

Effects of Phenyllactic Acid on Fermentation Parameters, Nitrogen Fractions and Bacterial Community of High-Moisture Stylo Silage


Abstract
:1. Introduction
2. Materials and Methods
2.1. Raw Material Preparation and Silage Processing
2.2. Determination of Microbial Population, Fermentation Parameters and Chemical Composition
2.3. Bacterial Community Analysis by 16S rDNA Sequencing Technology
2.4. Statistical Analysis
3. Results
3.1. Effect of Phenyllactic Acid on Dynamic Fermentation Characteristics of Stylo Silage
3.2. Effect of Phenyllactic Acid on Dynamic Nitrogen Distribution of Stylo Silage
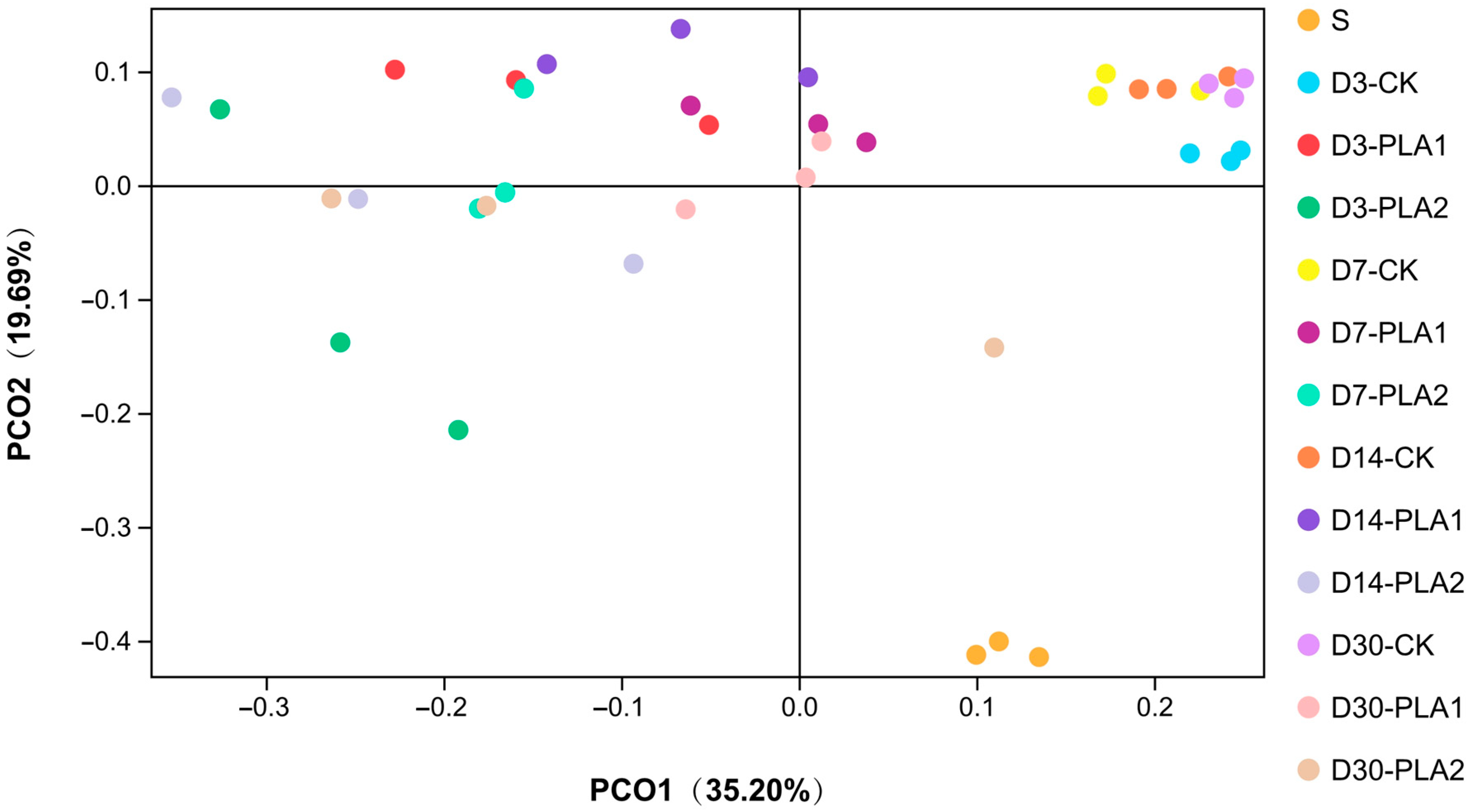
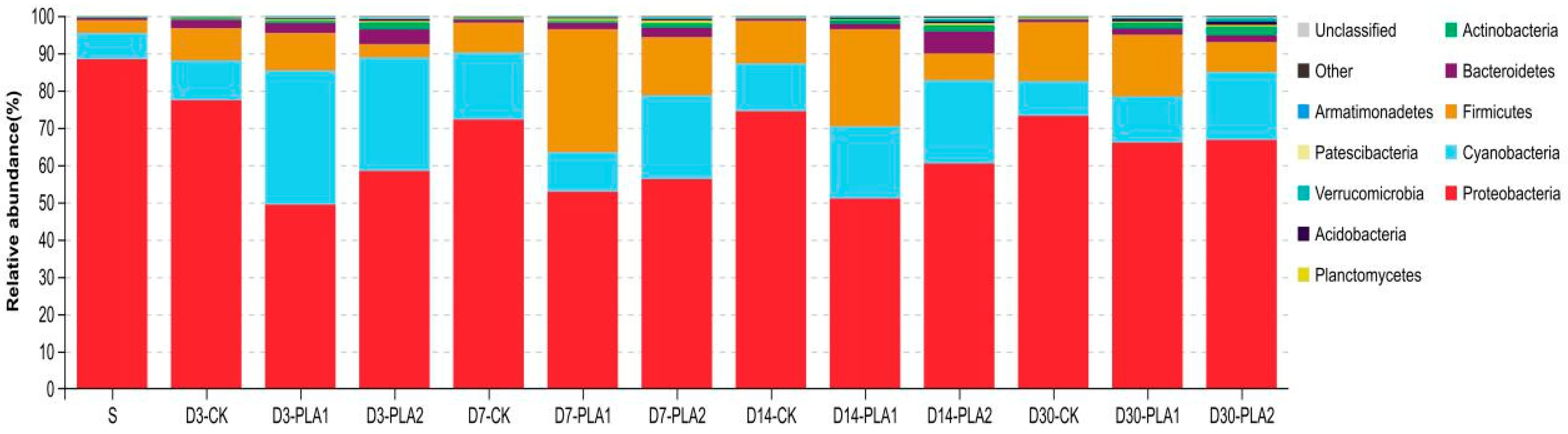
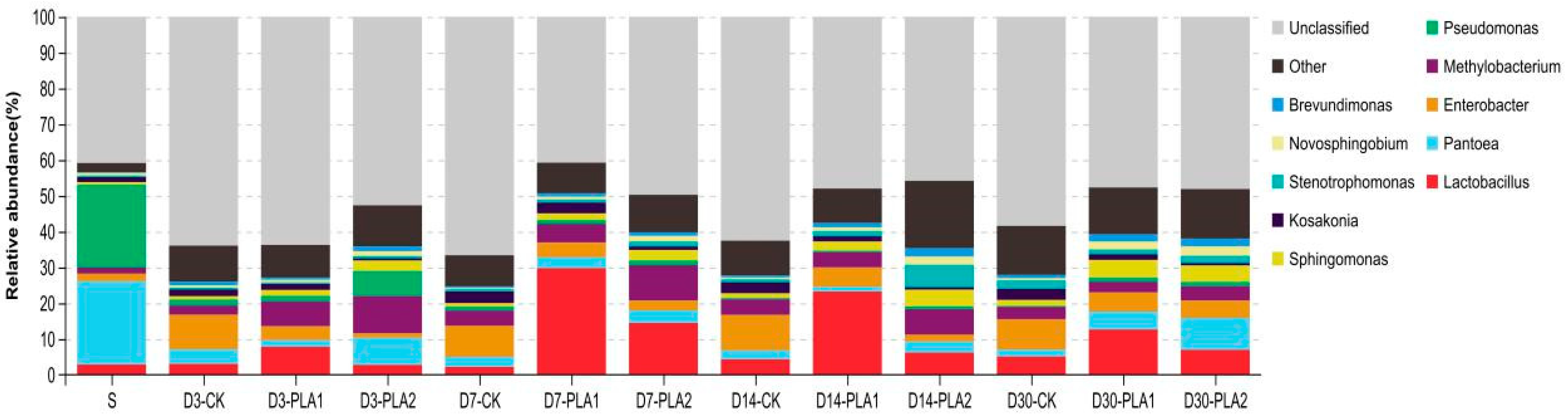
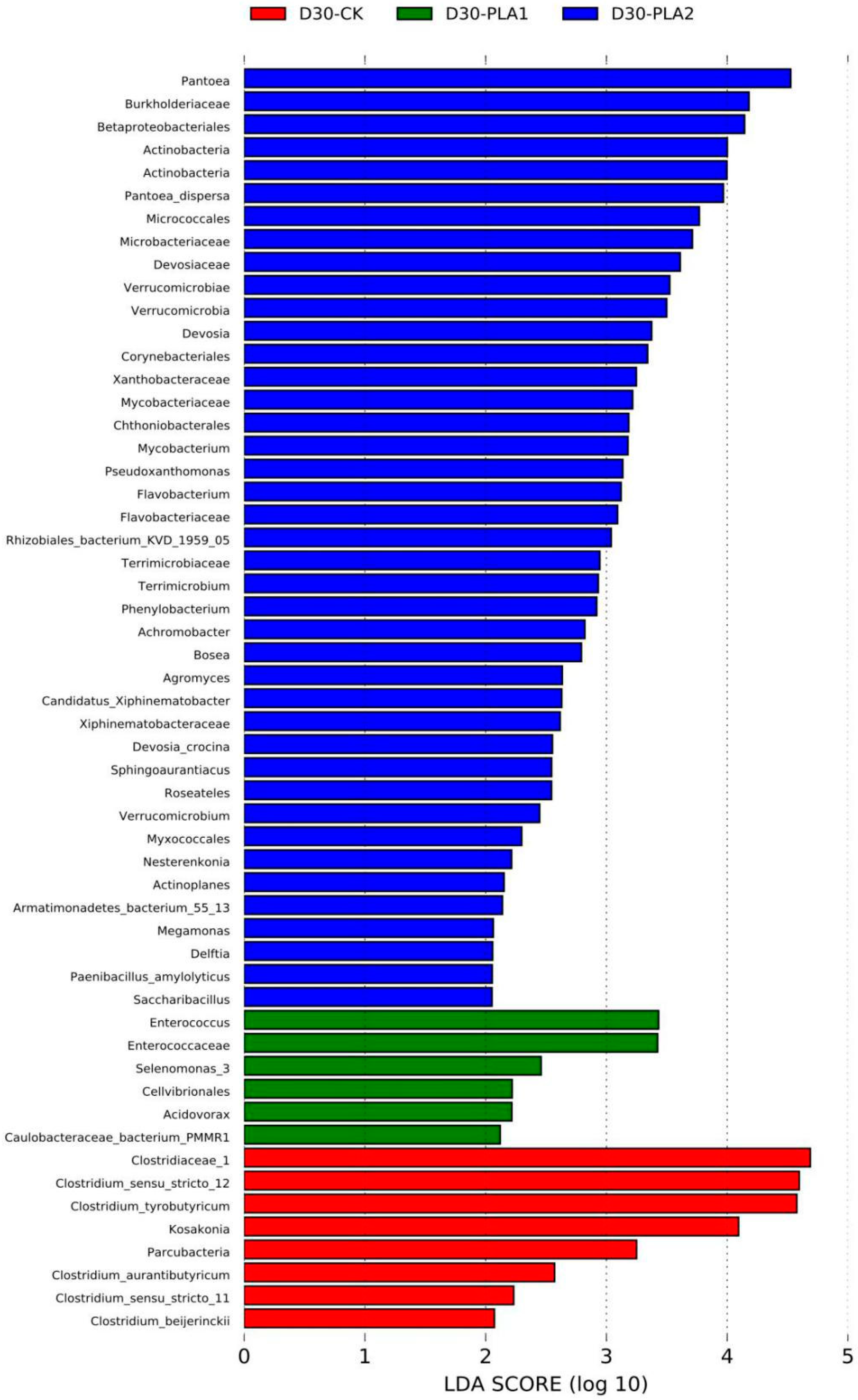
