
Fermentative L-Lactic Acid Production Using Bacillus coagulans from Corn Stalk Deconstructed by an Anaerobic Microbial Community
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
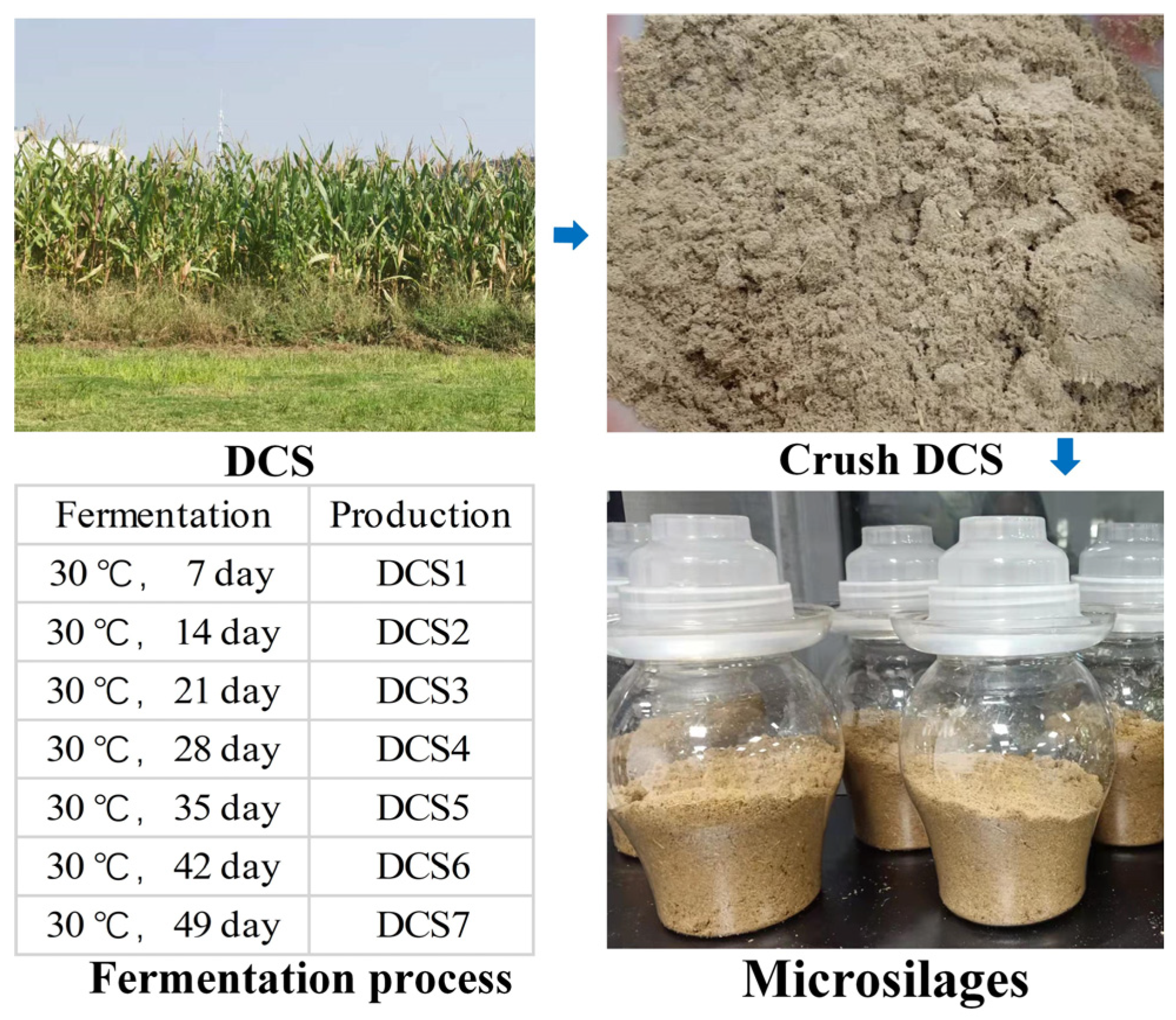
2.1. DCS and Bacterial Strains
2.2. Microbial Ensiling
2.3. Enzymatic Hydrolysis of Materials
2.4. LA Batch Fermentations
2.5. High-Throughput Sequencing Analysis
2.6. Chemical Composition of DCS Samples
2.7. Determination of Free Organic Acids and pH
2.8. HPLC Analysis Conditions
2.9. Optical Purity Calculation of LA
2.10. Characterization of DCS and Microsilage
2.11. Statistical Analysis
3. Results and Discussion
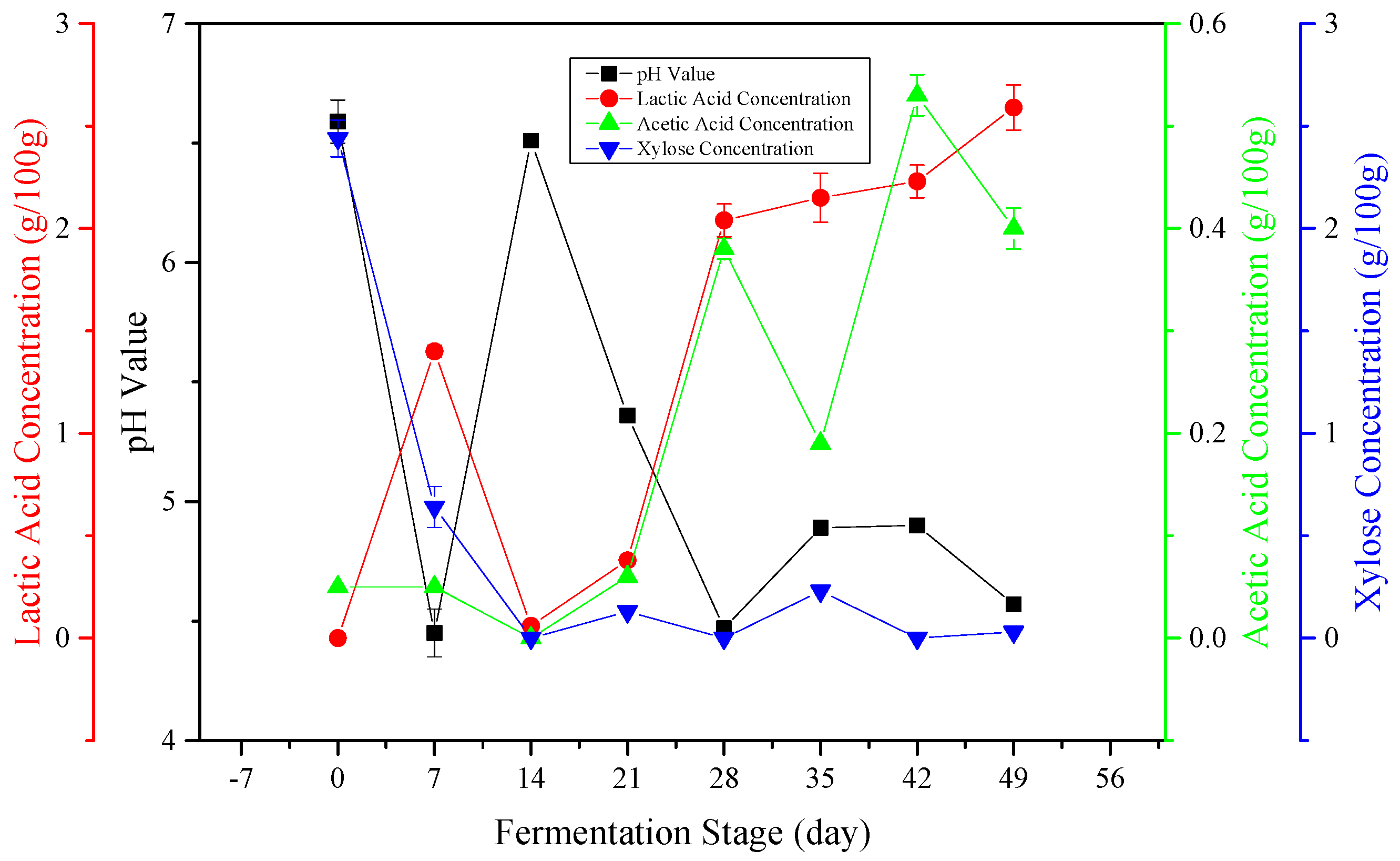
3.1. Characteristics of Microsilage
3.2. Effect of Ensiling on Carbohydrate Fractions of DCS
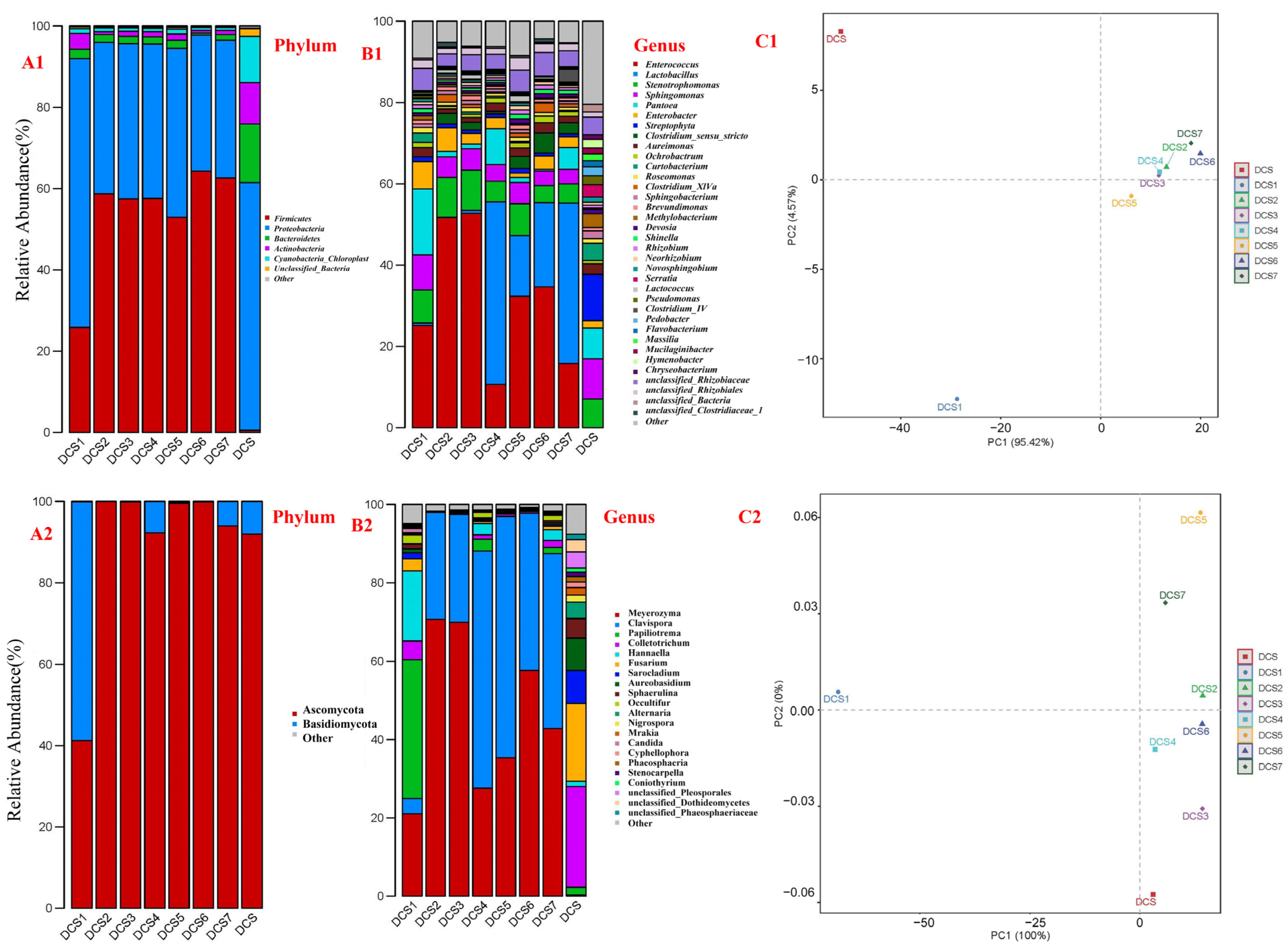
3.3. 16S rRNA Gene Sequencing Analysis of the Bacterial Community
3.4. ITS1–ITS2 Sequencing of the Fungal Community
3.5. Cell Wall of DCS Samples
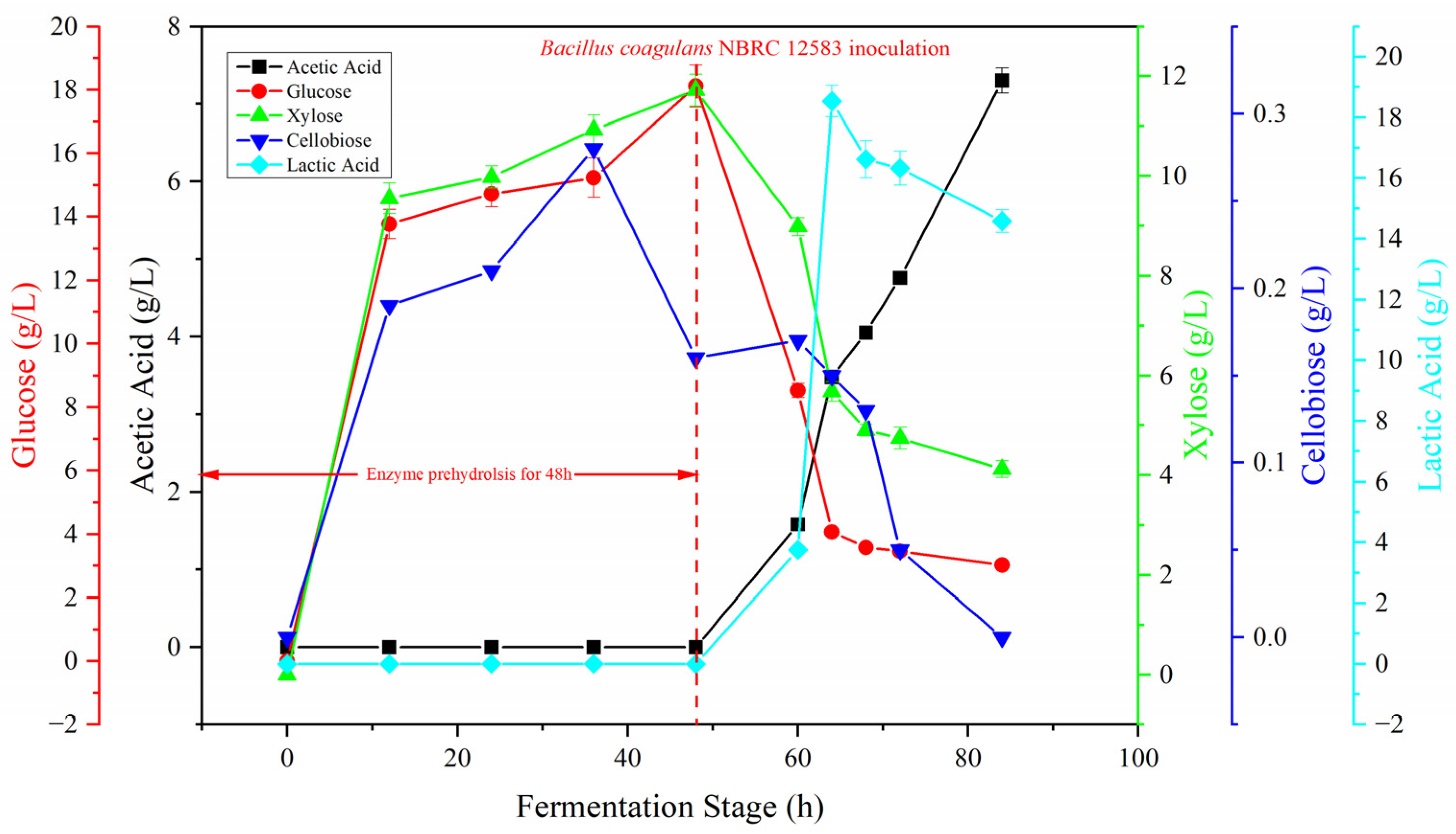
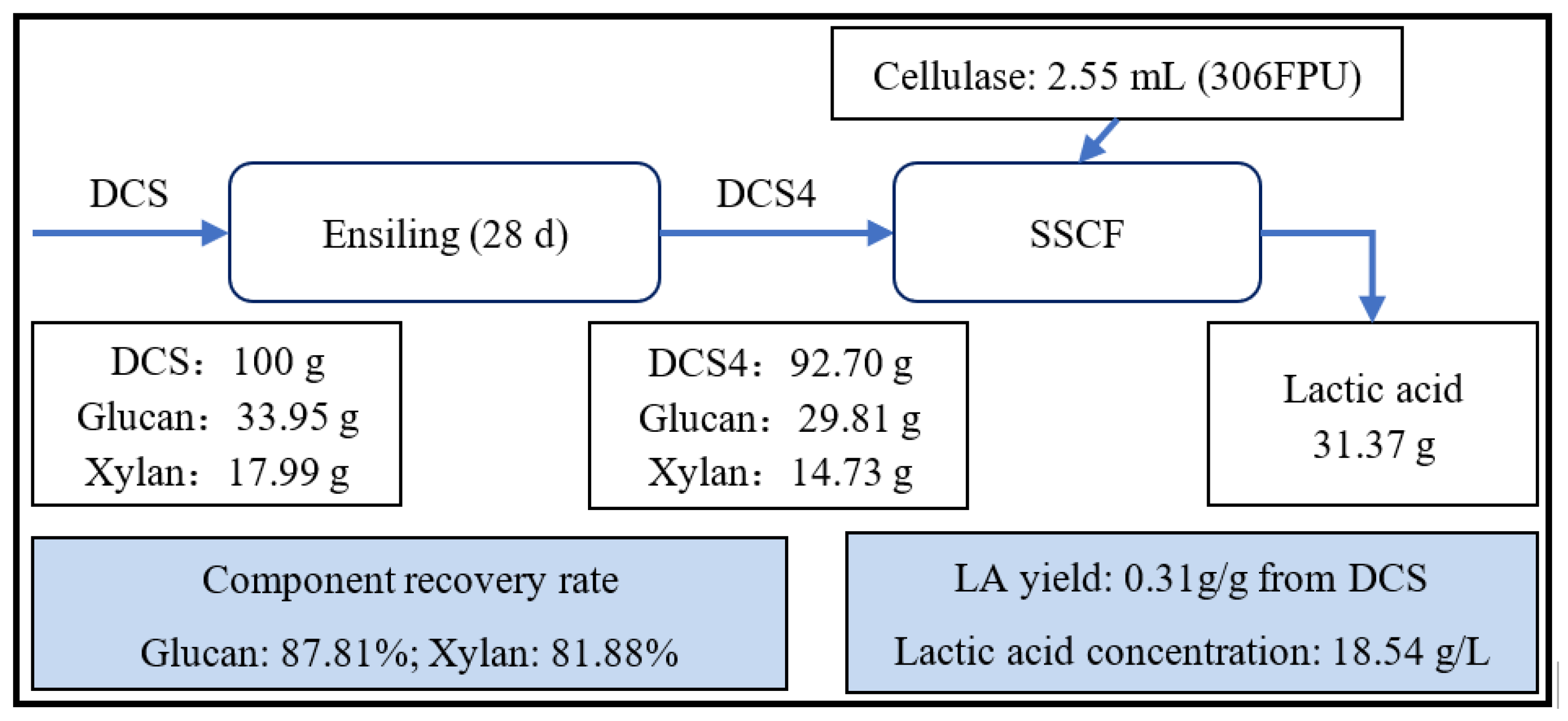
3.6. SSCF of Microsilage for LA Production
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Goldford, J.E.; Lu, N.X.; Bajic, D.; Estrela, S.; Tikhonov, M.; Sanchez-Gorostiaga, A.; Segre, D.; Mehta, P.; Sanchez, A. Emergent simplicity in microbial community assembly. Science 2018, 361, 469–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, X.J.; Chen, L.; Sui, Y.; Chen, C.; Zhang, W.M.; Zhou, J.; Dong, W.L.; Jiang, M.; Xin, F.X.; Ochsenreither, K. Biotechnological potential and applications of microbial consortia. Biotechnol. Adv. 2020, 40, 107500. [Google Scholar] [CrossRef] [PubMed]
