
Zinc Tolerance of Special Yeasts and Lactic Acid Bacteria for Use in the Food Industry

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Zinc-Enriched Microorganisms
2.3. OD600 Assay
2.4. Determination of Cell Viability via Colony-Forming Units
2.5. Determination of Cellular Zinc Concentration
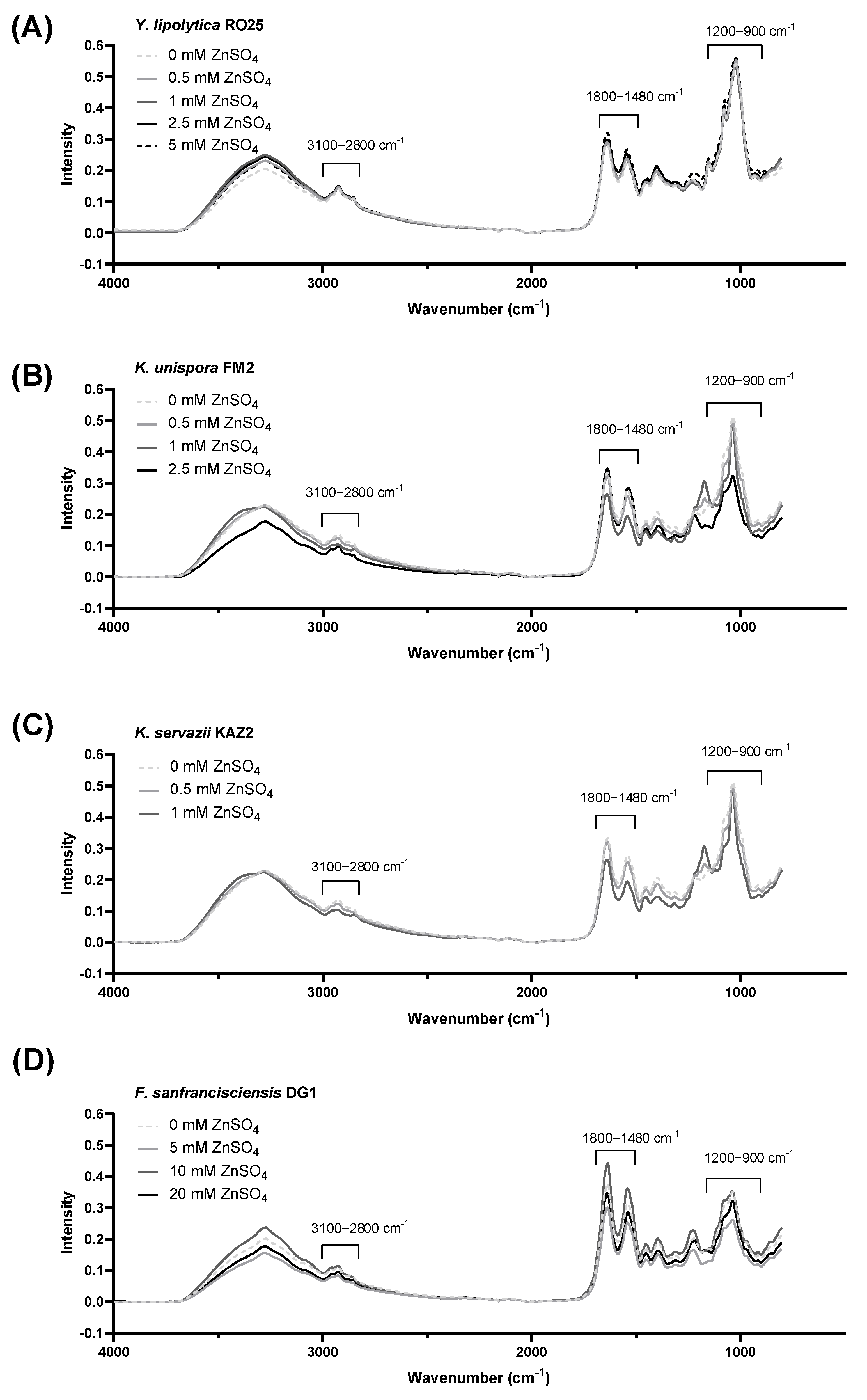
2.6. FTIR Analysis
2.7. Statistical Analyses
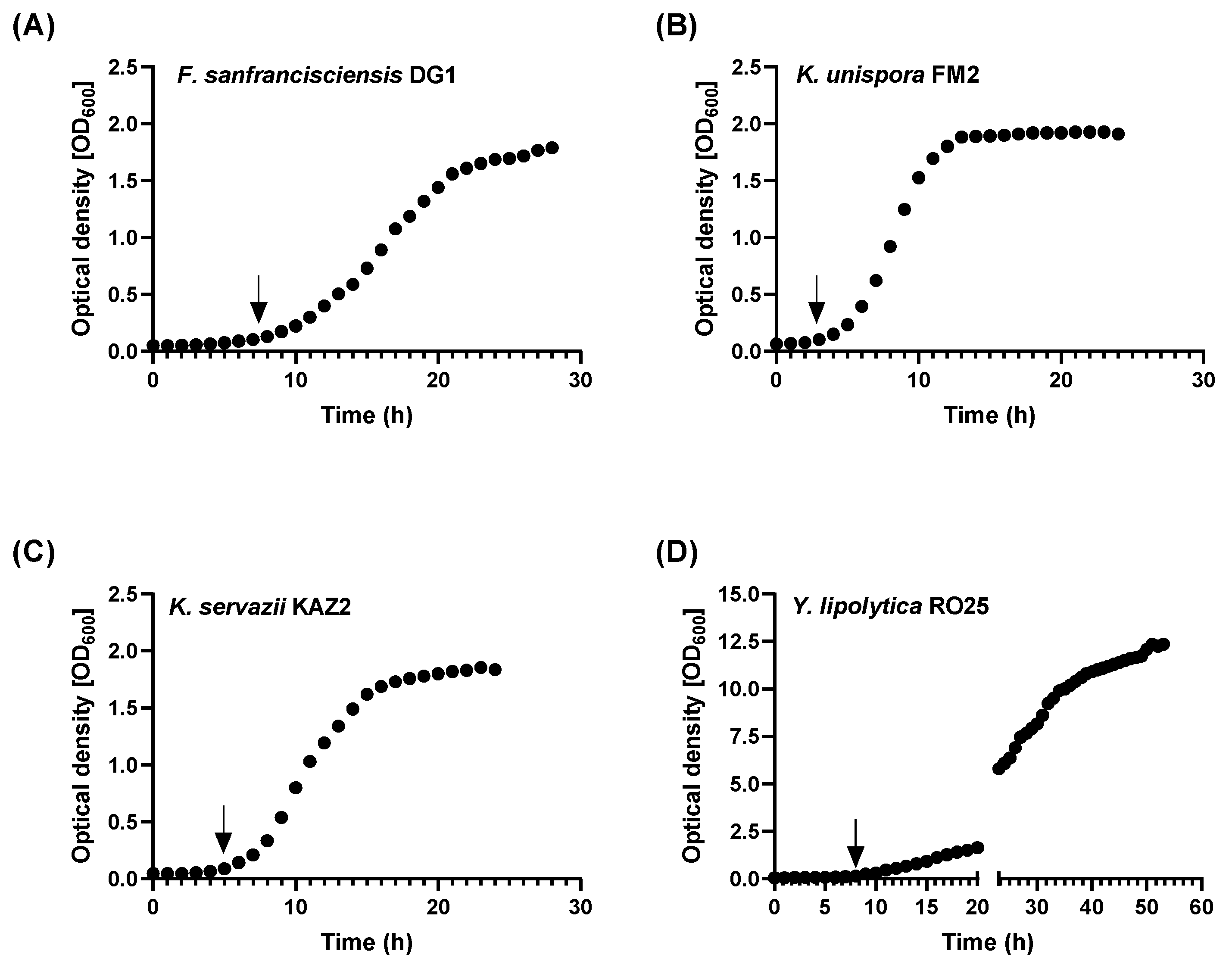
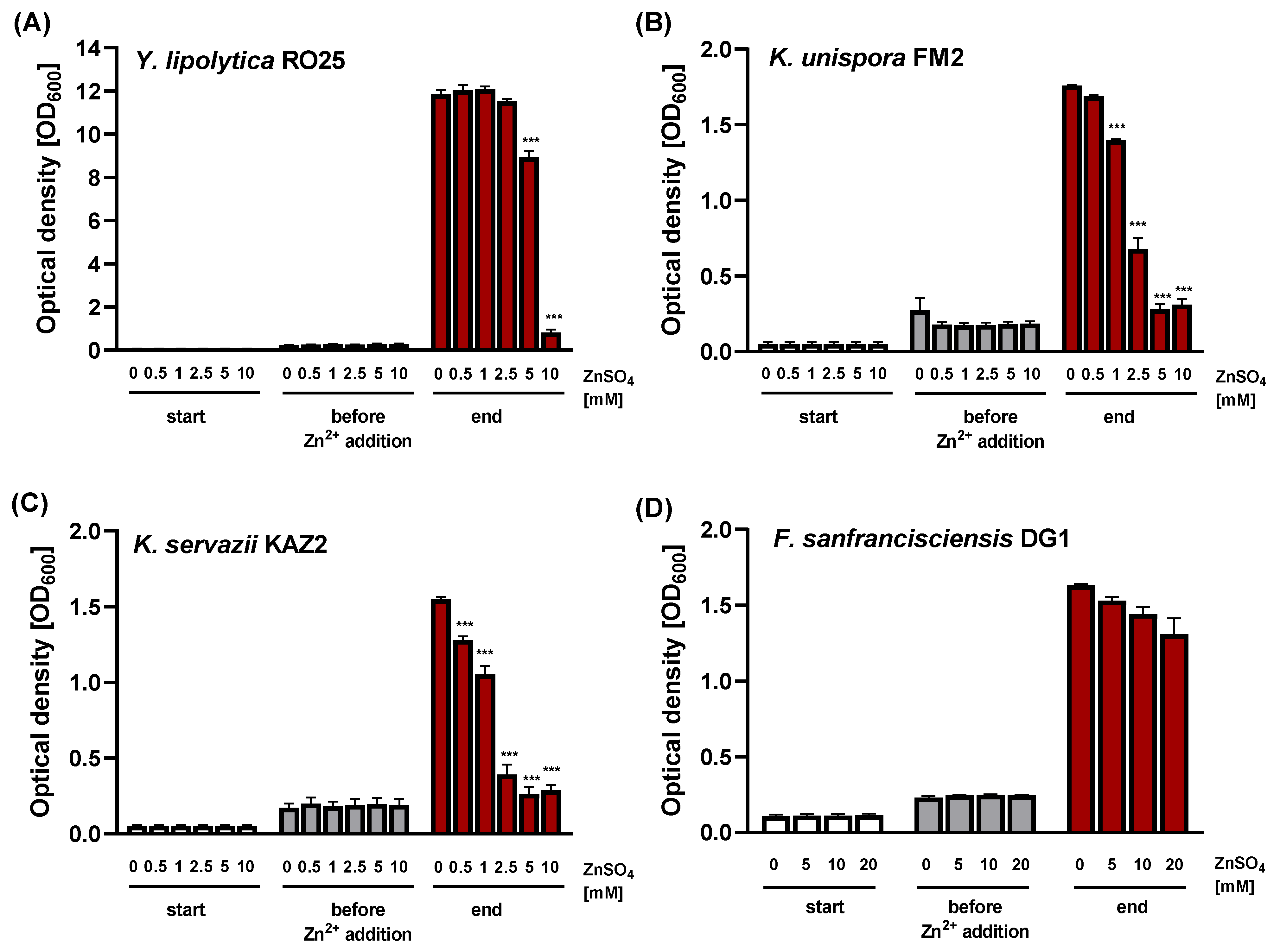
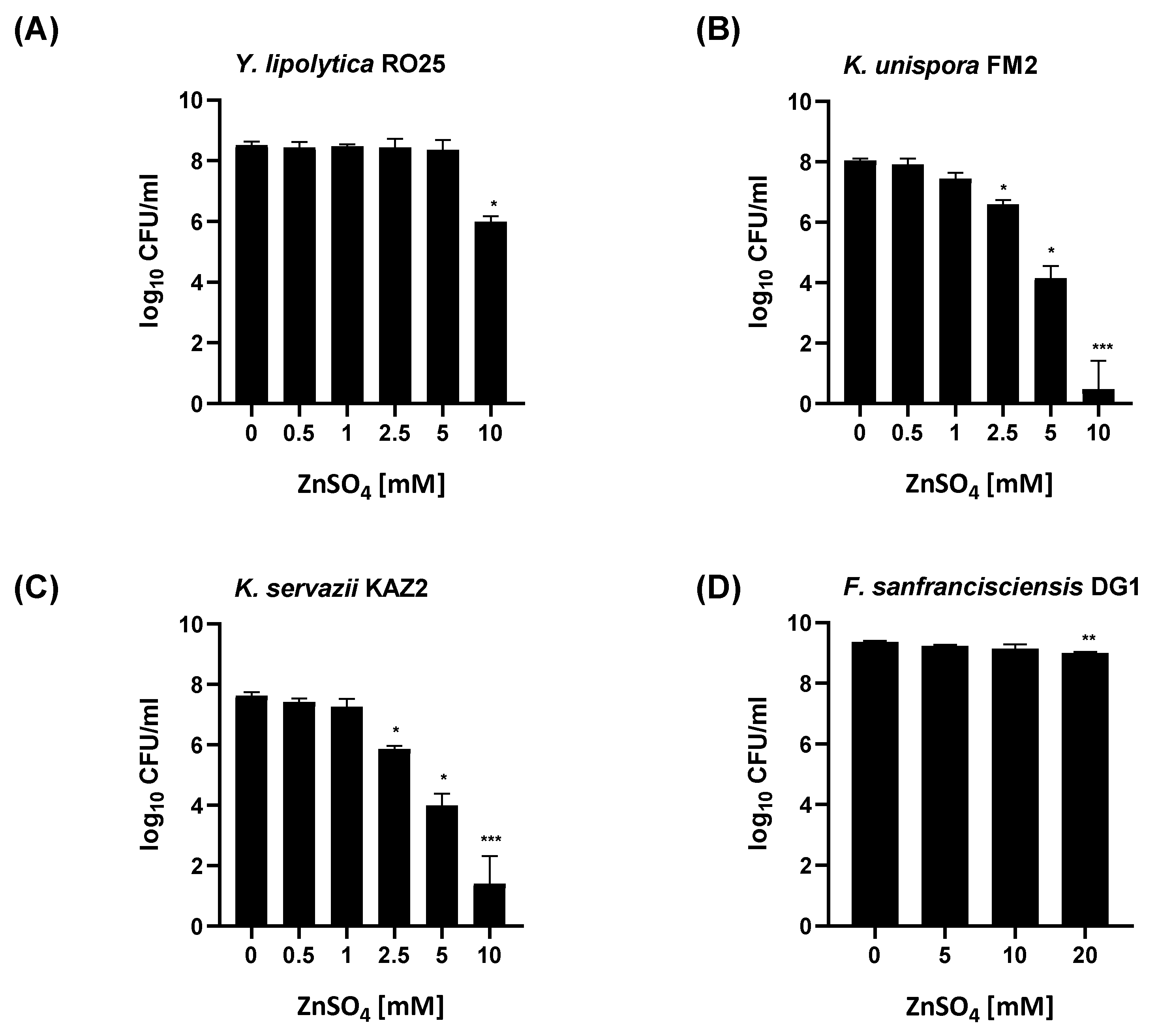
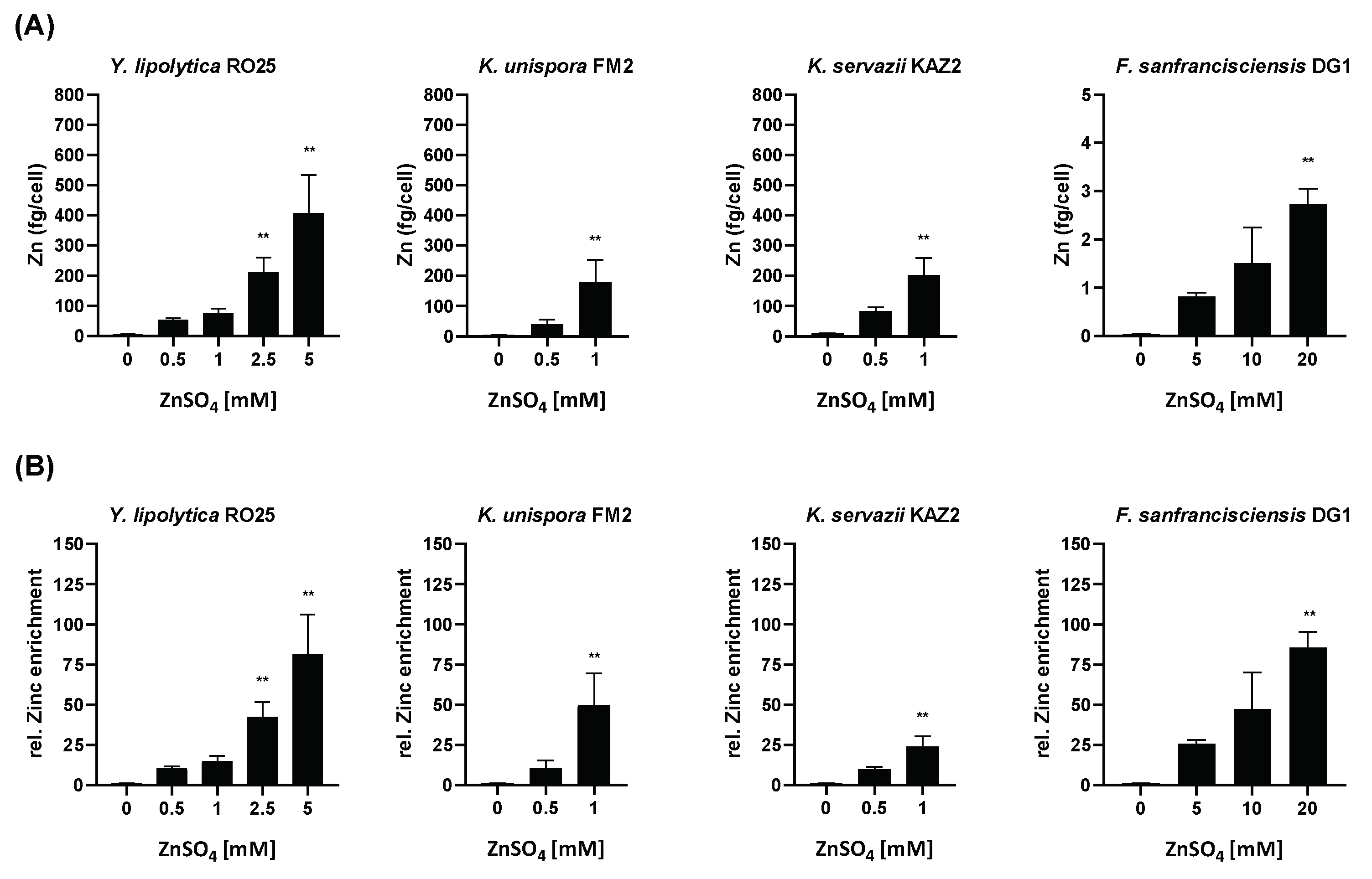
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ohanenye, I.C.; Emenike, C.U.; Mensi, A.; Medina-Godoy, S.; Jin, J.; Ahmed, T.; Sun, X.; Udenigwe, C.C. Food fortification technologies: Influence on iron, zinc and vitamin A bioavailability and potential implications on micronutrient deficiency in sub-Saharan Africa. Sci. Afr. 2021, 11, e00667. [Google Scholar] [CrossRef]
- Wessells, K.R.; Brown, K.H. Estimating the Global Prevalence of Zinc Deficiency: Results Based on Zinc Availability in National Food Supplies and the Prevalence of Stunting. PLoS ONE 2012, 7, e50568. [Google Scholar] [CrossRef]
- Beal, T.; Massiot, E.; Arsenault, J.E.; Smith, M.R.; Hijmans, R.J. Global trends in dietary micronutrient supplies and estimated prevalence of inadequate intakes. PLoS ONE 2017, 12, e0175554. [Google Scholar] [CrossRef]
- Wessels, I.; Rolles, B.; Slusarenko, A.J.; Rink, L. Zinc deficiency as a possible risk factor for increased susceptibility and severe progression of Corona Virus Disease 19. Br. J. Nutr. 2022, 127, 214–232. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.S. Discovery of Human Zinc Deficiency: Its Impact on Human Health and Disease. Adv. Nutr. Int. Rev. J. 2013, 4, 176–190. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.S. Lessons Learned from Experimental Human Model of Zinc Deficiency. J. Immunol. Res. 2020, 2020, 9207279. [Google Scholar] [CrossRef] [PubMed]
- Maares, M.; Haase, H. A Guide to Human Zinc Absorption: General Overview and Recent Advances of In Vitro Intestinal Models. Nutrients 2020, 12, 762. [Google Scholar] [CrossRef] [PubMed]
- Roohani, N.; Hurrell, R.; Kelishadi, R.; Schulin, R. Zinc and its importance for human health: An integrative review. J. Res. Med. Sci. 2013, 18, 144–157. [Google Scholar] [PubMed]
