
Effects of Guanidinoacetic Acid on Ruminal Fermentation and Greenhouse Gas Production Using Fresh Forage and Silage from Different Maize (Zea mays L.) Genotypes

 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Treatments
2.2. Fresh Forage and Silage Production
2.3. Chemical Composition
2.4. In Vitro Incubation
2.4.1. Production of Total Gas, Methane (CH4), Carbon Monoxide (CO), and Hydrogen Sulfide (H2S)
2.4.2. Ruminal Potential of Hydrogen (pH) and Dry Matter Degradability (DMD)
2.4.3. Calculations
2.5. Statistical Analysis
3. Results
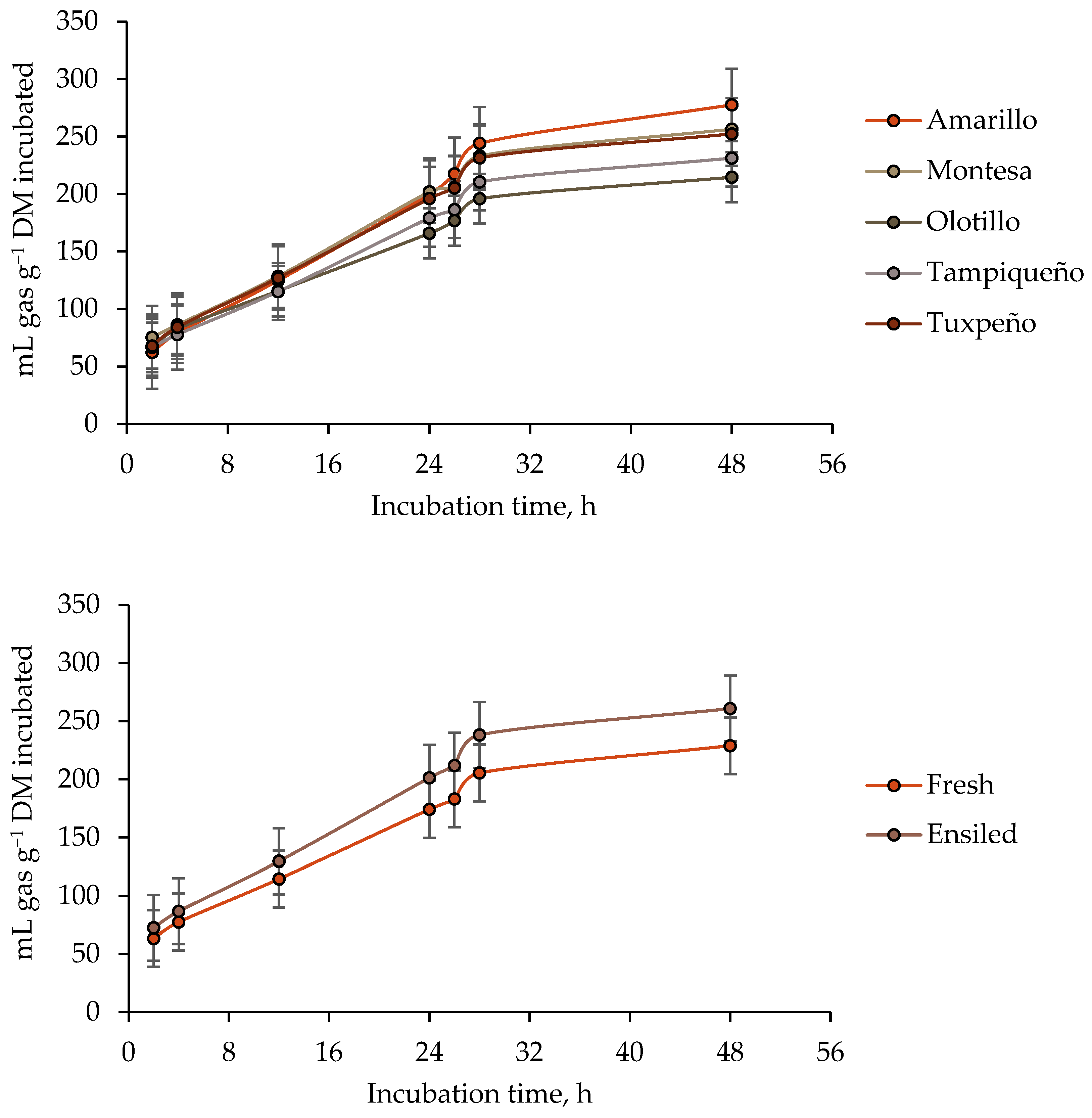
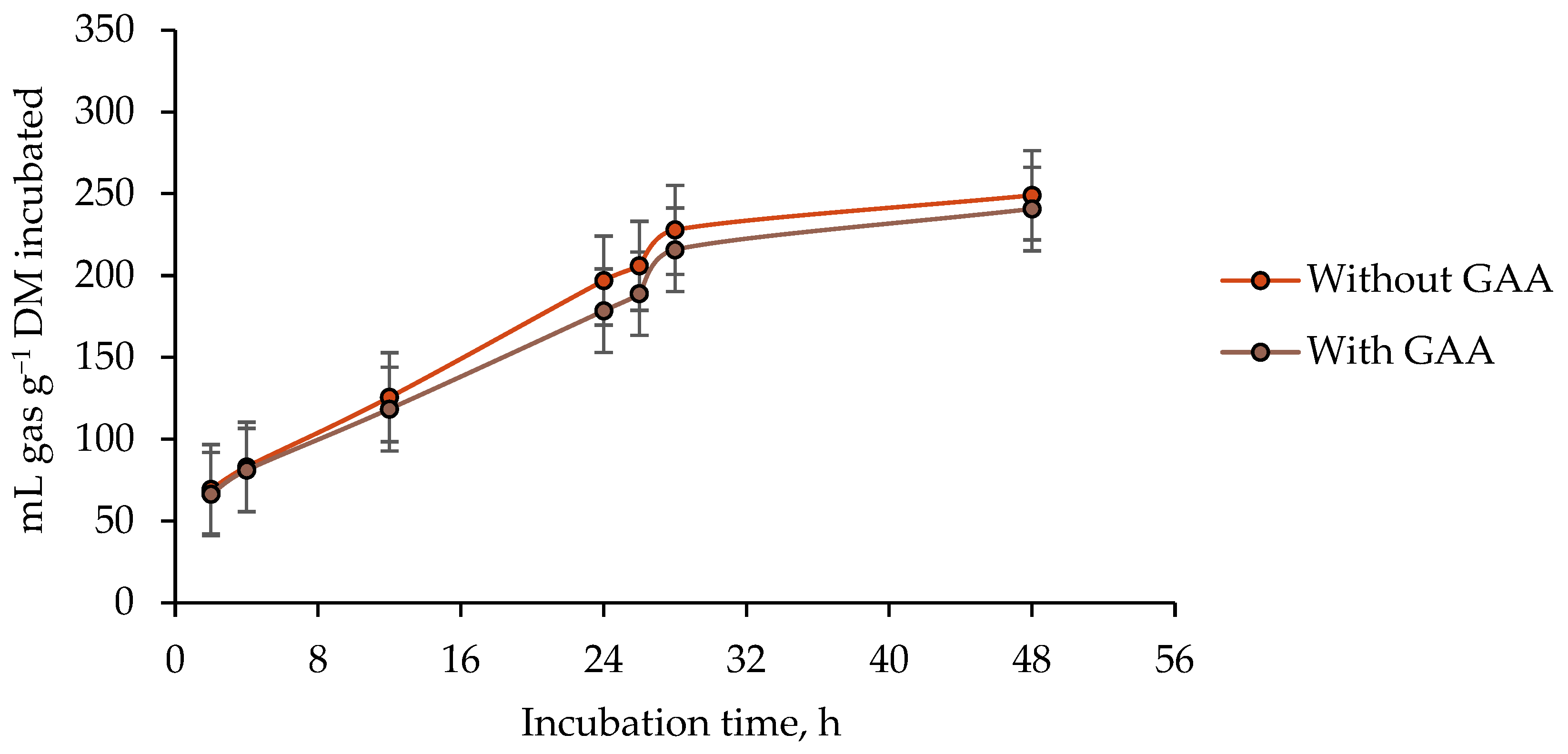
3.1. Ruminal Gas Total Production
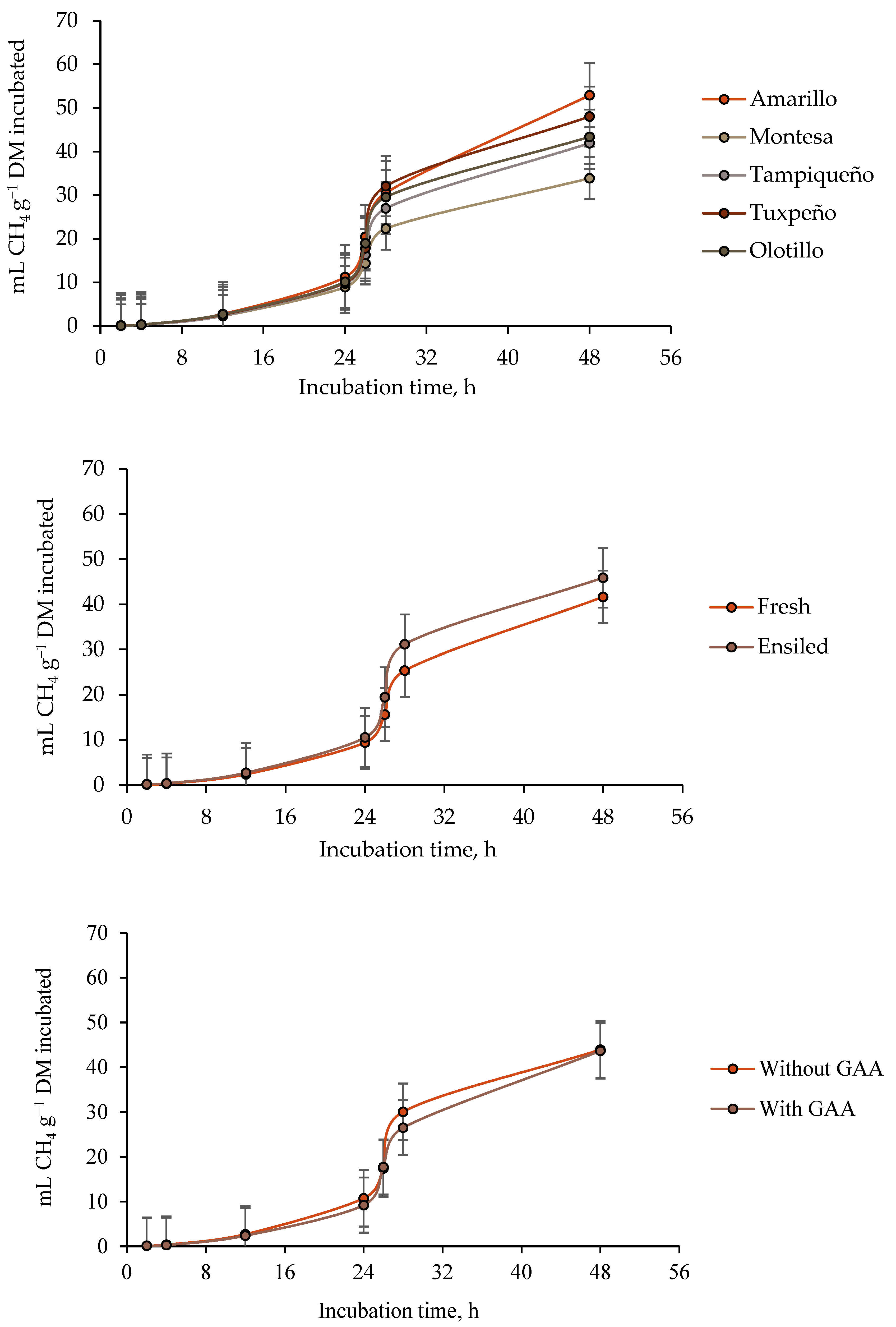
3.2. Ruminal Methane (CH4) Production
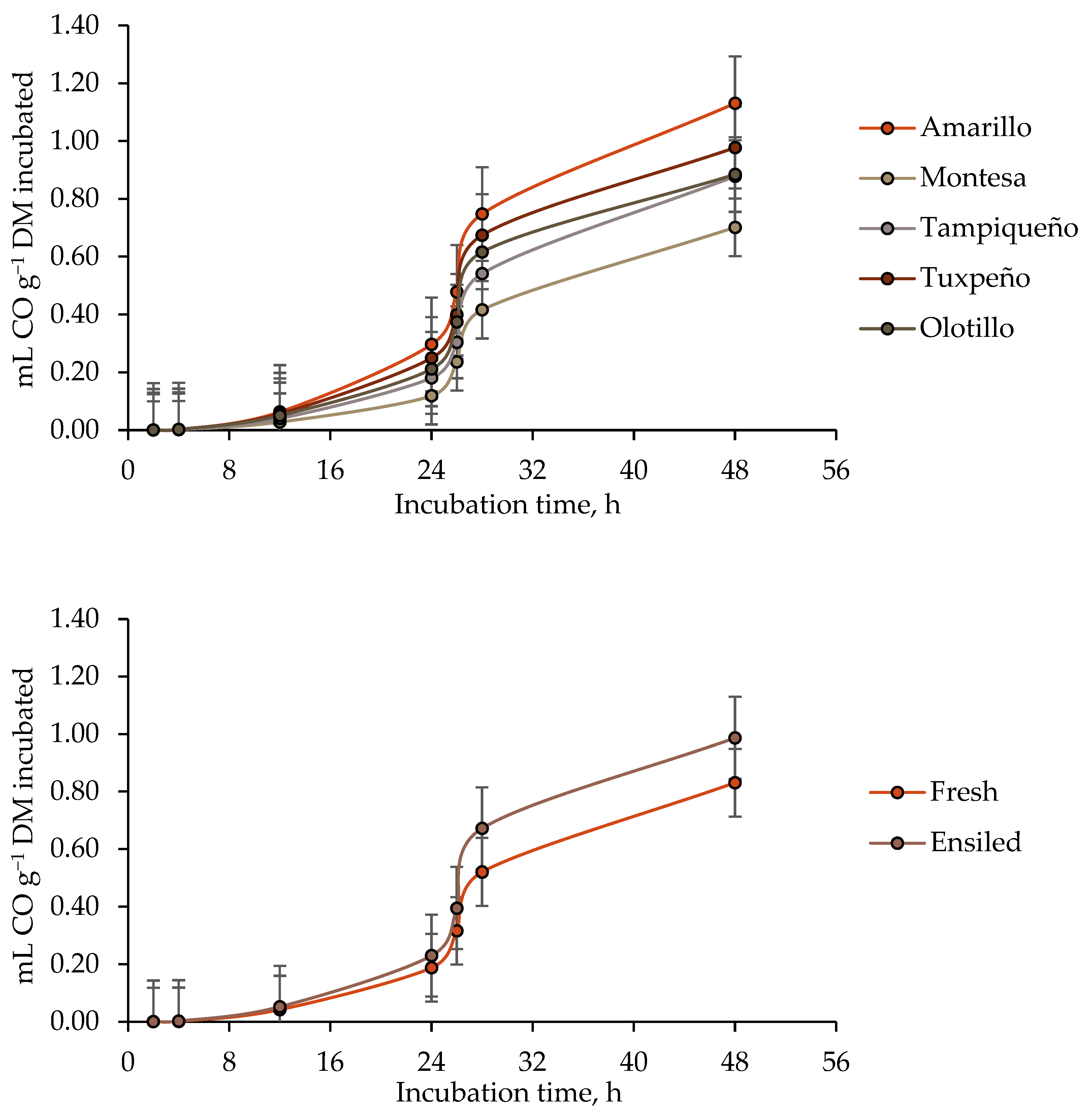
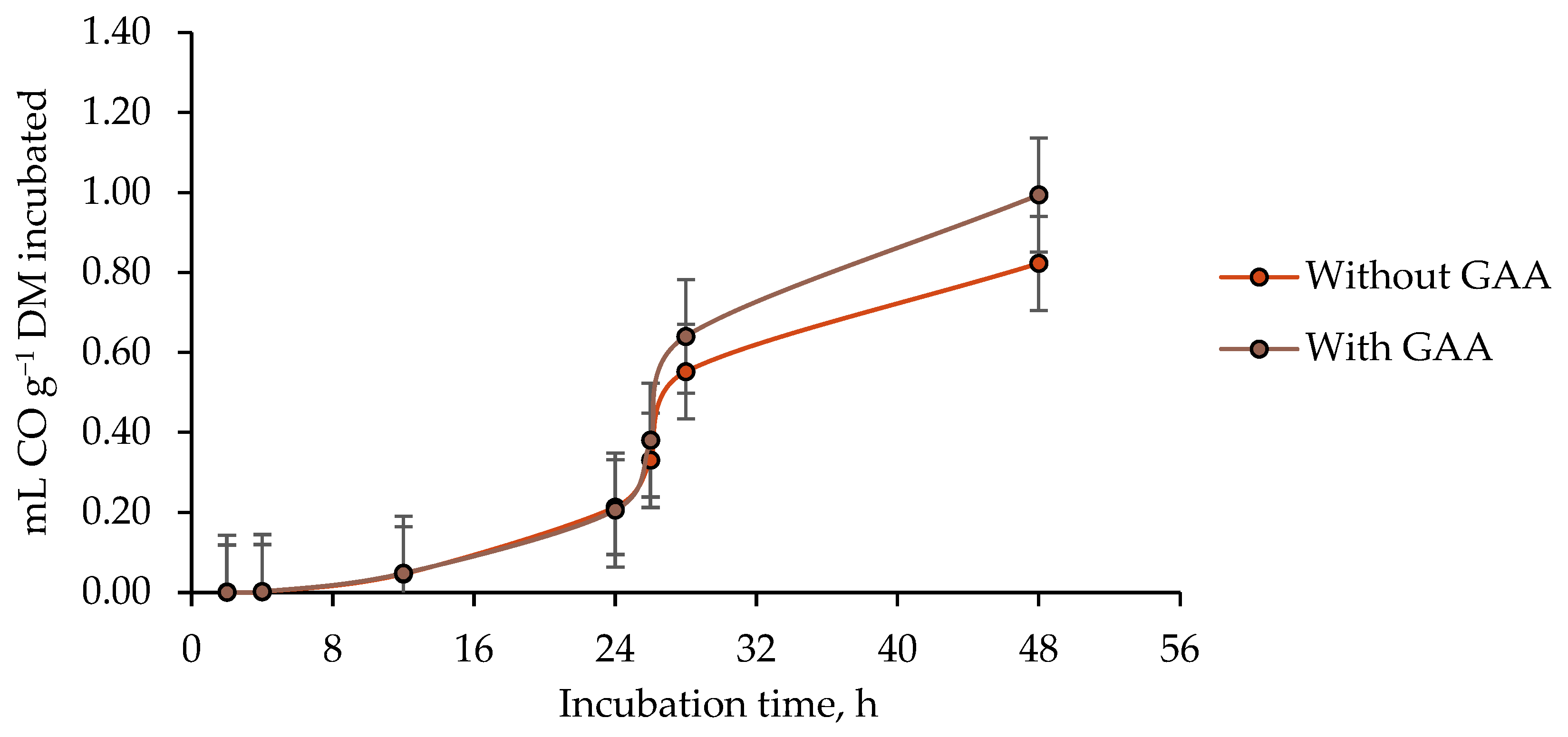
3.3. Ruminal Carbon Monoxide (CO) Production
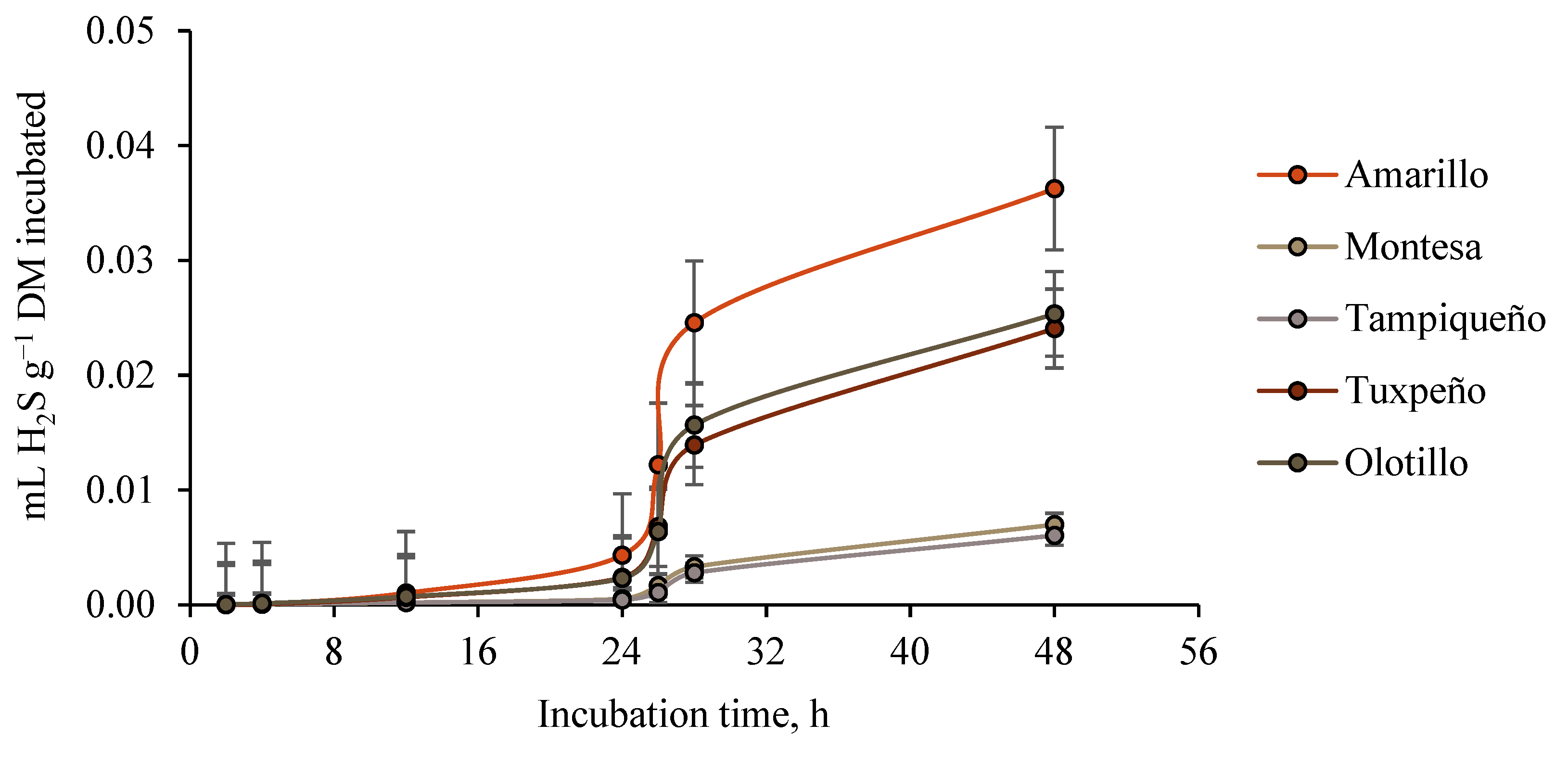
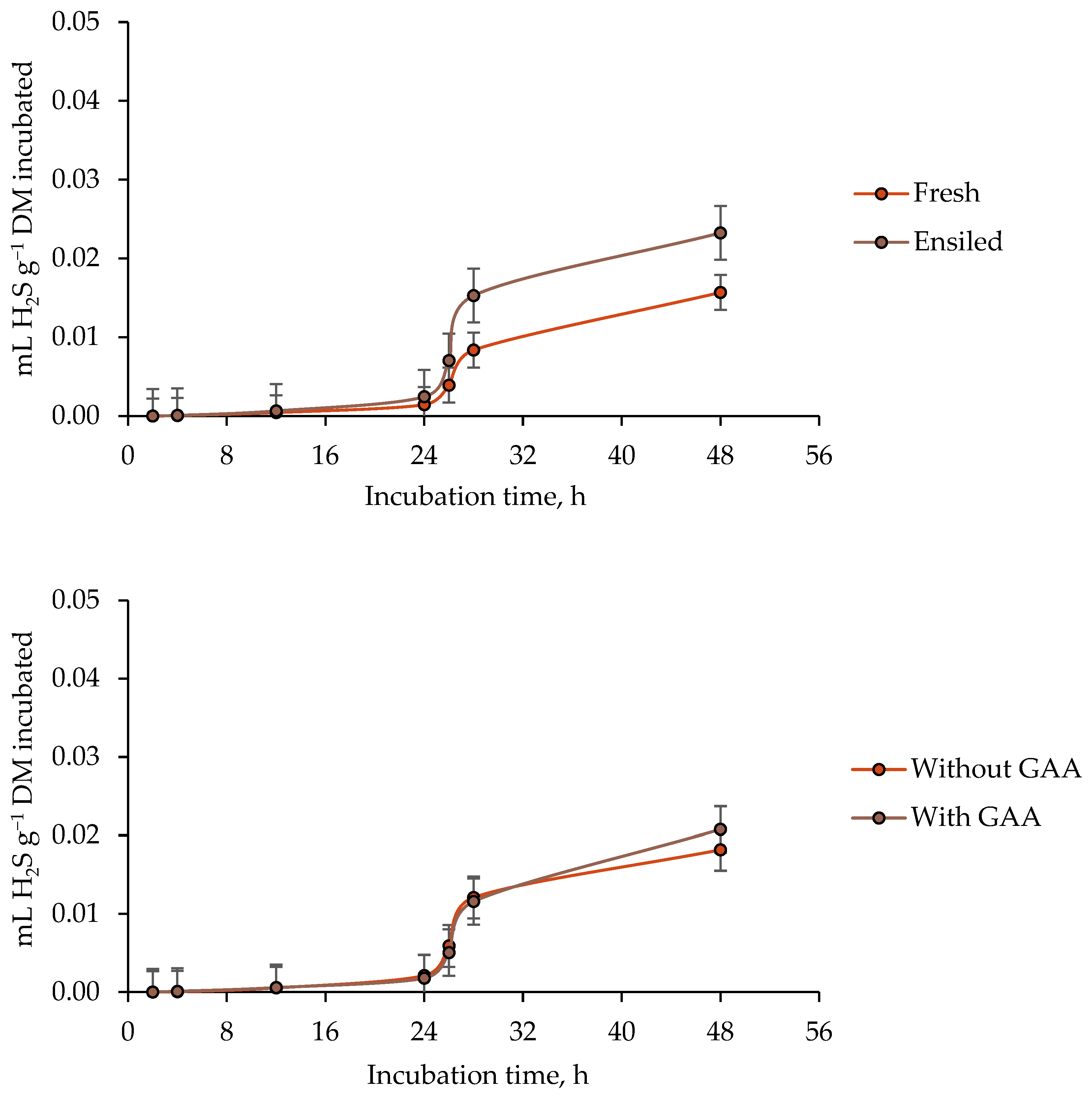
3.4. Ruminal Hydrogen Sulfide (H2S) Production
3.5. Fermentation Characteristics and CH4 Conversion Efficiency
4. Discussion
4.1. Ruminal Gas Total Production
4.2. Ruminal Methane (CH4) Production
4.3. Ruminal Carbon Monoxide (CO) Production
4.4. Ruminal Hydrogen Sulfide (H2S) Production
4.5. Fermentation Characteristics and CH4 Conversion Efficiency
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tseten, T.; Sanjorjo, R.A.; Kwon, M.; Kim, S.W. Strategies to mitigate enteric methane emissions from ruminant animals. J. Microbiol. Biotechnol. 2022, 32, 269–277. [Google Scholar] [CrossRef]
- Knapp, J.R.; Laur, G.L.; Vadas, P.A.; Weiss, W.P.; Tricarico, J.M. Invited review: Enteric methane in dairy cattle production: Quantifying the opportunities and impact of reducing emissions. J. Dairy Sci. 2014, 97, 3231–3261. [Google Scholar] [CrossRef] [PubMed]
- Pulina, G.; Francesconi, A.H.D.; Stefanon, B.; Sevi, A.; Calamari, L.; Lacetera, N.; Dell’Orto, V.; Pilla, F.; Marsan, P.A.; Mele, M.; et al. Sustainable ruminant production to help feed the planet. Ital. J. Anim. Sci. 2017, 16, 140–171. [Google Scholar] [CrossRef]
- Gerber, P.J.; Hristov, A.N.; Henderson, B.; Makkar, H.; Oh, J.; Lee, C.; Meinen, R.; Montes, F.; Ott, T.; Firkins, J.; et al. Technical options for the mitigation of direct methane and nitrous oxide emissions from livestock: A review. Animals 2013, 7, 220–234. [Google Scholar] [CrossRef]
- Pereira, A.M.; Dapkevicius, M.L.N.E.; Borba, A.E. Alternative pathways for hydrogen sink originated from the ruminal fermentation of carbohydrates: Which microorganisms are involved in lowering methane emission? Anim. Microbiome 2022, 4, 5. [Google Scholar] [CrossRef]
- Liu, W.; Hegglin, M.I.; Checa-Garcia, R.; Li, S.; Gillett, N.P.; Lyu, K.; Zhang, X.; Swart, N.C. Stratospheric ozone depletion and tropospheric ozone increases drive Southern Ocean interior warming. Nat. Clim. Chang. 2022, 12, 365–372. [Google Scholar] [CrossRef]
- Uniyal, S.; Chaudhary, L.C.; Kala, A.; Agarwal, N.; Chaturvedi, V.B. Effect of supplementing sulphate-reducing bacteria along with sulphur on growth performance, nutrient utilization and methane emission in goats. Trop Anim. Health Prod. 2022, 55, 3. [Google Scholar] [CrossRef]
- Drewnoski, M.; Beitz, D.C.; Loy, D.D.; Hansen, S.L.; Ensley, S.M. Factors affecting ruminal hydrogen sulfide concentration of cattle. Anim. Ind. Rep. 2011, 657, 11. [Google Scholar] [CrossRef]
- Rojas-Downing, M.M.; Nejadhashemi, A.P.; Harrigan, T.; Woznicki, S.A. Climate change and livestock: Impacts, adaptation, and mitigation. Clim. Risk Manag. 2017, 16, 145–163. [Google Scholar] [CrossRef]
- Arango, J.; Ruden, A.; Martinez-Baron, D.; Loboguerrero, A.M.; Berndt, A.; Chacón, M.; Torres, C.F.; Oyhantcabal, W.; Gomez, C.A.; Ricci, P.; et al. Ambition meets reality: Achieving GHG emission reduction targets in the livestock sector of Latin America. Front. Sustain. Food Syst. 2020, 4, 65. [Google Scholar] [CrossRef]
- Haque, M.N. Dietary manipulation: A sustainable way to mitigate methane emissions from ruminants. J. Anim. Sci. Technol. 2018, 60, 15. [Google Scholar] [CrossRef]
- Parra, C.S.; Bolson, D.C.; Jacovaci, F.A.; Nussio, L.G.; Jobim, C.C.; Daniel, J.L.P. Influence of soybean-crop proportion on the conservation of maize-soybean bi-crop silage. Anim. Feed. Sci. Tech. 2019, 257, 114295. [Google Scholar] [CrossRef]
- Rosa, L.O.; Pereira, O.G.; Ribeiro, K.G.; Filho, S.C.V.; Cecon, P.R. Chemical composition in soybean silages with inoculant and molasses. Cienc. Anim. Bras. 2020, 21, e-58211. [Google Scholar] [CrossRef]
- Zhao, C.; Wang, L.; Ma, G.; Jiang, X.; Yang, J.; Lv, J.; Zhang, Y. Cellulase interacts with lactic acid bacteria to affect fermentation quality, microbial community, and ruminal degradability in mixed silage of soybean residue and corn stover. Animals 2021, 11, 334. [Google Scholar] [CrossRef]