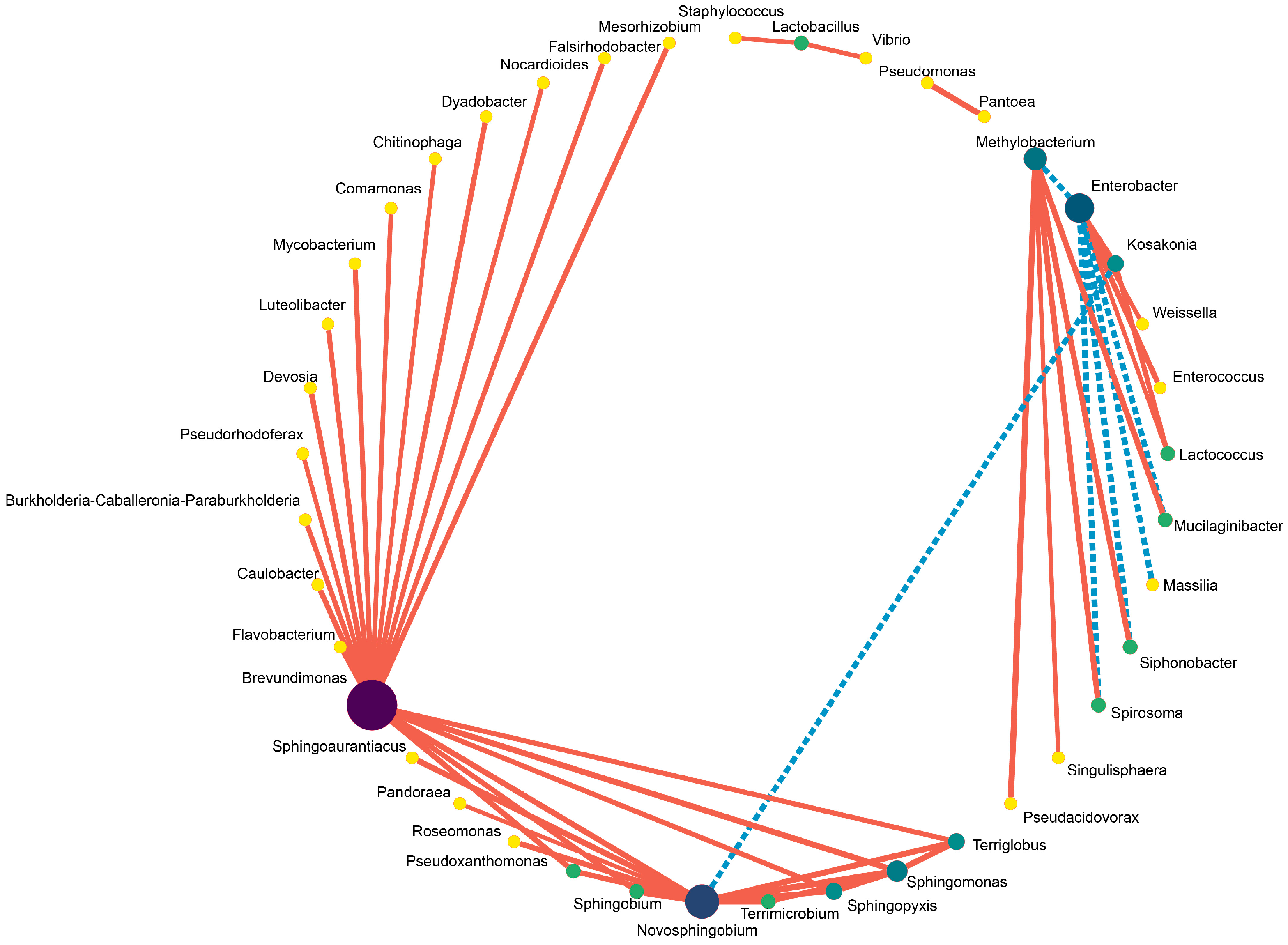
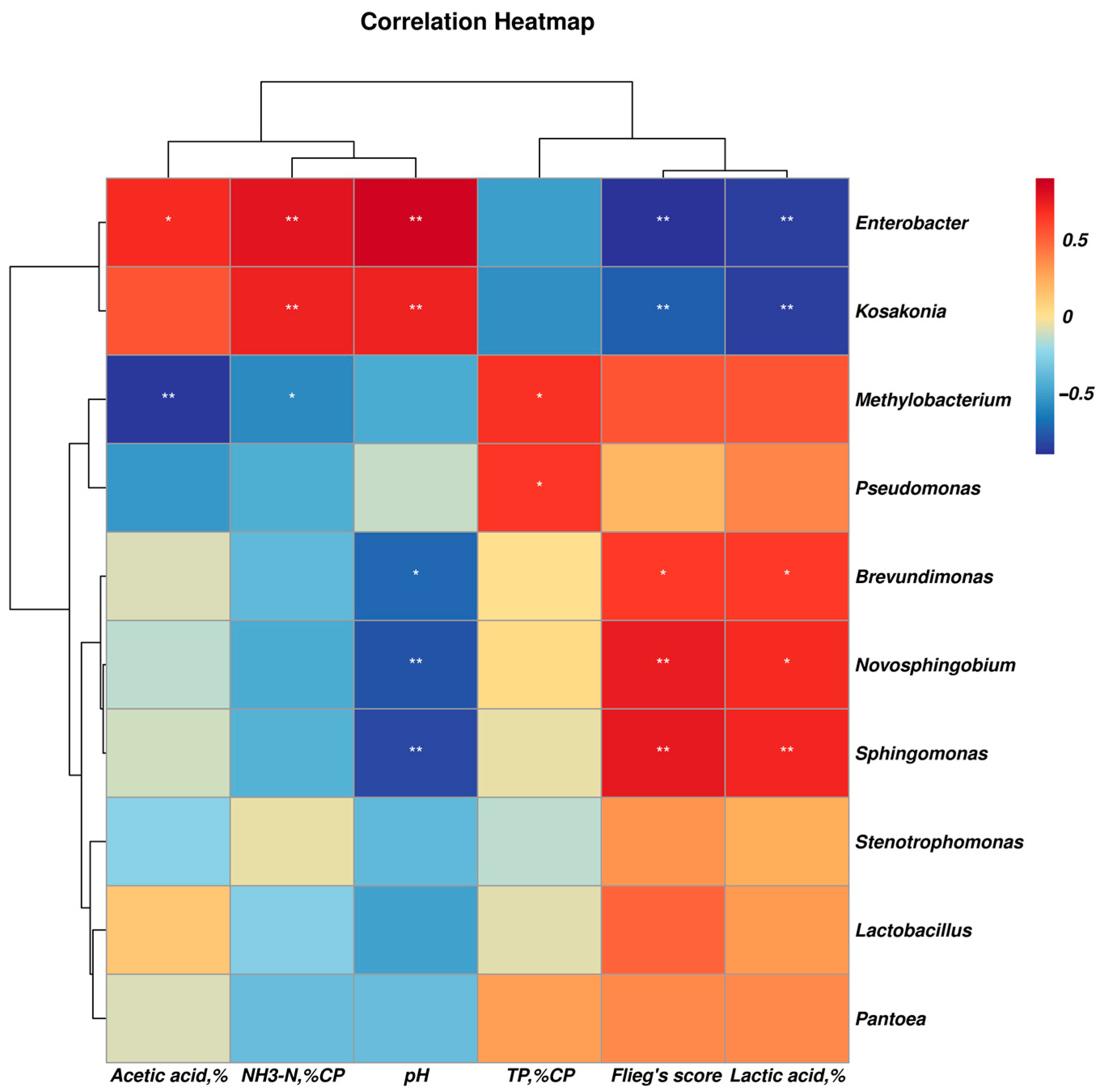
3.3. Effect of Phenyllactic Acid on Dynamic Bacterial Community of Stylo Silage
4. Discussion
4.1. General Characteristics of Fresh Stylo Used for Silage Production
4.2. Effect of Phenyllactic Acid on Dynamic Fermentation Characteristics of Stylo Silage
4.3. Effect of Phenyllactic Acid on Dynamic Nitrogen Distribution of Stylo Silage
4.4. Effect of Phenyllactic Acid on Dynamic Bacterial Community of Stylo Silage
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- HeuHeuzé, V.; Tran, G.; Boudon, A.; Labussière, E.; Bastianelli, D.; Lebas, F. Stylo (Stylosanthes guianensis). Feedipedia, a Programme by INRAE, CIRAD, AFZ and FAO. 2017. Available online: https://www.feedipedia.org/node/251 (accessed on 15 May 2023).
