
Dynamic Responses of Streptomyces albulus QLU58 and Its Acid-Tolerant Derivatives to the Autoacidification in ε-Poly-l-Lysine Production

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Inoculum Preparation
2.2. Screening of the Acid-Tolerant Mutants
2.3. Batch Fermentation
2.4. Measurement of ε-PL Production and Dry Cell Weights
2.5. Measurement of Cellular Respiratory Activity
2.6. Measurement of Intracellular pH
2.7. Measurement of Intracellular ATP and H+-ATPase
2.8. Intracellular Amino Acid Analyses
2.9. Analysis of Fatty Acids on Cell Membrane
2.10. Quantitative Reverse Transcriptase-PCR (qRT-PCR) Analysis
2.11. Statistical Analysis
3. Results and Discussion
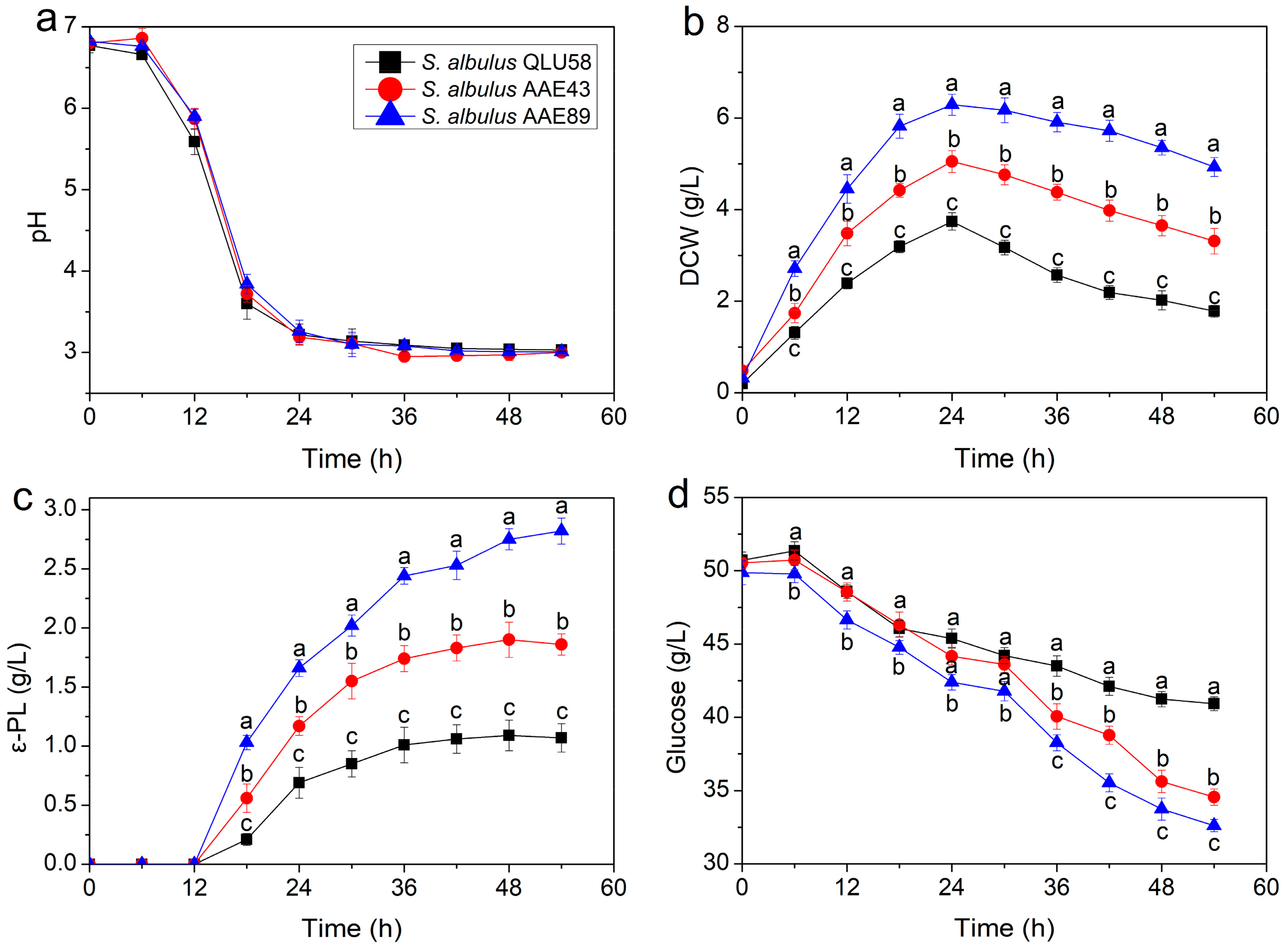
3.1. Changes in the Fermentation Parameters in Batch Fermentations for ε-PL Production by S. albulus
3.1.1. Kinetic Parameters
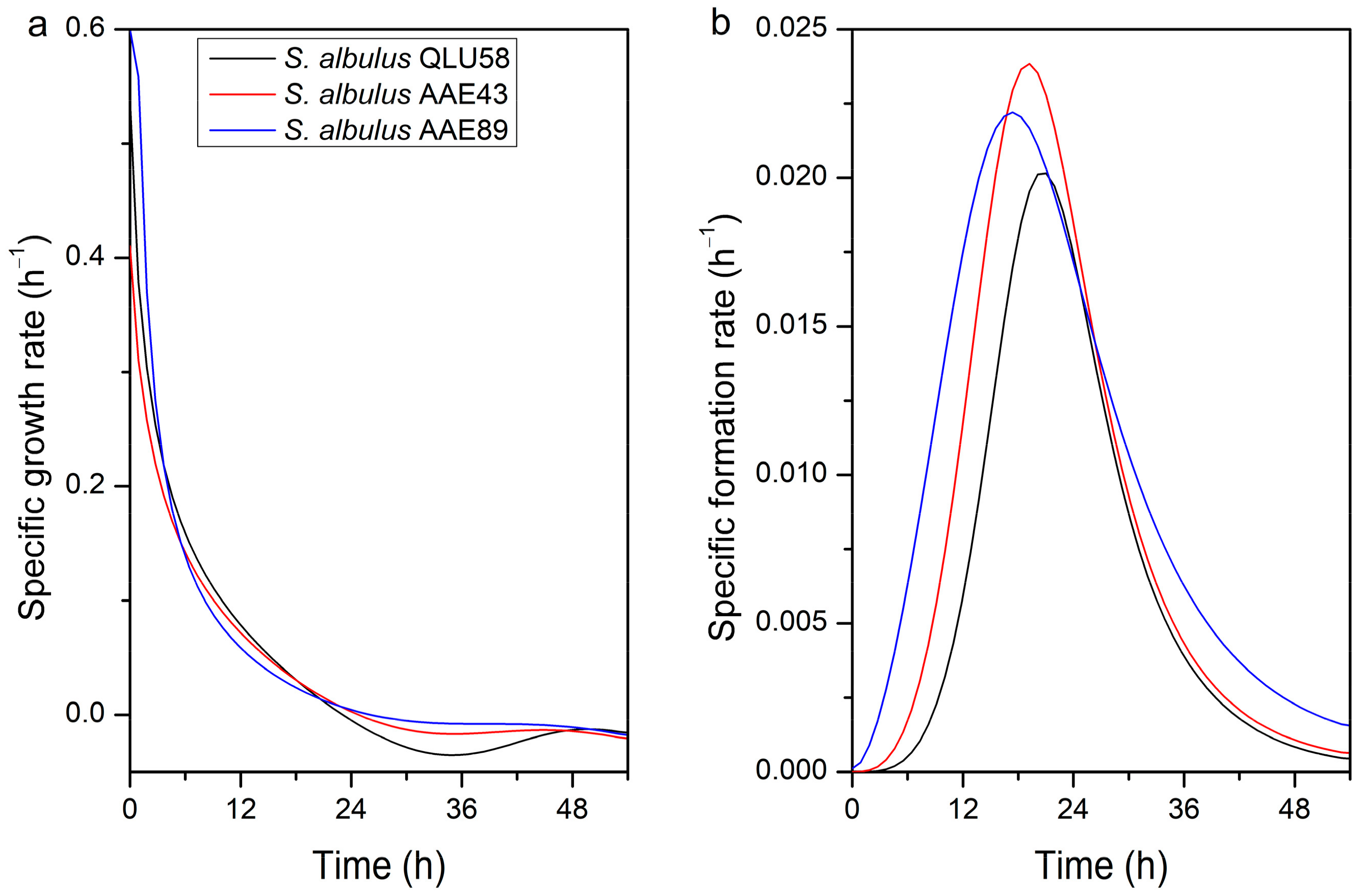
3.1.2. Specific Rates of Cell Growth and ε-PL Production
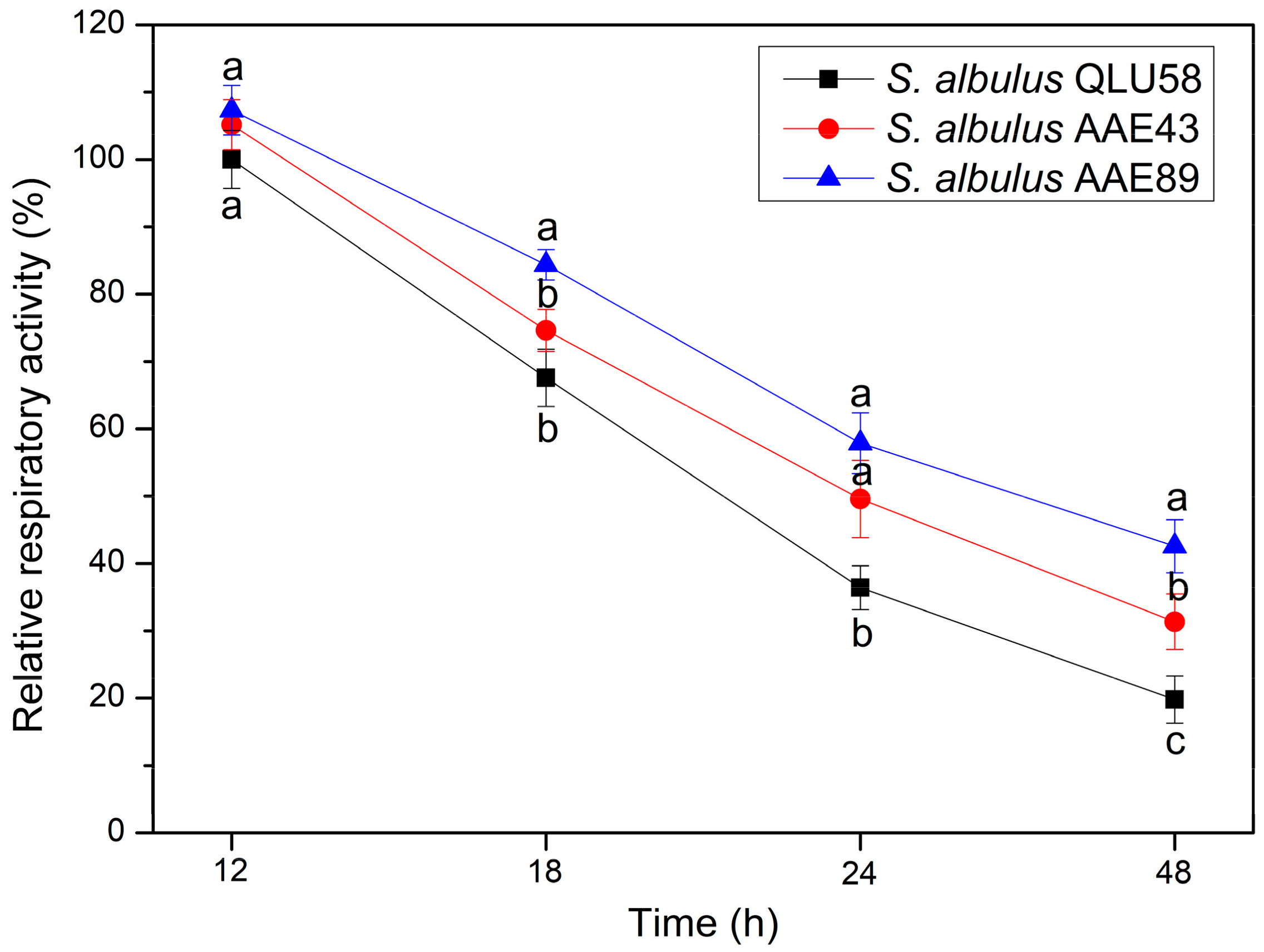
3.1.3. Respiratory Activity
3.2. Maintenance of Intracellular pH
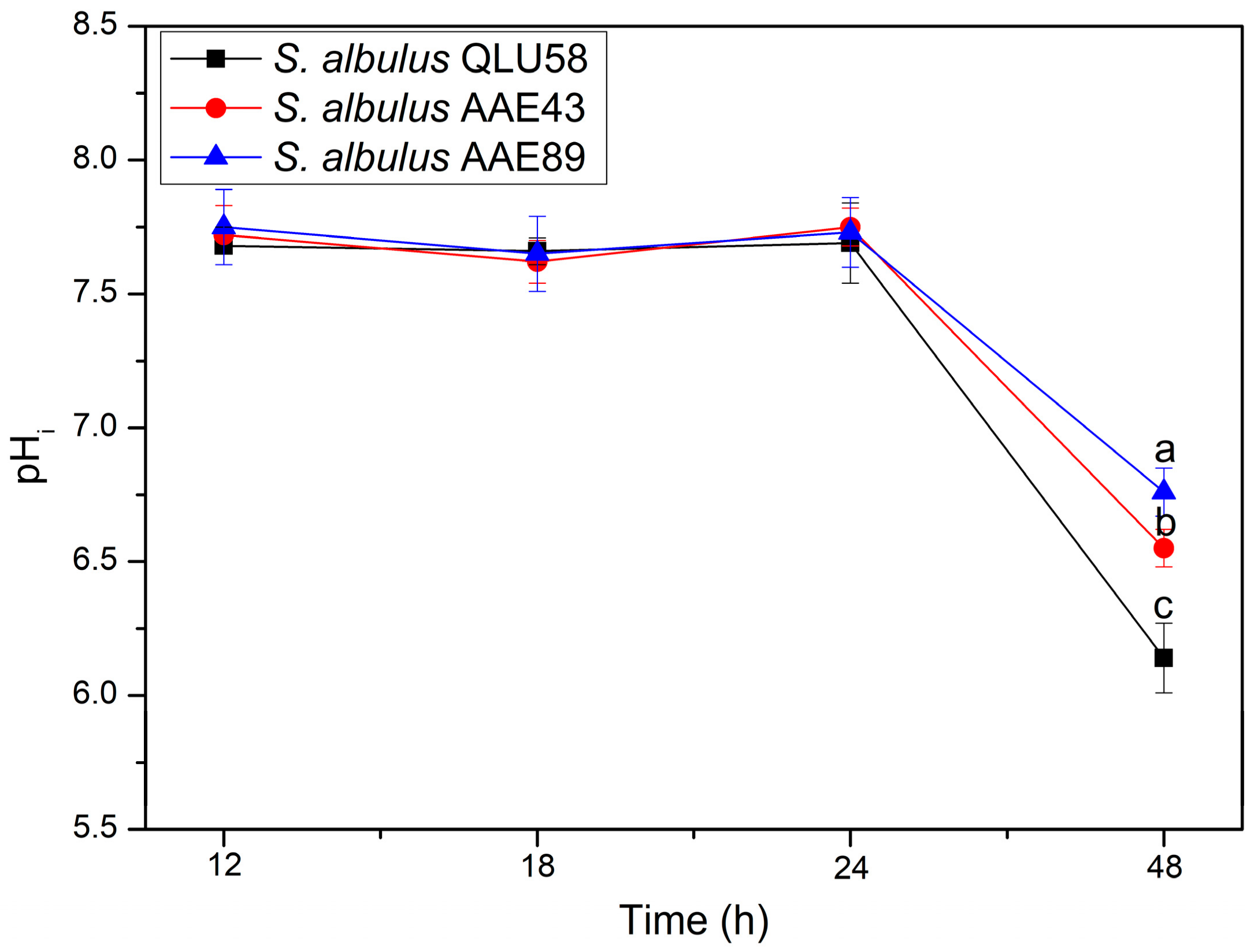
3.2.1. Intracellular pH
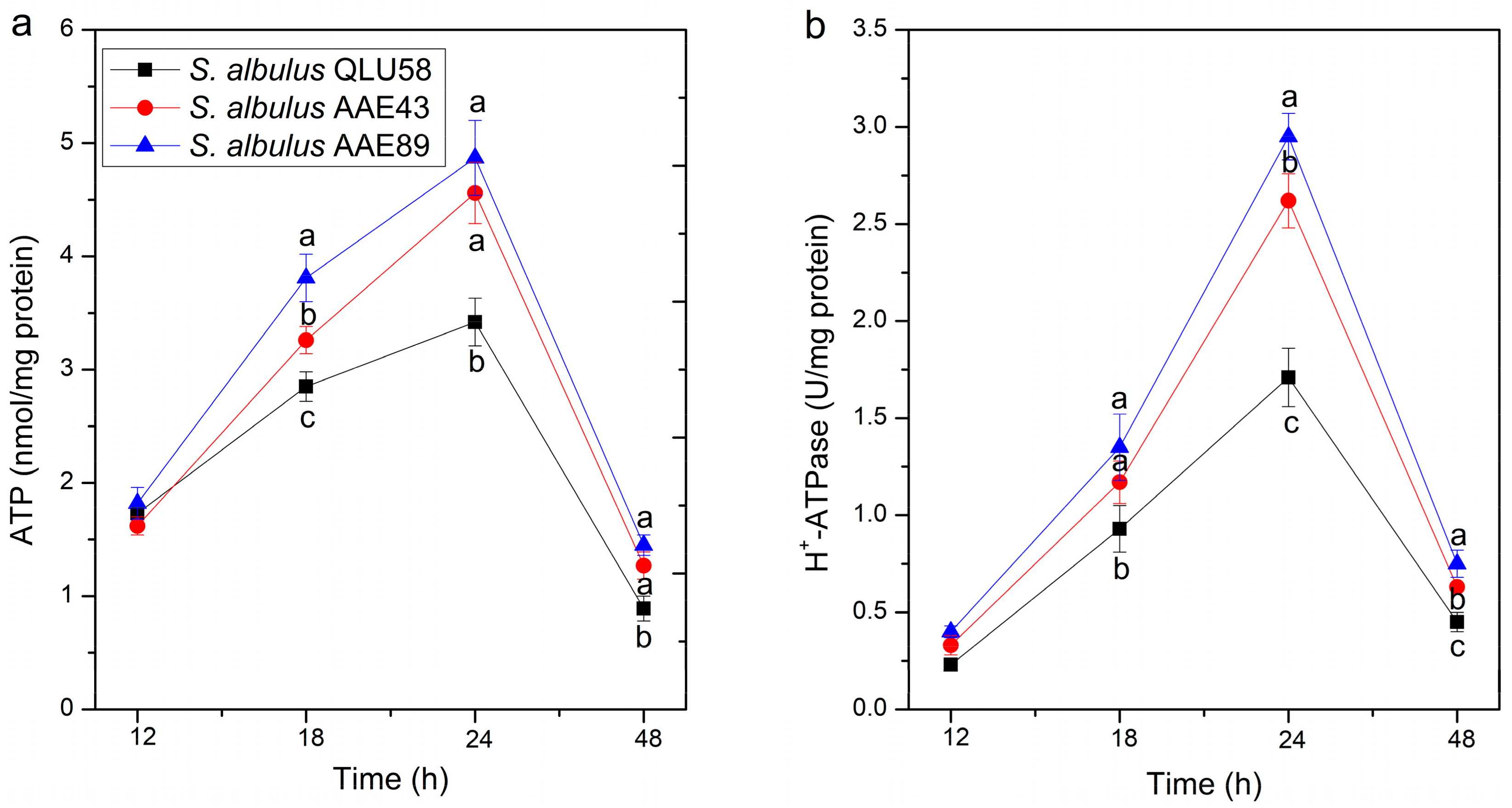
3.2.2. Intracellular ATP Concentration and H+-ATPase Activity
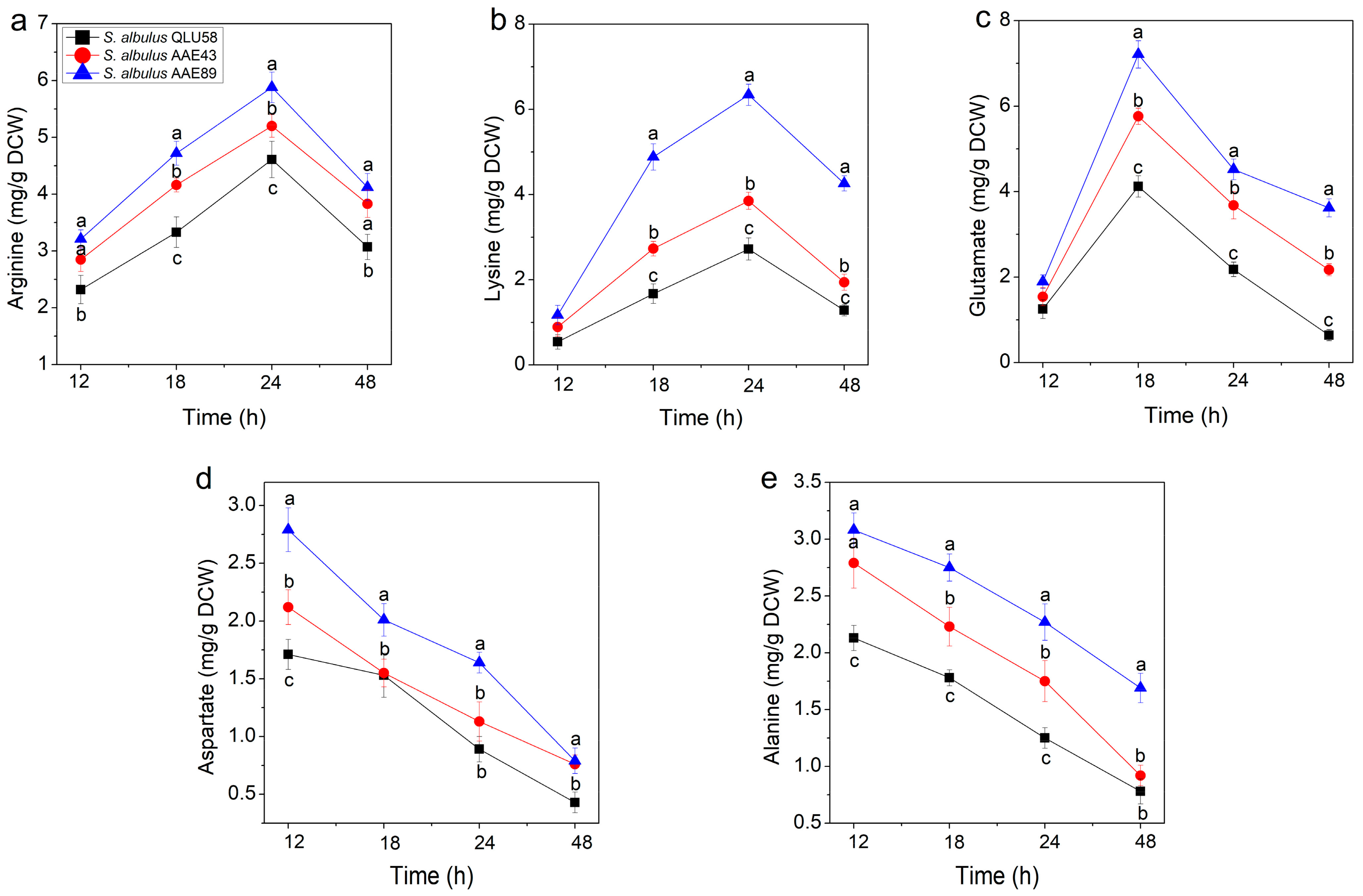
3.2.3. Intracellular Amino Acid Content
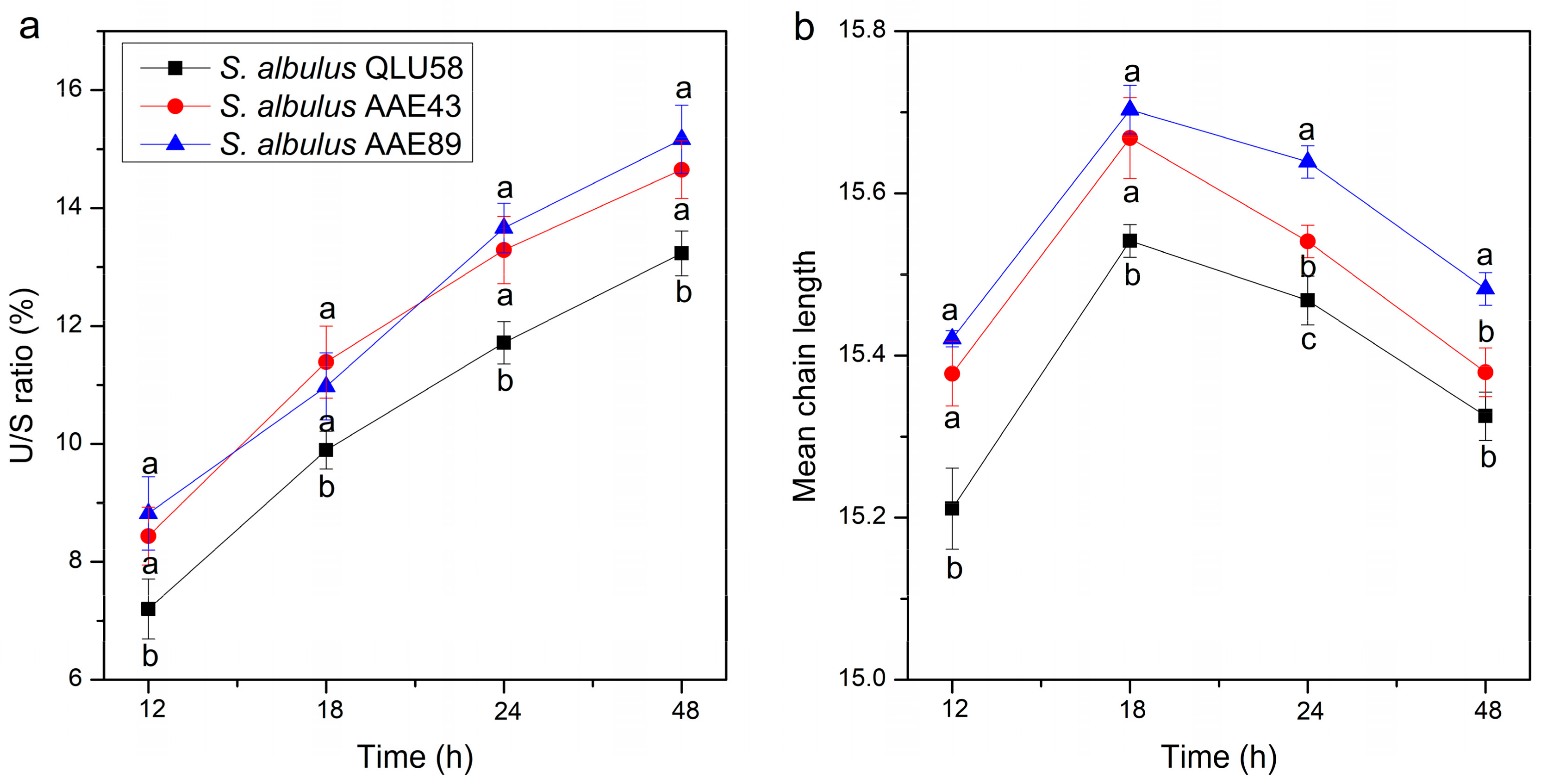
3.3. Changes in Cell Membrane Fatty Acids
3.4. Comparisons of the Transcriptional Level of Key Genes between Original and Mutational Strains during Autoacidification
