
Isolation and Optimal Fermentation Conditions of Bacillus licheniformis SFD-Y5 for a New Douchi Fibrinolytic Enzyme Producer
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Bacterial Diversity Analysis of Douchi Sample
2.3. Isolation of FE-Producing Bacteria
2.4. Identification of FE-Producing Bacteria
2.5. Measurement of Fibrinolytic Enzyme Activity
2.6. Optimization of the Culture Medium and Fermentation Parameters for Y5 FE Production
2.6.1. The Effect of Culture Medium on Y5 FE Production
2.6.2. The Effect of Fermentation Parameters on Y5 FE Production
2.6.3. Plackett–Burman, Steepest Ascent Experiment and Response Surface Experiment
2.7. The Physiological-Biochemical Characteristic of Y5 FE
2.7.1. The Effects of Protease Inhibitors on Y5 FE
2.7.2. The Effects of Metal Ions on Y5 FE
2.7.3. The Optimal pH and pH Stability of Y5 FE
2.7.4. The Optimal Temperature and Thermal Stability of Y5 FE
2.8. Statistical Analysis
3. Results and Discussion
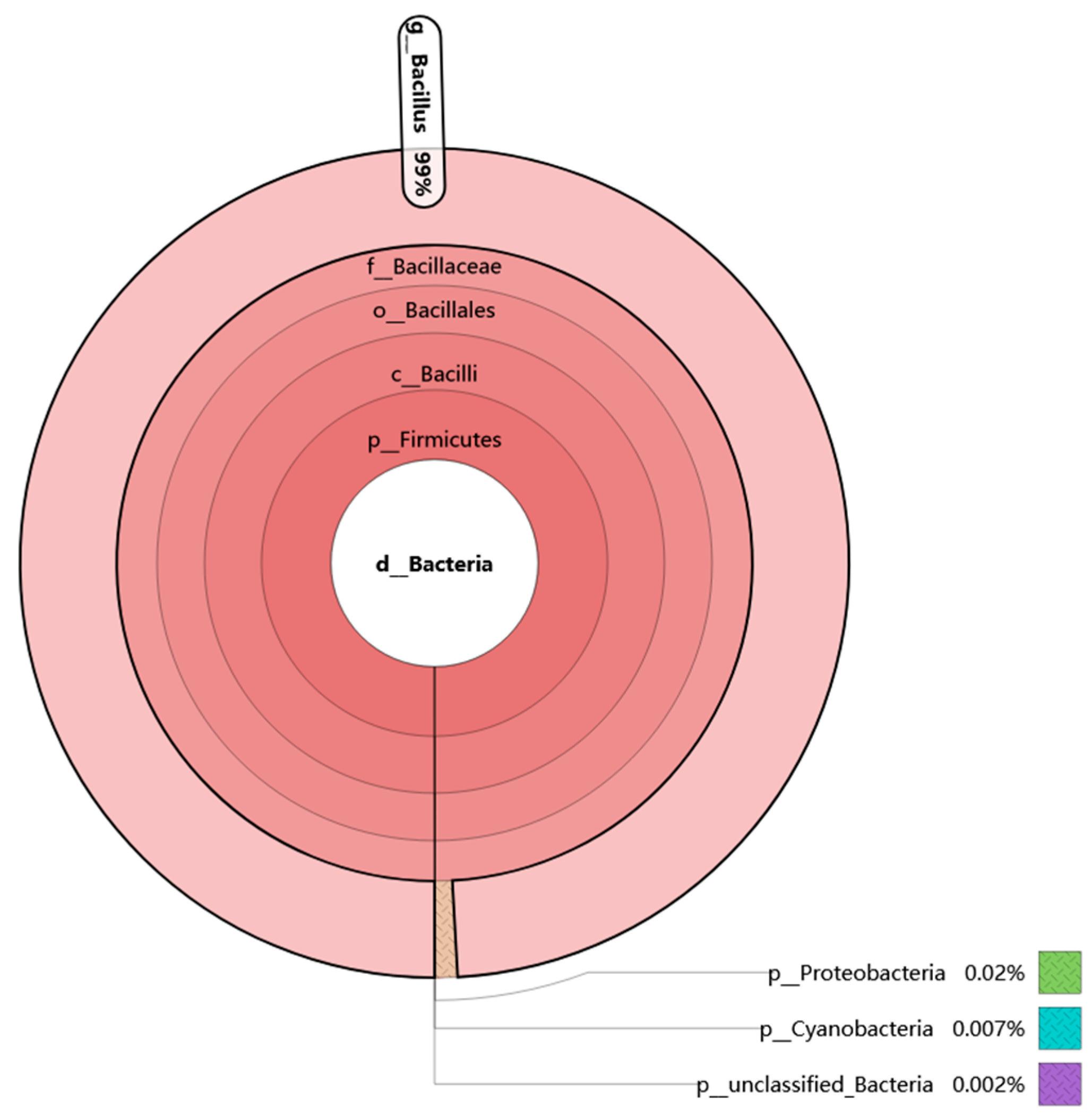
3.1. Bacterial Diversity Analysis of Douchi Sample