- Anil, L.; Park, J.; Phipps, R.H. The potential of forage–maize intercrops in ruminant nutrition. Anim. Feed. Sci. Tech. 2020, 86, 157–164. [Google Scholar] [CrossRef]
- Hristov, A.N.; Oh, J.; Firkins, L.; Dijkstra, J.; Kebreab, E.; Waghorn, G.; Makkar, H.P.S.; Adesogan, T.; Yang, W.; Lee, C.; et al. SPECIAL TOPICS-Mitigation of methane and nitrous oxide emissions from animal operations: I. A review of enteric methane mitigation options. J. Anim. Sci. 2013, 91, 5045–5069. [Google Scholar] [CrossRef]
- Lombardi, B.; Alvarado, P.I.; Ricci, P.; Guzmán, S.A.; Gonda, H.L.; Juliarena, M.P. Methane and nitrous oxide emissions from dung patches deposited by grazing cattle supplemented with maize grain. Anim. Feed. Sci. Tech. 2021, 279, 115029. [Google Scholar] [CrossRef]
- Dall-Orsoletta, A.C.; Oziemblowski, M.M.; Alexandre, B.; Ribeiro-Filho, M.M. Enteric methane emission from grazing dairy cows receiving corn silage or ground corn supplementation. Anim. Feed. Sci. Tech. 2019, 253, 65–73. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; Kreuzer, M.; O’mara, F.; McAllister, T.A. Nutritional management for enteric methane abatement: A review. Aust. J. Exp. Agric. 2008, 48, 21–27. [Google Scholar] [CrossRef]
- De Boever, J.L.; Goossens, K.; Peiren, N.; Swanckaert, J.; Ampe, B.; Reheul, D.; Vandaele, L. The effect of maize silage type on the performances and methane emission of dairy cattle. Anim. Physiol. Anim. Nutr. 2017, 101, e246–e256. [Google Scholar] [CrossRef]
- McGrath, J.; Duval, S.M.; Tamassia, L.F.; Kindermann, M.; Stemmler, R.T.; De Gouvea, V.N.; Acedo, T.S.; Immig, I.; Williams, S.N.; Celi, P. Nutritional strategies in ruminants: A lifetime approach. Res. Vet. Sci. 2018, 116, 28–39. [Google Scholar] [CrossRef]
- Cezimbra, I.M.; De Albuquerque-Nunes, P.A.; De Souza-Filho, W.; Tischler, M.R.; Genro, T.C.M.; Bayer, C.; Savian, J.V.; Francois-Bonnet, O.J.; Jean-Francois, S.; De Faccio-Carvalho, P.C. Potential of grazing management to improve beef cattle production and mitigate methane emissions in native grasslands of the Pampa biome. Sci. Total Environ. 2021, 780, 146582. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; Ungerfeld, E.M.; Eckard, R.J.; Wang, M. Fifty years of research on rumen methanogenesis: Lessons learned and future challenges for mitigation. Animal 2020, 14, s2–s16. [Google Scholar] [CrossRef]
- Ardalan, M.; Miesner, M.D.; Reinhardt, C.D.; Thomson, D.U.; Armendariz, C.K.; Titgemeyer, E. Guanidinoacetic acid as a precursor for creatine in steers. J. Anim. Sci. 2016, 94, 766. [Google Scholar] [CrossRef]
- Zhang, J.H.; Li, H.H.; Zhang, G.J.; Zhang, Y.H.; Liu, B.; Huang, S.; Guyader, J.; Zhong, R.Z. Supplementation of guanidinoacetic acid and rumen-protected methionine increased growth performance and meat quality of Tan lambs. Anim. Biosci. 2022, 35, 1556–1565. [Google Scholar] [CrossRef]
- Wyss, M.; Kaddurah-Daouk, R. Creatine and creatinine metabolism. Physiol. Rev. 2000, 80, 1107–1213. [Google Scholar] [CrossRef]
- Ostojic, S.M. Guanidinoacetic acid as a performance-enhancing agent. Amino Acids 2015, 48, 1867–1875. [Google Scholar] [CrossRef] [PubMed]
- Ardalan, M.; Miesner, M.D.; Reinhardt, C.D.; Thomson, D.U.; Armendariz, C.K.; Smith, J.S.; Titgemeyer, E.C. Effects of guanidinoacetic acid supplementation on nitrogen retention and methionine flux in cattle. J. Anim. Sci. 2021, 100, skac283. [Google Scholar] [CrossRef]
- Yan, Z.; Yan, Z.; Liu, S.; Yin, Y.; Yang, T.; Chen, Q. Regulative mechanism of guanidinoacetic acid on skeletal muscle development and its application prospects in animal husbandry: A review. Front. Nutr. 2021, 8, 714567. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP); Bampidis, V.; Azimonti, G.; Bastos, M.D.L.; Christensen, H.; Dusemund, B.; Fasmon-Durjava, M.; Kouba, M.; López-Alonso, M.; López-Puente, S.; et al. Safety and efficacy of a feed additive consisting of guanidinoacetic acid for all animal species (Alzchem Trostberg GmbH). EFSA J. 2022, 20, e07269. [Google Scholar] [CrossRef]
- Li, S.Y.; Wang, C.; Wu, Z.Z.; Liu, Q.; Guo, G.; Huo, W.J.; Zhang, J.; Chen, L.; Zhang, Y.L.; Pei, C.X.; et al. Effects of guanidinoacetic acid supplementation on growth performance, nutrient digestion, rumen fermentation and blood metabolites in Angus bulls. Animal 2020, 14, 2535–2542. [Google Scholar] [CrossRef]
- Li, Z.; Liang, H.; Xin, J.; Xu, L.; Li, M.; Yu, H.; Zhang, W.; Ge, Y.; Li, Y.; Qu, M. Effects of dietary guanidinoacetic acid on the feed efficiency, blood measures, and meat quality of jinjiang bulls. Front. Vet. Sci. 2021, 8, 684295. [Google Scholar] [CrossRef]