- Reshmy, R.; Paulose, T.A.P.; Philip, E.; Thomas, D.; Madhavan, A.; Sirohi, R.; Binod, P.; Awasthi, M.K.; Pandey, A.; Sindhu, R. Updates on high value products from cellulosic biorefinery. Fuel 2022, 308, 122056. [Google Scholar] [CrossRef]
- Zhu, J.J.; Jiao, N.X.; Zhang, H.; Xu, G.L.; Xu, Y. Detoxification of lignocellulosic prehydrolyzate by lignin nanoparticles prepared from biorefinery biowaste to improve the ethanol production. Bioproc. Biosyst. Eng. 2022, 45, 1011–1018. [Google Scholar] [CrossRef] [PubMed]
- Sorbara, M.T.; Pamer, E.G. Microbiome-based therapeutics. Nat. Rev. Microbiol. 2022, 20, 365–380. [Google Scholar] [CrossRef]
- Zheng, M.Y.; Zhou, W.; Zou, X.; Wu, S.; Chen, X.Y.; Zhang, Q. Fermentation Quality and Bacterial Diversity of Broussonetia papyrifera Leaves Ensiled with Lactobacillus plantarum and Stored at Different Temperatures. Agronomy 2022, 12, 986. [Google Scholar] [CrossRef]
- You, L.J.; Bao, W.C.; Yao, C.Q.; Zhao, F.Y.; Jin, H.; Huang, W.Q.; Li, B.H.; Kwok, L.Y.; Liu, W.J. Changes in chemical composition, structural and functional microbiome during alfalfa (Medicago sativa) ensilage with Lactobacillus plantarum PS-8. Anim. Nutr. 2022, 9, 100–109. [Google Scholar] [CrossRef]
- Liu, Z.H.; Le, R.K.; Kosa, M.; Yang, B.; Yuan, J.S.; Ragauskas, A.J. Identifying and creating pathways to improve biological lignin valorization. Renew Sust. Energ. Rev. 2019, 105, 349–362. [Google Scholar] [CrossRef]
- Lee, J.S.A.; Baugh, A.C.; Shevalier, N.J.; Strand, B.; Stolyar, S.; Marx, C.J. Cross-Feeding of a Toxic Metabolite in a Synthetic Lignocellulose-Degrading Microbial Community. Microorganisms 2021, 9, 321. [Google Scholar] [CrossRef]