3.2. Isolation and Identification of FE-Producing Bacteria
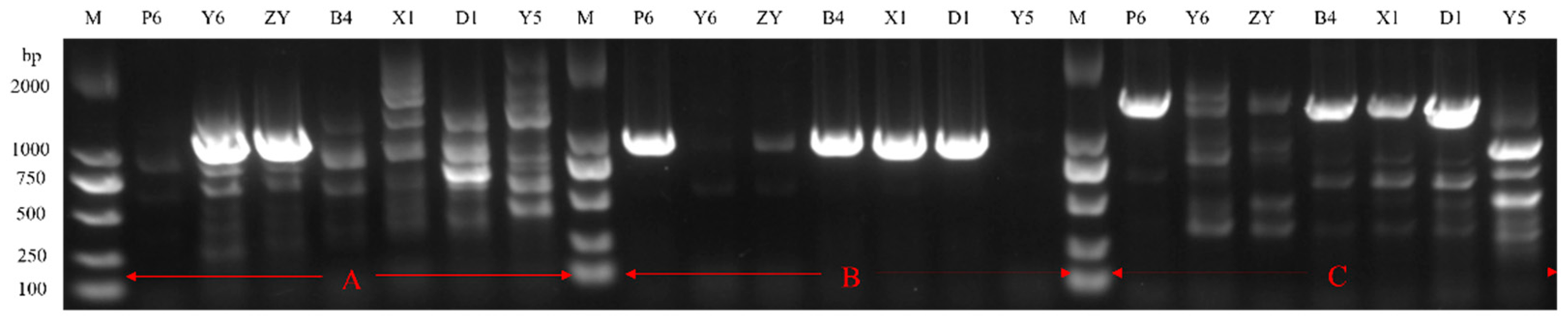
3.2.1. Isolation of FE-Producing Bacteria
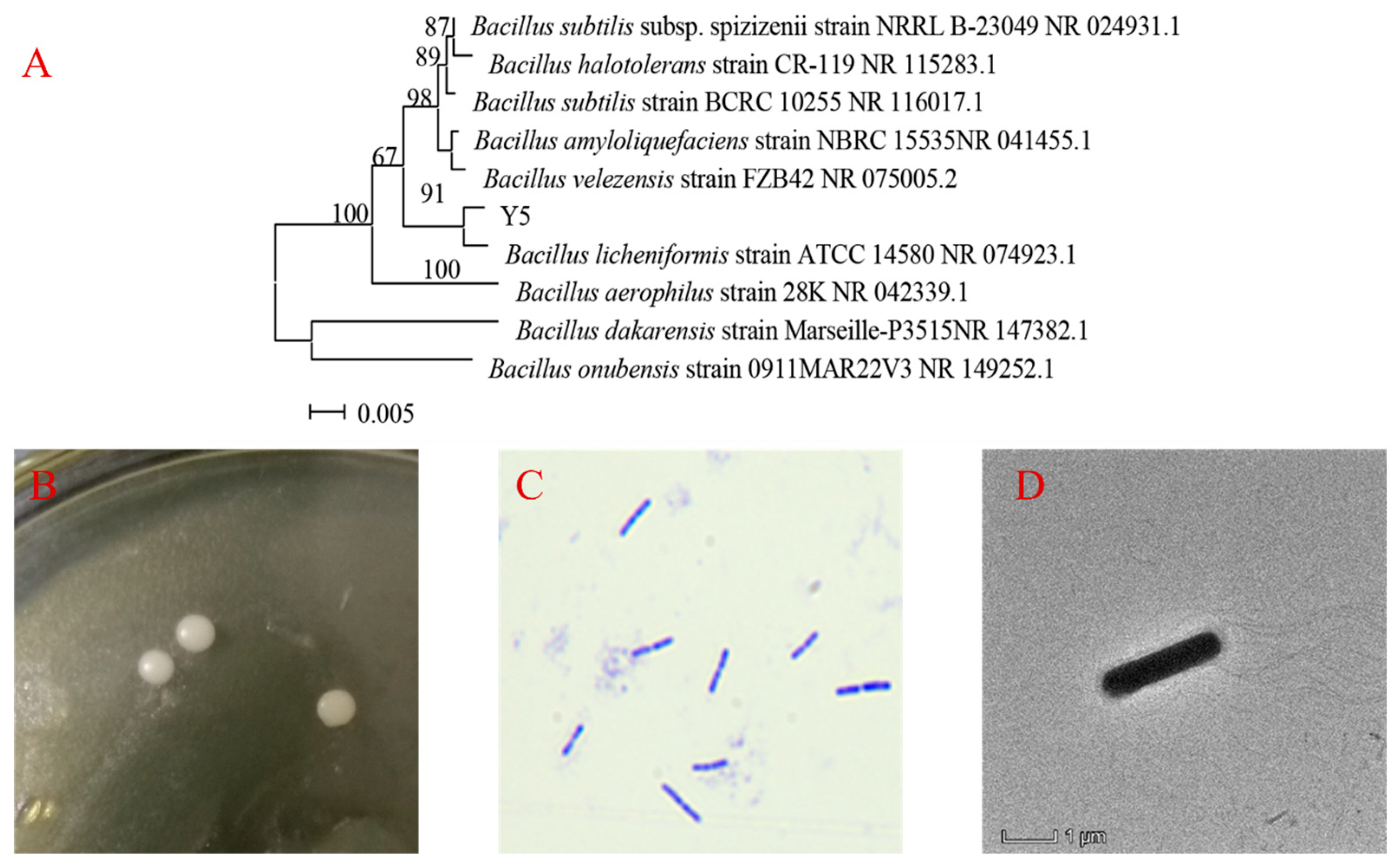
3.2.2. Further Identification of Y5
3.3. Optimization of the Culture Medium and Fermentation Parameters for Y5 FE Production
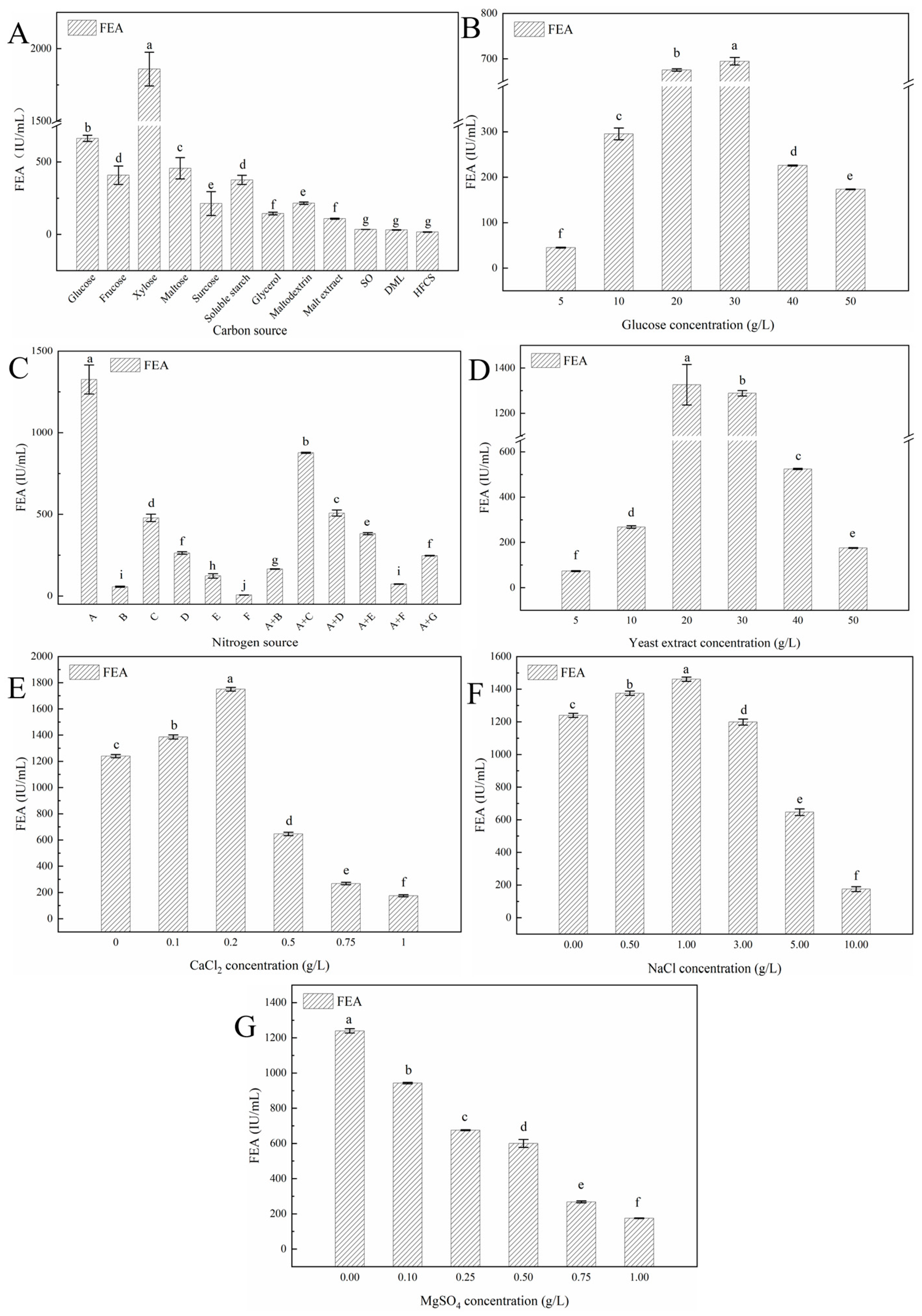
3.3.1. The Effects of Fermentation Medium Components on Y5 FE Production
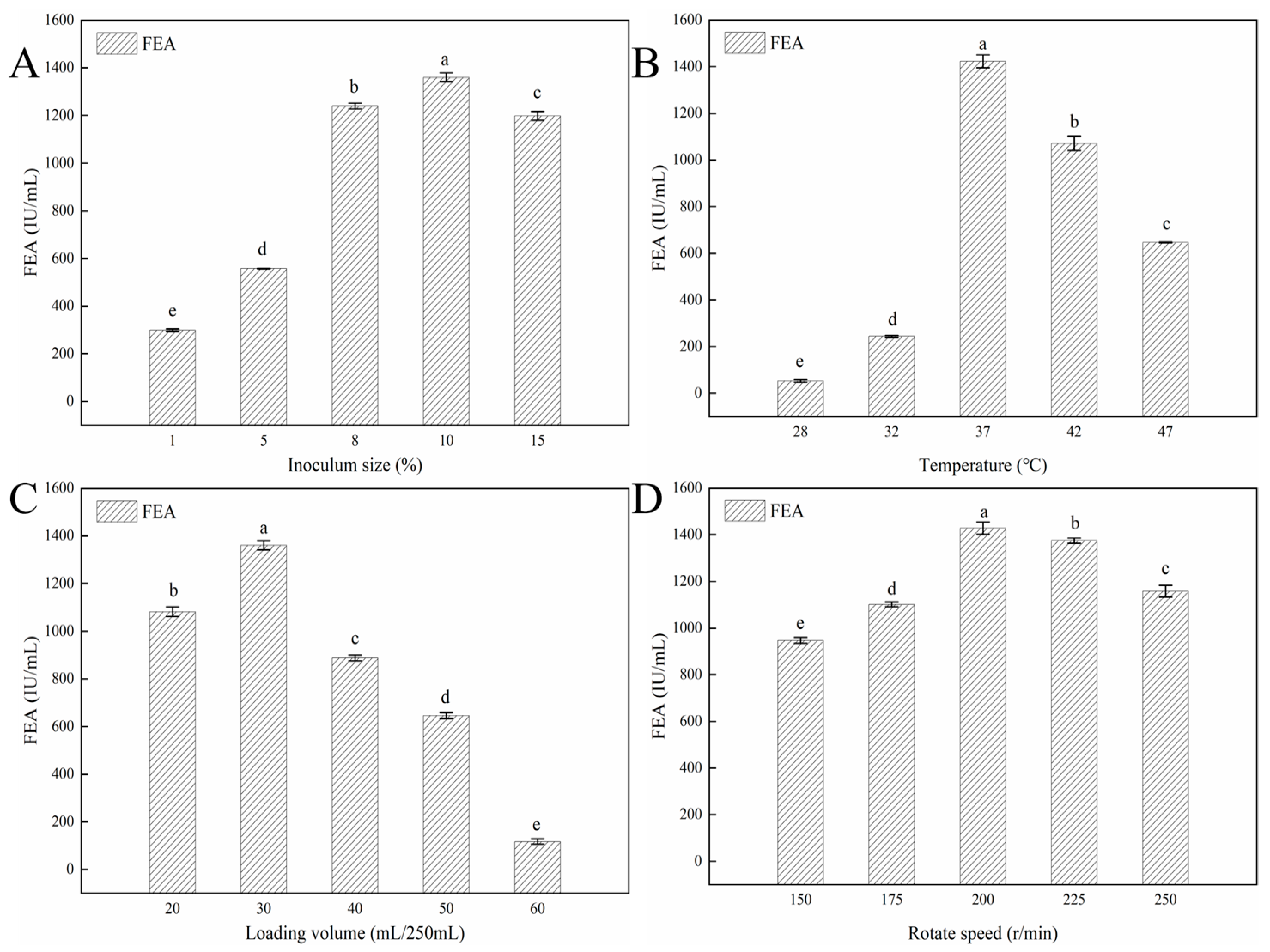
3.3.2. The Effects of Fermentation Parameters on Y5 FE Production
