
Effect of Potato Vine and Leaf Mixed Silage to Whole Corn Crops on Rumen Fermentation and the Microbe of Fatten Angus Bulls
 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Ensiling of Three Silages
2.2. In Vitro Incubation and Degradability Measurement
2.3. Animal Pens and Diets
2.4. Sample Collection and Analysis
2.5. DNA Extraction and 16s rRNA Gene Sequencing
2.6. Gas Production Kinetics
2.7. Statistical Analysis
3. Results
3.1. In Vitro Rumen Fermentation
3.2. In Vivo Rumen Fermentation
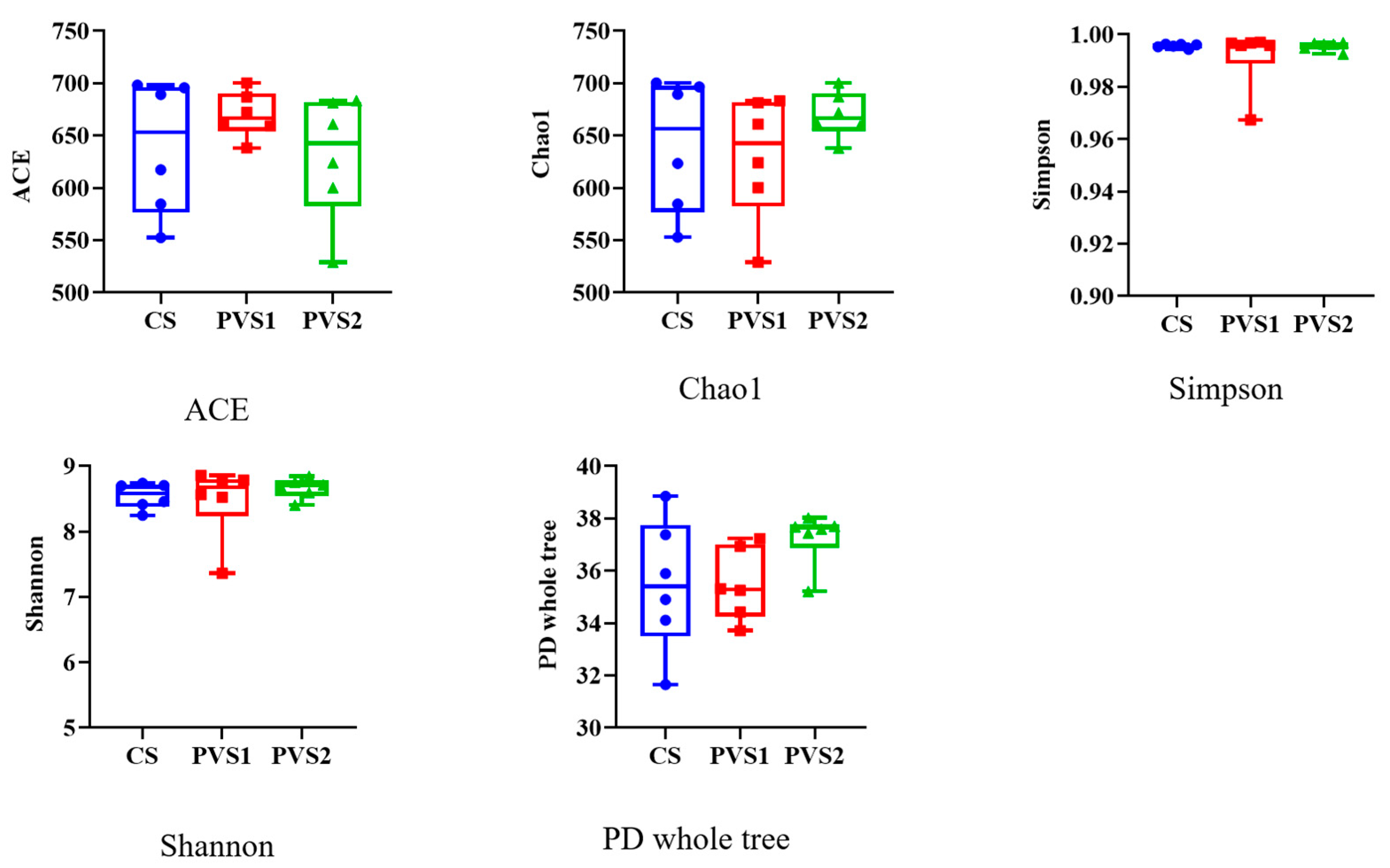
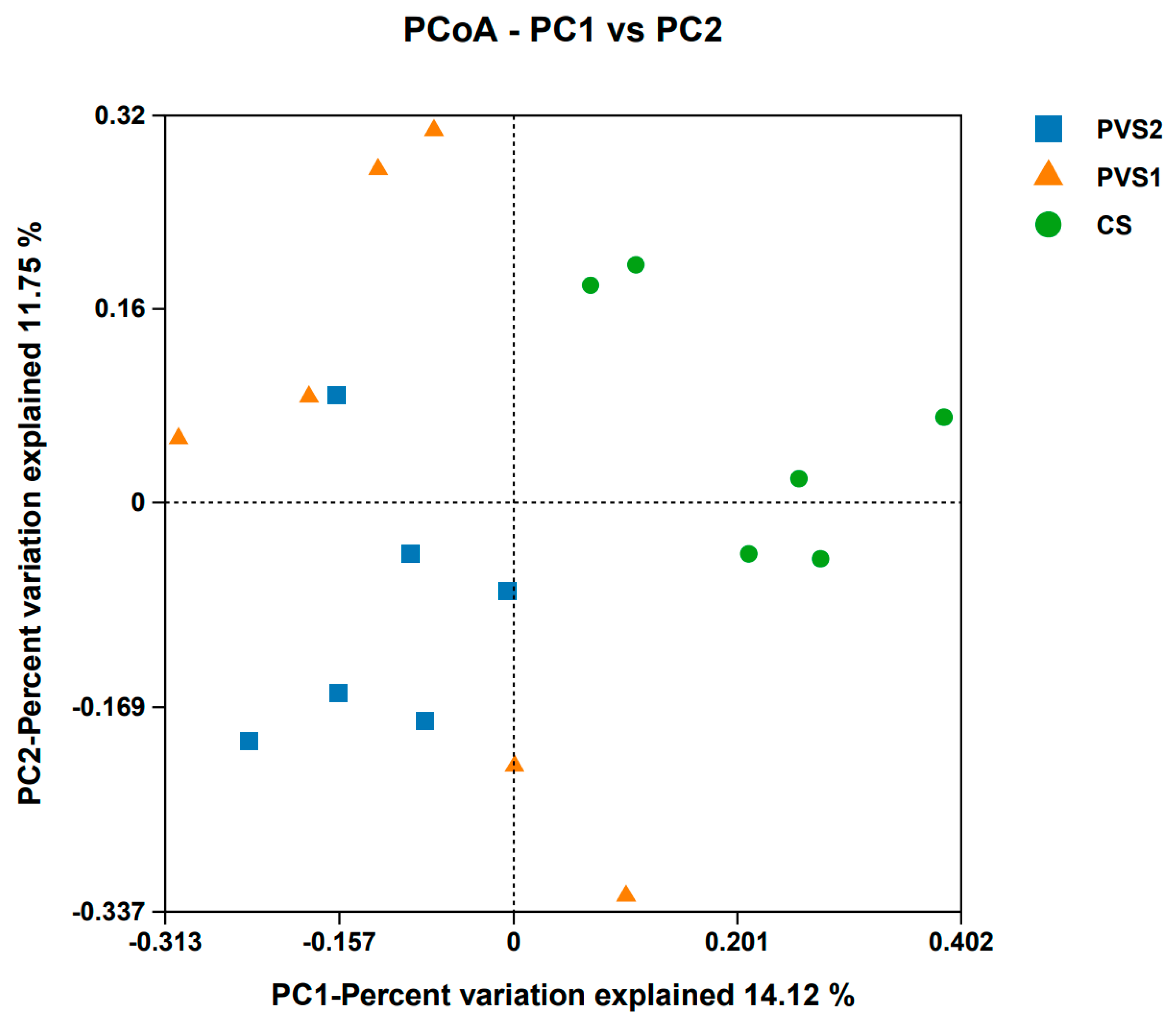
3.3. Rumen Microbial Diversities
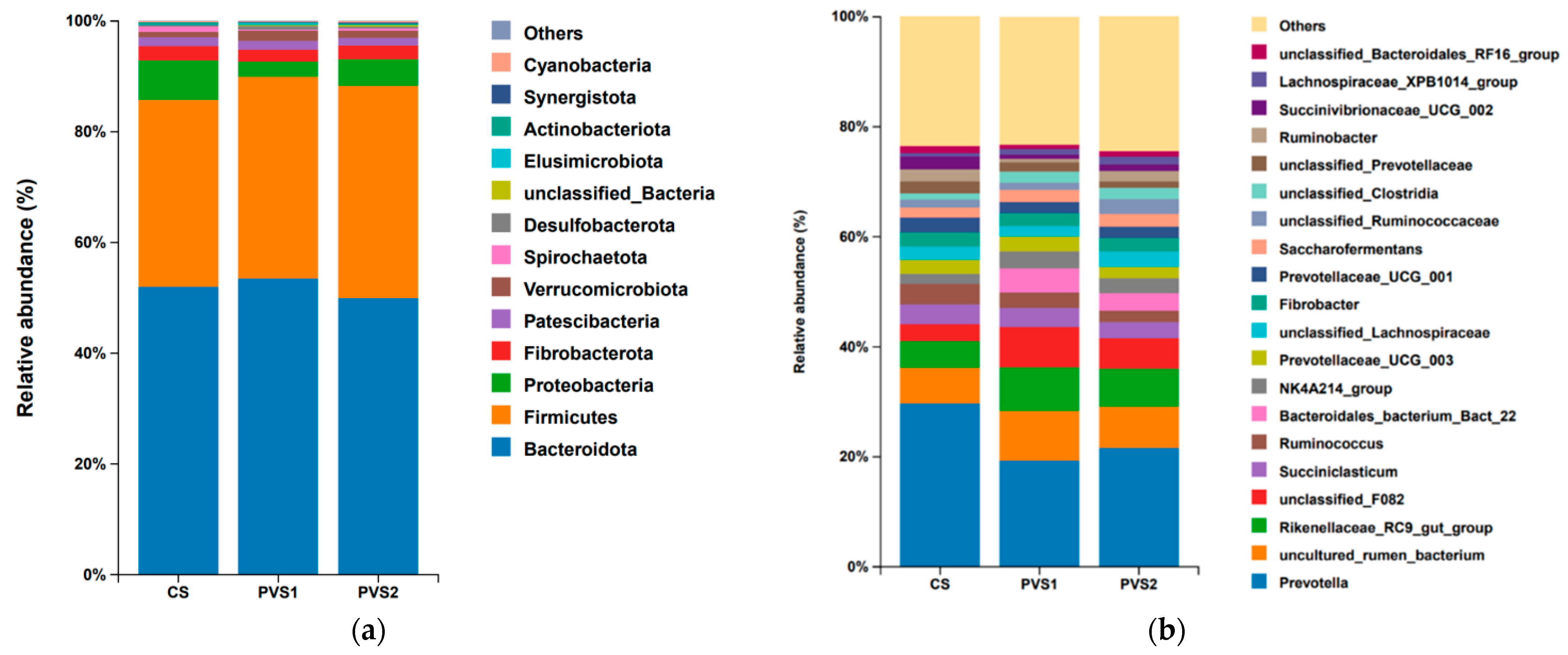
3.4. Compositions of Rumen Microbiota
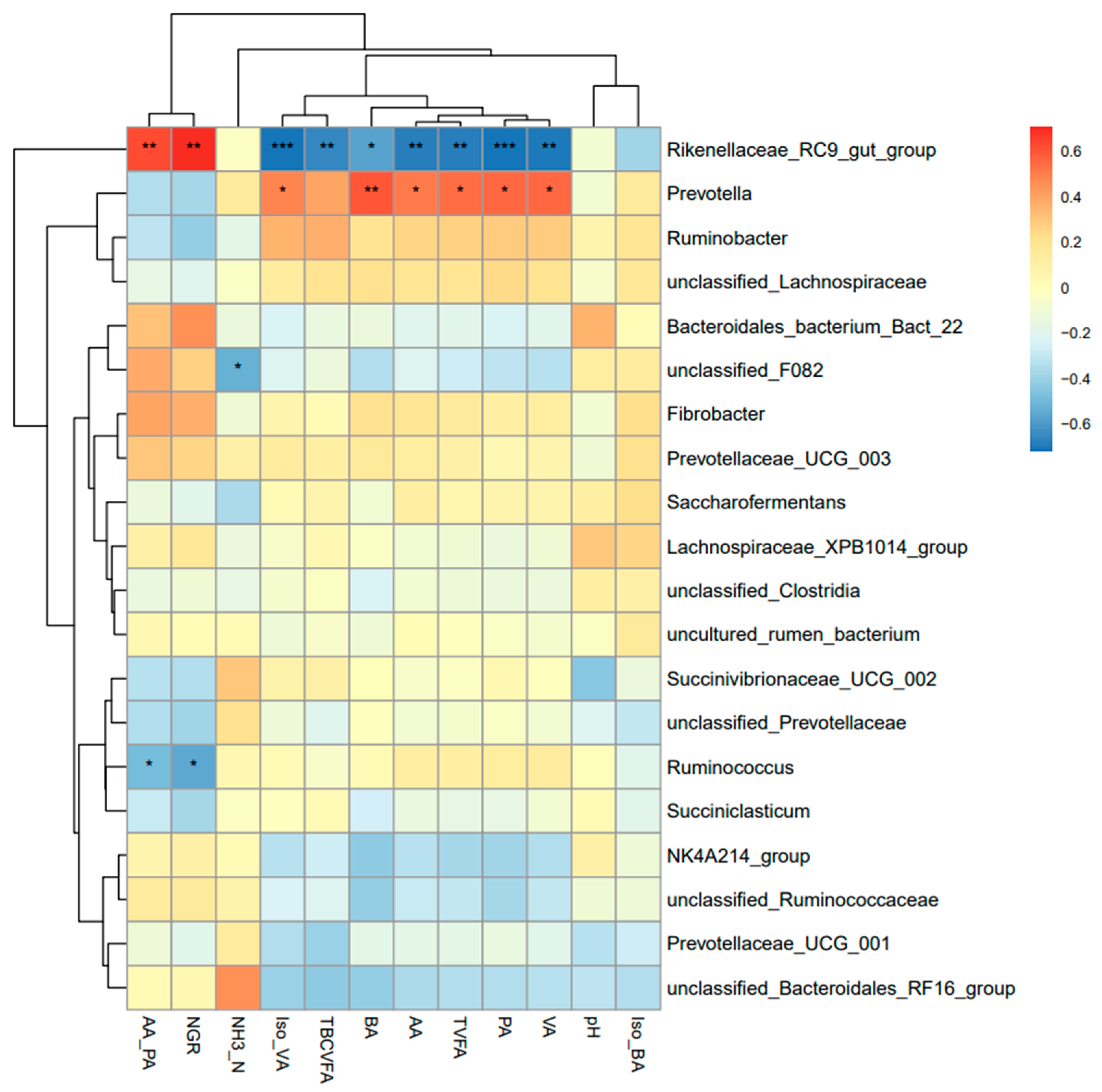
3.5. Correlation Analysis
3.6. Metabolic Functions
4. Discussion
4.1. In Vitro Rumen Fermentation Characteristic of Different Silages
4.2. Rumen Fermentation Parameter in Different Dietary Groups
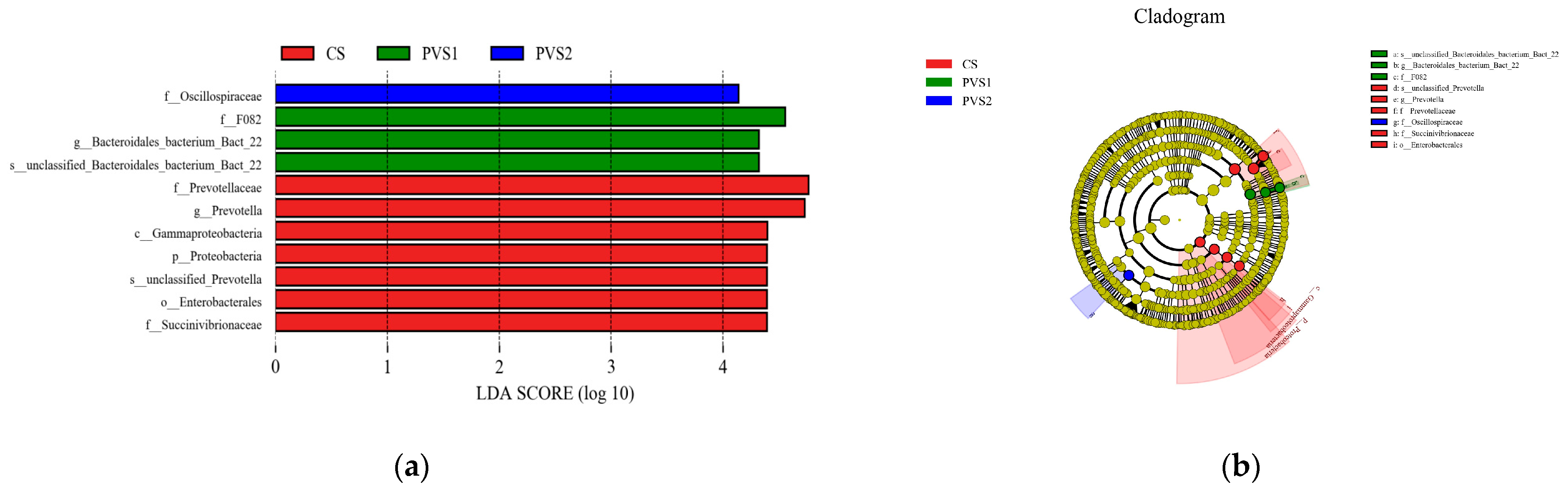
4.3. Rumen Bacteria Composition in Different Dietary Groups
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. Crops and Livestock Products. 2021. Available online: https://www.fao.org/faostat/zh/#data/QCL (accessed on 24 March 2023).
