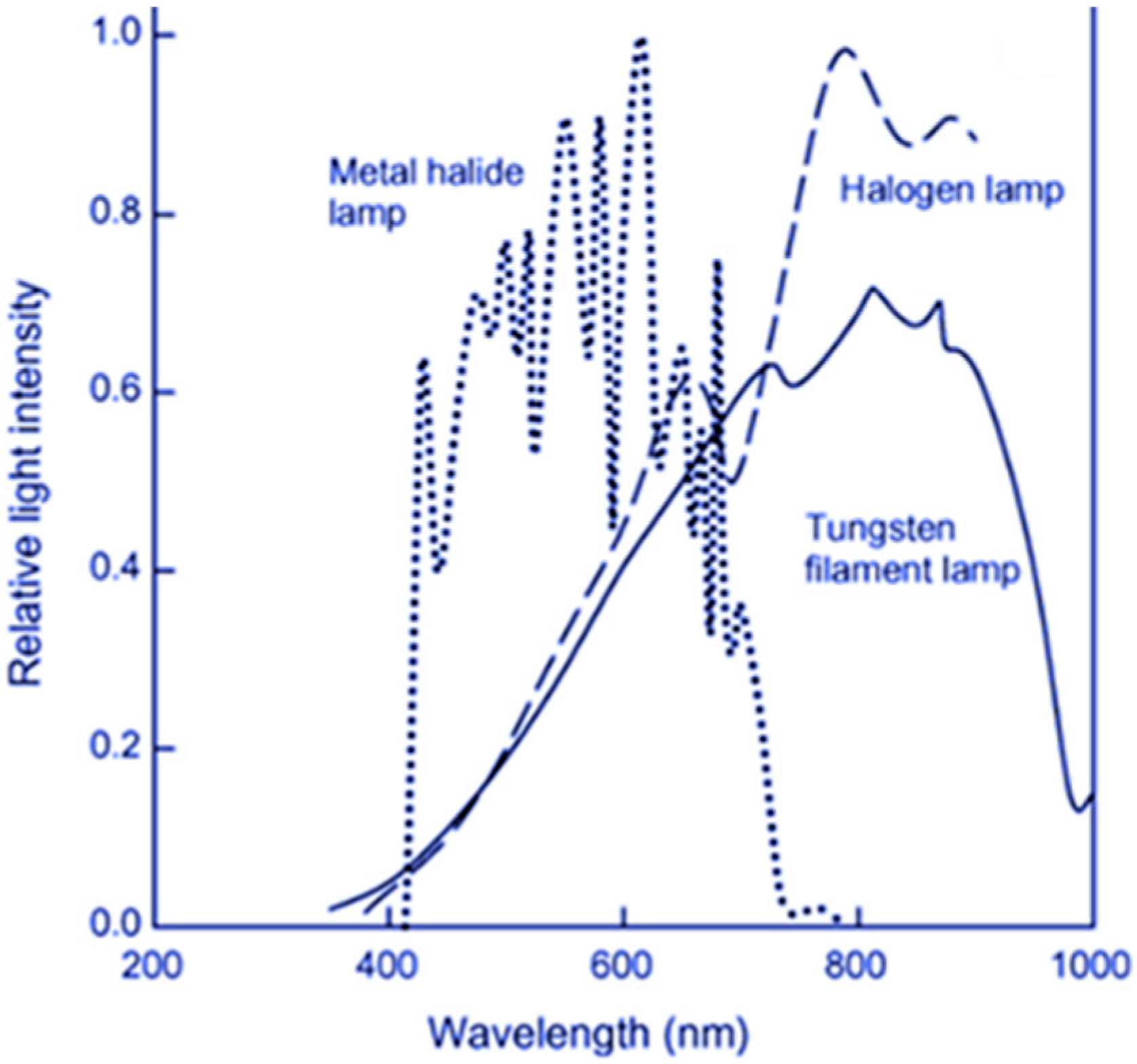
4.2. Influence of Light

5. Photobioreactors
5.1. Suspended-Culture PBRs
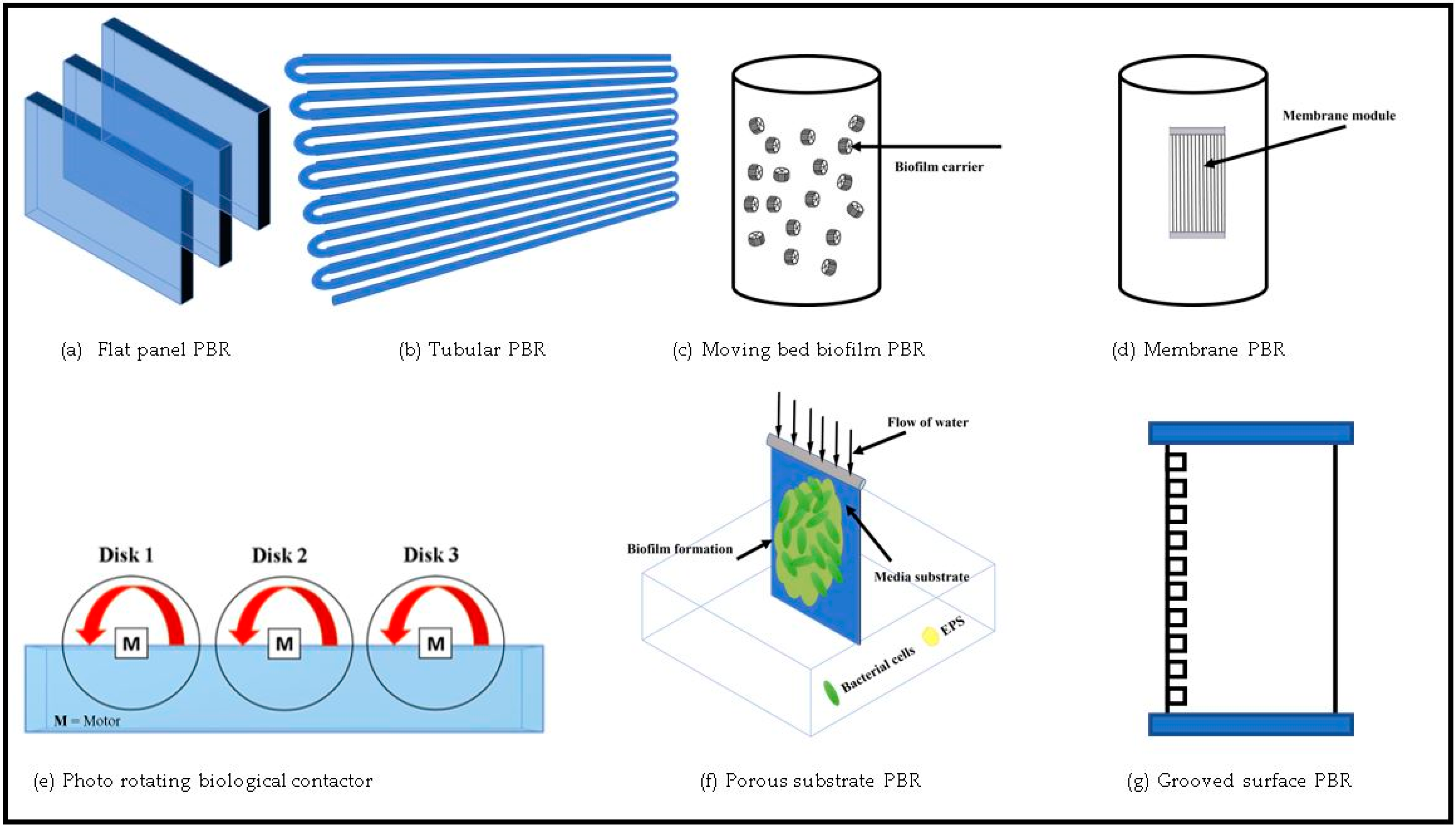
5.1.1. Flat-Panel (Plate) Photobioreactors
5.1.2. Tubular Photobioreactor
5.1.3. Membrane Photobioreactor (MPBR)
5.2. Immobilized Carrier Systems
5.2.1. Artificial Immobilization on or within Suspended Carriers
5.2.2. Self-Immobilized Granular Systems
5.2.3. Moving-Bed Biofilm Photobioreactor (MBBPR)
5.3. Fixed-Growth Biofilm Systems
5.3.1. Photo-Rotating Biological Contactor (PRBC)
5.3.2. Porous-Substrate Photobioreactor (PSBR)
5.3.3. Grooved-Surface PBR
5.3.4. Other Biofilm Photobioreactors
6. Summary and Future Directions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Geyer, R.; Jambeck, J.R.; Law, K.L. Production, Use, and Fate of All Plastics Ever Made. Sci. Adv. 2017, 3, e1700782. [Google Scholar] [CrossRef] [PubMed]
- Javaid, H.; Nawaz, A.; Riaz, N.; Mukhtar, H.; -Ul-Haq, I.; Shah, K.A.; Khan, H.; Naqvi, S.M.; Shakoor, S.; Rasool, A.; et al. Biosynthesis of Polyhydroxyalkanoates (PHAs) by the Valorization of Biomass and Synthetic Waste. Molecules 2020, 25, 5539. [Google Scholar] [CrossRef] [PubMed]
- Baierl, T.; Bogner, F.X. Plastic Pollution: Learning Activities from Production to Disposal—From Where Do Plastics Come & Where Do They Go? Am. Biol. Teach. 2021, 83, 320–324. [Google Scholar]
- Surendran, A.; Lakshmanan, M.; Chee, J.Y.; Sulaiman, A.M.; Thuoc, D.V.; Sudesh, K. Can Polyhydroxyalkanoates Be Produced Efficiently from Waste Plant and Animal Oils? Front. Bioeng. Biotechnol. 2020, 8, 169. [Google Scholar] [CrossRef]
- Gholami, A.; Mohkam, M.; Rasoul-Amini, S.; Ghasemi, Y. Industrial Production of Polyhydroxyalkanoates by Bacteria: Opportunities and Challenges. Minerva Biotecnol. 2016, 28, 59–74. [Google Scholar]
- Albuquerque, M.G.E.; Torres, C.A.V.; Reis, M.A.M. Polyhydroxyalkanoate (PHA) Production by a Mixed Microbial Culture Using Sugar Molasses: Effect of the Influent Substrate Concentration on Culture Selection. Water Res. 2010, 44, 3419–3433. [Google Scholar] [CrossRef]
- Ciesielski, S.; Mozejko, J.; Pisutpaisal, N. Plant Oils as Promising Substrates for Polyhydroxyalkanoates Production. J. Clean. Prod. 2015, 106, 408–421. [Google Scholar] [CrossRef]
- Basak, N.; Das, D. The Prospect of Purple Non-Sulfur (PNS) Photosynthetic Bacteria for Hydrogen Production: The Present State of the Art. World J. Microbiol. Biotechnol. 2007, 23, 31–42. [Google Scholar] [CrossRef]
- Alloul, A.; Cerruti, M.; Adamczyk, D.; Weissbrodt, D.; Vlaeminck, S. Control Tools to Selectively Produce Purple Bacteria for Microbial Protein in Raceway Reactors. bioRxiv 2020. [Google Scholar] [CrossRef]
- Ahmad, I.; Abdullah, N.; Koji, I.; Yuzir, A.; Eva Muhammad, S. Evolution of Photobioreactors: A Review Based on Microalgal Perspective. IOP Conf. Ser. Mater. Sci. Eng. 2021, 1142, 012004. [Google Scholar] [CrossRef]
- Brennan, L.; Owende, P. Biofuels from Microalgae-A Review of Technologies for Production, Processing, and Extractions of Biofuels and Co-Products. Renew. Sustain. Energy Rev. 2010, 14, 557–577. [Google Scholar] [CrossRef]