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xu, Z.; Xu, Z.; Feng, X.; Xu, D.; Liang, J.; Xu, H. Recent advances in the biotechnological production of microbial poly (ɛ-L-lysine) and understanding of its biosynthetic mechanism. Appl. Microbiol. Biotechnol. 2016, 100, 6619–6630. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.D.; Chen, X.S.; Zeng, X.; Wang, L.; Tang, L.; Mao, Z.G. Acidic pH shock induced overproduction of ε-poly-l-lysine in fed-batch fermentation by Streptomyces sp. M-Z18 from agro-industrial by-products. Bioprocess Biosyst. Eng. 2015, 38, 1113–1125. [Google Scholar] [CrossRef] [PubMed]
- Lund, P.; Tramonti, A.; De Biase, D. Coping with low pH: Molecular strategies in neutralophilic bacteria. FEMS Microbiol. Rev. 2014, 38, 1091–1125. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Ren, X.; Yu, C.; Wang, J.; Wang, L.; Zhuge, X.; Liu, X. Physiological and Transcriptional Responses of Streptomyces albulus to Acid Stress in the Biosynthesis of ε-Poly-l-lysine. Front. Microbiol. 2020, 11, 1379. [Google Scholar] [CrossRef]
- Wu, C.; Zhang, J.; Chen, W.; Wang, M.; Du, G.; Chen, J. A combined physiological and proteomic approach to reveal lactic-acid-induced alterations in Lactobacillus casei Zhang and its mutant with enhanced lactic acid tolerance. Appl. Microbiol. Biotechnol. 2012, 93, 707–722. [Google Scholar] [CrossRef]
- He, S.K.; Ye, B.N.; Zhang, Z.F.; Cui, Y.; Wang, S.Y.; Shi, X.M. Cross-protective effect of acid adaptation on ethanol tolerance in Salmonella enteritidis. Food Sci. Hum. Wellness 2023, 12, 1402–1407. [Google Scholar] [CrossRef]
- Itzhaki, R.F. Colorimetric method for estimating polylysine and polyarginine. Anal. Biochem. 1972, 50, 569–574. [Google Scholar] [CrossRef]
- Winding, A.; Binnerup, S.J.; Sørensen, J. Viability of indigenous soil bacteria assayed by respiratory activity and growth. Appl. Environ. Microbiol. 1994, 60, 2869–2875. [Google Scholar] [CrossRef]