- Liu, Y.J.; Chen, J.Z.; Wang, D.H.; Wu, M.J.; Zheng, C.; Wu, Z.Z.; Wang, C.; Lui, Q.; Zhang, J.; Guo, G.; et al. Effects of guanidinoacetic acid and coated folic acid supplementation on growth performance, nutrient digestion and hepatic gene expression in Angus bulls. Br. J. Nutr. 2020, 126, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wang, C.; Zhang, J.; Liu, Q.; Guo, G.; Huo, W.J.; Pei, C.X.; Chen, L.; Zhang, Y.L. Guanidinoacetic acid and betaine supplementation have positive effects on growth performance, nutrient digestion and rumen fermentation in Angus bulls. Anim. Feed Sci. Tech. 2021, 276, 114923. [Google Scholar] [CrossRef]
- Chao, Y.; Liu, B.; Kou, Q.; Niu, W.; Jiang, H.; Yang, L.; Zhang, G. Effects of guanidine acetic acid on growth performance, slaughter performance, fat deposition and nutritional components in muscle of stabling Tan sheep. Chin. J. Clin. Nutr. 2019, 31, 388–394. [Google Scholar] [CrossRef]
- Ren, G.; Hao, X.; Zhang, X.; Liu, S.; Zhang, J. Effects of guanidinoacetic acid and betaine on growth performance, energy and nitrogen metabolism, and rumen microbial protein synthesis in lambs. Anim. Feed. Sci. Tech. 2022, 292, 115402. [Google Scholar] [CrossRef]
- Wen-Juan, L.; Qi-Chao, W.; Zhao-Yang, C.; Yao-Wen, J.; Aisikaer, A.; Zhang, F.; He-Wei, C.; Wei-Kang, W.; Yan-Lu, W.; Liang-Kang, L.; et al. Guanidine acetic acid exhibited greater growth performance in younger (13–30 kg) than in older (30–50 kg) lambs under high-concentrate feedlotting pattern. Front. Vet. Sci. 2022, 1134, 954675. [Google Scholar] [CrossRef]
- Zhang, S.; Zang, C.; Pan, J.; Ma, C.; Wang, C.; Li, X.; Kai, W.; Yang, K. Effects of dietary guanidinoacetic acid on growth performance, guanidinoacetic acid absorption and creatine metabolism of lambs. PLoS ONE 2022, 17, e0264864. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, X.; Song, P.; Zhao, J.; Zhang, J.; Zhao, J. Skeletal muscle mass, meat quality and antioxidant status in growing lambs supplemented with guanidinoacetic acid. Meat Sci. 2022, 192, 108906. [Google Scholar] [CrossRef]
- Liu, B.; Zhong, R.; Kou, Q.F.; Niu, W.Z.; Zhang, G.J. Effects of combination of N-carbamyl glutamate and guanidine acetic acid on carcass characteristic and meat quality of Tan sheep. Chin. J. Ani. Nutr. 2019, 31, 5595–5600. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, J.; Wang, C.; Guo, G.; Huo, W.; Xia, C.; Chen, L.; Zhang, Y.; Pei, C.; Liu, Q. Effects of guanidinoacetic acid supplementation on lactation performance, nutrient digestion and rumen fermentation in Holstein dairy cows. J. Sci. Food Agric. 2022, 103, 1522–1529. [Google Scholar] [CrossRef]
- Vargas, V.; Hernández, M.E.; Gutiérrez, L.J.; Plácido, J.M.; Jiménez, C.A. Clasificación climática del Estado de Tamaulipas, México. Cienc. UAT 2007, 2, 15–19. [Google Scholar]
- Thiex, N.; Novotny, L.; Crawford, A. Determination of ash in animal feed: AOAC official method 942.05 revisited. J. AOAC Int. 2012, 95, 1392–1397. [Google Scholar] [CrossRef] [PubMed]
- Horneck, D.A.; Miller, R.O. Determination of total nitrogen in plant tissue. In Handbook of Reference Methods for Plant Analysis; Karla, Y.P., Ed.; CRC Press: Boca Raton, FL, USA, 1998; pp. 75–83. [Google Scholar]
- Van Soest, P.V.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and non-starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Faichney, G.; White, G. Methods for the Analysis of Feeds Eaten by Ruminants; Division of Animal Production, Ian Clunies Ross Animal Research Laboratory, Commonwealth Scientific and Industrial Research Organization: Melbourne, Australia, 1983. [Google Scholar]
- Padmore, J.M. Animal Feed. In Official Methods of Analysis of the Association of Official Analytical Chemists (AOAC); Helrick, K., Ed.; AOAC: Arlington, VA, USA, 1990; pp. 69–90. [Google Scholar]
- Mertens, D.R. Creating a system for meeting the fiber requirements of dairy cows. J. Dairy Sci. 1997, 8, 1463–1481. [Google Scholar] [CrossRef]
- Sniffen, C.J.; O’connor, J.D.; Van Soest, P.J.; Fox, D.G.; Russell, J.B. A net carbohydrate and protein system for evaluating cattle diets: II. Carbohydrate and protein availability. J. Anim. Sci. 1992, 70, 3562–3577. [Google Scholar] [CrossRef]
- Goering, M.K.; Van Soest, P.J. Forage Fibre Analysis (Apparatus, Reagents, Procedures and Some Applications); Agricultural Research Service USDA: Washington, DC, USA, 1970; pp. 1–24. [Google Scholar] [CrossRef]
- Theodorou, M.K.; Williams, B.A.; Dhanoa, M.S.; McAllan, A.B.; France, J. A simple gas production method using a pressure transducer to determine the fermentation kinetics of ruminant feeds. Anim. Feed. Sci. Tech. 1994, 48, 185–1977. [Google Scholar] [CrossRef]