- Corona, V.M.; Buitrón, G. Polyhydroxyalkanoates from Organic Waste Streams Using Purple Non-Sulfur Bacteria. Bioresour. Technol. 2021, 323, 124610. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.; Ganzerli, S.; Rugiero, I.; Pellizzari, S.; Pedrini, P.; Tamburini, E. Potential of Rhodobacter capsulatus Grown in Anaerobic-Light or Aerobic-Dark Conditions as Bioremediation Agent for Biological Wastewater Treatments. Water 2017, 9, 108. [Google Scholar] [CrossRef]
- Madigan, M.T.; Jung, D.O. An Overview of Purple Bacteria: Systematics, Physiology, and Habitats; Springer: Dordrecht, The Netherlands, 2009; pp. 1–15. ISBN 9781402088155. [Google Scholar]
- Reungsang, A.; Zhong, N.; Yang, Y.; Sittijunda, S.; Xia, A.; Liao, Q. Hydrogen from Photo Fermentation. In Green Energy and Technology; Springer: Berlin/Heidelberg, Germany, 2018; pp. 221–317. [Google Scholar]
- Lu, H.; Zhang, G.; Wan, T.; Lu, Y. Influences of Light and Oxygen Conditions on Photosynthetic Bacteria Macromolecule Degradation: Different Metabolic Pathways. Bioresour. Technol. 2011, 102, 9503–9508. [Google Scholar] [CrossRef]
- Chen, J.; Wei, J.; Ma, C.; Yang, Z.; Li, Z.; Yang, X.; Wang, M.; Zhang, H.; Hu, J.; Zhang, C. Photosynthetic Bacteria-Based Technology Is a Potential Alternative to Meet Sustainable Wastewater Treatment Requirement? Environ. Int. 2020, 137, 105417. [Google Scholar] [CrossRef]
- Mukhopadhyay, M.; Patra, A.; Paul, A.K. Phototrophic Growth and Accumulation of Poly(3-Hydroxybutyrate-Co-3-Hydroxyvalerate) by Purple Nonsulfur Bacterium Rhodopseudomonas palustris SP5212. J. Polym. 2013, 2013, 523941. [Google Scholar] [CrossRef]
- Gironi, F.; Piemonte, V. Bioplastics and Petroleum-Based Plastics: Strengths and Weaknesses. Energy Sources Part A Recovery Util. Environ. Eff. 2011, 33, 1949–1959. [Google Scholar] [CrossRef]
- Możejko-Ciesielska, J.; Kiewisz, R. Bacterial Polyhydroxyalkanoates: Still Fabulous? Microbiol. Res. 2016, 192, 271–282. [Google Scholar] [CrossRef]
- Rajan, K.P.; Thomas, S.P.; Gopanna, A.; Chavali, M. Polyhydroxybutyrate (PHB): A Standout Biopolymer for Environmental Sustainability. In Handbook of Ecomaterials; Springer International Publishing: Berlin/Heidelberg, Germany, 2018; pp. 1–23. [Google Scholar]
- Licciardello, G.; Catara, A.F.; Catara, V. Production of Polyhydroxyalkanoates and Extracellular Products Using Pseudomonas Corrugata and P. Mediterranea: A Review. Bioengineering 2019, 6, 105. [Google Scholar] [CrossRef]
- Zheng, Y.; Chen, J.C.; Ma, Y.M.; Chen, G.Q. Engineering Biosynthesis of Polyhydroxyalkanoates (PHA) for Diversity and Cost Reduction. Metab. Eng. 2020, 58, 82–93. [Google Scholar] [CrossRef]
- Aeschelmann, F.; Carus, M. Biobased Building Blocks and Polymers in the World: Capacities, Production, and Applications-Status Quo and Trends towards 2020. Ind. Biotechnol. 2015, 11, 154–159. [Google Scholar] [CrossRef]
- Market Polyhydroxyalkanoate (PHA) Market by Type, Production Method, Application, and Region—Global Forecast to 2025. Available online: https://www.prnewswire.com/news-releases/polyhydroxyalkanoate-pha-market-by-type-production-method-application-and-region---global-forecast-to-2025-301227618.html (accessed on 1 March 2021).