- Breeuwer, P.; Drocourt, J.l.; Rombouts, F.M.; Abee, T. A novel method for continuous determination of the intracellular pH in bacteria with the internally conjugated fluorescent probe 5 (and 6-)-carboxyfluorescein succinimidyl ester. Appl. Environ. Microbiol. 1996, 62, 178–183. [Google Scholar] [CrossRef]
- Cichna, M.; Raab, M.; Daxecker, H.; Griesmacher, A.; Müller, M.; Markl, P. Determination of fifteen nucleotides in cultured human mononuclear blood and umbilical vein endothelial cells by solvent generated ion-pair chromatography. J. Chromatogr. B 2003, 787, 381–391. [Google Scholar] [CrossRef]
- Fountoulakis, M.; Lahm, H.W. Hydrolysis and amino acid composition analysis of proteins. J. Chromatogr. A 1998, 826, 109–134. [Google Scholar] [CrossRef] [PubMed]
- Sasser, M. Identification of bacteria by gas chromatography of cellular fatty acids. In MIDI Technical Note 101; MIDI, Inc.: Newark, NJ, USA, 1990; pp. 1–7. [Google Scholar]
- Chen, X.S.; Wang, K.F.; Zheng, G.C.; Gao, Y.; Mao, Z.G. Preparation, characterization and antimicrobial activity of ε-poly-l-lysine with short chain length produced from glycerol by Streptomyces albulus. Process Biochem. 2018, 68, 22–29. [Google Scholar] [CrossRef]
- Kahar, P.; Iwata, T.; Hiraki, J.; Park, E.Y.; Okabe, M. Enhancement of ε-polylysine production by Streptomyces albulus strain 410 using pH control. J. Biosci. Bioeng. 2001, 91, 190–194. [Google Scholar] [CrossRef]
- Chen, X.S.; Li, S.; Liao, L.J.; Ren, X.D.; Li, F.; Tang, L.; Zhang, J.H.; Mao, Z.G. Production of ε-poly-l-lysine using a novel two-stage pH control strategy by Streptomyces sp. M-Z18 from glycerol. Bioprocess Biosyst. Eng. 2011, 34, 561–567. [Google Scholar] [CrossRef]