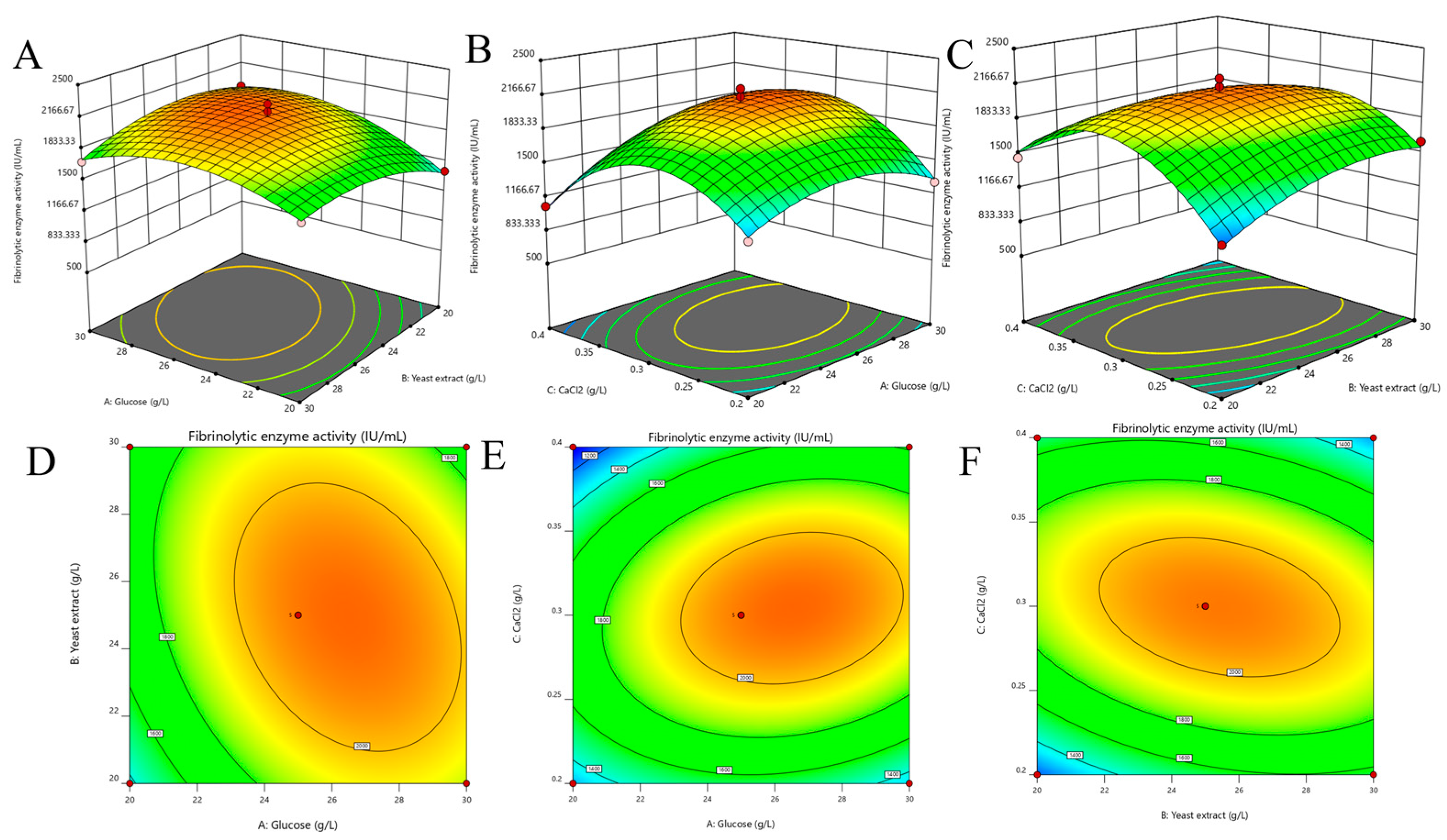
3.3.3. Optimization of Y5 FE Production by Statistical Experiments
3.4. The Physiological-Biochemical Characteristic of Y5 FE
3.4.1. The Effects of Protease Inhibitors and Metal Ions on Y5 FE
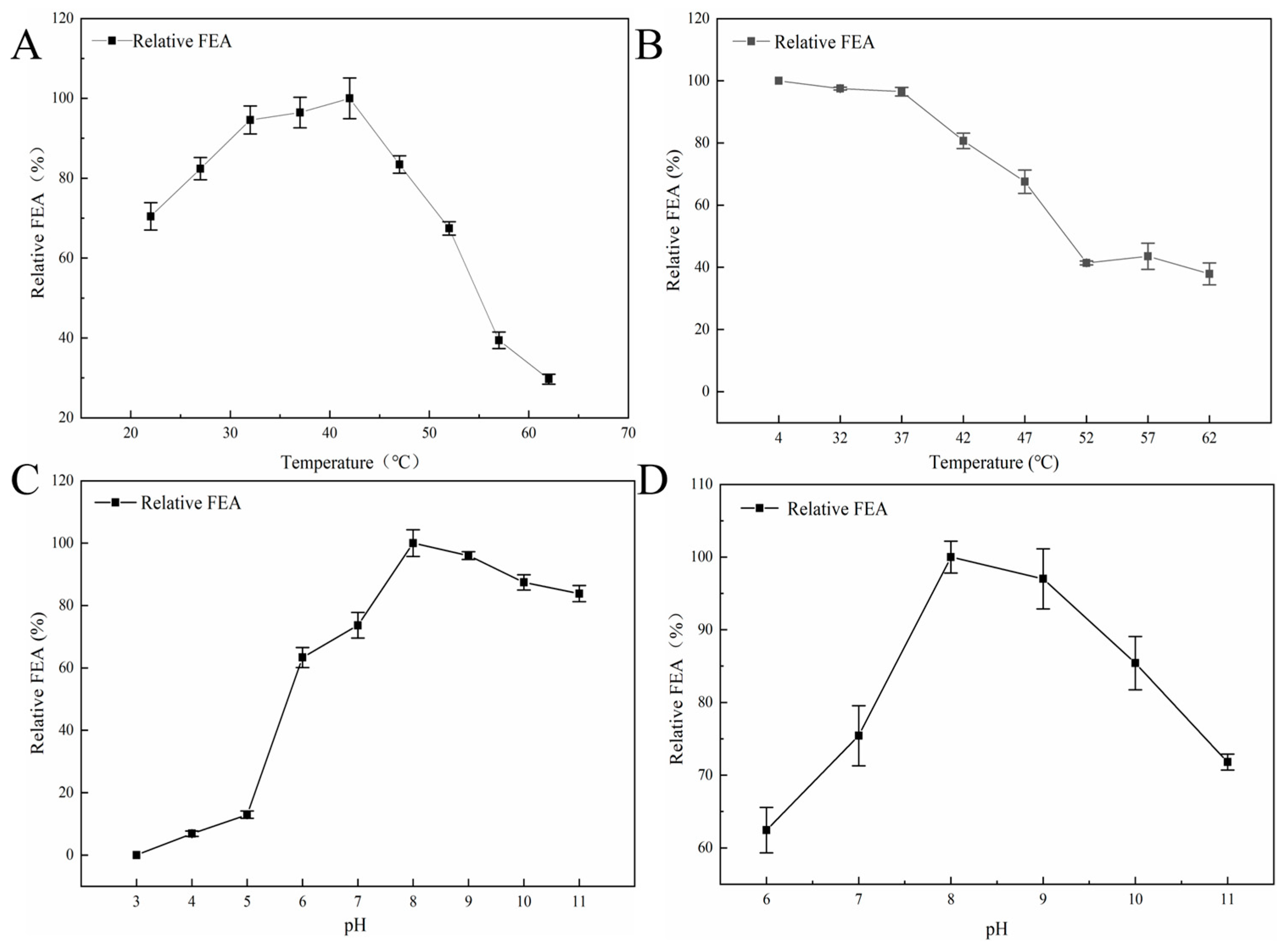
3.4.2. The Optimal Temperatures and Thermal Stability of Y5 FE
3.4.3. The Optimal pH and pH Stability of Y5 FE
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Noncommunicable Diseases: Mortality; WHO: Geneva, Switzerland, 2023.