- Lin, L. Bottom-up synthetic ecology study of microbial consortia to enhance lignocellulose bioconversion. Biotechnol. Biofuels Bioprod. 2022, 15, 14. [Google Scholar] [CrossRef]
- Lillington, S.P.; Leggieri, P.A.; Heom, K.A.; O’Malley, M.A. Nature’s recyclers: Anaerobic microbial communities drive crude biomass deconstruction. Curr. Opin. Biotech. 2020, 62, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Jomnonkhaow, U.; Sittijunda, S.; Reungsang, A. Assessment of organosolv, hydrothermal, and combined organosolv and hydrothermal with enzymatic pretreatment to increase the production of biogas from Napier grass and Napier silage. Renew Energ. 2022, 181, 1237–1249. [Google Scholar] [CrossRef]
- Peng, X.; Wilken, S.E.; Lankiewicz, T.S.; Gilmore, S.P.; Brown, J.L.; Henske, J.K.; Swift, C.L.; Salamov, A.; Barry, K.; Grigoriev, I.V.; et al. Genomic and functional analyses of fungal and bacterial consortia that enable lignocellulose breakdown in goat gut microbiomes. Nat. Microbiol. 2021, 6, 499–511. [Google Scholar] [CrossRef]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D.; Crocker, D. Determination of Structural Carbohydrates and Lignin in Biomass; National Renewable Energy Laboratory: Boulder, CO, USA, 2011.
- Yang, X.; Li, H.L.; Chang, C.; Chen, J.Y.; Ma, X.J. The integrated process of microbial ensiling and hot-washing pretreatment of dry corn stover for ethanol production. Waste Biomass Valori. 2018, 9, 2031–2040. [Google Scholar] [CrossRef]