- Yamanaka, K.; Kito, N.; Imokawa, Y.; Maruyama, C.; Utagawa, T.; Hamano, Y. Mechanism of ε-poly-l-lysine production and accumulation revealed by identification and analysis of an ε-poly-l-lysine-degrading enzyme. Appl. Environ. Microbiol. 2010, 76, 5669–5675. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Sun, Y.F.; Tang, X.; He, C.N.; Shao, Y.L.; Tang, Y.J.; Zhou, W.W. Alkaline pH shock enhanced production of validamycin A in fermentation of Streptomyces hygroscopicus. Bioresour. Technol. 2018, 249, 234–240. [Google Scholar] [CrossRef]
- Guan, N.; Liu, L.; Shin, H.-d.; Chen, R.R.; Zhang, J.; Li, J.; Du, G.; Shi, Z.; Chen, J. Systems-level understanding of how Propionibacterium acidipropionici respond to propionic acid stress at the microenvironment levels: Mechanism and application. J. Biotechnol. 2013, 167, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Cagnac, O.; Leterrier, M.; Yeager, M.; Blumwald, E. Identification and characterization of Vnx1p, a novel type of vacuolar monovalent cation/H+ antiporter of Saccharomyces cerevisiae. J. Biol. Chem. 2007, 282, 24284–24293. [Google Scholar] [CrossRef]
- Baker Austin, C.; Dopson, M. Life in acid: pH homeostasis in acidophiles. Trends Microbiol. 2007, 15, 165–171. [Google Scholar] [CrossRef]
- Xu, Y.; Zhao, Z.; Tong, W.; Ding, Y.; Liu, B.; Shi, Y.; Wang, J.; Sun, S.; Liu, M.; Wang, Y. An acid-tolerance response system protecting exponentially growing Escherichia coli. Nat. Commun. 2020, 11, 1496. [Google Scholar] [CrossRef]