- Acosta, J.A.D.; Elghandour, M.M.; Mariezcurrena-Berasain, M.D.; Adegbeye, M.J.; Fajemisin, A.N.; Pliego, A.B.; Salem, A.Z. Effect of mid-term dietary administration of the Caesalpinia coriaria extract on the sustainable mitigation of equine fecal methane, carbon monoxide and hydrogen sulfide production. J. Equine Vet. Sci. 2022, 115, 104021. [Google Scholar] [CrossRef] [PubMed]
- Tagliapietra, F.; Cattani, M.; Bailoni, L.; Schiavon, S. In Vitro rumen fermentation: Effect of headspace pressure on the gas production kinetics of corn meal and meadow hay. Anim. Feed. Sci. Tech. 2010, 158, 197–201. [Google Scholar] [CrossRef]
- Elghandour, M.M.; Chagoyán, J.C.V.; Salem, A.Z.; Kholif, A.E.; Castañeda, J.S.M.; Camacho, L.M.; Cerrillo-Soto, M.A. Effects of Saccharomyces cerevisiae at direct addition or pre-incubation on In Vitro gas production kinetics and degradability of four fibrous feeds. Ital. J. Anim. Sci. 2014, 13, 3075. [Google Scholar] [CrossRef]
- SAS. Statistical Analysis System. In User’s Guide: Statistics; Ver 9.0.; SAS Institute: Cary, NC, USA, 2002. [Google Scholar]
- France, J.; Dijkstra, J.; Dhanoa, M.S.; López, S.; Bannink, A. Estimating the extent of degradation of ruminant feeds from a description of their gas production profiles observed In Vitro: Derivation of models and other mathematical considerations. Br. J. Nutr. 2000, 83, 143–150. [Google Scholar] [CrossRef]
- Menke, K.H.; Raab, L.; Salewski, A.; Steingass, H.; Fritz, D.; Schneider, W. The estimation of the digestibility and metabolizable energy content of ruminant feedingstuffs from the gas production when they are incubated with rumen liquor In Vitro. J. Agric. Sci. 1979, 93, 217–222. [Google Scholar] [CrossRef]
- Getachew, G.; Makkar, H.P.S.; Becker, K. Tropical browses: Contents of phenolic compounds, In Vitro gas production and stoichiometric relationship between short chain fatty acid and In Vitro gas production. J. Agric. Sci. 2002, 139, 341–352. [Google Scholar] [CrossRef]
- Elghandour, M.M.Y.; Kholif, A.E.; Márquez-Molina, O.; Vázquez-Armijo, J.F.; Puniya, A.K.; Salem, A.Z.M. Influence of individual or mixed cellulase and xylanase mixture on In Vitro rumen gas production kinetics of total mixed rations with different maize silage and concentrate ratios. Turk. J. Vet. Anim. Sci. 2015, 39, 435–442. [Google Scholar] [CrossRef]
- Kholif, A.E.; Gouda, G.A.; Morsy, T.A.; Matloup, O.H.; Fahmy, M.; Gomaa, A.S.; Patra, A.K. Dietary date palm leaves ensiled with fibrolytic enzymes decreased methane production, and improved feed degradability and fermentation kinetics in a ruminal In Vitro system. Waste Biomass Valorization 2022, 13, 3475–3488. [Google Scholar] [CrossRef]
- Huang, Y.; Linag, L.; Dai, S.; Wu, C.; Chen, C.; Hao, J. Effect of different regions and ensiling periods on fermentation quality and bacteria community of whole-plant maize silage. Front. Microbiol. 2022, 12, 743695. [Google Scholar] [CrossRef]
- Giaveno, C.; Ferrero, J. Introduction of tropical maize genotypes to increase silage production in the central area of Santa Fe, Argentina. Crop. Breed. Appl. Biotechnol. 2003, 3, 89–94. [Google Scholar] [CrossRef]
- Ardalan, M.; Batista, E.D.; Titgemeyer, E.C. Effect of post-ruminal guanidinoacetic acid supplementation on creatine synthesis and plasma homocysteine concentrations in cattle. J. Anim. Sci. 2020, 98, skaa072. [Google Scholar] [CrossRef]
- Speer, H.F.; Pearl, K.A.; Titgemeyer, E.C. Relative bioavailability of guanidinoacetic acid delivered ruminally or abomasally to cattle. J. Anim. Sci. 2020, 98, skaa282. [Google Scholar] [CrossRef]
- Wojcieszak, M.; Pyzik, A.; Poszytek, K.; Krawczyk, P.S.; Sobczak, A.; Lipinski, L.; Roubinek, O.; Palige, J.; Sklodowska, A.; Drewniak, L. Adaptation of methanogenic inocula to anaerobic digestion of maize silage. Front. Microbiol. 2017, 8, 1881. [Google Scholar] [CrossRef]
- Zhu, M.; Xie, Y.; Li, S.; Chen, X.; Zhao, Z. Technical Measures for improving quality of silage maize in Southern Area. Anim. Husb. Feed Sci. 2010, 31, 31–34. [Google Scholar]
- Hu, W.; Schmidt, R.; Mcdonell, E.; Klingerman, C.M.; Niu, H. Diversity of bacteria community during ensiling and subsequent exposure to air in whole plant maize silage. Asian-Australas J. Anim. Sci. 2018, 31, 1464–1473. [Google Scholar] [CrossRef]
- Adjei-Fremah, S.; Ekwemalor, K.; Worku, M.; Ibrahim, S. Probiotics and Ruminant Health; IntechOpen: London, UK, 2018; Volume 8, pp. 133–150. [Google Scholar] [CrossRef]
- Takahashi, J. Lactic Acid Bacteria and Mitigation of GHG Emission from Ruminant Livestock; IntechOpen: London, UK, 2013; Volume 19, pp. 451–466. [Google Scholar] [CrossRef]
- Elghandour, M.M.M.Y.; Adegbeye, M.J.; Barbabosa-Pilego, A.; Perez, N.R.; Hernández, S.R.; Zaragoza-Bastida, A.; Salem, A.Z.M. Equine contribution in methane emission and its mitigation strategies. J. Equine Vet. Sci. 2019, 72, 56–63. [Google Scholar] [CrossRef]