- Singh, A.K.; Srivastava, J.K.; Chandel, A.K.; Sharma, L.; Mallick, N.; Singh, S.P. Biomedical Applications of Microbially Engineered Polyhydroxyalkanoates: An Insight into Recent Advances, Bottlenecks, and Solutions. Appl. Microbiol. Biotechnol. 2019, 103, 2007–2032. [Google Scholar] [CrossRef] [PubMed]
- Kourmentza, C.; Ntaikou, I.; Lyberatos, G.; Kornaros, M. Polyhydroxyalkanoates from Pseudomonas Sp. Using Synthetic and Olive Mill Wastewater under Limiting Conditions. Int. J. Biol. Macromol. 2015, 74, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Bugnicourt, E.; Cinelli, P.; Lazzeri, A.; Alvarez, V. Polyhydroxyalkanoate (PHA): Review of Synthesis, Characteristics, Processing and Potential Applications in Packaging. Express Polym. Lett. 2014, 8, 791–808. [Google Scholar] [CrossRef]
- Angelova, N.; Hunkeler, D. Rationalizing the Design of Polymeric Biomaterials. Trends Biotechnol. 1999, 17, 409–421. [Google Scholar] [CrossRef]
- Madison, L.L.; Huisman, G.W. Metabolic Engineering of Poly(3-Hydroxyalkanoates): From DNA to Plastic. Microbiol. Mol. Biol. Rev. 1999, 63, 21–53. [Google Scholar] [CrossRef] [PubMed]
- Noda, I. Films Comprising Biodegradable PHA Copolymers. U.S. Patent 6,174,990, 16 January 2001. [Google Scholar]
- Mudenur, C.; Mondal, K.; Singh, U.; Katiyar, V. Production of Polyhydroxyalkanoates and Its Potential Applications; Springer: Singapore, 2019; pp. 131–164. [Google Scholar]
- Jendrossek, D.; Schirmer, A.; Schlegel, H.G. Biodegradation of Polyhydroxyalkanoic Acids. Appl. Microbiol. Biotechnol. 1996, 46, 451–463. [Google Scholar] [CrossRef]
- Pötter, M.; Steinbüchel, A. Biogenesis and Structure of Polyhydroxyalkanoate Granules. In Inclusions in Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2006; pp. 109–136. [Google Scholar] [CrossRef]
- Fradinho, J.C.; Oehmen, A.; Reis, M.A.M. Improving Polyhydroxyalkanoates Production in Phototrophic Mixed Cultures by Optimizing Accumulator Reactor Operating Conditions. Int. J. Biol. Macromol. 2019, 126, 1085–1092. [Google Scholar] [CrossRef]
- Sangkharak, K.; Prasertsan, P. Nutrient Optimization for Production of Polyhydroxybutyrate from Halotolerant Photosynthetic Bacteria Cultivated under Aerobic-Dark Condition. Electron. J. Biotechnol. 2008, 11, 83–94. [Google Scholar] [CrossRef]
- Khatipov, E.; Miyake, M.; Miyake, J.; Asada, Y. Accumulation of Poly-β-Hydroxybutyrate by Rhodobacter sphaeroides on Various Carbon and Nitrogen Substrates. FEMS Microbiol. Lett. 1998, 162, 39–45. [Google Scholar] [CrossRef]
- Liu, J.; Zhao, Y.; Diao, M.; Wang, W.; Hua, W.; Wu, S.; Chen, P.; Ruan, R.; Cheng, Y. Poly(3-Hydroxybutyrate-Co-3-Hydroxyvalerate) Production by Rhodospirillum rubrum Using a Two-Step Culture Strategy. J. Chem. 2019, 2019, 8369179. [Google Scholar] [CrossRef]
- Segura, P.C.; Wattiez, R.; Vande Wouwer, A.; Leroy, B.; Dewasme, L. Dynamic Modeling of Rhodospirillum rubrum PHA Production Triggered by Redox Stress during VFA Photoheterotrophic Assimilations. J. Biotechnol. 2022, 360, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Bayon-Vicente, G.; Zarbo, S.; Deutschbauer, A.; Wattiez, R.; Leroy, B. Photoheterotrophic Assimilation of Valerate and Associated Polyhydroxyalkanoate Production by Rhodospirillum rubrum. Appl. Environ. Microbiol. 2020, 86, e00901-20. [Google Scholar] [CrossRef]
- Cerruti, M.; Ouboter, H.T.; Chasna, V.; van Loosdrecht, M.C.M.; Picioreanu, C.; Weissbrodt, D.G. Effects of Light/Dark Diel Cycles on the Photoorganoheterotrophic Metabolism of Rhodopseudomonas palustris for Differential Electron Allocation to PHAs and H2. bioRxiv 2020. bioRxiv:2020.08.19.258533. [Google Scholar]
- Imhoff, J.F. Rhodopseudomonas marina sp. Nov., a New Marine Phototrophic Purple Bacterium. Syst. Appl. Microbiol. 1983, 4, 512–521. [Google Scholar] [CrossRef]
- Almeida, J.R.; Fradinho, J.C.; Carvalho, G.; Oehmen, A.; Reis, M.A.M. Dynamics of Microbial Communities in Phototrophic Polyhydroxyalkanoate Accumulating Cultures. Microorganisms 2022, 10, 351. [Google Scholar] [CrossRef]
- Kompantseva, E.J. A New Species of Budding Purple Bacterium: Rhodopseudomonas julia sp. Nov. Microbiologiya 1989, 58, 254–259. [Google Scholar]
- Carlozzi, P.; Seggiani, M.; Cinelli, P.; Mallegni, N.; Lazzeri, A. Photofermentative Poly-3-Hydroxybutyrate Production by Rhodopseudomonas sp. S16-VOGS3 in a Novel Outdoor 70-L Photobioreactor. Sustainability 2018, 10, 3133. [Google Scholar] [CrossRef]
- Touloupakis, E.; Poloniataki, E.G.; Ghanotakis, D.F.; Carlozzi, P. Production of Biohydrogen and/or Poly-β-Hydroxybutyrate by Rhodopseudomonas sp. Using Various Carbon Sources as Substrate. Appl. Biochem. Biotechnol. 2021, 193, 307–318. [Google Scholar] [CrossRef]
- Higuchi-Takeuchi, M.; Morisaki, K.; Toyooka, K.; Numata, K. Synthesis of High-Molecular-Weight Polyhydroxyalkanoates by Marine Photosynthetic Purple Bacteria. PLoS ONE 2016, 11, e0160981. [Google Scholar] [CrossRef]
- Demiriz, B.Ö.; Kars, G.; Yücel, M.; Eroğlu, İ.; Gündüz, U. Hydrogen and Poly-Β-Hydroxybutyric Acid Production at Various Acetate Concentrations Using Rhodobacter capsulatus DSM 1710. Int. J. Hydrogen Energy 2019, 44, 17269–17277. [Google Scholar] [CrossRef]
- López-Abelairas, M.; García-Torreiro, M.; Lú-Chau, T.; Lema, J.M.; Steinbüchel, A. Comparison of Several Methods for the Separation of Poly(3-Hydroxybutyrate) from Cupriavidus Necator H16 Cultures. Biochem. Eng. J. 2015, 93, 250–259. [Google Scholar] [CrossRef]
- Monroy, I.; Buitron, G. Production of Polyhydroxybutyrate by Pure and Mixed Cultures of Purple Non-Sulfur Bacteria: A Review. J. Biotechnol. 2020, 317, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Carlozzi, P.; Touloupakis, E. Bioplastic Production by Feeding the Marine Rhodovulum Sulfidophilum DSM-1374 with Four Different Carbon Sources under Batch, Fed-Batch and Semi-Continuous Growth Regimes. New Biotechnol. 2021, 62, 10–17. [Google Scholar] [CrossRef]
- Schmid, M.; Raschbauer, M.; Song, H.; Bauer, C.; Neureiter, M. Effects of Nutrient and Oxygen Limitation, Salinity and Type of Salt on the Accumulation of Poly(3-Hydroxybutyrate) in Bacillus Megaterium Uyuni S29 with Sucrose as a Carbon Source. New Biotechnol. 2021, 61, 137–144. [Google Scholar] [CrossRef]
- Carlozzi, P.; Giovannelli, A.; Traversi, M.L.; Touloupakis, E.; Di Lorenzo, T. Poly-3-Hydroxybutyrate and H2 Production by Rhodopseudomonas sp. S16-VOGS3 Grown in a New Generation Photobioreactor under Single or Combined Nutrient Deficiency. Int. J. Biol. Macromol. 2019, 135, 821–828. [Google Scholar] [CrossRef]
- Alsafadi, D.; Al-Mashaqbeh, O. A One-Stage Cultivation Process for the Production of Poly-3-(Hydroxybutyrate-Co-Hydroxyvalerate) from Olive Mill Wastewater by Haloferax Mediterranei. New Biotechnol. 2017, 34, 47–53. [Google Scholar] [CrossRef]