- Wu, C.; Zhang, J.; Du, G.; Chen, J. Aspartate protects Lactobacillus casei against acid stress. Appl. Microbiol. Biotechnol. 2013, 97, 4083–4093. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wu, C.; Du, G.; Chen, J. Enhanced acid tolerance in Lactobacillus casei by adaptive evolution and compared stress response during acid stress. Biotechnol. Bioprocess Eng. 2012, 17, 283–289. [Google Scholar] [CrossRef]
- Eun Rhee, J.; Haeng Rhee, J.; Youl Ryu, P.; Ho Choi, S. Identification of the cadBA operon from Vibrio vulnificus and its influence on survival to acid stress. FEMS Microbiol. Lett. 2002, 208, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Budin Verneuil, A.; Pichereau, V.; Auffray, Y.; Ehrlich, D.S.; Maguin, E. Proteomic characterization of the acid tolerance response in Lactococcus lactis MG1363. Proteomics 2005, 5, 4794–4807. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Feng, S.; Tong, Y.; Yang, H. Adaptive mechanism of Acidithiobacillus thiooxidans CCTCC M 2012104 under stress during bioleaching of low-grade chalcopyrite based on physiological and comparative transcriptomic analysis. J. Ind. Microbiol. Biot. 2019, 46, 1643–1656. [Google Scholar] [CrossRef]
- Fernández, M.; Zúñiga, M. Amino acid catabolic pathways of lactic acid bacteria. Crit. Rev. Microbiol. 2006, 32, 155–183. [Google Scholar] [CrossRef] [PubMed]
- Diez Gonzalez, F.; Karaibrahimoglu, Y. Comparison of the glutamate-, arginine- and lysine- dependent acid resistance systems in Escherichia coli O157: H7. J. Appl. Microbiol. 2004, 96, 1237–1244. [Google Scholar] [CrossRef] [PubMed]
- Iyer, R.; Williams, C.; Miller, C. Arginine-agmatine antiporter in extreme acid resistance in Escherichia coli. J. Bacteriol. 2003, 185, 6556–6561. [Google Scholar] [CrossRef]