- Gupta, S.; Brazier, A.K.M.; Lowe, N.M. Zinc deficiency in low- and middle-income countries: Prevalence and approaches for mitigation. J. Hum. Nutr. Diet. 2020, 33, 624–643. [Google Scholar] [CrossRef]
- Shah, D.; Sachdev, H.S.; Gera, T.; De-Regil, L.M.; Peña-Rosas, J.P. Fortification of staple foods with zinc for improving zinc status and other health outcomes in the general population. Cochrane Database Syst. Rev. 2016, 2016, CD010697. [Google Scholar] [CrossRef]
- Gomes, M.J.C.; Martino, H.S.D.; Tako, E. Zinc-biofortified staple food crops to improve zinc status in humans: A systematic review. Crit. Rev. Food Sci. Nutr. 2021, 60, 1–13. [Google Scholar] [CrossRef]
- Tsang, B.L.; Holsted, E.; McDonald, C.M.; Brown, K.H.; Black, R.; Mbuya, M.N.N.; Grant, F.; A Rowe, L.; Manger, M.S. Effects of Foods Fortified with Zinc, Alone or Cofortified with Multiple Micronutrients, on Health and Functional Outcomes: A Systematic Review and Meta-Analysis. Adv. Nutr. Int. Rev. J. 2021, 12, 1821–1837. [Google Scholar] [CrossRef]
- Hall, A.G.; King, J.C. Zinc Fortification: Current Trends and Strategies. Nutrients 2022, 14, 3895. [Google Scholar] [CrossRef] [PubMed]
- de Romaña, D.L.; Lönnerdal, B.; Brown, K.H. Absorption of zinc from wheat products fortified with iron and either zinc sulfate or zinc oxide. Am. J. Clin. Nutr. 2003, 78, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Das, J.K.; Kumar, R.; Salam, R.A.; Bhutta, Z.A. Systematic Review of Zinc Fortification Trials. Ann. Nutr. Metab. 2013, 62, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Anjum, F.M.; Randhawa, M.A.; Farooq, U.; Akhtar, S.; Sultan, M.T. Effect of multiple fortification on the bioavailability of minerals in wheat meal bread. J. Food Sci. Technol. 2012, 49, 737–744. [Google Scholar] [CrossRef]
- Garnweidner-Holme, L.; Hallquist, M.; Watters, S.I.; Gjøvik, M.; Frederiksen, M.P.; Jonassen, S.; Ravnanger, I.; Henriksen, C.; Myhrstad, M.; Telle-Hansen, V. Attitudes towards and experiences with sourdough and baker’s yeast bread amongst par ticipants in a randomised controlled trial: A qualitative study. Food Nutr. Res. 2022, 66, 66. [Google Scholar] [CrossRef] [PubMed]
- Graça, C.; Lima, A.; Raymundo, A.; Sousa, I. Sourdough Fermentation as a Tool to Improve the Nutritional and Health-Promoting Properties of Its Derived-Products. Fermentation 2021, 7, 246. [Google Scholar] [CrossRef]