- Muck, R.E.; Weinberg, Z.G.; Rouse, D.I.; Igl, B.R. Ensiling of potato vines. Trans. ASABE 1999, 42, 565–572. [Google Scholar]
- Salehi, S.; Lashkari, S.; Abbasi, R.E.; Kamangar, H. Nutrient Digestibility and Chemical Composition of Potato (Solanum tuberosum L.) Vine as Alternative Forage in Ruminant Diets. Agric. Commun. 2014, 2, 63–66. [Google Scholar]
- Yang, Y.; Wang, C.; Cao, B. Effects of Different Additives on Silage Quality of Potato Stem and Leave. China Herbiv. Sci. 2015, 35, 34–49. [Google Scholar]
- Abong, G. A Review of Occurrence of Glycoalkaloids in Potato and Potato Products. Curr. Res. Nutr. Food Sci. 2016, 4, 95–202. [Google Scholar]
- Akiyama, R.; Watanabe, B.; Nakayasu, M.; Lee, H.J.; Kato, J.; Umemoto, N.; Muranaka, T.; Saito, K.; Sugimoto, Y.; Mizutani, M. The biosynthetic pathway of potato solanidanes diverged from that of spirosolanes due to evolution of a dioxygenase. Nat. Commun. 2021, 12, 1300. [Google Scholar] [CrossRef]
- Schrenk, D.; Bignami, M.; Bodin, L.; Chipman, J.K.; Mazo, J.D.; Hogstrand, C.; Hoogenboom, L.R.; Leblanc, J.; Nebbia, C.S.; Nielsen, E. Risk assessment of glycoalkaloids in feed and food, in particular in potatoes and potato-derived products. Efsa J. 2020, 18, e06222. [Google Scholar]