- Ghimire, A.; Valentino, S.; Frunzo, L.; Pirozzi, F.; Lens, P.N.L.; Esposito, G. Concomitant Biohydrogen and Poly-β-Hydroxybutyrate Production from Dark Fermentation Effluents by Adapted Rhodobacter sphaeroides and Mixed Photofermentative Cultures. Bioresour. Technol. 2016, 217, 157–164. [Google Scholar] [CrossRef]
- Kumar, G.; Ponnusamy, V.K.; Bhosale, R.R.; Shobana, S.; Yoon, J.J.; Bhatia, S.K.; Rajesh Banu, J.; Kim, S.H. A Review on the Conversion of Volatile Fatty Acids to Polyhydroxyalkanoates Using Dark Fermentative Effluents from Hydrogen Production. Bioresour. Technol. 2019, 287, 121427. [Google Scholar] [CrossRef]
- Fradinho, J.C.; Oehmen, A.; Reis, M.A.M. Photosynthetic Mixed Culture Polyhydroxyalkanoate (PHA) Production from Individual and Mixed Volatile Fatty Acids (VFAs): Substrate Preferences and Co-Substrate Uptake. J. Biotechnol. 2014, 185, 19–27. [Google Scholar] [CrossRef]
- Liu, S.; Daigger, G.T.; Kang, J.; Zhang, G. Effects of Light Intensity and Photoperiod on Pigments Production and Corresponding Key Gene Expression of Rhodopseudomonas palustris in a Photobioreactor System. Bioresour. Technol. 2019, 294, 122172. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Wang, Q.; Wu, Y.S.; Ai, X.C.; Zhang, X.J.; Huang, Y.G.; Zhang, X.K.; Zhang, J.P. Triplet Excitation Transfer between Carotenoids in the LH2 Complex from Photosynthetic Bacterium Rhodopseudomonas palustris. Photosynth. Res. 2004, 82, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Kuo, F.S.; Chien, Y.H.; Chen, C.J. Effects of Light Sources on Growth and Carotenoid Content of Photosynthetic Bacteria Rhodopseudomonas palustris. Bioresour. Technol. 2012, 113, 315–318. [Google Scholar] [CrossRef] [PubMed]
- Adessi, A.; Philipps, R. De Photosynthesis and Hydrogen Production in Purple Non Sulfur Bacteria: Fundamental and Applied Aspects. In Microbial BioEnergy: Hydrogen Production, Advances in Photosynthesis and Respiration; Springer: Dordrecht, The Netherlands, 2014; ISBN 978-94-017-8553-2. [Google Scholar]
- Adessi, A.; De Philippis, R. Photobioreactor Design and Illumination Systems for H2 Production with Anoxygenic Photosynthetic Bacteria: A Review. Int. J. Hydrogen Energy 2014, 39, 3127–3141. [Google Scholar] [CrossRef]
- Li, K.; Zhao, C.; Yue, H.; Yang, S. A Unique Low Light Adaptation Mechanism in Rhodobacter azotoformans. J. Basic Microbiol. 2014, 54, 1350–1357. [Google Scholar] [CrossRef] [PubMed]
- Brotosudarmo, T.H.P.; Limantara, L.; Heriyanto; Prihastyanti, M.N.U. Adaptation of the Photosynthetic Unit of Purple Bacteria to Changes of Light Illumination Intensities. Procedia Chem. 2015, 14, 414–421. [Google Scholar] [CrossRef]
- Kotecha, A.; Georgiou, T.; Papiz, M.Z. Evolution of Low-Light Adapted Peripheral Light-Harvesting Complexes in Strains of Rhodopseudomonas palustris. Photosynth. Res. 2013, 114, 155–164. [Google Scholar] [CrossRef]
- Akkerman, I.; Janssen, M.; Rocha, J.; Wijffels, R.H. Photobiological Hydrogen Production: Photochemical Efficiency and Bioreactor Design. Int. J. Hydrogen Energy 2002, 27, 1195–1208. [Google Scholar] [CrossRef]
- Asada, Y.; Miyake, J. Photobiological Hydrogen Production. J. Biosci. Bioeng. 1999, 88, 1–6. [Google Scholar] [CrossRef]
- Kawagoshi, Y.; Oki, Y.; Nakano, I.; Fujimoto, A.; Takahashi, H. Biohydrogen Production by Isolated Halotolerant Photosynthetic Bacteria Using Long-Wavelength Light-Emitting Diode (LW-LED). Int. J. Hydrogen Energy 2010, 35, 13365–13369. [Google Scholar] [CrossRef]
- Blanken, W.; Cuaresma, M.; Wijffels, R.H.; Janssen, M. Cultivation of Microalgae on Artificial Light Comes at a Cost. Algal Res. 2013, 2, 333–340. [Google Scholar] [CrossRef]
- George, D.M.; Vincent, A.S.; Mackey, H.R. An Overview of Anoxygenic Phototrophic Bacteria and Their Applications in Environmental Biotechnology for Sustainable Resource Recovery. Biotechnol. Rep. 2020, 28, e00563. [Google Scholar] [CrossRef] [PubMed]
- Capson-Tojo, G.; Batstone, D.J.; Grassino, M.; Hülsen, T. Light Attenuation in Enriched Purple Phototrophic Bacteria Cultures: Implications for Modelling and Reactor Design. Water Res. 2022, 219, 118572. [Google Scholar] [CrossRef]
- Yu, S.; Peng, L.; Xu, Y.; Song, S.; Xie, G.J.; Liu, Y.; Ni, B.J. Optimizing Light Sources for Selective Growth of Purple Bacteria and Efficient Formation of Value-Added Products. J. Clean. Prod. 2021, 280, 124493. [Google Scholar] [CrossRef]
- Chen, C.Y.; Lee, C.M.; Chang, J.S. Hydrogen Production by Indigenous Photosynthetic Bacterium Rhodopseudomonas palustris WP3-5 Using Optical Fiber-Illuminating Photobioreactors. Biochem. Eng. J. 2006, 32, 33–42. [Google Scholar] [CrossRef]
- Zhou, Q.; Zhang, P.; Zhang, G. Biomass and Pigments Production in Photosynthetic Bacteria Wastewater Treatment: Effects of Light Sources. Bioresour. Technol. 2015, 179, 505–509. [Google Scholar] [CrossRef] [PubMed]
- Muzziotti, D.; Adessi, A.; Faraloni, C.; Torzillo, G.; De Philippis, R. H2 Production in Rhodopseudomonas palustris as a Way to Cope with High Light Intensities. Res. Microbiol. 2016, 167, 350–356. [Google Scholar] [CrossRef]
- Higuchi-Takeuchi, M.; Numata, K. Acetate-Inducing Metabolic States Enhance Polyhydroxyalkanoate Production in Marine Purple Non-Sulfur Bacteria under Aerobic Conditions. Front. Bioeng. Biotechnol. 2019, 7, 118. [Google Scholar] [CrossRef]
- Montiel Corona, V.; Le Borgne, S.; Revah, S.; Morales, M. Effect of Light-Dark Cycles on Hydrogen and Poly-β-Hydroxybutyrate Production by a Photoheterotrophic Culture and Rhodobacter capsulatus Using a Dark Fermentation Effluent as Substrate. Bioresour. Technol. 2017, 226, 238–246. [Google Scholar] [CrossRef]
- Fradinho, J.C.; Oehmen, A.; Reis, M.A.M. Effect of Dark/Light Periods on the Polyhydroxyalkanoate Production of a Photosynthetic Mixed Culture. Bioresour. Technol. 2013, 148, 474–479. [Google Scholar] [CrossRef]
- Fradinho, J.C.; Domingos, J.M.B.; Carvalho, G.; Oehmen, A.; Reis, M.A.M. Polyhydroxyalkanoates Production by a Mixed Photosynthetic Consortium of Bacteria and Algae. Bioresour. Technol. 2013, 132, 146–153. [Google Scholar] [CrossRef]
- Lorrungruang, C.; Martthong, J.; Sasaki, K.; Noparatnaraporn, N. Selection of Photosynthetic Bacterium Rhodobacter sphaeroides 14F for Polyhydroxyalkanoate Production with Two-Stage Aerobic Dark Cultivation. J. Biosci. Bioeng. 2006, 102, 128–131. [Google Scholar] [CrossRef] [PubMed]
- Boran, E.; Özgür, E.; Van Der Burg, J.; Yücel, M.; Gündüz, U.; Eroglu, I. Biological Hydrogen Production by Rhodobacter capsulatus in Solar Tubular Photo Bioreactor. J. Clean. Prod. 2010, 18, S29–S35. [Google Scholar] [CrossRef]
- Basak, N.; Jana, A.K.; Das, D.; Saikia, D. Photofermentative Molecular Biohydrogen Production by Purple-Non-Sulfur (PNS) Bacteria in Various Modes: The Present Progress and Future Perspective. Int. J. Hydrogen Energy 2014, 39, 6853–6871. [Google Scholar] [CrossRef]
- Kumar, V.; Jain, S.M. Plants and Algae Species: Promising Renewable Energy Production Source. Emir. J. Food Agric. 2014, 26, 679–692. [Google Scholar] [CrossRef]
