
Sequential Bioprocess with Gluconobacter oxydans and Candida tropicalis for Gluconic Acid and Single-Cell Protein Production from Enzymatic Hydrolysate
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain Maintenance
2.2. Preparation of Enzymatic Hydrolysis of Pretreated Corncob Solids
2.3. Inoculum Preparation and Fermentations
2.4. Analytical Methods
3. Results and Discussion
3.1. Enzymatic Hydrolysis of Pretreated Corncob Solids
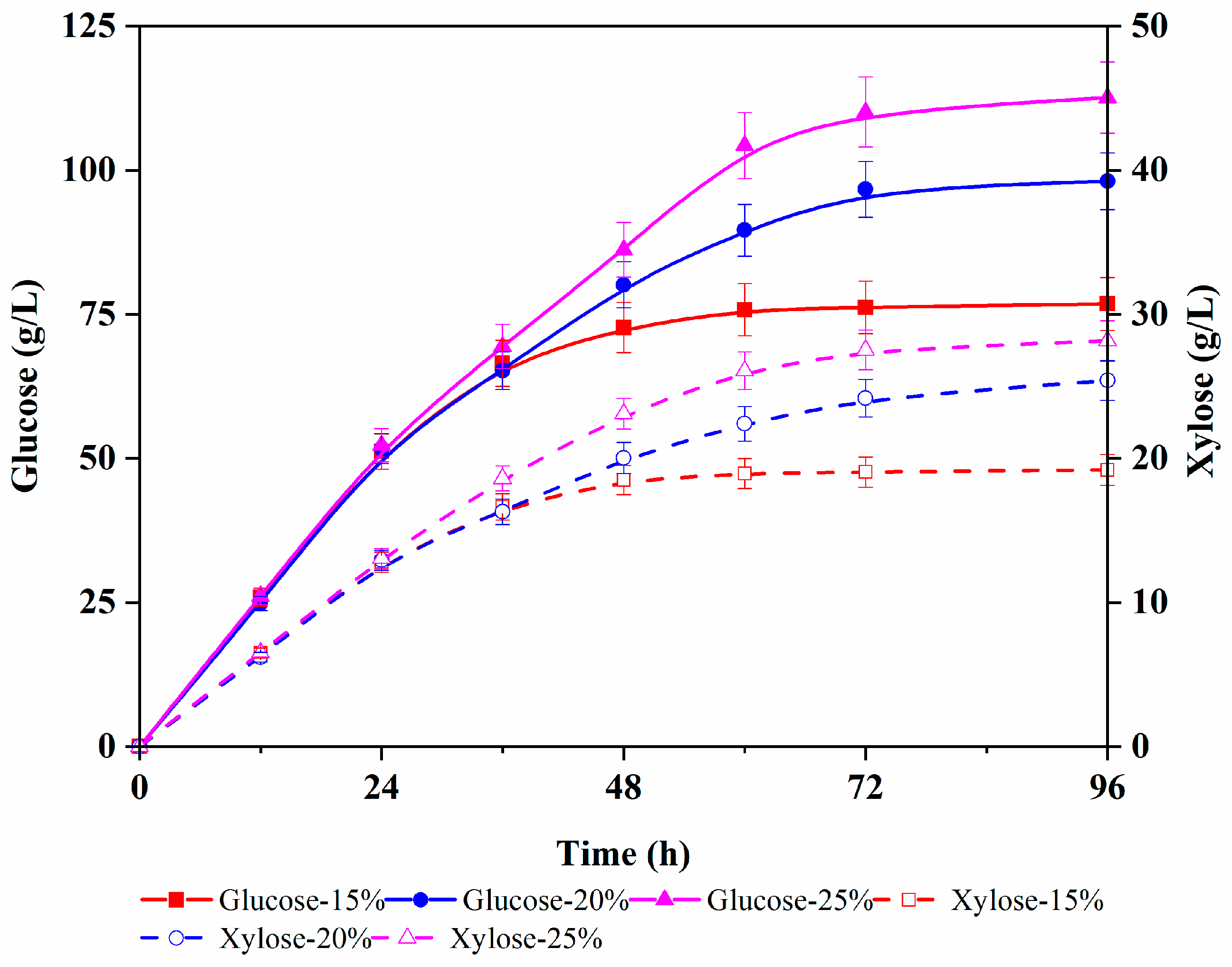
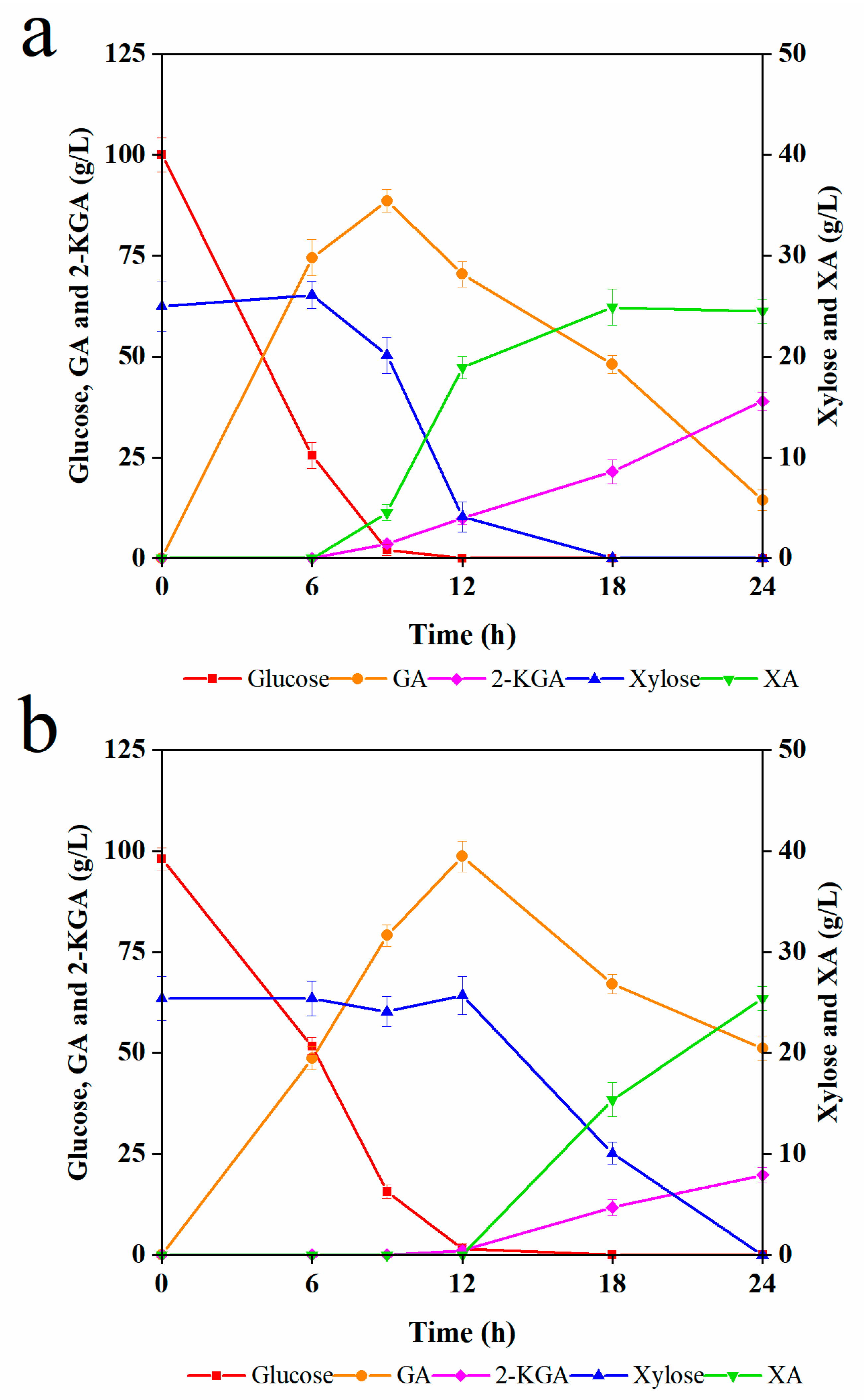
3.2. The Kinetics of G. oxydans for the Mixture Medium of Glucose and Xylose
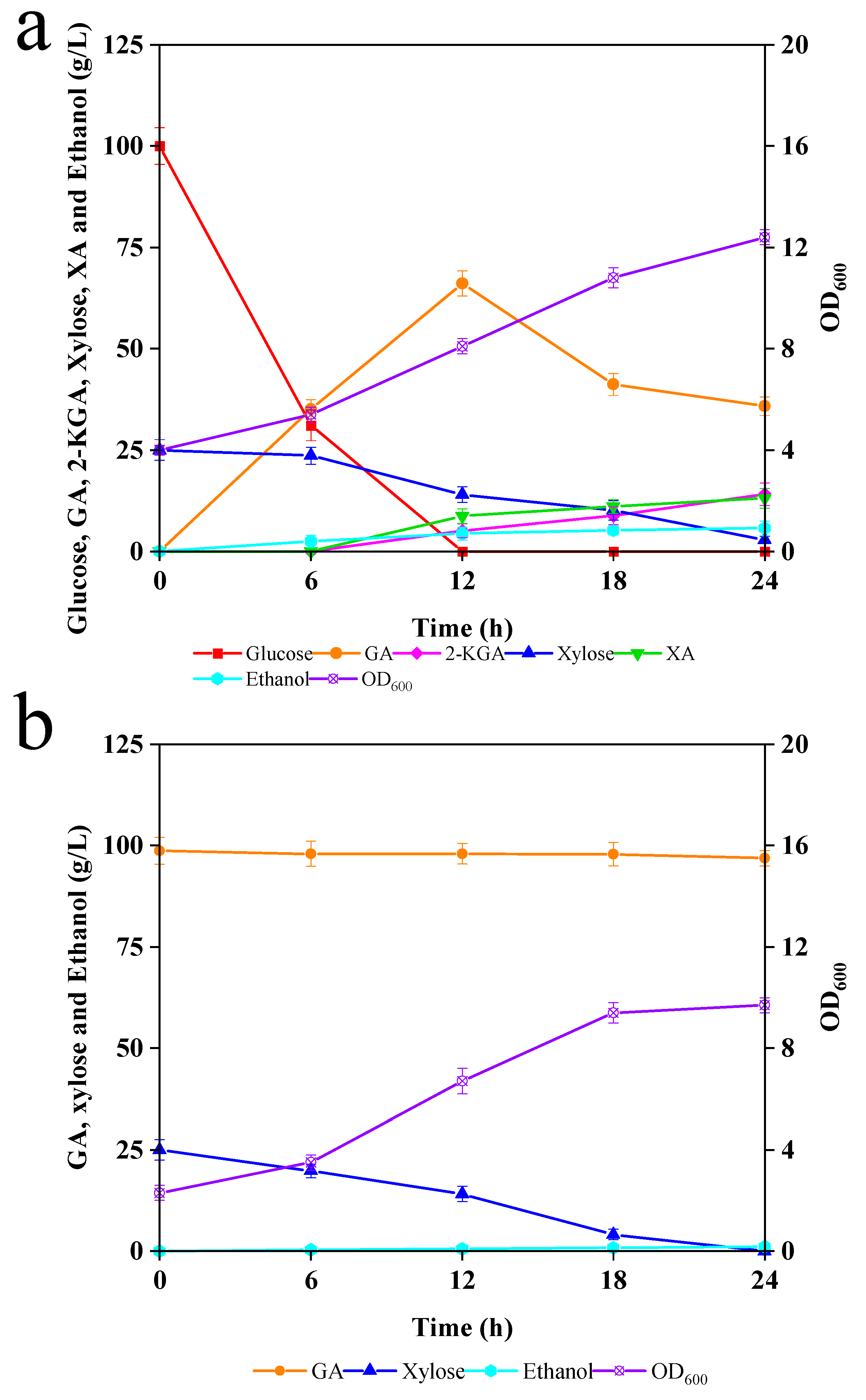
3.3. The Effects of G. oxydans and GA on C. tropicalis
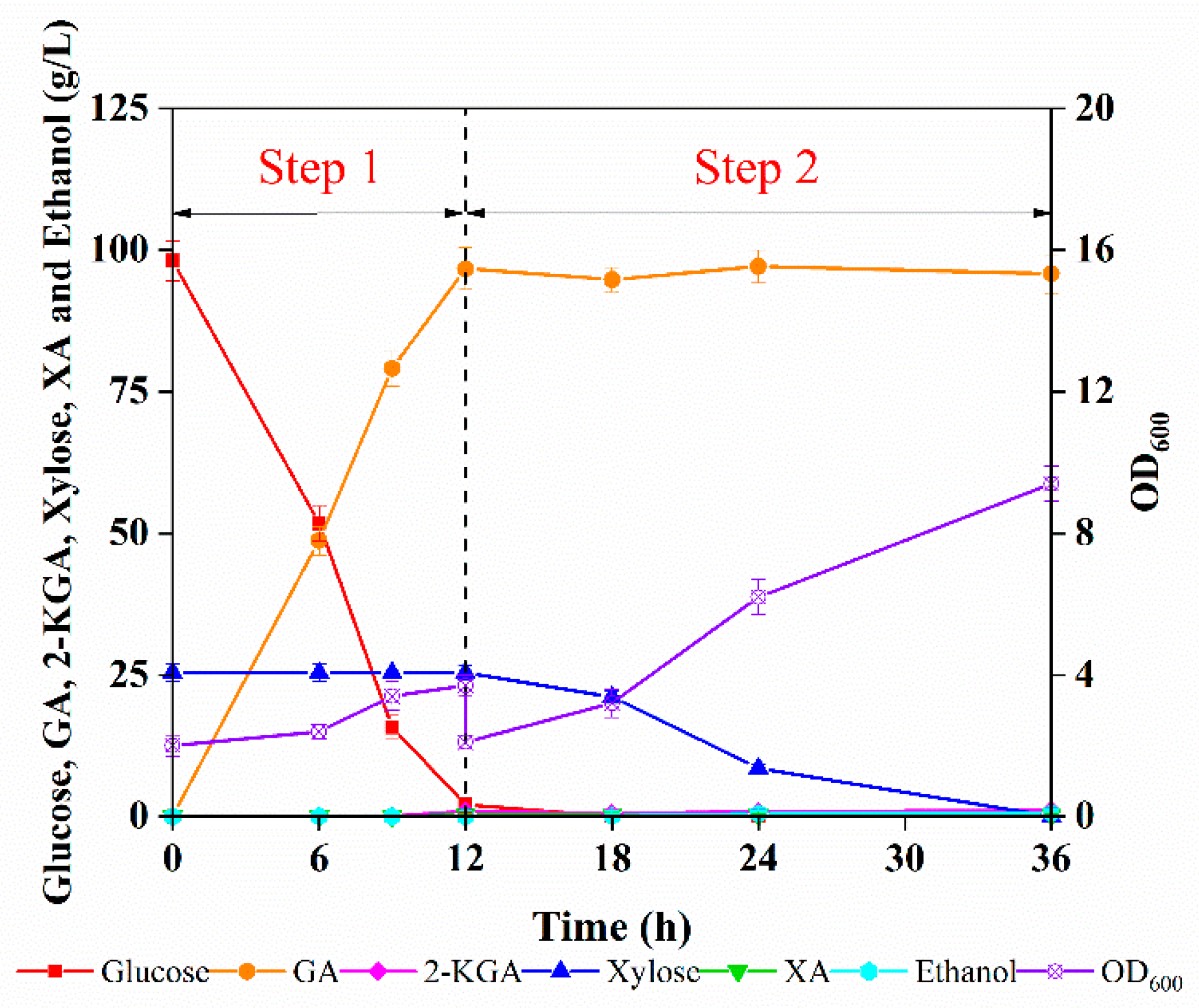
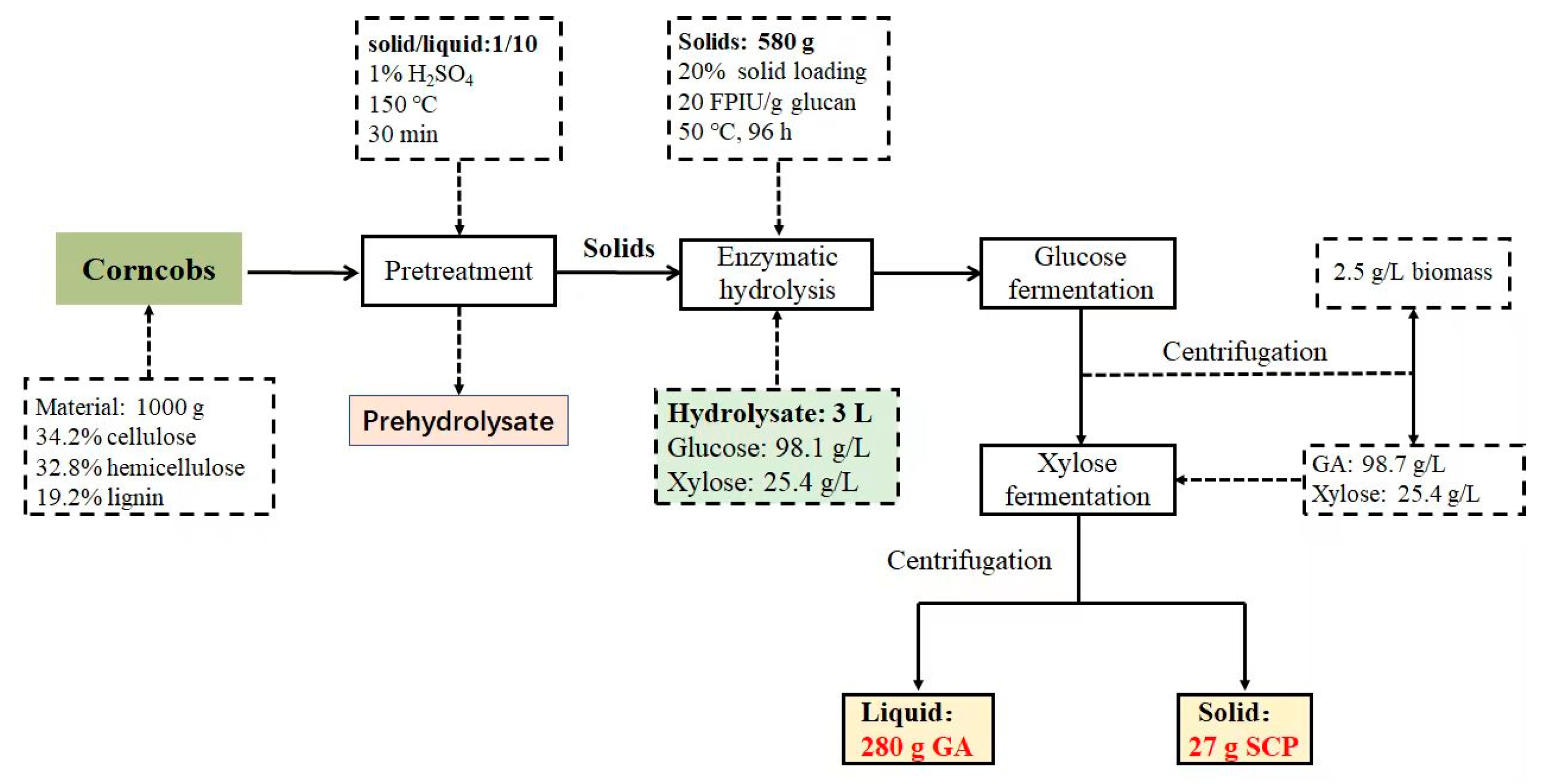
3.4. Sequential Bioprocess for Converting Enzymatic Hydrolysates into GA and Single-Cell Protein
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saravanan, A.; Senthil Kumar, P.; Jeevanantham, S.; Karishma, S.; Vo, D.N. Recent advances and sustainable development of biofuels production from lignocellulosic biomass. Bioresour. Technol. 2022, 344, 126203. [Google Scholar] [CrossRef] [PubMed]
- Deng, B.; Hou, Y.; Wang, F.; Bao, Y.; Zeng, F.; Qin, C.; Liang, C.; Huang, C.; Ma, J.; Yao, S. Highly selective separation of eucalyptus hemicellulose by salicylic acid treatment with both aromatic and hydroxy acids. Bioresour. Technol. 2022, 355, 127304. [Google Scholar] [CrossRef]