- Malecky, M.; Ghadbeigi, M.; Aliarabi, H.; Bahari, A.A.; Zaboli, K. Effect of replacing alfalfa with processed potato vines on growth performance, ruminal and total tract digestibility and blood metabolites in fattening lambs. Small Rumin. Res. 2016, 146, 13–22. [Google Scholar]
- Yupeng, H.E.; Guo, Y.; Qin, S.; Shumei, M.A.; Jinjiao, D.U.; Zheng, C.; Zhao, F. Effects of Adding Rice Bran and Wheat Bran on Silage Quality of Different Cultivars of Potato Vines. Chin. J. Anim. Nutr. 2015, 27, 3311–3318. [Google Scholar]
- Guo, Y.; Han, H.; Luo, R.; Zhao, F. Determining the ensiling characteristics of potato vine silage supplemented with rice bran and corn and evaluating their ruminal fermentation potential in vitro. N. Z. J. Agric. Res. 2018, 63, 179–188. [Google Scholar]
- Zhao, F.F.; Al-Marashdeh, O.; Zheng, C.; Guo, Y.L.; Cheng, L. Effect of adding wheat stalk and molasses to potato leaves and stems on silage chemical composition and fermentation quality. Philipp. Agric. Sci. 2019, 102, 174–179. [Google Scholar]
- Agency, I.E. Global Methane Tracker 2023. 2023. Available online: https://www.iea.org/reports/global-methane-tracker-2023 (accessed on 1 February 2023).