- Deo, D.; Ozgur, E.; Eroglu, I.; Gunduz, U.; Yucel, M. Photofermentative Hydrogen Production in Outdoor Conditions. In Hydrogen Energy—Challenges and Perspectives; Intech: London, UK, 2012. [Google Scholar] [CrossRef]
- Eroglu, I.; Özgür, E.; Eroglu, E.; Yücel, M.; Gündüz, U. Applications of Photofermentative Hydrogen Production; Springer: Dordrecht, The Netherlands, 2014; pp. 237–267. [Google Scholar] [CrossRef]
- Wang, B.; Lan, C.Q.; Horsman, M. Closed Photobioreactors for Production of Microalgal Biomasses. Biotechnol. Adv. 2012, 30, 904–912. [Google Scholar] [CrossRef]
- Avcioglu, S.G. Scale Up of Panel Photobioreactors for Hydrogen Production by PNS Bacteria. Master’s Thesis, Middle East Technical University, Ankara, Turkey, 2010. [Google Scholar]
- Flickinger, M.C. Upstream Industrial Biotechnology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013. [Google Scholar]
- Gilbert, J.J.; Ray, S.; Das, D. Hydrogen Production Using Rhodobacter sphaeroides (O.U. 001) in a Flat Panel Rocking Photobioreactor. Int. J. Hydrogen Energy 2011, 36, 3434–3441. [Google Scholar] [CrossRef]
- Eroǧlu, I.; Tabanoǧlu, A.; Gündüz, U.; Eroǧlu, E.; Yücel, M. Hydrogen Production by Rhodobacter sphaeroides O.U.001 in a Flat Plate Solar Bioreactor. Int. J. Hydrogen Energy 2008, 33, 531–541. [Google Scholar] [CrossRef]
- Carlozzi, P.; Sacchi, A. Biomass Production and Studies on Rhodopseudomonas Palustris Grown in an Outdoor, Temperature Controlled, Underwater Tubular Photobioreactor. J. Biotechnol. 2001, 88, 239–249. [Google Scholar] [CrossRef]
- Carlozzi, P.; Pushparaj, B.; Degl’Innocenti, A.; Capperucci, A. Growth Characteristics of Rhodopseudomonas palustris Cultured Outdoors, in an Underwater Tubular Photobioreactor, and Investigation on Photosynthetic Efficiency. Appl. Microbiol. Biotechnol. 2006, 73, 789–795. [Google Scholar] [CrossRef]
- Adessi, A.; Torzillo, G.; Baccetti, E.; De Philippis, R. Sustained Outdoor H 2 Production with Rhodopseudomonas palustris Cultures in a 50 L Tubular Photobioreactor. Int. J. Hydrogen Energy 2012, 37, 8840–8849. [Google Scholar] [CrossRef]
- Luo, Y.; Le-Clech, P.; Henderson, R.K. Assessment of Membrane Photobioreactor (MPBR) Performance Parameters and Operating Conditions. Water Res. 2018, 138, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Chitapornpan, S.; Chiemchaisri, C.; Chiemchaisri, W.; Honda, R.; Yamamoto, K. Organic Carbon Recovery and Photosynthetic Bacteria Population in an Anaerobic Membrane Photo-Bioreactor Treating Food Processing Wastewater. Bioresour. Technol. 2013, 141, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Sawayama, S.; Tsukahara, K.; Yagishita, T. Wastewater Treatment and Poly-β-Hydroxybutyrate Production Using Lighted Upflow Anaerobic Sludge Blanket Method. J. Biosci. Bioeng. 1999, 87, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Z.; Liao, Q.; Zhu, X.; Tian, X.; Zhang, C. Characteristics of Hydrogen Production and Substrate Consumption of Rhodopseudomonas palustris CQK 01 in an Immobilized-Cell Photobioreactor. Bioresour. Technol. 2010, 101, 4034–4041. [Google Scholar] [CrossRef]
- Elkahlout, K.; Alipour, S.; Eroglu, I.; Gunduz, U.; Yucel, M. Long-Term Biological Hydrogen Production by Agar Immobilized Rhodobacter capsulatus in a Sequential Batch Photobioreactor. Bioprocess Biosyst. Eng. 2017, 40, 589–599. [Google Scholar] [CrossRef]
- Sonwani, R.K.; Swain, G.; Giri, B.S.; Singh, R.S.; Rai, B.N. A Novel Comparative Study of Modified Carriers in Moving Bed Biofilm Reactor for the Treatment of Wastewater: Process Optimization and Kinetic Study. Bioresour. Technol. 2019, 281, 335–342. [Google Scholar] [CrossRef]
- Li, X.; Deng, Y.; Li, X.; Ma, X.; Wang, J.; Li, J. Integration of Marine Macroalgae (Chaetomorpha maxima) with a Moving Bed Bioreactor for Nutrient Removal from Maricultural Wastewater. Archaea 2020, 2020, 8848120. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Y.; Cheng, X.; Sun, D.; Ren, Y.; Xu, G. Formation Characteristics of an Anoxygenic Photosynthetic Bacterial Biofilm in a Photorotating Biological Contactor for Azo Dye Wastewater Treatment. J. Chem. Technol. Biotechnol. 2015, 90, 176–184. [Google Scholar] [CrossRef]
- Do, T.T.; Ong, B.N.; Tran, M.L.N.; Nguyen, D.; Melkonian, M.; Tran, H.D. Biomass and Astaxanthin Productivities of Haematococcus pluvialis in an Angled Twin-Layer Porous Substrate Photobioreactor: Effect of Inoculum Density and Storage Time. Biology 2019, 8, 68. [Google Scholar] [CrossRef]
- Zhang, C.; Zhu, X.; Liao, Q.; Wang, Y.; Li, J.; Ding, Y.; Wang, H. Performance of a Groove-Type Photobioreactor for Hydrogen Production by Immobilized Photosynthetic Bacteria. Int. J. Hydrogen Energy 2010, 35, 5284–5292. [Google Scholar] [CrossRef]
- Capson-tojo, G.; Zuo, A.; Gan, M.; Ledezma, P.; Damien, J.; Hülsen, T. Resource Recovery Using Enriched Purple Phototrophic Bacteria in an Outdoor Flat Plate Photobioreactor: Suspended vs. Attached Growth. Bioresour. Technol. 2023, 373, 128709. [Google Scholar] [CrossRef] [PubMed]
- Prachanurak, P.; Chiemchaisri, C.; Chiemchaisri, W.; Yamamoto, K. Modelling of Biofilm Growth for Photosynthetic Biomass Production in a Pipe-Overflow Recirculation Bioreactor. Biochem. Eng. J. 2019, 142, 50–57. [Google Scholar] [CrossRef]
- Machineni, L. Review on Biological Wastewater Treatment and Resources Recovery: Attached and Suspended Growth Systems. Water Sci. Technol. 2019, 80, 2013–2026. [Google Scholar] [CrossRef] [PubMed]
- Hoekema, S.; Bijmans, M.; Janssen, M.; Tramper, J.; Wijffels, R.H. A Pneumatically Agitated Flat-Panel Photobioreactor with Gas Re-Circulation: Anaerobic Photoheterotrophic Cultivation of a Purple Non-Sulfur Bacterium. Int. J. Hydrogen Energy 2002, 27, 1331–1338. [Google Scholar] [CrossRef]
- Madigan, M.T.; Gest, H. Growth of the Photosynthetic Bacterium Rhodopseudomonas capsulata Chemoautotrophically in Darkness with H2 as the Energy Source. J. Bacteriol. 1979, 137, 524–530. [Google Scholar] [CrossRef]
- Ruiz, J.; Álvarez-Díaz, P.D.; Arbib, Z.; Garrido-Pérez, C.; Barragán, J.; Perales, J.A. Performance of a Flat Panel Reactor in the Continuous Culture of Microalgae in Urban Wastewater: Prediction from a Batch Experiment. Bioresour. Technol. 2013, 127, 456–463. [Google Scholar] [CrossRef]
- Skjånes, K.; Andersen, U.; Heidorn, T.; Borgvang, S.A. Design and Construction of a Photobioreactor for Hydrogen Production, Including Status in the Field. J. Appl. Phycol. 2016, 28, 2205–2223. [Google Scholar] [CrossRef]
- Ghosh, S.; Dutta, S.; Chowdhury, R. Ameliorated Hydrogen Production through Integrated Dark-Photo Fermentation in a Flat Plate Photobioreactor: Mathematical Modelling and Optimization of Energy Efficiency. Energy Convers. Manag. 2020, 226, 113549. [Google Scholar] [CrossRef]