- Senouci Rezkallah, K.; Schmitt, P.; Jobin, M.P. Amino acids improve acid tolerance and internal pH maintenance in Bacillus cereus ATCC14579 strain. Food Microbiol. 2011, 28, 364–372. [Google Scholar] [CrossRef]
- Wu, H.; Zhao, Y.; Du, Y.; Miao, S.; Liu, J.; Li, Y.; Caiyin, Q.; Qiao, J. Quantitative proteomics of Lactococcus lactis F44 under cross-stress of low pH and lactate. J. Dairy Sci. 2018, 101, 6872–6884. [Google Scholar] [CrossRef]
- Higuchi, T.; Hayashi, H.; Abe, K. Exchange of glutamate and gamma-aminobutyrate in a Lactobacillus strain. J. Bacteriol. 1997, 179, 3362–3364. [Google Scholar] [CrossRef] [PubMed]
- De Biase, D.; Pennacchietti, E. Glutamate decarboxylase-dependent acid resistance in orally acquired bacteria: Function, distribution and biomedical implications of the gadBC operon. Mol. Microbiol. 2012, 86, 770–786. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Lu, P.; Zhang, Y.; Li, L.; Chen, S. Characterization of an aspartate-dependent acid survival system in Yersinia pseudotuberculosis. FEBS Lett. 2010, 584, 2311–2314. [Google Scholar] [CrossRef] [PubMed]
- Typas, A.; Banzhaf, M.; Gross, C.A.; Vollmer, W. From the regulation of peptidoglycan synthesis to bacterial growth and morphology. Nat. Rev. Microbiol. 2012, 10, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Boyd, D.A.; Cvitkovitch, D.G.; Bleiweis, A.S.; Kiriukhin, M.Y.; Debabov, D.V.; Neuhaus, F.C.; Hamilton, I.R. Defects in D-alanyl-lipoteichoic acid synthesis in Streptococcus mutans results in acid sensitivity. J. Bacteriol. 2000, 182, 6055–6065. [Google Scholar] [CrossRef] [PubMed]