- Pachauri, K.; Meyer, A. Climate Change 2014. Synthesis report. Environ. Sci. Policy 2014, 27, 408. [Google Scholar]
- Luo, R.; Guo, Y.; Han, H.; Taotao, L.I.; Sui, J.; Feng, P. A Study on in Vitro Rumen Fermentation Characteristics of Potato Vines and Leaves Mixed Silage Based on Rumen Simulation Technique. Chin. J. Anim. Nutr. 2018, 30, 1185–1191. [Google Scholar]
- NRC. Nutrient Requirements of Beef Cattle, 8th ed.; National Academies Press: Washington, DC, USA, 2016. [Google Scholar]
- Pang, D.G.; Yang, H.J.; Cao, B.B.; Wu, T.T.; Wang, J.Q. The beneficial effect of Enterococcus faecium on the in vitro ruminal fermentation rate and extent of three typical total mixed rations in northern China. Livest. Sci. 2014, 167, 154–160. [Google Scholar]
- Wang, Y.-L.; Wang, W.-K.; Wu, Q.-C.; Zhang, F.; Li, W.-J.; Yang, Z.-M.; Bo, Y.-K.; Yang, H.-J. The Effect of Different Lactic Acid Bacteria Inoculants on Silage Quality, Phenolic Acid Profiles, Bacterial Community and In Vitro Rumen Fermentation Characteristic of Whole Corn Silage. Fermentation 2022, 8, 285. [Google Scholar] [CrossRef]
- Verdouw, H.; Echteld, C.; Dekkers, E. Ammonia determination based on indophenol formation with sodium salicylate. Water Res. 1978, 12, 399–402. [Google Scholar]
- Makkar, H.P.S.; Sharma, O.P.; Dawra, R.K.; Negi, S.S. Simple Determination of Microbial Protein in Rumen Liquor. J. Dairy Sci. 1982, 65, 2170–2173. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Hao, F.; Fu, H.; Tian, G.; Zhang, Y.; Fu, K.; Qi, B. N-glycosylated intestinal protein BCF-1 shapes microbial colonization by binding bacteria via its fimbrial protein. Cell Rep. 2023, 42, 111993. [Google Scholar] [CrossRef] [PubMed]
- Groot, J.C.J.; Cone, J.W.; Williams, B.A.; Debersaques, F.M.A.; Lantinga, E.A. Multiphasic analysis of gas production kinetics for in vitro fermentation of ruminant feeds. Anim. Feed Sci. Technol. 1996, 64, 77–89. [Google Scholar] [CrossRef]
- Wolin, M.J. A Theoretical Rumen Fermentation Balance. J. Dairy Sci. 1960, 43, 1452–1459. [Google Scholar] [CrossRef]
- Ramin, M.; Huhtanen, P. Development of an in vitro method for determination of methane production kinetics using a fully automated in vitro gas system—A modelling approach. Anim. Feed Sci. Technol. 2012, 174, 190–200. [Google Scholar] [CrossRef]
- Noordar, H.; Malecky, M.; Jahanian Najafabadi, H.; Navidshad, B. Evaluating nutritional value of processed potato vines by in vitro gas production. N. Z. J. Agric. Res. 2017, 60, 189–204. [Google Scholar] [CrossRef]
- Zheng, Y.; Xue, S.; Zhao, Y.; Li, S. Effect of Cassava Residue Substituting for Crushed Maize on In Vitro Ruminal Fermentation Characteristics of Dairy Cows at Mid-Lactation. Animals 2020, 10, 893. [Google Scholar]
- Yu, Q. Effect of Different Sources of Starch on RumenFermentation, Digestion and Metabolism of Nutrients in Beef Cattle; Sichuan Agricultural University: Ya’an, China, 2011. [Google Scholar]
- Friedman, M. Potato glycoalkaloids and metabolites: Roles in the plant and in the diet. J. Agric. Food Chem. 2006, 54, 8655–8681. [Google Scholar] [PubMed]
- Nicholson, J.W.G.; Young, D.A.; McQueen, R.; De Jong, H.; Wood, F.A. The Feeding Value Potential of Potato Vines. Can. J. Anim. Sci. 1978, 58, 559–569. [Google Scholar]
- Dijkstra, J.; Van Gastelen, S.; Dieho, K.; Nichols, K.; Bannink, A. Rumen sensors: Data and interpretation for key rumen metabolic processes. Animal 2020, 14 (Suppl. S1), s176–s186. [Google Scholar]
- Bergman, E.N. Glucose metabolism in ruminants as related to hypoglycemia and ketosis. Cornell Vet. 1973, 63, 341–382. [Google Scholar]
- Lemosquet, S.; Raggio, G.; Lobley, G.; Rulquin, H.; Guinard-Flament, J.; Lapierre, H. Whole-body glucose metabolism and mammary energetic nutrient metabolism in lactating dairy cows receiving digestive infusions of casein and propionic acid. J. Dairy Sci. 2009, 92, 6068–6082. [Google Scholar]
- Liu, Q.; Wang, C.; Guo, G.; Huo, W.J.; Zhang, S.L.; Pei, C.X.; Zhang, Y.L.; Wang, H. Effects of branched-chain volatile fatty acids on lactation performance and mRNA expression of genes related to fatty acid synthesis in mammary gland of dairy cows. Animal 2018, 12, 2071–2079. [Google Scholar] [CrossRef]
- Zhang, H.L.; Lin, L.I.; Chen, Y.; Xiao-Li, X.U.; Yang, Y.X. Effects of Branched Chain Amino Acids or Fatty Acid Supplementation on in Vitro Fermentation of Corn Straw and Bacterial Diversity. J. Xinjiang Agric. Univ. 2012, 35, 5. [Google Scholar]
- Feng, Y. Ruminant Nutrition; China Science Publishing & Media Ltd.: Beijing, China, 2004. [Google Scholar]
- Lin, L.; Xu, F.; Ge, X.; Li, Y. Chapter Four—Biological treatment of organic materials for energy and nutrients production—Anaerobic digestion and composting. In Advances in Bioenergy; Li, Y., Ge, X., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; Volume 4, pp. 121–181. [Google Scholar]
- Deng-Pan, B.U.; De-Xun, L.U.; Cui, W.X.; Wang, J.Q. Advance in the Synchrony of Energy and Nitrogen Release in the Rumen on the Synthesis of Microbial Protein. China Anim. Husb. Vet. Med. 2008, 35, 5–11. [Google Scholar]
- Zhang, C.; Sun, Y.; Liu, X.; Sun, G. Effects of Rumen-Protected Methionine on Ruminal Microbial Protein Production, Milk Performance and Nitrogen Excretion of Dairy Cows. Chin. J. Anim. Nutr. 2017, 29, 1759–1766. [Google Scholar]
- Matras, J.; Bartle, S.J.; Preston, R.L. Nitrogen utilization in growing lambs: Effects of grain (starch) and protein sources with various rates of ruminal degradation. J. Anim. Sci. 1991, 69, 339–347. [Google Scholar]
- Liang, Y.; Zhao, Y.; Zhang, Y. Effects of potato leaf mixed with whole corn crop silage on performance, human environment and blood biochemistry of mutton sheep. China Feed 2020, 24, 112–115. [Google Scholar] [CrossRef]
- Jami, E.; Shterzer, N.; Yosef, E.; Nikbachat, M.; Miron, J.; Mizrahi, I. Effects of including NaOH-treated corn straw as a substitute for wheat hay in the ration of lactating cows on performance, digestibility, and rumen microbial profile. J. Dairy Sci. 2014, 97, 1623–1633. [Google Scholar] [PubMed] [Green Version]