- Dar, M.A.; Pawar, K.D.; Rajput, B.P.; Rahi, P.; Pandit, R.S. Purification of a cellulase from cellulolytic gut bacterium, Bacillus tequilensis G9 and its evaluation for valorization of agro-wastes into added value byproducts. Biocatal. Agric. Biotechnol. 2019, 20, 101219. [Google Scholar] [CrossRef]
- Liu, X.; Duan, X.; Wei, W.; Wang, S.; Ni, B. Photocatalytic conversion of lignocellulosic biomass to valuable products. Green Chem. 2019, 21, 4266–4289. [Google Scholar] [CrossRef]
- Liu, Y.; Nie, Y.; Lu, X.; Zhang, X.; He, H.; Pan, F.; Zhou, L.; Liu, X.; Ji, X.; Zhang, S. Cascade utilization of lignocellulosic biomass to high-value products. Green Chem. 2019, 21, 3499–3535. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, D.; Wang, K.; Jia, Y.; Yang, C.; Shen, B.; Lai, C.; Yong, Q. In-situ lignin modification with polyethylene glycol-epoxides to boost enzymatic hydrolysis of combined-pretreated masson pine. Bioresour. Technol. 2022, 344, 126315. [Google Scholar] [CrossRef]
- Dar, M.A.; Dhole, N.P.; Xie, R.; Pawar, K.D.; Ullah, K.; Rahi, P.; Pandit, R.S.; Sun, J. Valorization potential of a novel bacterial strain, Bacillus altitudinis RSP75, towards lignocellulose bioconversion: An assessment of symbiotic bacteria from the stored grain pest, Tribolium castaneum. Microorganisms 2021, 9, 1952. [Google Scholar] [CrossRef]