- Fernández-Peláez, J.; Paesani, C.; Gómez, M. Sourdough Technology as a Tool for the Development of Healthier Grain-Based Products: An Update. Agronomy 2020, 10, 1962. [Google Scholar] [CrossRef]
- Gobbetti, M.; De Angelis, M.; Di Cagno, R.; Calasso, M.; Archetti, G.; Rizzello, C.G. Novel insights on the functional/nutritional features of the sourdough fermentation. Int. J. Food Microbiol. 2019, 302, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Leenhardt, F.; Levrat-Verny, M.-A.; Chanliaud, E.; Rémésy, C. Moderate Decrease of pH by Sourdough Fermentation Is Sufficient To Reduce Phytate Content of Whole Wheat Flour through Endogenous Phytase Activity. J. Agric. Food Chem. 2005, 53, 98–102. [Google Scholar] [CrossRef]
- Yildirim, R.M.; Arici, M. Effect of the fermentation temperature on the degradation of phytic acid in whole-wheat sourdough bread. LWT 2019, 112, 108224. [Google Scholar] [CrossRef]
- Canesin, M.R.; Cazarin, C.B.B. Nutritional quality and nutrient bioaccessibility in sourdough bread. Curr. Opin. Food Sci. 2021, 40, 81–86. [Google Scholar] [CrossRef]
- Gobbetti, M.; Rizzello, C.G.; Di Cagno, R.; De Angelis, M. How the sourdough may affect the functional features of leavened baked goods. Food Microbiol. 2014, 37, 30–40. [Google Scholar] [CrossRef]
- Lopez, H.W.; Duclos, V.; Coudray, C.; Krespine, V.; Feillet-Coudray, C.; Messager, A.; Demigné, C.; Rémésy, C. Making bread with sourdough improves mineral bioavailability from reconstituted whole wheat flour in rats. Nutrition 2003, 19, 524–530. [Google Scholar] [CrossRef]
- Najafi, M.A.; Rezaei, K.; Safari, M.; Razavi, S.H. Use of sourdough to reduce phytic acid and improve zinc bioavailability of a traditional flat bread (sangak) from Iran. Food Sci. Biotechnol. 2012, 21, 51–57. [Google Scholar] [CrossRef]
- Nissen, L.; Samaei, S.P.; Babini, E.; Gianotti, A. Gluten free sourdough bread enriched with cricket flour for protein fortification: Antioxidant improvement and Volatilome characterization. Food Chem. 2020, 333, 127410. [Google Scholar] [CrossRef] [PubMed]