- Rajanikar, R.V.N.B. Phenyllactic acid: A green compound for food biopreservation. Food Control 2021, 128, 108184. [Google Scholar] [CrossRef]
- He, L.; Lv, H.; Xing, Y.; Wang, C.; You, X.; Chen, X.; Zhang, Q. The nutrients in Moringa oleifera leaf contribute to the improvement of stylo and alfalfa silage: Fermentation, nutrition and bacterial community. Bioresour. Technol. 2020, 301, 122733. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Li, S.; Wang, C.; Chen, X.; Zhang, Q. Effects of vanillic acid on dynamic fermentation parameter, nitrogen distribution, bacterial community, and enzymatic hydrolysis of stylo silage. Front. Microbiol. 2021, 12, 690801. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Chen, N.; Lv, H.; Wang, C.; Zhou, W.; Chen, X.; Zhang, Q. Gallic acid influencing fermentation quality, nitrogen distribution and bacterial community of high-moisture mulberry leaves and stylo silage. Bioresour. Technol. 2020, 295, 122255. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Li, Z.; Yu, Z.; Zhang, Q.; Li, X. Interactive effect of inoculant and dried jujube powder on the fermentation quality and nitrogen fraction of alfalfa silage. Anim. Sci. J. 2017, 88, 633–642. [Google Scholar] [CrossRef]
- Guo, J.; Xie, Y.; Yu, Z.; Meng, G.; Wu, Z. Effect of Lactobacillus plantarum expressing multifunctional glycoside hydrolases on the characteristics of alfalfa silage. Appl. Microbiol. Biotechnol. 2020, 19, 7983–7995. [Google Scholar] [CrossRef]
- Wang, Y.; He, L.; Xing, Y.; Zheng, Y.; Zhou, W.; Pian, R.; Yang, F.; Chen, X.; Zhang, Q. Dynamics of bacterial community and Dynamics of bacterial community and fermentation quality during ensiling of wilted and unwilted Moringa oleifera leaf silage with or without lactic acid bacterial inoculants. Msphere 2019, 4, e00341-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, D.; Dong, Z.; Wang, D.; Li, B.; Shi, P.; Yan, J.; Zhuang, D.; Shao, T.; Wang, W.; Gu, M. Evaluating the fermentation quality and bacterial community of high-moisture whole-plant quinoa silage ensiled with different additives. J. Appl. Microbiol. 2022, 132, 3578–3589. [Google Scholar] [CrossRef]
- Franco, M.; Ilma, T.; Arto, H.; Marketta, R. Fermentation quality and bacterial ecology of red clover dominated silage modulated by different management factors. Front. Anim. Sci. 2022, 3, 1080535. [Google Scholar] [CrossRef]
- Zhou, J. Effects of different additives on fermentation quality, microbial communities, and rumen degradation of alfalfa silage. Fermentation 2022, 8, 660–669. [Google Scholar]
- Wu, Z.; Xu, S.; Yun, Y.; Jia, T.; Yu, Z. Effect of 3-phenyllactic acid and 3-phenyllactic acid-producing lactic acid bacteria on the characteristics of alfalfa silage. Agriculture 2019, 10, 10. [Google Scholar] [CrossRef] [Green Version]
- Dieuleveux, V.; Lemarinier, S.; Guéguen, M. Antimicrobial spectrum and target site of D-3-phenyllactic acid. Int. J. Food Microbiol. 1998, 40, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Mu, W.; Yu, S.; Zhu, L.; Zhang, T.; Jiang, B. Recent research on 3-phenyllactic acid, a broad-spectrum antimicrobial compound. Appl. Microbiol. Biotech. 2012, 95, 1155–1163. [Google Scholar] [CrossRef]
- Prema, P.; Smila, D.; Palavesam, A.; Immanuel, G. Production and characterization of an antifungal compound (3-phenyllactic acid) produced by Lactobacillus plantarum strain. Food Bioprocess Technol. 2010, 3, 379–386. [Google Scholar] [CrossRef]
- Yanina, B.A.; Graciela, F.; Luciana, G.C. Optimization of phenyllactic acid production by Pediococcus acidilactici CRL 1753. Application of the formulated bio-preserver culture in bread. Biol. Control 2018, 123, 137–143. [Google Scholar]
- Li, P.; Lu, Y.; Zhao, M.; Chen, L.; Zhang, C.; Cheng, Q.; Chen, C. Effects of phenyllactic acid, lactic acid bacteria, and their mixture on fermentation characteristics and microbial community composition of timothy silage. Front. Microbiol. 2021, 12, 743433. [Google Scholar] [CrossRef] [PubMed]
- Broderick, G.A.; Kang, J.H. Automated simultaneous determination of ammonia and total amino acids in ruminal fluid and in vitro media. J. Dairy Sci. 1980, 63, 64–75. [Google Scholar] [CrossRef]