- Henderson, G.; Cox, F.; Ganesh, S.; Jonker, A.; Young, W.; Janssen, P.H. Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 2015, 5, 14567. [Google Scholar]
- Soest, P. Nutritional Ecology of the Ruminant; Cornell University Press: Ithaca, NY, USA, 1994; Volume 44, pp. 2552–2561. [Google Scholar]
- Luo, C. Effects of Dietary Crude Protein Level on Production Performance, Rumen Microflora and Metabolomics of Lactating Dairy Cows; Xinjiang Agricultural University: Urumqi, China, 2022. [Google Scholar]
- Bi, Y.; Zeng, S.; Zhang, R.; Diao, Q.; Tu, Y. Effects of dietary energy levels on rumen bacterial community composition in Holstein heifers under the same forage to concentrate ratio condition. BMC Microbiol. 2018, 18, 69. [Google Scholar] [CrossRef]
- Zhang, R.; Zhu, W.; Zhu, W.; Liu, J.; Mao, S. Effect of dietary forage sources on rumen microbiota, rumen fermentation and biogenic amines in dairy cows. J. Sci. Food Agric. 2014, 94, 1886–1895. [Google Scholar] [CrossRef]
- Singh, K.M.; Jisha, T.K.; Reddy, B.; Parmar, N.; Patel, A.; Patel, A.K.; Joshi, C.G. Microbial profiles of liquid and solid fraction associated biomaterial in buffalo rumen fed green and dry roughage diets by tagged 16S rRNA gene pyrosequencing. Mol. Biol. Rep. 2014, 42, 95–103. [Google Scholar]
- Pope, P.B.; Mackenzie, A.K.; Gregor, I.; Smith, W.; Sundset, M.A.; Mchardy, A.C.; Morrison, M.; Eijsink, V.G.H. Metagenomics of the Svalbard Reindeer Rumen Microbiome Reveals Abundance of Polysaccharide Utilization Loci. PLoS ONE 2012, 7, e38571. [Google Scholar]
- Naas, A.E.; Mackenzie, A.K.; Mravec, J.; Schückel, J.; Willats, W.G.; Eijsink, V.G.; Pope, P.B. Do rumen Bacteroidetes utilize an alternative mechanism for cellulose degradation? Mbio 2014, 5, e01401. [Google Scholar] [CrossRef] [Green Version]
- Broderick, G.A. Altering ruminal nitrogen metabolism to improve protein utilization. Introduction. J. Nutr. 1996, 126 (Suppl. S4), 1324s–1325s. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, W.; He, Y.; Wang, H.; Xia, C.; Shi, H.; Cao, B.; Su, H. Effects of Leymus chinensis replacement with whole-crop wheat hay on blood parameters, fatty acid composition, and microbiomes of Holstein bulls. J. Dairy Sci. 2018, 101, 246–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, E. The Family Prevotellaceae. In The Prokaryotes: Other Major Lineages of Bacteria and the Archaea; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 825–827. [Google Scholar]
- Bekele, A.Z.; Koike, S.; Kobayashi, Y. Genetic diversity and diet specificity of ruminal Prevotella revealed by 16S rRNA gene-based analysis. FEMS Microbiol. Lett. 2010, 305, 49–57. [Google Scholar] [CrossRef]
- Liu, H.; Xu, T.; Xu, S.; Ma, L.; Zhao, X. Effect of dietary concentrate to forage ratio on growth performance, rumen fermentation and bacterial diversity of Tibetan sheep under barn feeding on Qinghai-Tibetan plateau. PeerJ 2019, 7, e7462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.I.; Chunlei, L.I.; Zhao, H.; Zhang, Y. Rumen Degradation Characteristics and Intestinal Digestibility of Whole Corn Silages from Different Areas. Chin. J. Anim. Nutr. 2015, 27, 1641–1649. [Google Scholar]
- Dehority, B.A. Rumen Microbiology; Nottingham University: Nottingham, UK, 2003. [Google Scholar]
- Park, T.; Cersosimo, L.M.; Li, W.; Radloff, W.; Zanton, G.I. Pre-weaning Ruminal Administration of Differentially-Enriched, Rumen-Derived Inocula Shaped Rumen Bacterial Communities and Co-occurrence Networks of Post-weaned Dairy Calves. Front. Microbiol. 2021, 12, 625488. [Google Scholar] [CrossRef]
- Ortigues, I.; Doreau, M. Responses of the splanchnic tissues of ruminants to changes in intake: Absorption of digestion end products, tissue mass, metabolic activity and implications to whole animal energy metabolism. Ann. Zootech. 1995, 44, 321–346. [Google Scholar]
- Kirat, D.; Matsuda, Y.; Yamashiki, N.; Hayashi, H.; Kato, S. Expression, cellular localization, and functional role of monocarboxylate transporter 4 (MCT4) in the gastrointestinal tract of ruminants. Gene 2007, 391, 140–149. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, Z.; Hu, R.; Wang, X.; Li, F.; Zhang, X.; Zou, H.; Peng, Q.; Xue, B.; Wang, L. Comparative study of the bacterial communities throughout the gastrointestinal tract in two beef cattle breeds. Appl. Microbiol. Biotechnol. 2021, 105, 313–325. [Google Scholar] [CrossRef]
- Lv, W.; Liu, X.; Sha, Y.; Shi, H.; Wei, H.; Luo, Y.; Wang, J.; Li, S.; Hu, J.; Guo, X.; et al. Rumen Fermentation-Microbiota-Host Gene Expression Interactions to Reveal the Adaptability of Tibetan Sheep in Different Periods. Animals 2021, 11, 3529. [Google Scholar] [CrossRef]
- Zened, A.; Combes, S.; Cauquil, L.; Mariette, J.; Klopp, C.; Bouchez, O.; Troegeler-Meynadier, A.; Enjalbert, F. Microbial ecology of the rumen evaluated by 454 GS FLX pyrosequencing is affected by starch and oil supplementation of diets. FEMS Microbiol. Ecol. 2013, 83, 504–514. [Google Scholar] [CrossRef] [Green Version]
- Sha, Y.; Hu, J.; Shi, B.; Dingkao, R.; Wang, J.; Li, S.; Zhang, W.; Luo, Y.; Liu, X. Characteristics and Functions of the Rumen Microbial Community of Cattle-Yak at Different Ages. Biomed. Res. Int. 2020, 2020, 3482692. [Google Scholar] [CrossRef] [PubMed]
- van Gylswyk, N.O. Succiniclasticum ruminis gen. nov., sp. nov., a ruminal bacterium converting succinate to propionate as the sole energy-yielding mechanism. Int. J. Syst. Bacteriol. 1995, 45, 297–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Wang, H.; Li, C. Response of sheep rumen fermentation and microbial communities to feed infected with the endophyte Epichloë gansuensis as evaluated with rumen-simulating technology. J. Microbiol. 2021, 59, 718–728. [Google Scholar] [CrossRef]
- Amaretti, A.; Gozzoli, C.; Simone, M.; Raimondi, S.; Righini, L.; Pérez-Brocal, V.; García-López, R.; Moya, A.; Rossi, M. Profiling of Protein Degraders in Cultures of Human Gut Microbiota. Front. Microbiol. 2019, 10, 2614. [Google Scholar] [CrossRef] [PubMed]
- NCBI. LOC102595087 Mucin-17 [Solanum Tuberosum (Potato)]. 2019. Available online: https://www.ncbi.nlm.nih.gov/gene/102595087 (accessed on 3 June 2019).