- Zhao, Y.J.; Wexler, A.S.; Hase, F.; Pan, Y.; Mitloehner, F.M. Carbon Monoxide Emissions from Corn Silage. J. Environ. Prot. Sci. 2021, 12, 438–453. [Google Scholar] [CrossRef]
- Techtmann, S.M.; Colman, A.S.; Robb, F.T. ‘That which does not kill us only makes us stronger’: The role of carbon monoxide in thermophilic microbial consortia. Environ. Microbiol. 2009, 11, 1027–1037. [Google Scholar] [CrossRef]
- Drake, H.L.; Gößner, A.S.; Daniel, S.L. Old acetogens, new light. Ann. N. Y. Acad. Sci. 2008, 1125, 100–128. [Google Scholar] [CrossRef]
- Ragsdale, S.W. Enzymology of the Wood–Ljungdahl pathway of acetogenesis. Ann. N. Y. Acad. Sci. 2008, 1125, 129–136. [Google Scholar] [CrossRef]
- Kang, S.; Song, Y.; Jin, S.; Shin, J.; Bae, J.; Kim, D.R.; Lee, J.K.; Kim, S.C.; Cho, S.; Cho, B.K. Adaptive laboratory evolution of Eubacterium limosum ATCC 8486 on carbon monoxide. Front. Microbiol. 2020, 11, 402. [Google Scholar] [CrossRef]
- Wu, H.; Li, Y.; Meng, Q.; Zhou, Z. Effect of high sulfur diet on rumen fermentation, microflora, and epithelial barrier function in steers. Animals 2021, 11, 2545. [Google Scholar] [CrossRef]
- Castro, D.J.; Cerón-Cucchi, M.E.; Ortiz-Chura, A.; Depetris, G.J.; Irazoqui, J.M.; Amadio, A.F.; Cantón, G.J. Ruminal effects of excessive dietary sulphur in feedlot cattle. J. Anim. Physiol. Anim. Nutr. 2021, 106, 978–987. [Google Scholar] [CrossRef]
- Smith, D.R.; Dilorenzo, N.; Leibovich, J.; May, M.L.; Quinn, M.J.; Homm, J.W.; Galyean, M.L. Effects of sulfur and monensin concentrations on In Vitro dry matter disappearance, hydrogen sulfide production, and volatile fatty acid concentrations in batch culture ruminal fermentations. J. Anim. Sci. 2010, 88, 1503. [Google Scholar] [CrossRef]
- Mould, F.L.; Ørskov, E.R.; Mann, S.O. Associative effects of mixed feeds. I. Effects of type and level of supplementation and the influence of the rumen fluid pH on cellulolysis in vivo and dry matter digestion of various roughages. Anim. Feed. Sci. Tech. 1993, 10, 15–30. [Google Scholar] [CrossRef]
- Lyle, R.R.; Johnson, R.R.; Wilhite, J.V. Rumen characteristics in steers as affected by adaptation from forage to all concentrate diets. J. Anim. Sci. 1981, 53, 1383–1390. [Google Scholar] [CrossRef]
- Wang, S.; Giller, K.; Kreuzer, M.; Ulbrich, S.E.; Braun, U.; Schwarm, A. Contribution of ruminal fungi, archaea, protozoa, and bacteria to the methane suppression caused by oilseed supplemented diets. Front. Microbiol. 2017, 8, 1864. [Google Scholar] [CrossRef]
- Faniyi, T.O.; Adewumi, M.K.; Jack, M.K.; Adegbeye, M.J.; Elghandour, M.; Barbabosa-Pliego, A.; Salem, A.Z.M. Extracts of herbs and spices as feed additives mitigate ruminal methane production and improve fermentation characteristics in West African Dwarf sheep. Trop. Anim. Health Prod. 2021, 53, 312. [Google Scholar] [CrossRef]
- Rodriguez, G.B.; Carmona, D.A.; Elghandour, M.M.Y.; Salem, A.Z.M.; Soto, H.R.; Sanchez, R.R.; Adegbeye, M.J. Sustainable use of sodium butyrate as a source of bioactive additive: Impact on calf growth performance, rumen fermentation characteristics and microbial count. Biomass Conv. Bioref. 2022. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Maize Genotype and State of the Forage 2 | Component 1 | |||||||
|---|---|---|---|---|---|---|---|---|
| OM | CP | EE | NDF | ADF | ADL | NSC | TC | |
| Amarillo | ||||||||
| Fresh | 92.06 | 10.80 | 2.39 | 59.71 | 31.68 | 3.84 | 19.16 | 78.87 |
| Ensiled | 92.76 | 8.31 | 3.60 | 47.56 | 26.23 | 3.89 | 33.29 | 80.85 |
| Montesa | ||||||||
| Fresh | 92.72 | 10.54 | 2.59 | 60.63 | 30.27 | 3.67 | 18.96 | 79.59 |
| Ensiled | 93.26 | 8.29 | 3.90 | 50.35 | 26.90 | 4.12 | 30.72 | 81.07 |
| Olotillo | ||||||||
| Fresh | 92.76 | 10.29 | 2.64 | 66.23 | 36.37 | 4.41 | 13.60 | 79.83 |
| Ensiled | 92.99 | 8.35 | 3.80 | 59.82 | 36.08 | 4.89 | 21.02 | 80.84 |
| Tampiqueño | ||||||||
| Fresh | 92.12 | 10.49 | 2.19 | 61.66 | 35.05 | 4.25 | 17.78 | 79.45 |
| Ensiled | 93.62 | 8.56 | 3.40 | 59.46 | 32.31 | 4.86 | 22.19 | 81.66 |
| Tuxpeño | ||||||||
| Fresh | 92.08 | 10.26 | 2.45 | 58.86 | 30.52 | 3.70 | 20.50 | 79.36 |
| Ensiled | 91.63 | 9.58 | 3.60 | 51.49 | 28.88 | 4.21 | 26.95 | 78.45 |
| Maize Genotype | State of the Forage (SF) | Guanidinoacetic Acid (GAA) | Total Gas Production | |||||
|---|---|---|---|---|---|---|---|---|
| Kinetics Parameters 1 | mL gas g−1 DM Incubated | |||||||
| b | c | Lag | 4 h | 24 h | 48 h | |||
| Amarillo | Fresh | − | 234.85 | 0.0245 | 5.85 | 68.04 | 160.52 | 219.71 |
| + | 277.20 | 0.0170 | 14.37 | 85.72 | 168.94 | 227.60 | ||
| Ensiled | − | 307.45 | 0.0310 | 2.30 | 76.40 | 228.45 | 295.09 | |
| + | 414.90 | 0.0230 | 2.63 | 86.04 | 241.63 | 368.05 | ||
| SEM 2 | 29.32 | 0.0021 | 1.93 | 7.22 | 16.89 | 23.86 | ||