- Dar, M.A.; Shaikh, A.A.; Pawar, K.D.; Pandit, R.S. Exploring the gut of Helicoverpa armigera for cellulose degrading bacteria and evaluation of a potential strain for lignocellulosic biomass deconstruction. Process Biochem. 2018, 73, 142–153. [Google Scholar] [CrossRef]
- Huang, C.; Zhao, X.; Zheng, Y.; Lin, W.; Lai, C.; Yong, Q.; Ragauskas, A.J.; Meng, X. Revealing the mechanism of surfactant-promoted enzymatic hydrolysis of dilute acid pretreated bamboo. Bioresour. Technol. 2022, 360, 127524. [Google Scholar] [CrossRef]
- Ursachi, V.; Gutt, G. Production of cellulosic ethanol from enzymatically hydrolysed wheat straws. Appl. Sci. 2020, 10, 7638. [Google Scholar] [CrossRef]
- Zhang, Y.; Yoshida, M.; Vadlani, P.V. Biosynthesis of d-lactic acid from lignocellulosic biomass. Biotechnol. Lett. 2018, 40, 1167–1179. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.; Bao, J. Simultaneous saccharification and aerobic fermentation of high titer cellulosic citric acid by filamentous fungus Aspergillus niger. Bioresour. Technol. 2018, 253, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Xu, Y. Integrative process for sugarcane bagasse biorefinery to co-produce xylooligosaccharides and gluconic acid. Bioresour. Technol. 2019, 282, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Cañete-Rodríguez, A.M.; Santos-Dueñas, I.M.; Jiménez-Hornero, J.E.; Ehrenreich, A.; Liebl, W.; García-García, I. Gluconic acid: Properties, production methods and applications—An excellent opportunity for agro-industrial by-products and waste bio-valorization. Process Biochem. 2016, 51, 1891–1903. [Google Scholar] [CrossRef]