- He, L.; Zhou, W.; Wang, Y.; Wang, C.; Chen, X.; Zhang, Q. Effect of applying lactic acid bacteria and cellulase on the fermentation quality, nutritive value, tannins profile and in vitro digestibility of Neolamarckia cadamba leaves silage. J. Anim. Physiol. Anim. Nutr. 2018, 102, 1429–1436. [Google Scholar] [CrossRef]
- Licitra, G.; Hernandez, T.M.; Van Soest, P.J. Standardization of procedures for nitrogen fractionation of ruminant feeds. Anim. Feed Sci. Technol. 1996, 57, 347–358. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- MOA. Determination of soluble sugar in fruits and derived products—3,5-dinitrosalicylic acid colorimetry. In Agricultural Industry Standard of PRC NY/T 2742-2015; The Ministry of Agriculture: Beijing, China, 2015. [Google Scholar]
- Flieg, O. A key for the evaluation of silage samples. Futterb. Garfutterber. 1938, 1, 112–128. [Google Scholar]
- Wang, C.; He, L.; Xing, Y.; Zhou, W.; Yang, F.; Chen, X.; Zhang, Q. Fermentation quality and microbial community of alfalfa and stylo silage mixed with Moringa oleifera leaves. Bioresour. Technol. 2019, 284, 240–247. [Google Scholar] [CrossRef] [PubMed]
- NASEM. Nutrient Requirements of Beef Cattle: Eighth Revised Edition. In National Academies of Sciences, Engineering and Medicine; The National Academies Press: Washington, DC, USA, 2016. [Google Scholar]
- NRC. Nutrient Requirements of Small Ruminants: Sheep, Goats, Cervids, and New World Camelids; National Academy of Science: Washintgton, DC, USA, 2006. [Google Scholar]
- Mcdonald, P.; Henderson, A.R.; Heron, S.J.E. The Biochemistry of Silage; Chalcombe Publications: Southampton, UK, 1991. [Google Scholar]
- Lin, C.; Bolsen, K.K.; Brent, B.E.; Hart, R.A.; Dickerson, J.T.; Feyerherm, A.M.; Aimutis, W.R. Epiphytic microflora on alfalfa and whole-plant corn. J. Dairy Sci. 1992, 75, 2484–2493. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.H. Theoretical carbohydrate requirement for alfalfa silage production. Agron. J. 1962, 54, 291–293. [Google Scholar] [CrossRef]
- Guyader, J.; Baron, V.; Beauchemin, K. Corn forage yield and quality for silage in short growing season areas of the Canadian prairies. Agronomy 2018, 8, 164. [Google Scholar] [CrossRef] [Green Version]
- Pahlow, G.; Muck, R.E.; Driehuis, F.; Elferink, S.J.W.H.; Spoelstra, S.F. Microbiology of Ensiling. In Silage Science and Technology; Buxton, D.R., Muck, R.E., Harrison, J.H., Eds.; American Society of Agronomy, Inc.: Madison, WI, USA; Crop Science Society of America, Inc.: Madison, WI, USA; Soil Science Society of America, Inc. Publications: Madison, WI, USA, 2003; pp. 31–93. [Google Scholar]
- Kung, L.; Shaver, R.D.; Grant, R.J.; Schmidt, R.J. Silage review: Interpretation of chemical, microbial, and organoleptic components of silages. J. Dairy Sci. 2018, 101, 4020–4033. [Google Scholar] [CrossRef]
- Hutkins, R.W.; Nannen, N.L. pH Homeostasis in Lactic Acid Bacteria 1. J. Dairy Sci. 1993, 76, 2354–2365. [Google Scholar] [CrossRef]
- Kung, L.; Stokes, M.R.; Lin, C.J. Silage Additives. Silage Sci. Technol. 2003, 42, 305–360. [Google Scholar]
- Muck, R. Silage Microbiology and Its Control through Additives. Revista Brasileira Zootecnia 2010, 39, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Tabacco, E.; Borreani, G.; Crovetto, G.M.; Galassi, G.; Colombo, D.; Cavallarin, L. Effect of chestnut tannin on fermentation quality, proteolysis, and protein rumen degradability of alfalfa silage. J. Dairy Sci. 2006, 89, 4736–4746. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yang, J.; Wang, J.; Zheng, N.; Li, S.; Zhao, S.; Wen, F.; Wang, J. Progress assessment of chemical indicators of silage. Chin. J. Anim. Husb. 2016, 52, 37–42. [Google Scholar]