- Macfarlane, G.T.; Gibson, G.R. Carbohydrate Fermentation, Energy Transduction and Gas Metabolism in the Human Large Intestine. In Gastrointestinal Microbiology; Springer: Boston, MA, USA, 1997; pp. 269–318. [Google Scholar]
- Svartström, O.; Alneberg, J.; Terrapon, N.; Lombard, V.; de Bruijn, I.; Malmsten, J.; Dalin, A.-M.; El Muller, E.; Shah, P.; Wilmes, P.; et al. Ninety-nine de novo assembled genomes from the moose (Alces alces) rumen microbiome provide new insights into microbial plant biomass degradation. ISME J. 2017, 11, 2538–2551. [Google Scholar] [CrossRef]
- Liu, J.; Pu, Y.Y.; Xie, Q.; Wang, J.K.; Liu, J.X. Pectin induces an in vitro rumen microbial population shift attributed to the pectinolytic Treponema group. Curr. Microbiol. 2015, 70, 67–74. [Google Scholar] [CrossRef]
- Allison, M.J.; Mayberry, W.R.; McSweeney, C.S.; Stahl, D.A. Synergistes jonesii, gen. nov., sp.nov.: A Rumen Bacterium That Degrades Toxic Pyridinediols. Syst. Appl. Microbiol. 1992, 15, 522–529. [Google Scholar]
- Wang, X.; Lei, Z.; Shimizu, K.; Zhang, Z.; Lee, D.J. Improved methane production from corn straw using anaerobically digested sludge pre-augmented by nanobubble water. Bioresour. Technol. 2020, 311, 123479. [Google Scholar] [CrossRef]
- Wang, X.; Li, F.; Zhang, N.; Ungerfeld, E.; Guo, L.; Zhang, X.; Wang, M.; Ma, Z. Effects of supplementing a yeast culture in a pelleted total mixed ration on fiber degradation, fermentation parameters, and the bacterial community in the rumen of sheep. Anim. Feed Sci. Technol. 2023, 296, 115565. [Google Scholar] [CrossRef]
- Gharechahi, J.; Vahidi, M.F.; Bahram, M.; Han, J.L.; Ding, X.Z.; Salekdeh, G.H. Metagenomic analysis reveals a dynamic microbiome with diversified adaptive functions to utilize high lignocellulosic forages in the cattle rumen. ISME J. 2021, 15, 1108–1120. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item 1 | CS | PVS1 | PVS2 |
|---|---|---|---|
| pH-value | 4.48 | 4.69 | 4.87 |
| Nutritive values, % DM 2 | |||
| Dry matter | 30.07 | 25.59 | 35.71 |
| CP | 6.91 | 7.84 | 8.45 |
| EE | 2.46 | 3.30 | 3.09 |
| NDF | 44.04 | 43.42 | 41.79 |
| ADF | 24.45 | 30.00 | 27.48 |
| Ash | 4.94 | 10.42 | 18.48 |
| Starch | 31.81 | 19.95 | 24.15 |
| Metabolic energy, MJ/kg 3 | 11.08 | 10.51 | 9.38 |
| Item% | Content% |
|---|---|
| Ground corn | 15.0 |
| Dried distiller’s grains with soluble (DDGS) | 3.6 |
| Soybean meal | 4.2 |
| Jujube powder | 4.5 |
| Stone powder | 0.3 |
| NaHCO3 | 0.6 |
| NaCl | 0.3 |
| CaHPO3·2H2O | 0.3 |
| 5% Premix 1 | 1.2 |
| Wheat straw | 70.0 |
| Item% | CS | PVS 1 | PVS 2 |
|---|---|---|---|
| Flaked corn | 21.85 | 21.59 | 22.08 |
| Soybean meal | 5.59 | 5.36 | 5.79 |
| Wheat bran | 3.73 | 3.57 | 3.86 |
| Cotton meal | 2.98 | 2.86 | 3.09 |
| 5% Premix 1 | 2.98 | 2.86 | 3.09 |
| Dried distiller’s grains with soluble (DDGS) | 2.98 | 2.86 | 3.09 |
| Rapeseed meal | 2.01 | 1.93 | 2.08 |
| Probiotics 2 | 1.34 | 1.29 | 1.39 |
| Fermented feed 3 | 4.97 | 4.76 | 5.14 |
| Straw | 2.48 | 2.38 | 2.57 |
| Rice stalks | 6.21 | 5.95 | 6.43 |
| Silage | 42.89 | 45.25 | 40.83 |
| Nutritive values, % DM 2 | |||
| Dry matter | 57.2 | 54.4 | 62.5 |
| Crude protein | 11.24 | 13.17 | 13.6 |
| Ether extract | 7.87 | 5.75 | 5.53 |
| Neutral detergent fiber | 28.35 | 28.76 | 27.7 |