- Vasilica, B.; Chiș, M.S.; Alexa, E.; Pop, C.; Păucean, A.; Man, S.; Igual, M.; Haydee, K.M.; Dalma, K.E.; Stănilă, S.; et al. The Impact of Insect Flour on Sourdough Fermentation-Fatty Acids, Amino-Acids, Minerals and Volatile Profile. Insects 2022, 13, 576. [Google Scholar] [CrossRef] [PubMed]
- Hlongwane, Z.T.; Slotow, R.; Munyai, T.C. Indigenous Knowledge about Consumption of Edible Insects in South Africa. Insects 2021, 12, 22. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.R.; Stull, V.J.; A Patz, J.; Myers, S.S. Nutritional and environmental benefits of increasing insect consumption in Africa and Asia. Environ. Res. Lett. 2021, 16, 065001. [Google Scholar] [CrossRef]
- Svanberg, I.; Berggren, Å. Insects as past and future food in entomophobic Europe. Food Cult. Soc. 2021, 24, 624–638. [Google Scholar] [CrossRef]
- Kröger, T.; Dupont, J.; Büsing, L.; Fiebelkorn, F. Acceptance of Insect-Based Food Products in Western Societies: A Systematic Review. Front. Nutr. 2022, 8, 759885. [Google Scholar] [CrossRef]
- Steensels, J.; Snoek, T.; Meersman, E.; Nicolino, M.P.; Voordeckers, K.; Verstrepen, K.J. Improving industrial yeast strains: Exploiting natural and artificial diversity. FEMS Microbiol. Rev. 2014, 38, 947–995. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Semumu, T.; Gamero, A. Non-Conventional Yeasts as Alternatives in Modern Baking for Improved Performance and Aroma Enhancement. Fermentation 2021, 7, 102. [Google Scholar] [CrossRef]
- Landis, E.A.; Oliverio, A.M.; McKenney, E.A.; Nichols, L.M.; Kfoury, N.; Biango-Daniels, M.; Shell, L.; Madden, A.; Shapiro, L.; Sakunala, S.; et al. The diversity and function of sourdough starter microbiomes. elife 2021, 10, e61644. [Google Scholar] [CrossRef] [PubMed]
- Wittwer, A.; Howell, K. Rising stars in the bakery: Novel yeasts for modern bread. Microbiol. Aust. 2022, 43, 75–78. [Google Scholar] [CrossRef]
- Rossi, S.; Parrotta, L.; Del Duca, S.; Rosa, M.D.; Patrignani, F.; Schluter, O.; Lanciotti, R. Effect of Yarrowia lipolytica RO25 cricket-based hydrolysates on sourdough quality parameters. LWT 2021, 148, 111760. [Google Scholar] [CrossRef]
- Rossi, S.; Parrotta, L.; Gottardi, D.; Glicerina, V.; Del Duca, S.; Rosa, M.D.; Patrignani, F.; Schlüter, O.; Lanciotti, R. Unravelling the potential of cricket-based hydrolysed sourdough on the quality of an innovative bakery product. J. Insects Food Feed. 2022, 8, 921–935. [Google Scholar] [CrossRef]