| SF | 0.0230 | 0.0396 | 0.0166 | 0.5800 | 0.0141 | 0.0106 | ||
| GAA | 0.0630 | 0.0203 | 0.0834 | 0.1313 | 0.5575 | 0.1655 | ||
| SF × GAA | 0.3292 | 0.9100 | 0.1007 | 0.6068 | 0.8947 | 0.2445 | ||
| Montesa | Fresh | − | 230.90 | 0.0255 | 9.71 | 82.65 | 185.99 | 218.70 |
| + | 230.35 | 0.0225 | 8.61 | 69.31 | 161.27 | 210.39 | ||
| Ensiled | − | 327.70 | 0.0260 | 6.86 | 96.63 | 249.36 | 306.00 | |
| + | 329.75 | 0.0200 | 10.33 | 96.63 | 211.29 | 290.70 | ||
| SEM 2 | 30.35 | 0.0020 | 2.20 | 3.40 | 13.70 | 24.86 | ||
| SF | 0.0319 | 0.6483 | 0.8094 | 0.0037 | 0.0144 | 0.0280 | ||
| GAA | 0.9815 | 0.0911 | 0.6182 | 0.1216 | 0.0838 | 0.6596 | ||
| SF × GAA | 0.9679 | 0.5012 | 0.3577 | 0.1216 | 0.6518 | 0.8949 | ||
| Olotillo | Fresh | − | 226.60 | 0.0210 | 14.79 | 90.91 | 170.16 | 209.33 |
| + | 189.60 | 0.0150 | 20.98 | 82.76 | 121.45 | 165.92 | ||
| Ensiled | − | 257.95 | 0.0195 | 10.62 | 76.93 | 164.44 | 225.64 | |
| + | 236.70 | 0.0285 | 7.23 | 82.23 | 190.59 | 226.28 | ||
| SEM 2 | 9.51 | 0.0013 | 1.34 | 2.09 | 8.73 | 10.65 | ||
| SF | 0.0146 | 0.0093 | 0.0026 | 0.0254 | 0.0221 | 0.0228 | ||
| GAA | 0.0376 | 0.3046 | 0.3549 | 0.5313 | 0.2661 | 0.1151 | ||
| SF × GAA | 0.4542 | 0.0042 | 0.0232 | 0.0322 | 0.0128 | 0.1075 | ||
| Tampiqueño | Fresh | − | 261.10 | 0.0250 | 6.34 | 73.12 | 184.56 | 243.74 |
| + | 278.10 | 0.0220 | 4.77 | 63.38 | 167.62 | 243.43 | ||
| Ensiled | − | 253.70 | 0.0235 | 12.19 | 97.37 | 200.44 | 239.14 | |
| + | 214.40 | 0.0230 | 11.50 | 77.57 | 163.97 | 199.12 | ||
| SEM 2 | 21.56 | 0.0017 | 2.35 | 3.02 | 14.58 | 20.96 | ||
| SF | 0.1745 | 0.8887 | 0.0555 | 0.0031 | 0.6964 | 0.3081 | ||
| GAA | 0.6323 | 0.3556 | 0.6561 | 0.0081 | 0.1409 | 0.3905 | ||
| SF × GAA | 0.2616 | 0.4975 | 0.8595 | 0.1708 | 0.5396 | 0.3972 | ||
| Tuxpeño | Fresh | − | 307.40 | 0.0270 | 5.05 | 83.50 | 229.71 | 290.59 |
| + | 281.10 | 0.0245 | 5.57 | 75.98 | 191.54 | 259.63 | ||
| Ensiled | − | 263.45 | 0.0225 | 9.98 | 85.72 | 196.47 | 243.16 | |
| + | 228.20 | 0.0230 | 11.42 | 91.86 | 167.09 | 216.11 | ||
| SEM 2 | 14.71 | 0.0008 | 0.60 | 7.08 | 11.28 | 14.28 | ||
| SF | 0.0302 | 0.0192 | 0.0008 | 0.2704 | 0.0628 | 0.0334 | ||
| GAA | 0.1047 | 0.2746 | 0.1769 | 0.9271 | 0.0402 | 0.1121 | ||
| SF × GAA | 0.7762 | 0.1306 | 0.4848 | 0.3897 | 0.7167 | 0.8976 | ||
| Pooled SEM 2 | 22.60 | 0.0016 | 1.80 | 5.05 | 13.34 | 19.72 | ||
| p value | ||||||||
| Maize genotype | 0.0009 | 0.0828 | 0.0003 | 0.1336 | 0.0015 | 0.0008 | ||
| State of the forage | 0.0052 | 0.0416 | 0.1893 | 0.0006 | 0.0002 | 0.0017 | ||
| Guanidinoacetic acid | 0.9284 | 0.0015 | 0.1041 | 0.3907 | 0.0057 | 0.3528 | ||
| Maize genotype × State of the forage | 0.0001 | 0.0014 | <0.0001 | 0.0051 | 0.0003 | <0.0001 | ||
| Maize genotype × Guanidinoacetic acid | 0.0210 | 0.0081 | 0.3362 | 0.0120 | 0.1436 | 0.1317 | ||
| State of the forage × Guanidinoacetic acid | 0.8589 | 0.0315 | 0.1729 | 0.3343 | 0.3629 | 0.4607 | ||
| Maize genotype × State of the forage × Guanidinoacetic acid | 0.4613 | 0.0099 | 0.0436 | 0.2396 | 0.1358 | 0.3773 | ||
| Maize Genotype | State of the Forage (SF) | Guanidinoacetic Acid (GAA) | CH4 Production | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Kinetics Parameters 1 | mL gas g−1 DM Incubated | mL gas 100 mL−1 Gas | |||||||||
| b | c | Lag | 4 h | 24 h | 48 h | 4 h | 24 h | 48 h | |||
| Amarillo | Fresh | − | 38.27 | 0.1255 | 12.52 | 0.3400 | 8.97 | 38.13 | 0.5000 | 5.50 | 17.25 |
| + | 27.52 | 0.0650 | 9.86 | 0.3315 | 4.36 | 36.85 | 0.3750 | 2.56 | 16.19 | ||
| Ensiled | − | 58.92 | 0.1170 | 11.43 | 0.3820 | 19.18 | 58.55 | 0.5000 | 8.50 | 20.00 | |
| + | 78.71 | 0.0960 | 12.72 | 0.4300 | 12.44 | 78.10 | 0.5000 | 5.31 | 21.44 | ||
| SEM 2 | 5.20 | 0.0276 | 1.77 | 0.0749 | 2.89 | 3.60 | 0.0625 | 1.62 | 1.79 | ||
| SF | 0.0023 | 0.7044 | 0.6420 | 0.4016 | 0.0338 | 0.0010 | 0.3739 | 0.1505 | 0.0890 | ||
| GAA | 0.4338 | 0.2138 | 0.7192 | 0.8051 | 0.1201 | 0.0641 | 0.3739 | 0.1316 | 0.9216 | ||
| SF × GAA | 0.0426 | 0.5137 | 0.3262 | 0.7253 | 0.7302 | 0.0444 | 0.3739 | 0.9422 | 0.5233 | ||
| Montesa | Fresh | − | 42.97 | 0.1225 | 11.81 | 0.4135 | 12.46 | 42.86 | 0.5000 | 6.63 | 19.50 |
| + | 22.70 | 0.0480 | 4.87 | 0.2540 | 9.65 | 21.70 | 0.3750 | 6.06 | 10.35 | ||
| Ensiled | − | 36.45 | 0.1330 | 12.24 | 0.4830 | 7.17 | 36.29 | 0.5000 | 2.88 | 11.63 | |
| + | 34.94 | 0.0880 | 11.98 | 0.2415 | 6.51 | 34.69 | 0.2500 | 3.06 | 11.44 | ||
| SEM 2 | 10.81 | 0.0222 | 1.25 | 0.0365 | 1.79 | 10.78 | 0.0625 | 0.93 | 2.82 | ||
| SF | 0.8045 | 0.3191 | 0.0396 | 0.4787 | 0.0778 | 0.7803 | 0.3739 | 0.0220 | 0.2953 | ||
| GAA | 0.3707 | 0.0546 | 0.0453 | 0.0054 | 0.3861 | 0.3506 | 0.0399 | 0.8498 | 0.1733 | ||
| SF × GAA | 0.4345 | 0.5430 | 0.0560 | 0.3244 | 0.5803 | 0.4157 | 0.3739 | 0.7068 | 0.1874 | ||