- Luo, Z.; Wang, Y.; Liu, X.; Tian, C.; Wang, Y.; Mu, Y.; Zhang, M.; Hao, Y. Comparative investigation of effect of borax and sodium gluconate retarders on properties of magnesium phosphate cement. Arab. J. Sci. Eng. 2022, 47, 13187–13198. [Google Scholar] [CrossRef]
- Ma, S.; Li, W.; Zhang, S.; Ge, D.; Yu, J.; Shen, X. Influence of sodium gluconate on the performance and hydration of portland cement. Constr. Build. Mater. 2015, 91, 138–144. [Google Scholar] [CrossRef]
- Ye, Q.; Han, Y.; Liu, T.; Bai, Y.; Chen, Y.; Li, J.; Shi, S.Q. Magnesium oxychloride cement reinforced via d-gluconic acid sodium salt for slow-curing, with enhanced compressive strength and water resistance. Constr. Build. Mater. 2021, 280, 122487. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, J.; Bao, J. High titer gluconic acid fermentation by Aspergillus niger from dry dilute acid pretreated corn stover without detoxification. Bioresour. Technol. 2016, 203, 211–219. [Google Scholar] [CrossRef]
- Gupta, A.; Singh, V.K.; Qazi, G.N.; Kumar, A. Gluconobacter oxydans: Its biotechnological applications. J. Mol. Microbiol. Biotechnol. 2001, 3, 445–456. [Google Scholar]
- Zhang, H.; Liu, G.; Zhang, J.; Bao, J. Fermentative production of high titer gluconic and xylonic acids from corn stover feedstock by Gluconobacter oxydans and techno-economic analysis. Bioresour. Technol. 2016, 219, 123–131. [Google Scholar] [CrossRef]