- Iluz, D.; Abu-Ghosh, S. A Novel Photobioreactor Creating Fluctuating Light from Solar Energy for a Higher Light-to-Biomass Conversion Efficiency. Energy Convers. Manag. 2016, 126, 767–773. [Google Scholar] [CrossRef]
- Posten, C. Design Principles of Photo-Bioreactors for Cultivation of Microalgae. Eng. Life Sci. 2009, 9, 165–177. [Google Scholar] [CrossRef]
- Zhu, J.; Rong, J.; Zong, B. Factors in Mass Cultivation of Microalgae for Biodiesel. Cuihua Xuebao/Chin. J. Catal. 2013, 34, 80–100. [Google Scholar] [CrossRef]
- Vasumathi, K.K.; Premalatha, M.; Subramanian, P. Parameters Influencing the Design of Photobioreactor for the Growth of Microalgae. Renew. Sustain. Energy Rev. 2012, 16, 5443–5450. [Google Scholar] [CrossRef]
- De Vree, J.H.; Bosma, R.; Janssen, M.; Barbosa, M.J.; Wijffels, R.H. Comparison of Four Outdoor Pilot-Scale Photobioreactors. Biotechnol. Biofuels 2015, 8, 215. [Google Scholar] [CrossRef]
- Gupta, P.L.; Lee, S.M.; Choi, H.J. A Mini Review: Photobioreactors for Large Scale Algal Cultivation. World J. Microbiol. Biotechnol. 2015, 31, 1409–1417. [Google Scholar] [CrossRef] [PubMed]
- Luis, P. Fundamental Modeling of Membrane Systems: Membrane and Process Performance; Elsevier: Amsterdam, The Netherlands, 2018; ISBN 9780128134849. [Google Scholar]
- Praveen, P.; Heng, J.Y.P.; Loh, K.C. Tertiary Wastewater Treatment in Membrane Photobioreactor Using Microalgae: Comparison of Forward Osmosis & Microfiltration. Bioresour. Technol. 2016, 222, 448–457. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Le-Clech, P.; Henderson, R.K. Simultaneous Microalgae Cultivation and Wastewater Treatment in Submerged Membrane Photobioreactors: A Review. Algal Res. 2017, 24, 425–437. [Google Scholar] [CrossRef]
- Marbelia, L.; Bilad, M.R.; Passaris, I.; Discart, V.; Vandamme, D.; Beuckels, A.; Muylaert, K.; Vankelecom, I.F.J. Membrane Photobioreactors for Integrated Microalgae Cultivation and Nutrient Remediation of Membrane Bioreactors Effluent. Bioresour. Technol. 2014, 163, 228–235. [Google Scholar] [CrossRef]
- Liao, B.Q.; Allen, D.G.; Droppo, I.G.; Leppard, G.G.; Liss, S.N. Surface Properties of Sludge and Their Role in Bioflocculation and Settleability. Water Res. 2001, 35, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Alloul, A.; Wuyts, S.; Lebeer, S.; Vlaeminck, S.E. Volatile Fatty Acids Impacting Phototrophic Growth Kinetics of Purple Bacteria: Paving the Way for Protein Production on Fermented Wastewater. Water Res. 2019, 152, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Praveen, P.; Xiao, W.; Lamba, B.; Loh, K.C. Low-Retention Operation to Enhance Biomass Productivity in an Algal Membrane Photobioreactor. Algal Res. 2019, 40, 101487. [Google Scholar] [CrossRef]
- Parakh, S.K.; Praveen, P.; Loh, K.C.; Tong, Y.W. Integrating Gravity Settler with an Algal Membrane Photobioreactor for In Situ Biomass Concentration and Harvesting. Bioresour. Technol. 2020, 315, 123822. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.L.; Stadler, L.B.; Cao, L.; Love, N.G.; Raskin, L.; Skerlos, S.J. Navigating Wastewater Energy Recovery Strategies: A Life Cycle Comparison of Anaerobic Membrane Bioreactor and Conventional Treatment Systems with Anaerobic Digestion. Environ. Sci. Technol. 2014, 48, 5972–5981. [Google Scholar] [CrossRef] [PubMed]
- Ishizaki, S.; Fukushima, T.; Ishii, S.; Okabe, S. Membrane Fouling Potentials and Cellular Properties of Bacteria Isolated from Fouled Membranes in a MBR Treating Municipal Wastewater; Elsevier Ltd.: Amsterdam, The Netherlands, 2016; Volume 100, ISBN 8111706626. [Google Scholar]
- Liao, Y.; Bokhary, A.; Maleki, E.; Liao, B. A Review of Membrane Fouling and Its Control in Algal-Related Membrane Processes. Bioresour. Technol. 2018, 264, 343–358. [Google Scholar] [CrossRef]
- Lu, H.; Zhang, G.; Dai, X.; Schideman, L.; Zhang, Y.; Li, B.; Wang, H. A Novel Wastewater Treatment and Biomass Cultivation System Combining Photosynthetic Bacteria and Membrane Bioreactor Technology. Desalination 2013, 322, 176–181. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, H.; Zhang, Z.; Jiao, Y.; Zhang, Q. Effects of Mass Transfer and Light Intensity on Substrate Biological Degradation by Immobilized Photosynthetic Bacteria within an Annular Fiber-Illuminating Biofilm Reactor. J. Photochem. Photobiol. B 2014, 131, 113–119. [Google Scholar] [CrossRef]
- Tekucheva, D.N.; Laurinavichene, T.V.; Seibert, M.; Tsygankov, A.A. Immobilized Purple Bacteria for Light-Driven H 2 Production from Starch and Potato Fermentation Effluents. Biotechnol. Prog. 2011, 27, 1248–1256. [Google Scholar] [CrossRef]
- Nhi-Cong, L.T.; Lien, D.T.; Gupta, B.S.; Mai, C.T.N.; Ha, H.P.; Nguyet, N.T.M.; Duan, T.H.; Van Quyen, D.; Zaid, H.F.M.; Sankaran, R.; et al. Enhanced Degradation of Diesel Oil by Using Biofilms Formed by Indigenous Purple Photosynthetic Bacteria from Oil-Contaminated Coasts of Vietnam on Different Carriers. Appl. Biochem. Biotechnol. 2019, 191, 313–330. [Google Scholar] [CrossRef]
- Berillo, D.; Al-Jwaid, A.; Caplin, J. Polymeric Materials Used for Immobilisation of Bacteria for the Bioremediation of Contaminants in Water. Polymers 2021, 13, 1073. [Google Scholar] [CrossRef]
- Mehrotra, T.; Dev, S.; Banerjee, A.; Chatterjee, A.; Singh, R.; Aggarwal, S. Use of Immobilized Bacteria for Environmental Bioremediation: A Review. J. Environ. Chem. Eng. 2021, 9, 105920. [Google Scholar] [CrossRef]
- Skouteris, G.; Rodriguez-Garcia, G.; Reinecke, S.F.; Hampel, U. The Use of Pure Oxygen for Aeration in Aerobic Wastewater Treatment: A Review of Its Potential and Limitations. Bioresour. Technol. 2020, 312, 123595. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tay, J.H. The Essential Role of Hydrodynamic Shear Force in the Formation of Biofilm and Granular Sludge. Water Res. 2002, 36, 1653–1665. [Google Scholar] [CrossRef] [PubMed]
- Van der Star, W.R.L.; Abma, W.R.; Blommers, D.; Mulder, J.W.; Tokutomi, T.; Strous, M.; Picioreanu, C.; van Loosdrecht, M.C.M. Startup of Reactors for Anoxic Ammonium Oxidation: Experiences from the First Full-Scale Anammox Reactor in Rotterdam. Water Res. 2007, 41, 4149–4163. [Google Scholar] [CrossRef] [PubMed]
- Pat-Espadas, A.M.; Field, J.A.; Razo-Flores, E.; Cervantes, F.J.; Sierra-Alvarez, R. Continuous Removal and Recovery of Palladium in an Upflow Anaerobic Granular Sludge Bed (UASB) Reactor. J. Chem. Technol. Biotechnol. 2016, 91, 1183–1189. [Google Scholar] [CrossRef]
- Manirafasha, E.; Jiao, K.; Zeng, X.; Xu, Y.; Tang, X.; Sun, Y.; Lin, L.; Murwanashyaka, T.; Ndikubwimana, T.; Jing, K.; et al. Processing of Microalgae to Biofuels. In Microalgae Cultivation for Biofuels Production; Academic Press: Cambridge, MA, USA, 2019; pp. 111–128. [Google Scholar] [CrossRef]
- O’Flaherty, V.; Lens, P.N.L.; De Beer, D.; Colleran, E. Effect of Feed Composition and Upflow Velocity on Aggregate Characteristics in Anaerobic Upflow Reactors. Appl. Microbiol. Biotechnol. 1997, 47, 102–107. [Google Scholar] [CrossRef]