- Yao, M.; Yang, Y.; Fan, J.; Ma, C.; Liu, X.; Wang, Y.; Wang, B.; Sun, Z.; McClements, D.J.; Zhang, J.; et al. Production, purification, and functional properties of microbial fibrinolytic enzymes produced by microorganism obtained from soy-based fermented foods: Developments and challenges. Crit. Rev. Food Sci. Nutr. 2022, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.S.; Sabu, A. Fibrinolytic Enzymes for Thrombolytic Therapy [M]//LABROU N. In Therapeutic Enzymes: Function and Clinical Implications; Springer: Berlin/Heidelberg, Germany, 2019; pp. 345–381. [Google Scholar]
- Sharma, C.; Osmolovskiy, A.; Singh, R. Microbial Fibrinolytic Enzymes as Anti-Thrombotics: Production, Characterisation and Prodigious Biopharmaceutical Applications. Pharmaceutics 2021, 13, 1880. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tang, X.; Chen, L.; Xu, X.; Li, J. Characterization of a Nattokinase from the Newly Isolated Bile Salt-Resistant Bacillus mojavensis LY-06. Foods 2022, 11, 2403. [Google Scholar] [CrossRef] [PubMed]
- Kajihara, Y. Dabigatran-induced esophagitis. Clevel. Clin. J. Med. 2019, 86, 303–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barzkar, N.; Jahromi, S.T.; Vianello, F. Marine Microbial Fibrinolytic Enzymes: An Overview of Source, Production, Biochemical Properties and Thrombolytic Activity. Mar. Drugs 2022, 20, 46. [Google Scholar] [CrossRef] [PubMed]
- Pinontoan, R.; Elvina; Sanjaya, A.; Jo, J. Fibrinolytic characteristics of Bacillus subtilis G8 isolated from natto. Biosci. Microbiota Food Health 2021, 40, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Sumi, H.; Hamada, H.; Tsushima, H.; Mihara, H.; Muraki, H. A novel fibrinolytic enzyme (nattokinase) in the vegetable cheese Natto; a typical and popular soybean food in the Japanese diet. Experienti 1987, 43, 1110–1111. [Google Scholar] [CrossRef]
- Peng, Y.; Yang, X.; Zhang, Y. Microbial fibrinolytic enzymes: An overview of source, production, properties, and thrombolytic activity in vivo. Appl. Microbiol. Biotechnol. 2005, 69, 126–132. [Google Scholar] [CrossRef]
- Man, L.L.; Xiang, D.J.; Zhang, C.L. Strain Screening from Traditional Fermented Soybean Foods and Induction of Nattokinase Production in Bacillus subtilis MX-6. Probiotics Antimicrob. Proteins 2019, 11, 283–294. [Google Scholar] [CrossRef]
- Liu, X.L.; Kopparapu, N.K.; Zheng, H.C.; Katrolia, P.; Deng, Y.P.; Zheng, X.Q. Purification and characterization of a fibrinolytic enzyme from the food-grade fungus, Neurospora sitophila. J. Mol. Catal. B Enzym. 2016, 134, 98–104. [Google Scholar] [CrossRef]
- Wei, X.; Luo, M.; Xie, Y.; Yang, L.; Li, H.; Xu, L.; Liu, H. Strain screening, fermentation, separation, and encapsulation for production of nattokinase functional food. Appl. Biochem. Biotechnol. 2012, 168, 1753–1764. [Google Scholar] [CrossRef]
- Jeong, S.-J.; Heo, K.; Park, J.Y.; Lee, K.W.; Park, J.-Y.; Joo, S.H.; Kim, J.H. Characterization of AprE176, a fibrinolytic enzyme from Bacillus subtilis HK176. J. Microbiol. Biotechnol. 2015, 25, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Yao, Z.; Liu, X.; Shim, J.M.; Lee, K.W.; Kim, H.-J.; Kim, J.H. Properties of a Fibrinolytic Enzyme Secreted by Bacillus amyloliquefaciens RSB34, Isolated from Doenjang. J. Microbiol. Biotechnol. 2017, 27, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Bae, D.H.; Kwon, T.J.; Lee, S.B.; Lee, H.H.; Park, J.H.; Heo, S.; Johnson, M.G. Purification and characterization of a fibrinolytic enzyme from Bacillus sp. KDO-13 isolated from soybean paste. J. Microbiol. Biotechnol. 2001, 11, 845–852. [Google Scholar]
- Han, X.; Shiwa, Y.; Itoh, M.; Suzuki, T.; Yoshikawa, H.; Nakagawa, T.; Nagano, H. Molecular cloning and sequence analysis of an extracellular protease from four Bacillus subtilis strains. Biosci. Biotechnol. Biochem. 2013, 77, 870–873. [Google Scholar] [CrossRef] [PubMed]
- Huy, D.; Hao, P.A.; Hung, P.V. Screening and identification of Bacillus sp. isolated from traditional Vietnamese soybean-fermented products for high fibrinolytic enzyme production. Int. Food Res. J. 2016, 23, 326–331. [Google Scholar]
- Sewalt, V.; Shanahan, D.; Gregg, L.; La Marta, J.; Carrillo, R. The Generally Recognized as Safe (GRAS) Process for Industrial Microbial Enzymes. Ind. Biotechnol. 2016, 12, 295–302. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Chen, X.; Hao, L.; Zhang, G.; Jin, Z.; Li, C.; Yang, Y.; Rao, J.; Chen, B. Traditional fermented soybean products: Processing, flavor formation, nutritional and biological activities. Crit. Rev. Food Sci. Nutr. 2022, 62, 1971–1989. [Google Scholar] [CrossRef]