- Dzanaeva, L.; Kruk, B.; Ruchala, J.; Nielsen, J.; Sibirny, A.; Dmytruk, K. The role of peroxisomes in xylose alcoholic fermentation in the engineered Saccharomyces cerevisiae. Cell Biol. Int. 2020, 44, 1606–1615. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, F.; Zacchi, G.; Galbe, M.; Wallberg, O. Sequential targeting of xylose and glucose conversion in fed-batch simultaneous saccharification and co-fermentation of steam-pretreated wheat straw for improved xylose conversion to ethanol. BioEnergy Res. 2017, 10, 800–810. [Google Scholar] [CrossRef]
- Finneran, K.T.; Popovic, J. Solvent production from xylose. Appl. Microbiol. Biotechnol. 2018, 102, 8707–8715. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Xie, C.; Xia, Z.; Wu, Y.; Li, B.; Tang, Y. The effect of xylose reductase genes on xylitol production by industrial Saccharomyces cerevisiae in fermentation of glucose and xylose. Process Biochem. 2020, 95, 122–130. [Google Scholar] [CrossRef]
- Zhou, X.; Lü, S.; Xu, Y.; Mo, Y.; Yu, S. Improving the performance of cell biocatalysis and the productivity of xylonic acid using a compressed oxygen supply. Biochem. Eng. J. 2015, 93, 196–199. [Google Scholar] [CrossRef]
- Kochova, K.; Schollbach, K.; Gauvin, F.; Brouwers, H.J.H. Effect of saccharides on the hydration of ordinary portland cement. Constr. Build. Mater. 2017, 150, 268–275. [Google Scholar] [CrossRef]