- Grice, E.A.; Kong, H.H.; Conlan, S.; Deming, C.B.; Davis, J.; Young, A.C.; Segre, J.A. Topographical and temporal diversity of the human skin microbiome. Science 2009, 324, 1190–1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogunade, I.M.; Jiang, Y.; Cervantes, A.P.; Kim, D.H.; Oliveira, A.S.; Vyas, D.; Weinberg, Z.G.; Jeong, K.C.; Adesogan, A.T. Bacterial diversity and composition of alfalfa silage as analyzed by Illumina MiSeq sequencing: Effects of Escherichia coli O157: H7 and silage additives. J. Dairy Sci. 2018, 101, 2048–2059. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Qiu, C.; Wang, Y.; Zhang, W.; He, L. Effect of tea polyphenols on the fermentation quality, protein preservation, antioxidant capacity and bacterial community of stylo silage. Front. Microbiol. 2022, 13, 993750. [Google Scholar] [CrossRef] [PubMed]
- Guan, H.; Yan, Y.; Li, X.; Li, X.; Shuai, Y.; Feng, G.; Ran, Q.; Cai, Y.; Li, Y.; Zhang, X. Microbial communities and natural fermentation of corn silages prepared with farm bunker-silo in Southwest China. Bioresour. Technol. 2018, 265, 282–290. [Google Scholar] [CrossRef]
- Brenner, D.J.; Krieg, N.R.; Staley, J.T. The proteobacteria, volume two, part B, the gammaproteo bacteria. In Berge’s Manual of Systematic Bacteriology, 2nd ed.; Garrity, G.M., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2005; pp. 382–400. [Google Scholar]
- Li, X.; Li, Y.; Li, Y.; Wu, J. Myriophyllum elatinoides growth and rhizosphere bacterial community structure under different nitrogen concentrations in swine wastewater. Bioresour. Technol. 2020, 301, 122776. [Google Scholar] [CrossRef]
- Nie, Y.; Wang, M.; Zhang, W.; Ni, Z.; Hashidoko, Y.; Shen, W. Ammonium nitrogen content is a dominant predictor of bacterial community composition in an acidic forest soil with exogenous nitrogen enrichment. Sci. Total Environ. 2018, 624, 407–415. [Google Scholar] [CrossRef]
- Li, Y. Classification of Bacteria of the Genus Kosakonia Based on Genome-Wide Sequence Systems. Master’s Thesis, Zhejiang University, Hangzhou, China, 2016. [Google Scholar]
- Doronina, N.V.; Trotsenko, Y.A.; Kuznetsov, B.B.; Tourova, T.P.; Salkinoja-Salonen, M.S. Methylobacterium suomiense sp. nov. and Methylobacterium lusitanum sp. nov., aerobic, pink-pigmented, facultatively methylotrophic bacteria. Int. J. Syst. Evol. Microbiol. 2002, 52, 773–776. [Google Scholar]
- White, D.C.; Sutton, S.D.; Ringelberg, D.B. The genus Sphingomonas: Physiology and ecology. Curr. Opin. Biotech. 1996, 7, 301–306. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, Y.; Guo, J.; Shen, Y.; Yang, J. The correlations and spatial characteristics of microbiome and silage quality by reusing of citrus waste in a family-scale bunker silo. J. Clean. Prod. 2019, 226, 407–418. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Fresh Stylo | Item | Fresh Stylo |
|---|---|---|---|
| Dry matter, % | 24.82 | Lactic acid bacteria, log10 CFU/g FM | 6.05 |
| Crude protein, % DM | 13.35 | Yeast, log10 CFU/g FM | 3.88 |
| True protein, % DM | 10.96 | Mold, log10 CFU/g FM | 3.71 |
| Nonprotein-nitrogen, % DM | 2.38 | Coliform bacteria, log10 CFU/g FM | 6.75 |
| True protein proportion, % CP | 82.13 | Sobs | 1291 |
| Nonprotein-nitrogen, % CP | 17.87 | Shannon | 5.86 |
| Neutral detergent fiber, % DM | 53.06 | Simpson | 0.96 |
| Acid detergent fiber, % DM | 44.66 | Chao | 2087 |
| Acid detergent lignin, % DM | 7.33 | Ace | 2171 |
| Water soluble carbohydrate, % DM | 1.54 | Coverage | 0.9926 |
| Item | Trt | Days of Ensiling | SEM | p-Value | Contrast | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 3 | 7 | 14 | 30 | D | T | D × T | L | Q | |||
| Dry matter, % | CK | 25.20 Ca | 25.67 Ca | 24.29 Bb | 21.59 Cc | 0.21 | <0.01 | <0.01 | 0.04 | <0.01 | 0.36 |
| 1% | 26.40 Ba | 26.43 Ba | 25.41 Ab | 23.14 Bc | |||||||
| 2% | 27.42 Aa | 27.75 Aa | 25.56 Ab | 24.29 Ac | |||||||
| Dry matter loss, % | CK | - | - | - | 15.36 A | 0.85 | - | <0.01 | - | <0.01 | 0.65 |
| 1% | - | - | - | 9.05 B | |||||||
| 2% | - | - | - | 4.69 C | |||||||
| pH | CK | 5.74 | 5.60 | 5.66 | 5.43 | 0.06 | <0.01 | <0.01 | 0.23 | <0.01 | <0.01 |
| 1% | 4.92 | 4.66 | 4.57 | 4.50 | |||||||
| 2% | 4.71 | 4.33 | 4.41 | 4.32 | |||||||
| Lactic acid bacteria, log10 CFU/g FM | CK | 8.23 Aa | 8.14 Aa | 7.93 Bb | 7.60 Ac | 0.09 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| 1% | 7.63 Bb | 8.31 Aa | 8.19 Aa | 7.62 Ab | |||||||
| 2% | 6.59 Cc | 7.75 Ba | 7.12 Cb | 6.68 Bc | |||||||