- Zhang, X.; Yun, L.-J.; Peng, L.-B.; Lu, Y.; Ma, K.-P.; Tang, F. Optimization of Douchi fibrinolytic enzyme production by statistical experimental methods. J. Huazhong Univ. Sci. Technol. Med. Sci. 2013, 33, 153–158. [Google Scholar] [CrossRef]
- Nakamura, T.; Yamagata, Y.; Ichishima, E. Nucleotide Sequence of the Subtilisin NAT Gene, aprN, of Bacillus subtilis (natto). Biosci. Biotechnol. Biochem. 1992, 56, 1869–1871. [Google Scholar] [CrossRef] [Green Version]
- Jo, H.-D.; Kwon, G.-H.; Park, J.-Y.; Cha, J.; Song, Y.-S.; Kim, J.H. Cloning and overexpression of aprE3-17 encoding the major fibrinolytic protease of Bacillus licheniformis CH 3-17. Biotechnol. Bioprocess Eng. 2011, 16, 352–359. [Google Scholar] [CrossRef]
- Yang, H.; Liu, Y.; Ning, Y.; Wang, C.; Zhang, X.; Weng, P.; Wu, Z. Characterization of an Intracellular Alkaline Serine Protease from Bacillus velezensis SW5 with Fibrinolytic Activity. Curr. Microbiol. 2020, 77, 1610–1621. [Google Scholar] [CrossRef] [PubMed]
- Astrup, T.; Müllertz, S. The Fibrin Plate Method for Estimating Fibrinolytic Activity. Arch. Biochem. Biophys. 1952, 40, 346–351. [Google Scholar] [CrossRef]
- Li, G.; Liu, X.; Cong, S.; Deng, Y.; Zheng, X. A novel serine protease with anticoagulant and fibrinolytic activities from the fruiting bodies of mushroom Agrocybe aegerita. Int. J. Biol. Macromol. 2021, 168, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Salunke, A.S.; Nile, S.H.; Kharat, A.S. A comparative study on fibrinolytic enzymes extracted from six Bacillus spp. isolated from fruit-vegetable waste biomass. Food Biosci. 2022, 50, 102149. [Google Scholar] [CrossRef]
- Yang, L.; Yang, H.-L.; Tu, Z.-C.; Wang, X.-L. High-Throughput Sequencing of Microbial Community Diversity and Dynamics during Douchi Fermentation. PLoS ONE 2016, 11, e0168166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, R.; Chen, G.; Pan, S.; Zeng, J.; Liang, Z. Cost-effective fibrinolytic enzyme production by Bacillus subtilis WR350 using medium supplemented with corn steep powder and sucrose. Sci. Rep. 2019, 9, 6824. [Google Scholar] [CrossRef] [Green Version]
- Yao, Z.; Kim, J.A.; Kim, J.H. Characterization of a Fibrinolytic Enzyme Secreted by Bacillus velezensis BS2 Isolated from Sea Squirt Jeotgal. J. Microbiol. Biotechnol. 2019, 29, 347–356. [Google Scholar] [CrossRef]
- Xin, X.; Ambati, R.R.; Cai, Z.; Lei, B. Purification and characterization of fibrinolytic enzyme from a bacterium isolated from soil. 3 Biotech 2018, 8, 90. [Google Scholar] [CrossRef]
- Qiuling, X. Optimization of Conditions in Liqiuid Fermentation to Produce Nattokinase and Research about Metabolic Regulation of Nattokinase Biosynthesis. Ph.D. Thesis, South China University of Technology, Guangzhou, China, 1999. [Google Scholar]
- Singh, K.; Richa, K.; Bose, H.; Karthik, L.; Kumar, G.; Rao, K.V.B. Statistical media optimization and cellulase production from marine Bacillus VITRKHB. 3 Biotech 2014, 4, 591–598. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Liu, X.; Zhang, L.; Ding, Z.; Xu, S.; Gu, Z.; Shi, G. Transcriptional Changes in the Xylose Operon in Bacillus licheniformis and Their Use in Fermentation Optimization. Int. J. Mol. Sci. 2019, 20, 4615. [Google Scholar] [CrossRef] [Green Version]
- Vijayaraghavan, P.; Arasu, M.V.; Rajan, R.A.; Al-Dhabi, N. Enhanced production of fibrinolytic enzyme by a new Xanthomonas oryzae IND3 using low-cost culture medium by response surface methodology. Saudi J. Biol. Sci. 2019, 26, 217–224. [Google Scholar] [CrossRef]
- Li, D.; Hou, L.; Hu, M.; Gao, Y.; Tian, Z.; Fan, B.; Li, S.; Wang, F. Recent Advances in Nattokinase-Enriched Fermented Soybean Foods: A Review. Foods 2022, 11, 1867. [Google Scholar] [CrossRef] [PubMed]
- Ju, S.; Cao, Z.; Wong, C.; Liu, Y.; Foda, M.F.; Zhang, Z.; Li, J. Isolation and Optimal Fermentation Condition of the Bacillus subtilis Subsp. natto Strain WTC016 for Nattokinase Production. Ermentation 2019, 5, 92. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Zhan, C.; Guo, G.; Liu, Z.; Hao, N.; Ouyang, P. Tofu processing wastewater as a low-cost substrate for high activity nattokinase production using Bacillus subtilis. BMC Biotechnol. 2021, 21, 57. [Google Scholar] [CrossRef]