- Zheng, Y.; Lee, C.; Yu, C.; Cheng, Y.S.; Simmons, C.W.; Zhang, R.; Jenkins, B.M.; VanderGheynst, J.S. Ensilage and bioconversion of grape pomace into fuel ethanol. J. Agric. Food Chem. 2012, 60, 11128–11134. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.B.; de Jonge, N.; Dolriis, M.D.; Kragelund, C.; Fischer, C.H.; Eskesen, M.R.; Noer, K.; Moller, H.B.; Ottosen, L.D.M.; Nielsen, J.L.; et al. Cellulolytic and Xylanolytic Microbial Communities Associated with Lignocellulose-Rich Wheat Straw Degradation in Anaerobic Digestion. Front. Microbiol. 2021, 12, 645174. [Google Scholar] [CrossRef]
- Zhu, L.; O’Dwyer, J.P.; Chang, V.S.; Granda, C.B.; Holtzapple, M.T. Structural features affecting biomass enzymatic digestibility. Bioresour. Technol. 2008, 99, 3817–3828. [Google Scholar] [CrossRef]
- Wang, S.; Li, F.; Wu, D.; Zhang, P.; Wang, H.; Tao, X.; Ye, J.; Nabi, M. Enzyme Pretreatment Enhancing Biogas Yield from Corn Stover: Feasibility, Optimization, and Mechanism Analysis. J. Agric. Food Chem. 2018, 66, 10026–10032. [Google Scholar] [CrossRef]
- Wen, J.L.; Sun, S.N.; Yuan, T.Q.; Xu, F.; Sun, R.C. Fractionation of bamboo culms by autohydrolysis, organosolv delignification and extended delignification: Understanding the fundamental chemistry of the lignin during the integrated process. Bioresour. Technol. 2013, 150, 278–286. [Google Scholar] [CrossRef]
- He, L.W.; Lv, H.J.; Wang, C.; Zhou, W.; Pian, R.Q.; Zhang, Q.; Chen, X.Y. Dynamics of fermentation quality, physiochemical property and enzymatic hydrolysis of high-moisture corn stover ensiled with sulfuric acid or sodium hydroxide. Bioresour. Technol. 2020, 298, 122510. [Google Scholar] [CrossRef]