| Olotillo | Fresh | − | 21.71 | 0.0760 | 10.70 | 0.2270 | 4.47 | 19.86 | 0.2500 | 2.63 | 9.50 |
| + | 25.89 | 0.1185 | 12.37 | 0.4140 | 5.56 | 25.89 | 0.5000 | 4.56 | 15.56 | ||
| Ensiled | − | 58.90 | 0.1565 | 13.79 | 0.3850 | 8.40 | 58.88 | 0.5000 | 4.94 | 25.56 | |
| + | 64.43 | 0.1280 | 11.52 | 0.4110 | 21.67 | 64.09 | 0.5000 | 11.31 | 28.19 | ||
| SEM 2 | 11.37 | 0.0203 | 1.01 | 0.0102 | 3.08 | 11.40 | 0.0000 | 1.58 | 4.03 | ||
| SF | 0.0291 | 0.0910 | 0.3261 | 0.0016 | 0.0313 | 0.0276 | <0.0001 | 0.0453 | 0.0235 | ||
| GAA | 0.6915 | 0.7477 | 0.7798 | 0.0005 | 0.0800 | 0.6476 | <0.0001 | 0.0579 | 0.3413 | ||
| SF × GAA | 0.9555 | 0.1553 | 0.1216 | 0.0014 | 0.1192 | 0.9734 | <0.0001 | 0.2322 | 0.6914 | ||
| Tampiqueño | Fresh | − | 64.20 | 0.1090 | 12.29 | 0.3655 | 18.38 | 63.78 | 0.5000 | 9.25 | 24.63 |
| + | 43.67 | 0.0955 | 11.80 | 0.1585 | 9.37 | 43.30 | 0.2500 | 5.50 | 17.75 | ||
| Ensiled | − | 33.17 | 0.1085 | 10.91 | 0.4870 | 6.92 | 32.84 | 0.5000 | 3.44 | 13.31 | |
| + | 27.72 | 0.1485 | 13.34 | 0.1940 | 4.41 | 27.75 | 0.2500 | 2.69 | 13.94 | ||
| SEM 2 | 18.90 | 0.0278 | 1.73 | 0.0137 | 6.82 | 18.78 | 0.0000 | 3.19 | 5.82 | ||
| SF | 0.2817 | 0.3979 | 0.9644 | 0.0046 | 0.2953 | 0.2835 | <0.0001 | 0.2478 | 0.2639 | ||
| GAA | 0.5296 | 0.6581 | 0.6047 | <0.0001 | 0.4462 | 0.5334 | <0.0001 | 0.5195 | 0.6200 | ||
| SF × GAA | 0.7103 | 0.3898 | 0.4482 | 0.0347 | 0.6587 | 0.7029 | <0.0001 | 0.6627 | 0.5547 | ||
| Tuxpeño | Fresh | − | 60.04 | 0.0845 | 12.02 | 0.4175 | 15.65 | 59.33 | 0.5000 | 7.00 | 20.75 |
| + | 64.88 | 0.1665 | 14.25 | 0.1900 | 5.44 | 64.97 | 0.2500 | 2.87 | 24.62 | ||
| Ensiled | − | 29.10 | 0.1860 | 13.75 | 0.4285 | 5.96 | 28.66 | 0.5000 | 3.06 | 11.81 | |
| + | 39.27 | 0.1210 | 11.39 | 0.4590 | 12.89 | 39.14 | 0.5000 | 7.75 | 18.12 | ||
| SEM 2 | 16.99 | 0.0085 | 0.16 | 0.0277 | 4.91 | 17.05 | 0.0000 | 2.34 | 5.83 | ||
| SF | 0.1714 | 0.0303 | 0.0223 | 0.0072 | 0.8306 | 0.1728 | <0.0001 | 0.8512 | 0.2563 | ||
| GAA | 0.6815 | 0.3750 | 0.6958 | 0.0237 | 0.7547 | 0.6612 | <0.0001 | 0.9103 | 0.4318 | ||
| SF × GAA | 0.8828 | 0.0010 | 0.0001 | 0.0096 | 0.1558 | 0.8940 | <0.0001 | 0.1333 | 0.8447 | ||
| Pooled SEM 2 | 13.56 | 0.0224 | 1.32 | 0.0400 | 4.28 | 13.43 | 0.0395 | 2.08 | 4.37 | ||
| p value | |||||||||||
| Maize genotype | 0.4842 | 0.1032 | 0.1094 | 0.1104 | 0.9620 | 0.3673 | 0.0330 | 0.9465 | 0.2759 | ||
| State of the forage | 0.4214 | 0.0135 | 0.0882 | 0.0003 | 0.5637 | 0.4893 | 0.0104 | 0.9683 | 0.9733 | ||
| Guanidinoacetic acid | 0.8197 | 0.1676 | 0.2278 | 0.0001 | 0.4347 | 0.9645 | <0.0001 | 0.8217 | 0.8531 | ||
| Maize genotype × State of the forage | 0.0041 | 0.8829 | 0.2206 | 0.4349 | 0.0228 | 0.0052 | 0.0119 | 0.0280 | 0.0086 | ||
| Maize genotype × Guanidinoacetic acid | 0.7263 | 0.1011 | 0.1933 | <0.0001 | 0.2354 | 0.6304 | <0.0001 | 0.1635 | 0.4339 | ||
| State of the forage × Guanidinoacetic acid | 0.2549 | 0.3519 | 0.4050 | 0.9362 | 0.0760 | 0.3314 | 1.0000 | 0.0869 | 0.3685 | ||
| Maize genotype × State of the forage × Guanidinoacetic acid | 0.9455 | 0.0196 | 0.0233 | 0.0115 | 0.5353 | 0.9716 | 0.0020 | 0.5793 | 0.8646 | ||
| Maize Genotype | State of the Forage (SF) | Guanidinoacetic Acid (GAA) | CO production | |||||
|---|---|---|---|---|---|---|---|---|
| Kinetics Parameters 1 | mL gas g−1 DM Incubated | |||||||
| b | c | Lag | 4 h | 24 h | 48 h | |||
| Amarillo | Fresh | − | 6.1784 | 0.0272 | 4.90 | 0.0008 | 0.2536 | 0.7511 |
| + | 3.9989 | 0.0003 | 2.90 | 0.0019 | 0.0817 | 0.6090 | ||
| Ensiled | − | 3.1119 | 0.0008 | 7.55 | 0.0019 | 0.5838 | 1.4391 | |
| + | 3.1728 | 0.0020 | 5.81 | 0.0030 | 0.2685 | 1.7249 | ||
| SEM 2 | 1.3634 | 0.0047 | 2.15 | 0.0002 | 0.0316 | 0.1129 | ||
| SF | 0.2266 | 0.0570 | 0.2662 | 0.0085 | 0.0012 | 0.0013 | ||
| GAA | 0.4806 | 0.0508 | 0.4337 | 0.0100 | 0.0015 | 0.5592 | ||
| SF × GAA | 0.4574 | 0.0393 | 0.9551 | 0.9198 | 0.0855 | 0.1311 | ||
| Montesa | Fresh | − | 6.3918 | 0.0003 | 4.20 | 0.0010 | 0.1599 | 0.6604 |
| + | 2.2404 | 0.0003 | 5.10 | 0.0019 | 0.1214 | 0.5415 | ||
| Ensiled | − | 0.6923 | 0.0171 | 4.31 | 0.0021 | 0.1026 | 0.7018 | |
| + | 0.9613 | 0.0109 | 6.25 | 0.0030 | 0.0928 | 0.9027 | ||
| SEM 2 | 1.1929 | 0.0100 | 2.35 | 0.0001 | 0.0387 | 0.3252 | ||
| SF | 0.0430 | 0.2416 | 0.8012 | 0.0002 | 0.3297 | 0.5694 | ||
| GAA | 0.1790 | 0.7695 | 0.5781 | 0.0003 | 0.5668 | 0.9056 | ||
| SF × GAA | 0.1376 | 0.7722 | 0.8359 | 0.5614 | 0.7298 | 0.6487 | ||
| Olotillo | Fresh | − | 1.4100 | 0.0004 | 5.07 | 0.0021 | 0.0785 | 0.2683 |
| + | 0.2768 | 0.0446 | 7.95 | 0.0022 | 0.1266 | 0.5658 | ||
| Ensiled | − | 1.8519 | 0.0008 | 2.40 | 0.0019 | 0.1801 | 1.1317 | |
| + | 4.3147 | 0.0151 | 7.28 | 0.0028 | 0.4182 | 1.4497 | ||
| SEM 2 | 1.9449 | 0.0228 | 0.99 | 0.0003 | 0.0493 | 0.0703 | ||