| Acid detergent fiber | 17.00 | 17.00 | 15.41 |
| Ash | 8.51 | 9.11 | 10.81 |
| Ca 4, % | 0.41 | 0.39 | 0.42 |
| P 4, % | 0.15 | 0.14 | 0.15 |
| Starch | 31.81 | 19.95 | 24.57 |
| Metabolic energy, MJ/kg 5 | 12.17 | 11.83 | 11.37 |
| Items 1 | CS | PVS1 | PVS2 | SEM | p-Value 2 |
|---|---|---|---|---|---|
| Ruminal disappearance | |||||
| IVDMD24/% | 66.66 a | 68.49 a | 64.13 b | 0.524 | 0.001 |
| IVDMD48/% | 77.19 a | 73.74 b | 71.68 c | 0.519 | <0.001 |
| IVNDFD24/% | 37.86 b | 41.80 a | 34.47 b | 0.923 | 0.002 |
| IVNDFD48/% | 57.37 a | 50.62 b | 45.51 c | 1.088 | <0.001 |
| IVADFD24/% | 31.40 b | 41.99 a | 29.82 b | 1.338 | <0.001 |
| IVADFD48/% | 52.45 a | 49.79 b | 38.94 c | 1.290 | <0.001 |
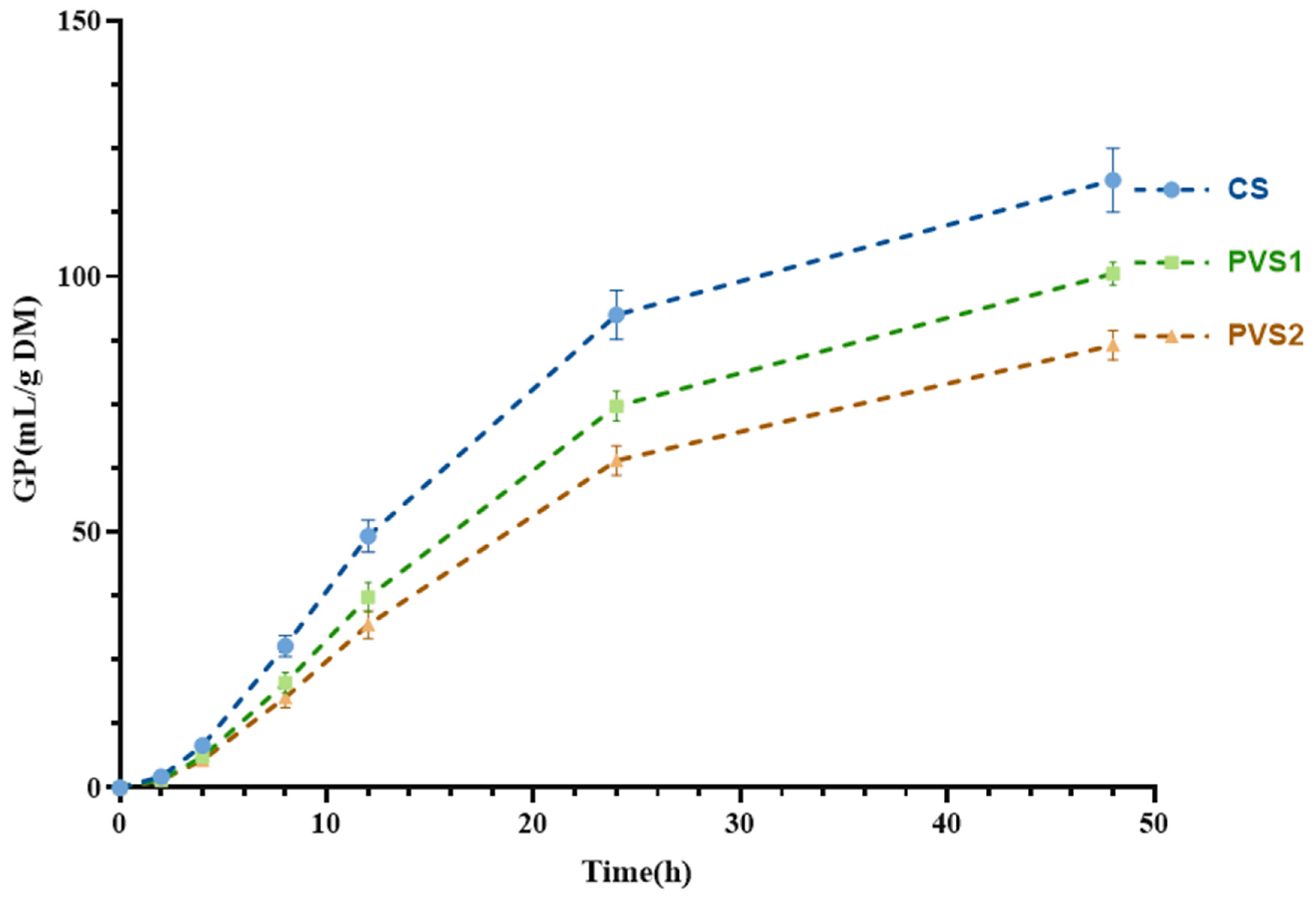
| GP48/(mL/g DM) 3 | 118.81 a | 100.49 ab | 88.70 b | 3.487 | 0.001 |
| Predicted CH4 (48 h) (mL/g DM) | 21.42 a | 19.00 b | 14.99 c | 0.666 | <0.001 |
| Gas production kinetics | |||||
| A/(mL/g DM) | 131.40 a | 115.24 ab | 100.77 b | 3.865 | 0.005 |
| B | 2.01 | 1.97 | 2.03 | 0.021 | 0.530 |
| C/(h) | 15.56 | 17.44 | 17.59 | 0.407 | 0.078 |
| TRmaxG/(h) | 8.96 | 10.10 | 10.04 | 0.356 | 0.410 |
| RmaxG/(mL/h) | 5.51 a | 4.32 b | 3.67 b | 0.204 | <0.001 |
| TRmaxS/(h) | 15.19 | 17.47 | 17.37 | 0.574 | 0.195 |
| RmaxS/(h) | 0.065 a | 0.058 b | 0.057 b | 0.001 | 0.012 |
| AGPR /(mL/h) | 4.24 a | 3.32 b | 2.86 b | 0.149 | 0.001 |
| Items 1 | CS | PVS1 | PVS2 | SEM | p-Value |
|---|---|---|---|---|---|
| Ruminal fermentation profile | |||||
| pH24 | 6.27 c | 6.40 b | 6.54 a | 0.024 | <0.001 |
| pH48 | 6.19 c | 6.42 b | 6.54 a | 0.030 | <0.001 |
| NH3-N24/(mg/dL) | 6.55 ab | 5.88 b | 7.73 a | 0.240 | 0.007 |
| NH3-N48/(mg/dL) | 8.48 ab | 8.04 b | 10.57 a | 0.421 | 0.020 |
| MCP24/(μg/mL) | 331.31 | 334.33 | 345.83 | 3.385 | 0.173 |
| MCP48/(μg/mL) | 308.71 b | 318.71 ab | 324.00 a | 2.564 | 0.040 |
| VFA pattern/(mmol/L) | |||||
| Acetate24 | 19.38 a | 16.62 ab | 13.41 b | 0.831 | 0.007 |
| Acetate48 | 25.46 a | 22.57 a | 18.96 b | 0.785 | 0.001 |
| Propionate24 | 9.92 a | 7.39 b | 6.03 b | 0.495 | 0.002 |
| Propionate48 | 12.20 a | 9.61 b | 8.43 b | 0.417 | <0.001 |
| Isobutyrate24 | 0.15 | 0.15 | 0.10 | 0.015 | 0.341 |
| Isobutyrate48 | 0.27 | 0.22 | 0.25 | 0.010 | 0.198 |
| Butyrate24 | 2.40 a | 1.89 b | 1.80 b | 0.106 | 0.036 |
| Butyrate48 | 3.45 a | 2.69 b | 2.55 b | 0.116 | 0.001 |
| Isovalerate24 | 0.45 a | 0.28 b | 0.28 b | 0.024 | 0.001 |
| Isovalerate48 | 0.65 a | 0.49 b | 0.53 b | 0.023 | 0.004 |
| Valerate24 | 0.29 a | 0.20 b | 0.17 b | 0.016 | 0.003 |
| Valerate48 | 0.41 a | 0.29 b | 0.28 b | 0.016 | <0.001 |
| Total VFA24 | 32.60 a | 26.53 b | 21.79 b | 1.458 | 0.005 |
| Total VFA48 | 42.43 a | 35.89 b | 30.99 c | 1.331 | <0.001 |
| Acetate to propionate24 | 1.97 b | 2.28 a | 2.23 a | 0.044 | 0.003 |
| Acetate to propionate48 | 2.09 b | 2.34 a | 2.25 a | 0.029 | <0.001 |
| TBCVFA24 | 0.60 a | 0.43 b | 0.38 b | 0.033 | 0.009 |
| TBCVFA48 | 0.92 a | 0.72 b | 0.78 b | 0.031 | 0.012 |
| NGR24 | 2.41 b | 2.75 a | 2.77 a | 0.051 | 0.001 |
| NGR48 | 2.61 b | 2.85 a | 2.80 a | 0.033 | 0.011 |
| Items | CS | PVS1 | PVS2 | SEM | p-Value |
|---|---|---|---|---|---|
| pH | 6.49 | 6.60 | 6.64 | 0.07 | 0.652 |
| NH3-N mg/dL | 7.03 | 6.42 | 6.37 | 0.21 | 0.391 |