- Haase, H.; Jordan, L.; Keitel, L.; Keil, C.; Mahltig, B. Comparison of methods for determining the effectiveness of antibacterial functionalized textiles. PLoS ONE 2017, 12, e0188304. [Google Scholar] [CrossRef]
- Gottardi, D.; Ciccone, M.; Siroli, L.; Lanciotti, R.; Patrignani, F. Use of Yarrowia lipolytica to Obtain Fish Waste Functional Hydrolysates Rich in Flavoring Compounds. Fermentation 2022, 8, 708. [Google Scholar] [CrossRef]
- Alker, W.; Schwerdtle, T.; Schomburg, L.; Haase, H. A Zinpyr-1-based Fluorimetric Microassay for Free Zinc in Human Serum. Int. J. Mol. Sci. 2019, 20, 4006. [Google Scholar] [CrossRef]
- Maares, M.; Keil, C.; Pallasdies, L.; Schmacht, M.; Senz, M.; Nissen, J.; Kieserling, H.; Drusch, S.; Haase, H. Zinc availability from zinc-enriched yeast studied with an in vitro digestion/Caco-2 cell culture model. J. Trace Elements Med. Biol. 2022, 71, 126934. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Weisenhorn, E.; MacDiarmid, C.W.; Andreini, C.; Bucci, M.; Taggart, J.; Banci, L.; Russell, J.; Coon, J.J.; Eide, D.J. The cellular economy of the Saccharomyces cerevisiae zinc proteome. Metallomics 2018, 10, 1755–1776. [Google Scholar] [CrossRef]
- Santivarangkna, C.; Wenning, M.; Foerst, P.; Kulozik, U. Damage of cell envelope of Lactobacillus helveticus during vacuum drying. J. Appl. Microbiol. 2007, 102, 748–756. [Google Scholar] [CrossRef]
- Mihoubi, W.; Sahli, E.; Gargouri, A.; Amiel, C. FTIR spectroscopy of whole cells for the monitoring of yeast apoptosis mediated by p53 over-expression and its suppression by Nigella sativa extracts. PLoS ONE 2017, 12, e0180680. [Google Scholar] [CrossRef]
- Erenstein, O.; Jaleta, M.; Mottaleb, K.A.; Sonder, K.; Donovan, J.; Braun, H.-J. Global Trends in Wheat Production, Consumption and Trade. In Wheat Improvement; Reynolds, M.P., Braun, H.-J., Eds.; Springer: Cham, Switzerland, 2022; pp. 47–66. [Google Scholar] [CrossRef]
- Chen, X.-P.; Zhang, Y.-Q.; Tong, Y.-P.; Xue, Y.-F.; Liu, D.-Y.; Zhang, W.; Deng, Y.; Meng, Q.-F.; Yue, S.-C.; Yan, P.; et al. Harvesting more grain zinc of wheat for human health. Sci. Rep. 2017, 7, 1–8. [Google Scholar] [CrossRef]
- Wang, M.; Kong, F.; Liu, R.; Fan, Q.; Zhang, X. Zinc in Wheat Grain, Processing, and Food. Front. Nutr. 2020, 7, 124. [Google Scholar] [CrossRef]