- Zhang, Y.; Xia, Y.; Lai, P.F.-H.; Liu, X.; Xiong, Z.; Liu, J.; Ai, L. Fermentation conditions of serine/alkaline milk-clotting enzyme production by newly isolated Bacillus licheniformis BL312. Ann. Microbiol. 2019, 69, 1289–1300. [Google Scholar] [CrossRef]
- KyHwang, K.-J.; Choi, K.-H.; Kim, M.-J.; Park, C.-S.; Cha, J. Purification and characterization of a new fibrinolytic enzyme of Bacillus licheniformis KJ-31, isolated from Korean traditional Jeot-gal. J. Microbiol. Biotechnol. 2007, 17, 1469–1476. [Google Scholar]
- Afifah, D.N.; Rustanti, N.; Anjani, G.; Syah, D.; Yanti; Suhartono, M.T. Proteomics study of extracellular fibrinolytic proteases from Bacillus licheniformis RO3 and Bacillus pumilus 2.g isolated from Indonesian fermented food. IOP Conf. Ser. Earth Environ. Sci. 2017, 55, 012025. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Primers | References |
|---|---|---|
| Bacillus subtilis | F1: 5′-GTGAGAAGCAAAAAATTGTG-3′ R1: 5′-TTATTGTGCAGCTGCTTGTA-3′ | [22] |
| Bacillus licheniformis | F2: 5′-AGGATCCCAAGAGAGCGATTGCGGCTGTGTAC-3 R2: 5′-ACTCGAGTTCAGAGGGAGCCACCCGTCGATCA-3 | [23] |
| Bacillus velezensis | F3: 5′-CGACGGGATCCAATGAATGGTGAAATGCATTTGA-3′ R3: 5′-GCCGCTCGAGGAAAGACAGCAGCTGTGCCT-3′ | [24] |
| Strain | Primary (mm2) | Secondary (mm2) | 16S rDNA |
|---|---|---|---|
| B4 | 162.79 ± 12.65 b | 169.24 ± 10.12 ab | Bacillus velezensis |
| D1 | 165.44 ± 2.04 ab | 159.56 ± 6.43 c | Bacillus velezensis |
| P6 | 146.88 ± 9.48 c | 148.74 ± 5.64 d | Bacillus subtilis |
| X1 | 150.64 ± 7.76 c | 160.13 ± 4.98 c | Bacillus velezensis |
| Y5 | 156.95 ± 5.18 bc | 164.67 ± 5.43 bc | Bacillus licheniformis |
| Y6 | 148.61 ± 7.48 c | 142.61 ± 7.62 e | Bacillus subtilis |
| ZY | 175.06 ± 11.12 a | 173.16 ± 8.46 a | Bacillus subtilis |
| HBI G14 Kit | API 50 CHB | ||||||
|---|---|---|---|---|---|---|---|
| Test | Results | Substrate | Results | Substrate | Results | Substrate | Results |
| V-P | + | Glycerol | + | Methyl-α-D- glucopyranoside | + | Salicin | − |
| Citrate | − | Erythritol | − | N-Acetylglucosamine | − | D-Cellobiose | + |
| Propionate | − | D-Arabinose | − | Amygdalin | + | D-Maltose | + |
| D-xylose | − | L-Arabinose | + | Arbutin | + | D-Lactose (bovine origin) | − |
| L-arabinose | + | D-Ribose | + | Esculin ferric citrate | + | D-Melibiose | − |
| D-mannitol | + | D-Xylose | + | L-Arabitol | − | D-Sucrose | + |
| Gelatin liquefaction | + | L-Xylose | − | Potassium gluconate | − | D-Fucose | − |
| 7% NaCl | − | D-Adonitol | − | Potassium 2- Ketogluconate | − | Inulin | − |
| pH 5.7 | + | Methyl-β-D- xylopyranoside | − | Potassium 5- Ketogluconate | − | D-Melezitose | − |
| Nitrate reduction | + | D-Galactose | + | Sorbitol | + | D-Raffinose | − |
| Amylohydrolysis | + | D-Glucose | + | Methyl-α-D- mannopyranoside | − | Amidon (starch) | − |
| D-Fructose | + | D-Fucose | − | Glycogen | + | ||
| D-Mannose | + | L-Fucose | − | Xylitol | − | ||
| L-Sorbose | − | D-Arabitol | − | D-Gentiobiose | + | ||
| L-Rhamnose | − | D-Mannitol | + | D-Turanose | + | ||
| Dulcitol | − | D-Lyxose | − | ||||
| Inositol | + | D-Tagatose | + | ||||
| (a) | ||||||||||||
| Num | Glu (g/L) | Yeast (g/L) | V 1 | IS (%) | CaCl2 (g/L) | V 2 | Temperature (°C) | NaCl (g/L) | V 3 | LV (mL) | RS (r/min) | FEA (IU/mL) |
| 1 | 40 | 20 | 1 | 15 | 0.2 | 1 | 42 | 3 | −1 | 30 | 200 | 1070.27 ± 20.37 |
| 2 | 40 | 40 | −1 | 15 | 0.5 | 1 | 37 | 1 | −1 | 50 | 200 | 747.81 ± 10.84 |
| 3 | 20 | 40 | 1 | 10 | 0.5 | 1 | 42 | 1 | −1 | 30 | 225 | 618.45 ± 9.56 |
| 4 | 20 | 40 | −1 | 15 | 0.5 | −1 | 42 | 3 | 1 | 30 | 200 | 608.16 ± 7.91 |
| 5 | 40 | 20 | 1 | 15 | 0.5 | −1 | 37 | 1 | 1 | 30 | 225 | 1139.15 ± 17.43 |
| 6 | 40 | 40 | −1 | 10 | 0.2 | 1 | 37 | 3 | 1 | 30 | 225 | 522.06 ± 9.32 |
| 7 | 20 | 40 | 1 | 15 | 0.2 | −1 | 37 | 3 | −1 | 50 | 225 | 207.06 ± 5.62 |
| 8 | 20 | 20 | −1 | 15 | 0.2 | 1 | 42 | 1 | 1 | 50 | 225 | 745.82 ± 16.15 |
| 9 | 20 | 20 | 1 | 10 | 0.5 | 1 | 37 | 3 | 1 | 50 | 200 | 1042.34 ± 10.29 |
| 10 | 40 | 20 | −1 | 10 | 0.5 | −1 | 42 | 3 | −1 | 50 | 225 | 1558.31 ± 36.71 |
| 11 | 40 | 40 | 1 | 10 | 0.2 | −1 | 42 | 1 | 1 | 50 | 200 | 758.94 ± 14.34 |
| 12 | 20 | 20 | −1 | 10 | 0.2 | −1 | 37 | 1 | −1 | 30 | 200 | 555.56 ± 19.48 |
| (b) | ||||||||||||
| Factor | Standardized Effect | Contribution (%) | df | p-Value | Significance | |||||||