- Ren, H.; Feng, Y.; Pei, J.; Li, J.; Wang, Z.; Fu, S.; Zheng, Y.; Li, Z.; Peng, Z. Effects of Lactobacillus plantarum additive and temperature on the ensiling quality and microbial community dynamics of cauliflower leaf silages. Bioresour. Technol. 2020, 307, 123238. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.D.; Zhang, L.Q.; Sun, Y.; Xu, G.B.; Wang, W.D.; Piao, R.Z.; Cui, Z.J.; Zhao, H.Y. Degradation of lignocelluloses in straw using AC-1, a thermophilic composite microbial system. PeerJ 2021, 9, 12364. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.L.; Yuan, X.J.; Li, J.F.; Dong, Z.H.; Shao, T. Dynamics of microbial community and fermentation quality during ensiling of sterile and nonsterile alfalfa with or without Lactobacillus plantarum inoculant. Bioresour. Technol. 2019, 275, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Bai, J.; Huang, W.; Li, F.; Ke, W.; Zhang, Y.; Xie, D.; Zhang, B.; Guo, X. Characterization of a novel beta-cypermethrin-degrading strain of Lactobacillus pentosus 3–27 and its effects on bioremediation and the bacterial community of contaminated alfalfa silage. J. Hazard. Mater. 2022, 423, 127101. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Garcia, L.; Huang, S.X.; Sproer, C.; Sierra-Ramirez, R.; Bunk, B.; Overmann, J.; Jimenez, D.J. Dilution-to-Stimulation/Extinction Method: A Combination enrichment strategy to develop a minimal and versatile lignocellulolytic bacterial consortium. Appl. Environ. Microb. 2021, 87, e02427-20. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, D.J.; Korenblum, E.; van Elsas, J.D. Novel multispecies microbial consortia involved in lignocellulose and 5-hydroxymethylfurfural bioconversion. Appl. Microbiol. Biot. 2014, 98, 2789–2803. [Google Scholar] [CrossRef] [Green Version]