- Beun, J.J.; Hendriks, A.; Van Loosdrecht, M.C.M.; Morgenroth, E.; Wilderer, P.A.; Heijnen, J.J. Aerobic Granulation in a Sequencing Batch Reactor. Water Res. 1999, 33, 2283–2290. [Google Scholar] [CrossRef]
- Van der Last, A.R.M.; Lettinga, G. Anaerobic Treatment of Domestic Sewage under Moderate Climatic (Dutch) Conditions Using Upflow Reactors at Increased Superficial Velocities. Water Sci. Technol. 1992, 25, 167–178. [Google Scholar] [CrossRef]
- Sawayama, S.; Tsukahara, K.; Yagishita, T.; Hanada, S. Characterization of Lighted Upflow Anaerobic Sludge Blanket (LUASB) Method under Sulfate-Rich Conditions. J. Biosci. Bioeng. 2001, 91, 195–201. [Google Scholar] [CrossRef]
- Lv, L.; Li, W.; Wu, C.; Meng, L.; Qin, W. Microbial Community Composition and Function in a Pilot-Scale Anaerobic-Anoxic-Aerobic Combined Process for the Treatment of Traditional Chinese Medicine Wastewater. Bioresour. Technol. 2017, 240, 84–93. [Google Scholar] [CrossRef]
- Cerruti, M.; Stevens, B.; Ebrahimi, S.; Alloul, A.; Vlaeminck, S.E.; Weissbrodt, D.G. Enrichment and Aggregation of Purple Non-Sulfur Bacteria in a Mixed-Culture Sequencing-Batch Photobioreactor for Biological Nutrient Removal from Wastewater. Front. Bioeng. Biotechnol. 2020, 8, 557234. [Google Scholar] [CrossRef]
- Stegman, S.; Batstone, D.J.; Rozendal, R.; Jensen, P.D.; Hülsen, T. Purple Phototrophic Bacteria Granules under High and Low Upflow Velocities. Water Res. 2021, 190, 116760. [Google Scholar] [CrossRef] [PubMed]
- Blansaer, N.; Alloul, A.; Verstraete, W.; Vlaeminck, S.E.; Smets, B.F. Aggregation of Purple Bacteria in an Upflow Photobioreactor to Facilitate Solid/Liquid Separation: Impact of Organic Loading Rate, Hydraulic Retention Time and Water Composition. Bioresour. Technol. 2022, 348, 126806. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.C.; He, P.J.; Wang, G.Z.; Yu, G.H.; Shao, L.M. Enhanced Storage Stability of Aerobic Granules Seeded with Pellets. Bioresour. Technol. 2010, 101, 8031–8037. [Google Scholar] [CrossRef]
- Wang, L.; Liu, X.; Lee, D.J.; Tay, J.H.; Zhang, Y.; Wan, C.L.; Chen, X.F. Recent Advances on Biosorption by Aerobic Granular Sludge. J. Hazard. Mater. 2018, 357, 253–270. [Google Scholar] [CrossRef]
- Safwat, S.M. Moving Bed Biofilm Reactors for Wastewater Treatment: A Review of Basic Concepts International Journal of Research Moving Bed Biofilm Reactors for Wastewater Treatment: A Review of Basic Concepts. Int. J. Res. 2019, 6, 85–90. [Google Scholar]
- Ashrafi, E.; Allahyari, E.; Torresi, E.; Andersen, H.R. Effect of Slow Biodegradable Substrate Addition on Biofilm Structure and Reactor Performance in Two MBBRs Filled with Different Support Media. Environ. Technol. 2020, 41, 2750–2759. [Google Scholar] [CrossRef]
- Martínez-Huerta, G.; Prendes-Gero, B.; Ortega-Fernández, F.; Mesa-Fernández, J.M. Design of a Carrier for Wastewater Treatment Using Moving Bed Bioreactor. In Proceedings of the 2nd International Conference on Environmental and Geological Science and Engineering, EG ’09, Brasov, Romania, 24–26 September 2009; pp. 44–49. [Google Scholar]
- Aldris, B.; Farhoud, N. Wastewater Treatment Efficiency of an Experimental MBBR System under Different Influent Concentrations. DYSONA-Appl. Sci. 2020, 1, 20–28. [Google Scholar] [CrossRef]
- Shaikh, S.; Rashid, N.; Onwusogh, U.; McKay, G.; Mackey, H.R. Effect of Nutrients Deficiency on Biofilm Formation and Single Cell Protein Production with a Purple Non-Sulphur Bacteria Enriched Culture. Biofilms 2023, 5, 100098. [Google Scholar] [CrossRef]
- Shaikh, S.; Rashid, N.; McKay, G.; Liberski, A.R.; Mackey, H.R. Nitrogen Influence on Suspended vs Biofilm Growth and Resource Recovery Potential of Purple Non-Sulfur Bacteria Treating Fuel Synthesis Wastewater. Biochem. Eng. J. 2023, 190, 108754. [Google Scholar] [CrossRef]
- De Silva, J.K.A.; Karunaratne, A.K.; Sumanasinghe, V.A. Wastewater Treatment Using Attached Growth Microbial Biofilms on Coconut Fiber: A Short Review. J. Agric. Value Addit. 2019, 2, 61–70. [Google Scholar]
- Li, T.; Strous, M.; Melkonian, M. Biofilm-Based Photobioreactors: Their Design and Improving Productivity through Efficient Supply of Dissolved Inorganic Carbon. FEMS Microbiol. Lett. 2017, 364, fnx218. [Google Scholar] [CrossRef] [PubMed]
- Pragya, N.; Pandey, K.K.; Sahoo, P.K. A Review on Harvesting, Oil Extraction and Biofuels Production Technologies from Microalgae. Renew. Sustain. Energy Rev. 2013, 24, 159–171. [Google Scholar] [CrossRef]
- Singh, G.; Patidar, S.K. Microalgae Harvesting Techniques: A Review. J. Environ. Manag. 2018, 217, 499–508. [Google Scholar] [CrossRef]
- Hülsen, T.; Sander, E.M.; Jensen, P.D.; Batstone, D.J. Application of Purple Phototrophic Bacteria in a Biofilm Photobioreactor for Single Cell Protein Production: Biofilm vs Suspended Growth. Water Res. 2020, 181, 115909. [Google Scholar] [CrossRef] [PubMed]
- Blanken, W.; Janssen, M.; Cuaresma, M.; Libor, Z.; Bhaiji, T.; Wijffels, R.H. Biofilm Growth of Chlorella Sorokiniana in a Rotating Biological Contactor Based Photobioreactor. Biotechnol. Bioeng. 2014, 111, 2436–2445. [Google Scholar] [CrossRef]
- Cortez, S.; Teixeira, P.; Oliveira, R.; Mota, M. Rotating Biological Contactors: A Review on Main Factors Affecting Performance. Rev. Environ. Sci. Biotechnol. 2008, 7, 155–172. [Google Scholar] [CrossRef]
- Sawada, H.; Rogers, P.L. Photosynthetic Bacteria in Waste Treatment—Mixed Culture Studies with Rhodopseudomonas capsulata. J. Ferment. Technol. 1977, 55, 311–325. [Google Scholar]
- Wang, X.; Cheng, X.; Sun, D.; Ren, Y.; Xu, G. Simultaneous Nutrient and Carbon Removal from Azo Dye Wastewater Using a Photorotating Biological Contactor Reactor. J. Chem. Technol. Biotechnol. 2014, 89, 1545–1552. [Google Scholar] [CrossRef]
- Naumann, T.; Çebi, Z.; Podola, B.; Melkonian, M. Growing Microalgae as Aquaculture Feeds on Twin-Layers: A Novel Solid-State Photobioreactor. J. Appl. Phycol. 2013, 25, 1413–1420. [Google Scholar] [CrossRef]
- Podola, B.; Li, T.; Melkonian, M. Porous Substrate Bioreactors: A Paradigm Shift in Microalgal Biotechnology? Trends Biotechnol. 2017, 35, 121–132. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, L.; Wang, J.; Chen, Y.; Gao, X.; Zhang, Z.; Liu, T. Attached Cultivation for Improving the Biomass Productivity of Spirulina Platensis. Bioresour. Technol. 2015, 181, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Wang, J.; Hu, Q.; Cheng, P.; Ji, B.; Liu, J.; Chen, Y.; Zhang, W.; Chen, X.; Chen, L.; et al. Attached Cultivation Technology of Microalgae for Efficient Biomass Feedstock Production. Bioresour. Technol. 2013, 127, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Podola, B.; Melkonian, M. Investigating Dynamic Processes in a Porous Substrate Biofilm Photobioreactor—A Modeling Approach. Algal Res. 2016, 13, 30–40. [Google Scholar] [CrossRef]