- WHO Guideline: Fortification of Maize Flour and Corn Meal with Vitamins and Minerals; World Health Organization: Geneva, Switzerland, 2016.
- Guideline: Fortification of Wheat Flour with Vitamins and Minerals as a Public Health Strategy; World Health Organization: Geneva, Switzerland, 2022.
- GFDX. Global Fortification Data Exchange. Nutrient Levels in Fortification Standards (Mid-Range or Average). Available online: https://fortificationdata.org/map-nutrient-levels-in-fortification-standards/ (accessed on 21 January 2023).
- Borges, M.M.; da Costa, D.V.; Trombete, F.M.; Câmara, A.K.F.I. Edible insects as a sustainable alternative to food products: An insight into quality aspects of reformulated bakery and meat products. Curr. Opin. Food Sci. 2022, 46, 100864. [Google Scholar] [CrossRef]
- Kröncke, N.; Grebenteuch, S.; Keil, C.; Demtröder, S.; Kroh, L.; Thünemann, A.F.; Benning, R.; Haase, H. Effect of Different Drying Methods on Nutrient Quality of the Yellow Mealworm (Tenebrio molitor L.). Insects 2019, 10, 84. [Google Scholar] [CrossRef] [PubMed]
- Bankar, A.; Zinjarde, S.; Telmore, A.; Walke, A.; Ravikumar, A. Morphological response of Yarrowia lipolytica under stress of heavy metals. Can. J. Microbiol. 2018, 64, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Bankar, A.; Zinjarde, S.; Shinde, M.; Gopalghare, G.; Ravikumar, A. Heavy metal tolerance in marine strains of Yarrowia lipolytica. Extremophiles 2018, 22, 617–628. [Google Scholar] [CrossRef] [PubMed]
- García-Béjar, B.; Arévalo-Villena, M.; Guisantes-Batan, E.; Rodríguez-Flores, J.; Briones, A. Study of the bioremediatory capacity of wild yeasts. Sci. Rep. 2020, 10, 11265. [Google Scholar] [CrossRef] [PubMed]
- de Nicola, R.; Hall, N.; Melville, S.G.; Walker, G.M. Influence of Zinc on Distiller’s Yeast: Cellular Accumulation of Zinc and Impact on Spirit Congeners. J. Inst. Brew. 2009, 115, 265–271. [Google Scholar] [CrossRef]
- Simm, C.; Lahner, B.; Salt, D.; LeFurgey, A.; Ingram, P.; Yandell, B.; Eide, D.J. Saccharomyces cerevisiae Vacuole in Zinc Storage and Intracellular Zinc Distribution. Eukaryot. Cell 2007, 6, 1166–1177. [Google Scholar] [CrossRef]
- MacDiarmid, C.W.; Milanick, M.A.; Eide, D.J. Induction of the ZRC1 Metal Tolerance Gene in Zinc-limited Yeast Confers Resistance to Zinc Shock. J. Biol. Chem. 2003, 278, 15065–15072. [Google Scholar] [CrossRef]