| Glucose | 336.523 | 24.487 | 2 | 0.0001 | ** | |||||||
| Yeast extract | −441.492 | 42.145 | 1 | <0.0001 | ** | |||||||
| Inoculum size | −89.565 | 1.734 | 6 | 0.007 | ** | |||||||
| CaCl2 | 309.083 | 20.656 | 3 | 0.0002 | ** | |||||||
| Temperature | 190.995 | 7.888 | 4 | 0.0008 | ** | |||||||
| NaCl | 73.743 | 1.176 | 7 | 0.012 | * | |||||||
| Loading volume | 91.105 | 1.795 | 5 | 0.0066 | ** | |||||||
| Rotate speed | 1.295 | 0.0003 | 8 | 0.9296 | ||||||||
| Num | Glucose (g/L) | Yeast Extract (g/L) | CaCl2 (g/L) | FEA (IU/mL) |
|---|---|---|---|---|
| 1 | 15 | 35 | 0.1 | 638.59 ± 18.24 |
| 2 | 20 | 30 | 0.2 | 1240.76 ± 29.67 |
| 3 | 25 | 25 | 0.3 | 1742.63 ± 43.24 |
| 4 | 30 | 20 | 0.4 | 1647.68 ± 27.28 |
| 5 | 35 | 15 | 0.5 | 762.48 ± 24.38 |
| (a) | ||||||
| Num. | Glucose (g/L) | Yeast Extract (g/L) | CaCl2 (g/L) | FEA (IU/Ml) | ||
| 1 | 20 | 25 | 0.4 | 1076.39 ± 22.64 | ||
| 2 | 25 | 25 | 0.3 | 2013.45 ± 19.48 | ||
| 3 | 25 | 30 | 0.2 | 1612.31 ± 34.28 | ||
| 4 | 25 | 25 | 0.3 | 1976.54 ± 9.98 | ||
| 5 | 25 | 30 | 0.4 | 1235.18 ± 20.18 | ||
| 6 | 25 | 25 | 0.3 | 2138.25 ± 16.45 | ||
| 7 | 20 | 20 | 0.3 | 1396.79 ± 23.57 | ||
| 8 | 30 | 25 | 0.4 | 1621.48 ± 21.46 | ||
| 9 | 20 | 25 | 0.2 | 1239.37 ± 17.69 | ||
| 10 | 25 | 20 | 0.2 | 1196.43 ± 21.28 | ||
| 11 | 25 | 25 | 0.3 | 2069.17 ± 31.42 | ||
| 12 | 20 | 30 | 0.3 | 1643.19 ± 19.34 | ||
| 13 | 25 | 25 | 0.3 | 2216.45 ± 14.68 | ||
| 14 | 30 | 20 | 0.3 | 1901.27 ± 22.35 | ||
| 15 | 30 | 30 | 0.3 | 1698.64 ± 20.17 | ||
| 16 | 25 | 20 | 0.4 | 1462.34 ± 27.63 | ||
| 17 | 30 | 25 | 0.2 | 1287.62 ± 30.21 | ||
| (b) | ||||||
| Source | Sum of Squares | df | Mean Square | F-Value | p-Value | Significance |
| Model | 2.09 × 106 | 9 | 2.33 × 105 | 32.61 | <0.0001 | ** |
| A-glucose | 1.66 × 105 | 1 | 1.66 × 105 | 23.31 | 0.0019 | ** |
| B-yeast extract | 1.08 × 105 | 1 | 1.08 × 105 | 18.69 | 0.0362 | * |
| C-CaCl2 | 5.34 × 104 | 1 | 5.34 × 104 | 6.43 | 0.0415 | * |
| AB | 5.04 × 104 | 1 | 5.04 × 104 | 7.07 | 0.0326 | * |
| AC | 6.17 × 104 | 1 | 6.17 × 104 | 8.65 | 0.0217 | * |
| BC | 1.03 × 105 | 1 | 1.03 × 105 | 14.49 | 0.0067 | ** |
| A2 | 2.56 × 105 | 1 | 2.56 × 105 | 35.88 | 0.0005 | ** |
| B2 | 1.31 × 105 | 1 | 1.31 × 105 | 18.33 | 0.0036 | ** |
| C2 | 1.18 × 106 | 1 | 1.18 × 106 | 165.78 | <0.0001 | ** |
| Residual | 4.99 × 104 | 7 | 7.13 × 103 | |||
| Lack of fit | 1.27 × 104 | 3 | 4.24 × 103 | 0.4554 | 0.7279 | |
| Pure error | 3.72 × 104 | 4 | 9.30 × 104 | |||
| Cor Total | 2.14 × 106 | 16 | ||||
| R2 = 0.9767, R2Adj = 0.9467 | ||||||
| Inhibitors | Concentration | Relative FEA (%) |
|---|---|---|
| Control | - | 100.00% |
| EDTA | 5 mM | 0 |
| EDTA | 1 mM | 0 |
| PMSF | 10 mM | 0 |
| PMSF | 5 mM | 0 |
| PMSF | 1 mM | 34.89% |
| Pepstine A | 5 mM | 98.99% |
| Pepstine A | 1 mM | 101.12% |
| DTT | 5 mM | 96.36% |
| DTT | 1 mM | 98.51% |
| EDTA | 5 mM | 0 |
| Metal Ions | Concentration | Relative FEA (%) | Metal Ions | Concentration | Relative FEA (%) |
|---|---|---|---|---|---|
| Control | - | 100.00% | |||
| K+ | 10 mM | 113.36% | K+ | 5 mM | 58.54% |
| Ca2+ | 10 mM | 155.94% | Ca2+ | 5 mM | 75.54% |
| Cu2+ | 10 mM | 34.89% | Cu2+ | 5 mM | 25.55% |
| Na+ | 10 mM | 98.52% | Na+ | 5 mM | 99.85% |
| Zn2+ | 10 mM | 176.53% | Zn2+ | 5 mM | 122.38% |
| Mg2+ | 10 mM | 130.94% | Mg2+ | 5 mM | 52.87% |
| Co2+ | 10 mM | 82.80% | Co2+ | 5 mM | 79.29% |
| Fe2+ | 10 mM | 36.80% | Fe2+ | 5 mM | 34.77% |
| Mn2+ | 10 mM | 130.77% | Mn2+ | 5 mM | 88.78% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, M.; Ma, C.; Bian, X.; Yang, Y.; Xu, Y.; Wu, Q.; Xu, X.; Li, L.; Zhang, N.; Tian, Y. Isolation and Optimal Fermentation Conditions of Bacillus licheniformis SFD-Y5 for a New Douchi Fibrinolytic Enzyme Producer. Fermentation 2023, 9, 668. https://doi.org/10.3390/fermentation9070668
Yao M, Ma C, Bian X, Yang Y, Xu Y, Wu Q, Xu X, Li L, Zhang N, Tian Y. Isolation and Optimal Fermentation Conditions of Bacillus licheniformis SFD-Y5 for a New Douchi Fibrinolytic Enzyme Producer. Fermentation. 2023; 9(7):668. https://doi.org/10.3390/fermentation9070668
Chicago/Turabian StyleYao, Mingjing, Chunmin Ma, Xin Bian, Yang Yang, Yue Xu, Qiaoyan Wu, Xinyu Xu, Lulu Li, Na Zhang, and Yanjun Tian. 2023. "Isolation and Optimal Fermentation Conditions of Bacillus licheniformis SFD-Y5 for a New Douchi Fibrinolytic Enzyme Producer" Fermentation 9, no. 7: 668. https://doi.org/10.3390/fermentation9070668