- Jard, G.; Dumas, C.; Delgenes, J.P.; Marfaing, H.; Sialve, B.; Steyer, J.P.; Carrere, H. Effect of thermochemical pretreatment on the solubilization and anaerobic biodegradability of the red macroalga Palmaria palmata. Biochem. Eng. J. 2013, 79, 253–258. [Google Scholar] [CrossRef]
- Li, P.P.; He, C.; Li, G.; Ding, P.; Lan, M.M.; Gao, Z.; Jiao, Y.Z. Biological pretreatment of corn straw for enhancing degradation efficiency and biogas production. Bioengineered 2020, 11, 251–260. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.B.; Wang, L.; Liu, D.H. Peracetic acid pretreatment of sugarcane bagasse for enzymatic hydrolysis: A continued work. J. Chem. Technol. Biot. 2008, 83, 950–956. [Google Scholar] [CrossRef]
- Xu, C.Y.; Ma, F.Y.; Zhang, X.Y.; Chen, S.L. Biological Pretreatment of Corn Stover by Irpex lacteus for Enzymatic Hydrolysis. J. Agr. Food Chem. 2010, 58, 10893–10898. [Google Scholar] [CrossRef]
- Wang, J.F.; Xin, D.L.; Hou, X.C.; Wu, J.Y.; Fan, X.F.; Li, K.N.; Zhang, J.H. Structural properties and hydrolysabilities of Chinese Pennisetum and Hybrid Pennisetum: Effect of aqueous ammonia pretreatment. Bioresour. Technol. 2016, 199, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.D.; Ye, L.D.; Wu, J.C. Efficient production of l-lactic acid by newly isolated thermophilic Bacillus coagulans WCP10-4 with high glucose tolerance. Appl. Microbiol. Biot. 2013, 97, 4309–4314. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Shi, S.A.; Wang, Y.; Jiang, Z.H. Integrated production of optically pure L-lactic acid from paper mill sludge by simultaneous saccharification and co-fermentation (SSCF). Waste Manag. 2021, 129, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Chacon, M.G.; Ibenegbu, C.; Leak, D.J. Simultaneous saccharification and lactic acid fermentation of the cellulosic fraction of municipal solid waste using Bacillus smithii. Biotechnol. Lett. 2021, 43, 667–675. [Google Scholar] [CrossRef]