- Takehara, M.; Hibino, A.; Saimura, M.; Hirohara, H. High-yield production of short chain length poly (ε-L-lysine) consisting of 5-20 residues by Streptomyces aureofaciens, and its antimicrobial activity. Biotechnol. Lett. 2010, 32, 1299–1303. [Google Scholar] [CrossRef]
- Yamanaka, K.; Maruyama, C.; Takagi, H.; Hamano, Y. ε-Poly-l-lysine dispersity is controlled by a highly unusual nonribosomal peptide synthetase. Nat. Chem. Biol. 2008, 4, 766–772. [Google Scholar] [CrossRef]
- Tran, N.T.; Huang, X.; Hong, H.J.; Bush, M.J.; Chandra, G.; Pinto, D.; Bibb, M.J.; Hutchings, M.I.; Mascher, T.; Buttner, M.J. Defining the regulon of genes controlled by σE, a key regulator of the cell envelope stress response in Streptomyces coelicolor. Mol. Microbiol. 2019, 112, 461–481. [Google Scholar] [CrossRef]
- Giotis, E.S.; McDowell, D.A.; Blair, I.S.; Wilkinson, B.J. Role of branched-chain fatty acids in pH stress tolerance in Listeria monocytogenes. Appl. Environ. Microbiol. 2007, 73, 997–1001. [Google Scholar] [CrossRef]
- Denich, T.; Beaudette, L.; Lee, H.; Trevors, J. Effect of selected environmental and physico-chemical factors on bacterial cytoplasmic membranes. J. Microbiol. Methods 2003, 52, 149–182. [Google Scholar] [CrossRef]
- Fozo, E.M.; Quivey, R.G., Jr. The fabM gene product of Streptococcus mutans is responsible for the synthesis of monounsaturated fatty acids and is necessary for survival at low pH. J. Bacteriol. 2004, 186, 4152–4158. [Google Scholar] [CrossRef] [PubMed]
- Liao, G.; Liu, Q.; Xie, J. Transcriptional analysis of the effect of exogenous decanoic acid stress on Streptomyces roseosporus. Microb. Cell Fact. 2013, 12, 19. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Zhang, J.; Wang, M.; Du, G.; Chen, J. Lactobacillus casei combats acid stress by maintaining cell membrane functionality. J. Ind. Microbiol. Biot. 2012, 39, 1031–1039. [Google Scholar] [CrossRef] [PubMed]