- Huang, Q.; Jiang, F.; Wang, L.; Yang, C. Design of Photobioreactors for Mass Cultivation of Photosynthetic Organisms. Engineering 2017, 3, 318–329. [Google Scholar] [CrossRef]
- Zhang, C.; Ma, S.; Wang, G.; Guo, Y.P. Enhancing Continuous Hydrogen Production by Photosynthetic Bacterial Biofilm Formation within an Alveolar Panel Photobioreactor. Int. J. Hydrogen Energy 2019, 44, 27248–27258. [Google Scholar] [CrossRef]
- Huang, Y.; Zheng, Y.; Li, J.; Liao, Q.; Fu, Q.; Xia, A.; Fu, J.; Sun, Y. Enhancing Microalgae Biofilm Formation and Growth by Fabricating Microgrooves onto the Substrate Surface. Bioresour. Technol. 2018, 261, 36–43. [Google Scholar] [CrossRef]
- Chen, X.; Stewart, P.S. Biofilm Removal Caused by Chemical Treatments. Water Res. 2000, 34, 4229–4233. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aspect | Characteristics |
|---|---|
| Typical habitats | Organic polluted water (waste lagoons, sewage), soil, acidic and alkali springs, soda lakes |
| Photosynthetic system | Intracytoplasmic membrane |
| Metabolism | Photoheterotrophy (primary), photoautotroph, chemoheterotrophy |
| Types of bacteriochlorophylls | Bacteriochlorophylls (BChls) a and b |
| Form and motility | Motile, rod, spherical |
| Carbon source | Organic carbon, especially short-chain volatile fatty acids |
| Sulfur utilization | Low concentration of sulfur |
| Light–oxygen demand | Light–anaerobic, Light–microaerobic, Light–aerobic, Dark–aerobic, Dark–anaerobic |
| Preferred electron donor for phototrophic growth | Organic compounds, H2, sulfide, ferrous iron |
| Salinity tolerance | 0–3% |
| Typical bacterial genera | Rhodopseudomonas, Rhodobacter, Rhodospirillum |
| Property | Homopolymer (PHAs) | Copolymer (PHAs) | Polypropylene | ||
|---|---|---|---|---|---|
| - | Scl | Mcl | P(3HB-co-3HV) | P(3HB-co-6%3HD) | - |
| Tensile strength (Mpa) | 5 | 20 | up to 690 | 17 | 400 |
| Young’s modulus (Gpa) | 3.5 | - | 0.7–2.9 | - | 1.7 |
| Elongation to break (%) | 40 | 300 | 30–38 | 680 | 38 |
| Melting temperature (°C) | 179 | 80 | 137–170 | 130 | 176 |
| Glass transition temperature (°C) | 4 | −40 | 10 to −6 | −8 | −10 |
| PBR | Purpose | Strain | Illumination | Scale | References |
|---|---|---|---|---|---|
| Flat-panel (plate) PBR | Hydrogen production | Rhodobacter sphaeroides (O.U. 001) | Artificial | Laboratory-scale | [89] |
| Hydrogen and PHB production | Rhodobacter sphaeroides (O.U. 001) | Solar | Laboratory-scale | [90] | |
| Tubular PBR | PHB production | Rhodopseudomonas sphaeroides S16-VOGS3 | Solar | Laboratory-scale | [45] |
| PHB production | Rhodopseudomonas palustris 42OL | Solar | Laboratory-scale | [91] | |
| Biomass production | Rhodopseudomonas palustris | Artificial/Solar | Laboratory-scale | [92] | |
| Hydrogen production | Rhodopseudomonas palustris | Solar | Laboratory-scale | [93] | |
| Hydrogen production | Rhodobacter capsulatus | Solar | Pilot-scale | [81] | |
| Membrane PBR | Wastewater treatment | Microalgae (Chlorella vulgaris) | Artificial | Laboratory-scale | [94] |
| Wastewater treatment | Rhodopseudomonas palustris | Artificial | Laboratory-scale | [95] | |
| Up-flow anaerobic sludge blanket (UASB) PBR | Wastewater treatment and PHB production | Mixed culture of phototrophic bacteria | Artificial | Laboratory-scale | [96] |
| Artificial immobilization system | Hydrogen production (gel) | Rhodopseudomonas palustris CQK 01 | Artificial | Laboratory-scale | [97] |
| Hydrogen production (agar) | Rhodobacter capsulatus (DSM 1710 wild-type strain) and YO3 (Hup-mutant of MT1131 strain) | Artificial | Laboratory-scale | [98] | |
| Hydrogen production (silica gel, clay and activated carbon) | Rhodopseudomonas palustris WP3-5 | Artificial | Laboratory-scale | [73] | |
| Moving-bed Biofilm PBR | Wastewater treatment | NA 1 | NA | Laboratory-scale | [99] |
| Wastewater treatment | Marine macroalgae | Artificial | Laboratory-scale | [100] | |
| Photo-rotating biological contactor | Wastewater treatment | Anoxygenic photosynthetic bacteria | Artificial | Laboratory-scale | [101] |
| Porous-substrate PBR | Astaxanthin production | Microalga Haematococcus pluvialis | Artificial | Laboratory-scale | [102] |
| Grooved-surface PBR | Hydrogen production | Rhodopseudomonas palustris CQK 01 | Artificial | Laboratory-scale | [103] |
| Flat-panel (plate) biofilm PBR | Protein production | Mixed culture of purple bacteria | Solar | Pilot-scale | [104] |
| Pipe-overflow recirculation biofilm PBR | Verification of mathematical model for substrate consumption and biofilm productivity | Mixed culture of photosynthetic bacteria | Artificial | Laboratory-scale | [105] |
| Suspended PBR | Advantages | Relevance to PNSB-PHA Production | Disadvantages | Relevance to PNSB-PHA Production |
|---|---|---|---|---|
| Flat-panel |
|
|
|
|
| Tubular |
|
|
|
|
| Membrane |
|
|
|
|
| Immobilized PBRs System | Advantages | Relevance to PNSB-PHA Production | Disadvantages | Relevance to PNSB-PHA Production |
|---|---|---|---|---|
| Artificial immobilization on or within suspended carriers |
|
|
|
|
| Self-immobilized granular systems |
|
|
|
|
| MBBPR |
|
|
|
|
| Biofilm System | Advantages | Relevance to PNSB-PHA Production | Disadvantages | Relevance to PNSB-PHA Production |
|---|---|---|---|---|
| PRBC |
|
|
|
|
| PSBR |
|
|
|
|
| Grooved |
|
|
|
|
| System | Illumination | Control of Nutrients | Mixing | Temperature Control | Biomass Production | Biomass Harvesting | Overall |
|---|---|---|---|---|---|---|---|
| Flat-panel PBR | 4 | 4 | 4 | 4 | 4 | 4 | 4/5 |
| Tubular PBR | 4 | 4 | 4 | 4 | 4 | 3 | 3.8/5 |
| Membrane PBR | 4 | 4 | 4 | 4 | 4 | 3 | 3.8/5 |
| Artificial immobilization on or within suspended carriers | 3 | 4 | 4 | 4 | 4 | 2 | 3.5/5 |
| Self-immobilized granular system | 4 | 4 | 4 | 4 | 5 | 4 | 4.2/5 |
| Moving-bed biofilm photobioreactor (MBBPR) | 4 | 4 | 4 | 4 | 4 | 2 | 3.6/5 |
| Photo-rotating biological contactor (PRBC) | 3 | 4 | 4 | 4 | 3 | 3 | 3.5/5 |
| Porous-substrate photobioreactor (PSBR) | 4 | 4 | 4 | 2 | 3 | 3 | 3.3/5 |
| Grooved-surface PBR | 5 | 4 | 4 | 4 | 3 | 2 | 3.3/5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shaikh, S.; Rashid, N.; McKay, G.; Mackey, H.R. Photobioreactor Design for Polyhydroxyalkanoate Production Using Anoxygenic Photoheterotrophs: A Review. Fermentation 2023, 9, 778. https://doi.org/10.3390/fermentation9080778
Shaikh S, Rashid N, McKay G, Mackey HR. Photobioreactor Design for Polyhydroxyalkanoate Production Using Anoxygenic Photoheterotrophs: A Review. Fermentation. 2023; 9(8):778. https://doi.org/10.3390/fermentation9080778
Chicago/Turabian StyleShaikh, Sultan, Naim Rashid, Gordon McKay, and Hamish Robert Mackey. 2023. "Photobioreactor Design for Polyhydroxyalkanoate Production Using Anoxygenic Photoheterotrophs: A Review" Fermentation 9, no. 8: 778. https://doi.org/10.3390/fermentation9080778