- Zheng, Z.J.; Jiang, T.; Zou, L.H.; Ouyang, S.P.; Zhou, J.; Lin, X.; He, Q.; Wang, L.M.; Yu, B.; Xu, H.J.; et al. Simultaneous consumption of cellobiose and xylose by Bacillus coagulans to circumvent glucose repression and identification of its cellobiose-assimilating operons. Biotechnol. Biofuels 2018, 11, 320. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, S.; Zou, L.; Qiao, H.; Shi, J.; Zheng, Z.; Ouyang, J. One-pot process for lactic acid production from wheat straw by an adapted Bacillus coagulans and identification of genes related to hydrolysate-tolerance. Bioresour. Technol. 2020, 315, 123855. [Google Scholar] [CrossRef]
- Aulitto, M.; Fusco, S.; Nickel, D.B.; Bartolucci, S.; Contursi, P.; Franzen, C.J. Seed culture pre-adaptation of Bacillus coagulans MA-13 improves lactic acid production in simultaneous saccharification and fermentation. Biotechnol. Biofuels 2019, 12, 45. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.M.; Chen, X.R.; Luo, J.Q.; Qi, B.K.; Wan, Y.H. An efficient process for lactic acid production from wheat straw by a newly isolated Bacillus coagulans strain IPE22. Bioresour. Technol. 2014, 158, 396–399. [Google Scholar] [CrossRef]
- Ashokkumar, V.; Venkatkarthick, R.; Jayashree, S.; Chuetor, S.; Dharmaraj, S.; Kumar, G.; Chen, W.H.; Ngamcharussrivichai, C. Recent advances in lignocellulosic biomass for biofuels and value-added bioproducts—A critical review. Bioresour. Technol. 2022, 344, 126195. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cellulose | Hemicellulose | AIL | ASL | Ash | |