| Coliform bacteria, log10 CFU/g FM | CK | 7.58 Aa | 7.51 Aa | 5.56 b | 2.00 c | 0.14 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| 1% | 6.62 B | 4.44 B | <2.00 | <2.00 | |||||||
| 2% | 5.99 C | 4.50 B | <2.00 | <2.00 | |||||||
| Lactic acid, % DM | CK | 0.73 Ba | 0.40 Cb | 0.26 Bc | 0.22 Cc | 0.09 | 0.11 | <0.01 | <0.01 | <0.01 | <0.01 |
| 1% | 1.09 ABab | 0.87 Bb | 1.17 Aab | 1.36 Aa | |||||||
| 2% | 1.33 A | 1.37 A | 1.34 A | 1.03 B | |||||||
| Acetic acid, % DM | CK | 0.35 | 0.44 | 0.58 | 0.65 | - | - | - | - | - | - |
| 1% | 0.20 | 0.41 | 0.56 | 0.63 | |||||||
| 2% | ND | ND | ND | 0.60 | |||||||
| Butyric acid, % DM | CK | ND | ND | 0.41 | 0.76 | - | - | - | - | - | - |
| 1% | ND | ND | ND | ND | |||||||
| 2% | ND | ND | ND | ND | |||||||
| Flieg’s score | CK | 25.67 | 32.47 | 27.05 | 30.98 | 2.50 | <0.01 | <0.01 | 0.23 | <0.01 | <0.01 |
| 1% | 61.01 | 71.46 | 73.01 | 71.41 | |||||||
| 2% | 71.57 | 87.17 | 79.85 | 80.64 | |||||||
| Item | Trt | Days of Ensiling | SEM | p-Value | Contrast | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 3 | 7 | 14 | 30 | D | T | D × T | L | Q | |||
| CP, % DM | CK | 12.88 | 14.06 | 13.25 | 12.79 | 0.30 | 0.12 | 0.20 | 0.33 | 0.13 | 0.31 |
| 1% | 13.38 | 13.60 | 13.00 | 13.04 | |||||||
| 2% | 12.60 | 12.98 | 13.15 | 12.99 | |||||||
| TP, % DM | CK | 8.72 | 8.14 | 7.38 | 6.48 | 0.21 | <0.01 | 0.02 | 0.13 | <0.01 | 0.28 |
| 1% | 8.58 | 8.24 | 7.44 | 7.01 | |||||||
| 2% | 8.76 | 8.46 | 8.15 | 7.63 | |||||||
| NH3-N, % DM | CK | 0.71 Ad | 1.29 Ac | 1.76 Ab | 2.37 Aa | 0.04 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| 1% | 0.22 Bd | 0.48 Bc | 0.66 Bb | 0.83 Ba | |||||||
| 2% | 0.13 Cc | 0.20 Cbc | 0.28 Cb | 0.49 Ca | |||||||
| TP, % CP | CK | 66.75 | 57.87 | 55.70 | 50.66 | 1.33 | <0.01 | <0.01 | 0.42 | <0.01 | 0.10 |
| 1% | 64.24 | 60.70 | 58.50 | 53.77 | |||||||
| 2% | 69.48 | 65.23 | 61.98 | 58.85 | |||||||
| NH3-N, % CP | CK | 5.55 Ad | 9.16 Ac | 13.28 Ab | 18.53 Aa | 0.30 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| 1% | 1.65 Bd | 3.57 Bc | 5.06 Bb | 6.38 Ba | |||||||
| 2% | 1.05 Bd | 1.55 Cc | 2.13 Cb | 3.74 Ca | |||||||
| Item | Trt | Days of Ensiling | SEM | p-Value | Contrast | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 3 | 7 | 14 | 30 | D | T | D × T | L | Q | |||
| Sobs | CK | 1460 | 1387 | 1339 | 1369 | 50 | 0.02 | <0.01 | 0.10 | <0.01 | 0.37 |
| 1% | 1618 | 1495 | 1442 | 1407 | |||||||
| 2% | 1649 | 1771 | 1689 | 1539 | |||||||
| Shannon | CK | 5.73 | 5.52 | 5.76 | 5.91 | 0.26 | 0.01 | <0.01 | 0.47 | <0.01 | 0.17 |
| 1% | 5.56 | 6.55 | 6.37 | 6.82 | |||||||
| 2% | 6.07 | 6.60 | 6.76 | 6.78 | |||||||
| Simpson | CK | 0.93 | 0.92 | 0.94 | 0.95 | 0.02 | 0.03 | 0.73 | 0.49 | 0.54 | 0.64 |
| 1% | 0.88 | 0.97 | 0.95 | 0.97 | |||||||
| 2% | 0.91 | 0.95 | 0.95 | 0.96 | |||||||
| Chao | CK | 2312 | 2186 | 2195 | 2176 | 74 | 0.04 | 0.03 | 0.38 | 0.02 | 0.10 |
| 1% | 2410 | 2244 | 2105 | 2091 | |||||||
| 2% | 2412 | 2508 | 2385 | 2321 | |||||||
| Ace | CK | 2376 | 2251 | 2277 | 2191 | 74 | <0.01 | 0.13 | 0.14 | 0.12 | 0.16 |
| 1% | 2463 | 2294 | 2139 | 2085 | |||||||
| 2% | 2367 | 2538 | 2378 | 2245 | |||||||
| Coverage | CK | 0.9928 | 0.9934 | 0.9929 | 0.9935 | 0.0003 | 0.02 | 0.05 | 0.20 | 0.47 | 0.02 |
| 1% | 0.9927 | 0.9921 | 0.9923 | 0.9932 | |||||||
| 2% | 0.9932 | 0.9924 | 0.9936 | 0.9939 | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, C.; Liu, P.; Wang, X.; Zhang, W.; He, L. Effects of Phenyllactic Acid on Fermentation Parameters, Nitrogen Fractions and Bacterial Community of High-Moisture Stylo Silage. Fermentation 2023, 9, 572. https://doi.org/10.3390/fermentation9060572
Dong C, Liu P, Wang X, Zhang W, He L. Effects of Phenyllactic Acid on Fermentation Parameters, Nitrogen Fractions and Bacterial Community of High-Moisture Stylo Silage. Fermentation. 2023; 9(6):572. https://doi.org/10.3390/fermentation9060572
Chicago/Turabian StyleDong, Chenxi, Ping Liu, Xianliu Wang, Wei Zhang, and Liwen He. 2023. "Effects of Phenyllactic Acid on Fermentation Parameters, Nitrogen Fractions and Bacterial Community of High-Moisture Stylo Silage" Fermentation 9, no. 6: 572. https://doi.org/10.3390/fermentation9060572