- Strouhal, M.; Kizek, R.; Vacek, J.; Trnková, L.; Němec, M. Electrochemical study of heavy metals and metallothionein in yeast Yarrowia lipolytica. Bioelectrochemistry 2003, 60, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Eide, D.J. Transcription factors and transporters in zinc homeostasis: Lessons learned from fungi. Crit. Rev. Biochem. Mol. Biol. 2020, 55, 88–110. [Google Scholar] [CrossRef]
- Nguyen, T.Q.; Dziuba, N.; Lindahl, P.A. Isolated Saccharomyces cerevisiae vacuoles contain low-molecular-mass transition-metal polyphosphate complexes. Metallomics 2019, 11, 1298–1309. [Google Scholar] [CrossRef]
- Krężel, A.; Maret, W. The biological inorganic chemistry of zinc ions. Arch. Biochem. Biophys. 2016, 611, 3–19. [Google Scholar] [CrossRef]
- Sun, J.; Xu, S.; Du, Y.; Yu, K.; Jiang, Y.; Weng, H.; Yuan, W. Accumulation and Enrichment of Trace Elements by Yeast Cells and Their Applications: A Critical Review. Microorganisms 2022, 10, 1746. [Google Scholar] [CrossRef]
- Bird, A.J.; Wilson, S. Zinc homeostasis in the secretory pathway in yeast. Curr. Opin. Chem. Biol. 2020, 55, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.; Li, R.; Huang, Y. Zinc homeostasis in Schizosaccharomyces pombe. Arch. Microbiol. 2023, 205, 126. [Google Scholar] [CrossRef]
- Zhao, Y.-Y.; Cao, C.-L.; Liu, Y.-L.; Wang, J.; Li, J.; Li, S.-Y.; Deng, Y. Identification of the Genetic Requirements for Zinc Tolerance and Toxicity in Saccharomyces cerevisiae. G3 Genes|Genomes|Genetics 2020, 10, 479–488. [Google Scholar] [CrossRef]
- Leonardi, A.; Zanoni, S.; De Lucia, M.; Amaretti, A.; Raimondi, S.; Rossi, M. Zinc Uptake by Lactic Acid Bacteria. ISRN Biotechnol. 2013, 2013, 312917. [Google Scholar] [CrossRef]
- Yusof, H.M.; Mohamad, R.; Zaidan, U.H.; Rahman, N.A. Sustainable microbial cell nanofactory for zinc oxide nanoparticles production by zinc-tolerant probiotic Lactobacillus plantarum strain TA4. Microb. Cell Factories 2020, 19, 10. [Google Scholar] [CrossRef]
- Papadimitriou, K.; Alegría, Á.; Bron, P.A.; de Angelis, M.; Gobbetti, M.; Kleerebezem, M.; Lemos, J.A.; Linares, D.M.; Ross, P.; Stanton, C.; et al. Stress Physiology of Lactic Acid Bacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 837–890. [Google Scholar] [CrossRef] [PubMed]