|---|---|---|---|---|---|
| DCS | 33.95 ± 0.93 | 17.99 ± 0.55 | 27.72 ± 0.79 | 2.71 ± 0.03 | 2.25 ± 0.01 |
| DCS1 | 33.31 ± 0.86 | 17.57 ± 0.63 | 27.14 ± 0.85 | 3.10 ± 0.03 | 2.23 ± 0.01 |
| DCS2 | 33.71 ± 0.97 | 17.39 ± 0.47 | 28.48 ± 0.91 | 3.15 ± 0.02 | 2.56 ± 0.03 |
| DCS3 | 32.65 ± 0.73 | 16.98 ± 0.51 | 28.66 ± 0.73 | 3.06 ± 0.03 | 2.45 ± 0.01 |
| DCS4 | 32.16 ± 0.82 | 15.89 ± 0.33 | 27.41 ± 0.56 | 2.97 ± 0.08 | 2.32 ± 0.06 |
| DCS5 | 32.85 ± 0.96 | 15.65 ± 0.41 | 28.65 ± 0.81 | 3.08 ± 0.02 | 2.46 ± 0.02 |
| DCS6 | 32.66 ± 0.63 | 15.14 ± 0.45 | 27.53 ± 0.63 | 3.15 ± 0.07 | 2.48 ± 0.02 |
| DCS7 | 32.12 ± 0.76 | 14.85 ± 0.21 | 26.99 ± 0.58 | 3.15 ± 0.02 | 2.45 ± 0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Shi, Z.; Wang, T.; Meng, X.; Song, L.; Zhang, Z.; Zhang, J.; Wei, T. Fermentative L-Lactic Acid Production Using Bacillus coagulans from Corn Stalk Deconstructed by an Anaerobic Microbial Community. Fermentation 2023, 9, 611. https://doi.org/10.3390/fermentation9070611
Yang X, Shi Z, Wang T, Meng X, Song L, Zhang Z, Zhang J, Wei T. Fermentative L-Lactic Acid Production Using Bacillus coagulans from Corn Stalk Deconstructed by an Anaerobic Microbial Community. Fermentation. 2023; 9(7):611. https://doi.org/10.3390/fermentation9070611
Chicago/Turabian StyleYang, Xu, Zhiyuan Shi, Tongyu Wang, Xiangyu Meng, Lili Song, Zhiping Zhang, Jingnan Zhang, and Tao Wei. 2023. "Fermentative L-Lactic Acid Production Using Bacillus coagulans from Corn Stalk Deconstructed by an Anaerobic Microbial Community" Fermentation 9, no. 7: 611. https://doi.org/10.3390/fermentation9070611