- Yang, Z.; Jiang, L.; Zhang, M.; Deng, Y.; Suo, W.; Zhang, H.; Wang, C.; Li, H. Bioconversion of apple pomace into microbial protein feed based on extrusion pretreatment. Appl. Biochem. Biotechnol. 2022, 194, 1496–1509. [Google Scholar] [CrossRef]
- Sánchez, S.; Bravo, V.; García, J.F.; Cruz, N.; Cuevas, M. Fermentation of d-glucose and d-xylose mixtures by Candida tropicalis NBRC 0618 for xylitol production. World J. Microbiol. Biotechnol. 2008, 24, 709–716. [Google Scholar] [CrossRef]
- Wang, S.; Cheng, G.; Joshua, C.; He, Z.; Sun, X.; Li, R.; Liu, L.; Yuan, Q. Furfural tolerance and detoxification mechanism in Candida tropicalis. Biotechnol. Biofuels 2016, 9, 250. [Google Scholar] [CrossRef]
- Xu, L.; Liu, L.; Li, S.; Zheng, W.; Cui, Y.; Liu, R.; Sun, W. Xylitol production by Candida tropicalis 31949 from sugarcane bagasse hydrolysate. Sugar Tech 2019, 21, 341–347. [Google Scholar] [CrossRef]
- Zhou, X.; Xu, Y.; Yu, S. Simultaneous bioconversion of xylose and glycerol to xylonic acid and 1,3-dihydroxyacetone from the mixture of pre-hydrolysates and ethanol-fermented waste liquid by Gluconobacter oxydans. Appl. Biochem. Biotechnol. 2016, 178, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Alvira, P.; Tomás-Pejó, E.; Ballesteros, M.; Negro, M.J. Pretreatment technologies for an efficient bioethanol production process based on enzymatic hydrolysis: A review. Bioresour. Technol. 2010, 101, 4851–4861. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.; Kadhum, H.J.; Murthy, G.S.; Dien, B.S.; Singh, V. High solids loading biorefinery for the production of cellulosic sugars from bioenergy sorghum. Bioresour. Technol. 2020, 318, 124051. [Google Scholar] [CrossRef]
- Chaiyaso, T.; Manowattana, A.; Techapun, C.; Watanabe, M. Efficient bioconversion of enzymatic corncob hydrolysate into biomass and lipids by oleaginous yeast Rhodosporidium paludigenum KM281510. Prep. Biochem. Biotechnol. 2019, 49, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Guo, X.; Feng, X.; Li, C. An environment friendly and efficient process for xylitol bioconversion from enzymatic corncob hydrolysate by adapted Candida tropicalis. Chem. Eng. J. 2015, 263, 249–256. [Google Scholar] [CrossRef]
- Cai, X.; Hu, C.; Wang, J.; Zeng, X.; Luo, J.; Li, M.; Liu, Z.; Zheng, Y. Efficient high-solids enzymatic hydrolysis of corncobs by an acidic pretreatment and a fed-batch feeding mode. Bioresour. Technol. 2021, 326, 124768. [Google Scholar] [CrossRef]
- Bommarius, A.S.; Katona, A.; Cheben, S.E.; Patel, A.S.; Ragauskas, A.J.; Knudson, K.; Pu, Y. Cellulase kinetics as a function of cellulose pretreatment. Metab. Eng. 2008, 10, 370–381. [Google Scholar] [CrossRef]
- Lian, Z.; Zhang, Q.; Xu, Y.; Zhou, X.; Jiang, K. Biorefinery cascade processing for converting corncob to xylooligosaccharides and glucose by maleic acid pretreatment. Appl. Biochem. Biotechnol. 2022, 194, 4946–4958. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, X.; Cao, R.; Tao, Y.; Xu, Y.; Yu, S. Characteristics and kinetics of the aldonic acids production using whole-cell catalysis of Gluconobacter oxydans. BioResources 2015, 10, 4277–4286. [Google Scholar] [CrossRef]
- Zhou, X.; Zhou, X.; Huang, L.; Cao, R.; Xu, Y. Efficient coproduction of gluconic acid and xylonic acid from lignocellulosic hydrolysate by Zn(ii)-selective inhibition on whole-cell catalysis by Gluconobacter oxydans. Bioresour. Technol. 2017, 243, 855–859. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, L.; Huang, R.; Ying, J.; Fu, Y.; Zhou, X.; Jiang, K. Sequential Bioprocess with Gluconobacter oxydans and Candida tropicalis for Gluconic Acid and Single-Cell Protein Production from Enzymatic Hydrolysate. Fermentation 2023, 9, 562. https://doi.org/10.3390/fermentation9060562
Cheng L, Huang R, Ying J, Fu Y, Zhou X, Jiang K. Sequential Bioprocess with Gluconobacter oxydans and Candida tropicalis for Gluconic Acid and Single-Cell Protein Production from Enzymatic Hydrolysate. Fermentation. 2023; 9(6):562. https://doi.org/10.3390/fermentation9060562
Chicago/Turabian StyleCheng, Lu, Rong Huang, Jiaxun Ying, Yixiu Fu, Xin Zhou, and Kankan Jiang. 2023. "Sequential Bioprocess with Gluconobacter oxydans and Candida tropicalis for Gluconic Acid and Single-Cell Protein Production from Enzymatic Hydrolysate" Fermentation 9, no. 6: 562. https://doi.org/10.3390/fermentation9060562