- Huynh, U.; Qiao, M.; King, J.; Trinh, B.; Valdez, J.; Haq, M.; Zastrow, M.L. Differential Effects of Transition Metals on Growth and Metal Uptake for Two Distinct Lactobacillus Species. Microbiol. Spectr. 2022, 10, e0100621. [Google Scholar] [CrossRef]
- Mrvčić, J.; Stanzer, D.; Šolić, E.; Stehlik-Tomas, V. Interaction of lactic acid bacteria with metal ions: Opportunities for improving food safety and quality. World J. Microbiol. Biotechnol. 2012, 28, 2771–2782. [Google Scholar] [CrossRef] [PubMed]
- Clément, H.; Prost, C.; Chiron, H.; Bonnand-Ducasse, M.; Della Valle, G.; Courcoux, P.; Onno, B. The effect of organic wheat flour by-products on sourdough performances assessed by a multi-criteria approach. Food Res. Int. 2018, 106, 974–981. [Google Scholar] [CrossRef]
- Meng, Y.; Liang, Z.; Yi, M.; Tan, Y.; Li, Z.; Du, P.; Li, A.; Li, C.; Liu, L. Enrichment of zinc in Lactobacillus plantarum DNZ-4: Impact on its characteristics, metabolites and antioxidant activity. LWT 2022, 153, 112462. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward power | 1550 W |
| Cool gas flow | 15 L min−1 |
| Auxiliary gas flow | 0.9 L min−1 (Argon) |
| Nebulizer gas flow | 0.9 L min−1 (Argon) |
| Nebulizer type | MicroMist |
| Quadrupole (m/z) | 66 (Zn), 103 (Rh) |
| Limit of quantitation | 0.6 µg L−1 (Zn) |
| Calibration range | 1–100 µg L−1 (Zn) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossi, S.; Maares, M.; Kieserling, H.; Rohn, S.; Schlüter, O.; Patrignani, F.; Lanciotti, R.; Haase, H.; Keil, C. Zinc Tolerance of Special Yeasts and Lactic Acid Bacteria for Use in the Food Industry. Fermentation 2023, 9, 521. https://doi.org/10.3390/fermentation9060521
Rossi S, Maares M, Kieserling H, Rohn S, Schlüter O, Patrignani F, Lanciotti R, Haase H, Keil C. Zinc Tolerance of Special Yeasts and Lactic Acid Bacteria for Use in the Food Industry. Fermentation. 2023; 9(6):521. https://doi.org/10.3390/fermentation9060521
Chicago/Turabian StyleRossi, Samantha, Maria Maares, Helena Kieserling, Sascha Rohn, Oliver Schlüter, Francesca Patrignani, Rosalba Lanciotti, Hajo Haase, and Claudia Keil. 2023. "Zinc Tolerance of Special Yeasts and Lactic Acid Bacteria for Use in the Food Industry" Fermentation 9, no. 6: 521. https://doi.org/10.3390/fermentation9060521