- Kolbeck, S.; Behr, J.; Vogel, R.F.; Ludwig, C.; Ehrmann, M.A. Acid stress response of Staphylococcus xylosus elicits changes in the proteome and cellular membrane. J. Appl. Microbiol. 2019, 126, 1480–1495. [Google Scholar] [CrossRef]
- Mykytczuk, N.; Trevors, J.; Leduc, L.; Ferroni, G. Fluorescence polarization in studies of bacterial cytoplasmic membrane fluidity under environmental stress. Prog. Biophys. Mol. Bio. 2007, 95, 60–82. [Google Scholar] [CrossRef]
- Cao Hoang, L.; Marechal, P.A.; Lê Thanh, M.; Gervais, P.; Waché, Y. Fluorescent probes to evaluate the physiological state and activity of microbial biocatalysts: A guide for prokaryotic and eukaryotic investigation. Biotechnol. J. 2008, 3, 890–903. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, X.; Chen, Y.; Guo, Y.; Li, K.; Wang, C.; Liu, X. Dynamic Responses of Streptomyces albulus QLU58 and Its Acid-Tolerant Derivatives to the Autoacidification in ε-Poly-l-Lysine Production. Fermentation 2023, 9, 459. https://doi.org/10.3390/fermentation9050459
Ren X, Chen Y, Guo Y, Li K, Wang C, Liu X. Dynamic Responses of Streptomyces albulus QLU58 and Its Acid-Tolerant Derivatives to the Autoacidification in ε-Poly-l-Lysine Production. Fermentation. 2023; 9(5):459. https://doi.org/10.3390/fermentation9050459
Chicago/Turabian StyleRen, Xidong, Yan Chen, Yangzi Guo, Kunpeng Li, Chenying Wang, and Xinli Liu. 2023. "Dynamic Responses of Streptomyces albulus QLU58 and Its Acid-Tolerant Derivatives to the Autoacidification in ε-Poly-l-Lysine Production" Fermentation 9, no. 5: 459. https://doi.org/10.3390/fermentation9050459