| VFA pattern (mmol/L) | |||||
| Acetate | 31.20 | 27.59 | 30.26 | 3.08 | 0.897 |
| Propionate | 8.71 | 6.83 | 7.49 | 0.88 | 0.704 |
| Isobutyrate | 0.28 | 0.31 | 0.32 | 0.28 | 0.830 |
| Butyrate | 4.92 | 4.20 | 4.55 | 0.49 | 0.852 |
| Isovalerate | 0.74 | 0.57 | 0.57 | 0.07 | 0.499 |
| Valerate | 0.52 | 0.43 | 0.46 | 0.04 | 0.583 |
| Total VFA | 46.38 | 39.94 | 43.65 | 4.54 | 0.860 |
| Acetate to propionate | 3.66 | 4.02 | 4.27 | 0.11 | 0.076 |
| TBCVFA | 1.02 | 0.88 | 0.89 | 0.09 | 0.788 |
| NGR | 4.55 | 5.02 | 5.27 | 0.13 | 0.062 |
| Items | CS | PVS1 | PVS2 | SEM | p-Value |
|---|---|---|---|---|---|
| Metabolic pathways | 17.62 a | 17.49 b | 17.45 b | 0.028 | 0.021 |
| Biosynthesis of secondary metabolites | 8.01 a | 7.93 b | 7.95 b | 0.014 | 0.006 |
| Microbial metabolism in diverse environments | 3.69 b | 3.76 a | 3.74 a | 0.009 | <0.001 |
| Carbon metabolism | 2.57 b | 2.64 a | 2.62 a | 0.010 | 0.002 |
| Ribosome | 2.55 b | 2.62 a | 2.60 a | 0.010 | 0.007 |
| Amino sugar and nucleotide sugar metabolism | 1.24 a | 1.19 b | 1.21 ab | 0.008 | 0.046 |
| Aminoacyl-tRNA biosynthesis | 1.08 b | 1.12 a | 1.12 a | 0.005 | 0.001 |
| Alanine, aspartate and glutamate metabolism | 1.00 a | 0.97 b | 0.97 b | 0.004 | 0.002 |
| Carbon fixation pathways in prokaryotes | 0.94 b | 0.98 a | 0.96 a | 0.006 | 0.003 |
| Starch and sucrose metabolism | 0.99 a | 0.89 b | 0.91 b | 0.013 | 0.001 |
| Glycine, serine and threonine metabolism | 0.82 b | 0.84 a | 0.83 ab | 0.003 | 0.043 |
| Peptidoglycan biosynthesis | 0.75 b | 0.76 a | 0.76 a | 0.001 | 0.041 |
| Fructose and mannose metabolism | 0.77 a | 0.73 b | 0.73 b | 0.008 | 0.018 |
| Galactose metabolism | 0.72 a | 0.65 b | 0.67 b | 0.010 | 0.002 |
| Protein export | 0.60 b | 0.62 a | 0.61 a | 0.002 | 0.005 |
| RNA degradation | 0.59 b | 0.61 a | 0.60 a | 0.004 | 0.031 |
| Lysine biosynthesis | 0.60 a | 0.58 b | 0.60 ab | 0.003 | 0.041 |
| Pantothenate and CoA biosynthesis | 0.60 a | 0.59 b | 0.58 b | 0.002 | 0.009 |
| Fatty acid biosynthesis | 0.57 b | 0.60 a | 0.59 a | 0.006 | 0.025 |
| Fatty acid metabolism | 0.55 b | 0.60 a | 0.59 a | 0.006 | 0.003 |
| Terpenoid backbone biosynthesis | 0.55 b | 0.56 a | 0.56 a | 0.002 | 0.001 |
| Nicotinate and nicotinamide metabolism | 0.55 a | 0.54 b | 0.54 b | 0.002 | 0.015 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Deng, J.; Cui, Y.; Wang, L.; Li, Y.; Wang, X.; Min, S.; Wang, H.; Zhang, Q.; Li, P.; et al. Effect of Potato Vine and Leaf Mixed Silage to Whole Corn Crops on Rumen Fermentation and the Microbe of Fatten Angus Bulls. Fermentation 2023, 9, 704. https://doi.org/10.3390/fermentation9080704
Zhang S, Deng J, Cui Y, Wang L, Li Y, Wang X, Min S, Wang H, Zhang Q, Li P, et al. Effect of Potato Vine and Leaf Mixed Silage to Whole Corn Crops on Rumen Fermentation and the Microbe of Fatten Angus Bulls. Fermentation. 2023; 9(8):704. https://doi.org/10.3390/fermentation9080704
Chicago/Turabian StyleZhang, Siyu, Jiajie Deng, Yafang Cui, Lina Wang, Yingqi Li, Xianli Wang, Shengnan Min, Huili Wang, Qianzi Zhang, Peiqi Li, and et al. 2023. "Effect of Potato Vine and Leaf Mixed Silage to Whole Corn Crops on Rumen Fermentation and the Microbe of Fatten Angus Bulls" Fermentation 9, no. 8: 704. https://doi.org/10.3390/fermentation9080704