| SF | 0.3136 | 0.5590 | 0.1679 | 0.6041 | 0.0163 | 0.0002 | ||
| GAA | 0.7497 | 0.2699 | 0.0174 | 0.1343 | 0.0439 | 0.0119 | ||
| SF × GAA | 0.4076 | 0.5477 | 0.3717 | 0.2084 | 0.1260 | 0.8917 | ||
| Tampiqueño | Fresh | − | 2.5560 | 0.0009 | 2.74 | 0.0011 | 0.2323 | 1.0656 |
| + | 0.9572 | 0.0004 | 5.90 | 0.0015 | 0.2012 | 1.1168 | ||
| Ensiled | − | 1.0592 | 0.0034 | 7.11 | 0.0030 | 0.0990 | 0.6514 | |
| + | 1.5083 | 0.0010 | 6.14 | 0.0023 | 0.1915 | 0.6809 | ||
| SEM 2 | 0.6003 | 0.0014 | 1.04 | 0.0003 | 0.0787 | 0.2833 | ||
| SF | 0.4750 | 0.3266 | 0.0902 | 0.0132 | 0.4149 | 0.2079 | ||
| GAA | 0.3925 | 0.3473 | 0.3490 | 0.6728 | 0.7167 | 0.8937 | ||
| SF × GAA | 0.1633 | 0.5122 | 0.1177 | 0.1707 | 0.4761 | 0.9712 | ||
| Tuxpeño | Fresh | − | 2.3052 | 0.0052 | 5.75 | 0.0018 | 0.2971 | 1.0811 |
| + | 3.1465 | 0.0037 | 6.15 | 0.0021 | 0.3330 | 1.6444 | ||
| Ensiled | − | 3.3266 | 0.0001 | 6.55 | 0.0027 | 0.1452 | 0.4791 | |
| + | 5.0119 | 0.0171 | 4.20 | 0.0027 | 0.2238 | 0.7056 | ||
| SEM 2 | 1.8381 | 0.0024 | 0.97 | 0.0003 | 0.1306 | 0.3833 | ||
| SF | 0.4762 | 0.1614 | 0.5864 | 0.0870 | 0.3742 | 0.1148 | ||
| GAA | 0.5297 | 0.0319 | 0.3731 | 0.6540 | 0.6839 | 0.3610 | ||
| SF × GAA | 0.8297 | 0.0185 | 0.2305 | 0.6540 | 0.8779 | 0.6830 | ||
| Pooled SEM 2 | 1.4699 | 0.0114 | 1.62 | 0.0003 | 0.0751 | 0.2648 | ||
| p value | ||||||||
| Maize genotype | 0.1204 | 0.5711 | 0.9685 | 0.1154 | 0.0330 | 0.2576 | ||
| State of the forage | 0.5062 | 0.7723 | 0.3500 | <0.0001 | 0.2251 | 0.2018 | ||
| Guanidinoacetic acid | 0.6217 | 0.4514 | 0.3390 | 0.0004 | 0.8290 | 0.1638 | ||
| Maize genotype × State of the forage | 0.0687 | 0.3949 | 0.2884 | 0.0254 | 0.0043 | 0.0005 | ||
| Maize genotype × Guanidinoacetic acid | 0.5531 | 0.1415 | 0.1461 | 0.0179 | 0.0198 | 0.8052 | ||
| State of the forage × Guanidinoacetic acid | 0.0592 | 0.8699 | 0.6269 | 0.5256 | 0.4799 | 0.7331 | ||
| Maize genotype × State of the forage × Guanidinoacetic acid | 0.9205 | 0.4370 | 0.6375 | 0.1880 | 0.6021 | 0.8569 | ||
| Maize Genotype | State of the Forage (SF) | Guanidinoacetic Acid (GAA) | H2S Production | |||||
|---|---|---|---|---|---|---|---|---|
| Kinetics Parameters 1 | mL gas g−1 DM Incubated | |||||||
| b | c | Lag | 4 h | 24 h | 48 h | |||
| Amarillo | Fresh | − | 0.0236 | 0.0002 | 5.90 | 0.0001 | 0.0058 | 0.0458 |
| + | 0.0041 | 1.6232 | 6.04 | 0.0001 | 0.0011 | 0.0068 | ||
| Ensiled | − | 0.0590 | 0.0015 | 3.80 | 0.0001 | 0.0093 | 0.0768 | |
| + | 0.0057 | 0.1531 | 5.22 | 0.0001 | 0.0011 | 0.0157 | ||
| SEM 2 | 0.0060 | 0.7424 | 1.84 | 0.0000 | 0.0002 | 0.0035 | ||
| SF | 0.0361 | 0.3785 | 0.4698 | <0.0001 | 0.0008 | 0.0049 | ||
| GAA | 0.0037 | 0.2980 | 0.6930 | <0.0001 | <0.0001 | 0.0001 | ||
| SF × GAA | 0.0475 | 0.3778 | 0.7464 | <0.0001 | 0.0008 | 0.0358 | ||
| Montesa | Fresh | − | 0.4036 | 0.0027 | 3.40 | 0.0000 | 0.0007 | 0.0125 |
| + | 0.0682 | 0.0195 | 2.43 | 0.0001 | 0.0005 | 0.0052 | ||
| Ensiled | − | 0.0030 | 0.1610 | 5.34 | 0.0000 | 0.0005 | 0.0057 | |
| + | 0.0319 | 0.0099 | 4.43 | 0.0001 | 0.0006 | 0.0047 | ||
| SEM 2 | 0.1507 | 0.0224 | 1.31 | 0.0000 | 0.0001 | 0.0005 | ||
| SF | 0.2208 | 0.0295 | 0.2057 | <0.0001 | 0.6213 | 0.0022 | ||
| GAA | 0.3667 | 0.0401 | 0.5115 | <0.0001 | 0.6213 | 0.0013 | ||
| SF × GAA | 0.2932 | 0.0200 | 0.9843 | <0.0001 | 0.3453 | 0.0038 | ||
| Olotillo | Fresh | − | 0.0049 | 0.1863 | 3.59 | 0.0000 | 0.0017 | 0.0062 |
| + | 0.0171 | 0.0001 | 1.35 | 0.0001 | 0.0029 | 0.0356 | ||
| Ensiled | − | 0.0214 | 0.3001 | 3.93 | 0.0001 | 0.0011 | 0.0060 | |
| + | 0.0239 | 0.0006 | 9.65 | 0.0003 | 0.0027 | 0.0482 | ||
| SEM 2 | 0.0120 | 0.1502 | 0.53 | 0.0000 | 0.0003 | 0.0015 | ||
| SF | 0.3873 | 0.7228 | 0.0012 | 0.0075 | 0.2931 | 0.0128 | ||
| GAA | 0.5759 | 0.1812 | 0.0300 | 0.0075 | 0.0084 | <0.0001 | ||
| SF × GAA | 0.7092 | 0.7251 | 0.0017 | 0.3739 | 0.5275 | 0.0115 | ||
| Tampiqueño | Fresh | − | 0.0255 | 0.0009 | 5.53 | 0.0002 | 0.0010 | 0.0072 |
| + | 0.0608 | 0.0862 | 4.39 | 0.0000 | 0.0004 | 0.0085 | ||
| Ensiled | − | 0.0391 | 0.0044 | 6.88 | 0.0000 | 0.0002 | 0.0045 | |
| + | 0.0461 | 0.0086 | 6.12 | 0.0001 | 0.0002 | 0.0041 | ||
| SEM 2 | 0.0296 | 0.0417 | 3.26 | 0.0001 | 0.0004 | 0.0009 | ||
| SF | 0.9848 | 0.4242 | 0.6605 | 0.7556 | 0.3046 | 0.0171 | ||
| GAA | 0.5148 | 0.3434 | 0.7856 | 0.7556 | 0.4766 | 0.6489 | ||
| SF × GAA | 0.6579 | 0.3857 | 0.9565 | 0.1709 | 0.4766 | 0.4059 | ||
| Tuxpeño | Fresh | − | 0.0144 | 0.0725 | 1.65 | 0.0000 | 0.0003 | 0.0072 |
| + | 0.0318 | 0.2500 | 2.64 | 0.0001 | 0.0005 | 0.0221 | ||
| Ensiled | − | 0.1363 | 0.0067 | 4.91 | 0.0001 | 0.0008 | 0.0100 | |
| + | 0.0258 | 0.0070 | 8.25 | 0.0001 | 0.0081 | 0.0570 | ||
| SEM 2 | 0.0267 | 0.1271 | 1.82 | 0.0000 | 0.0029 | 0.0055 | ||
| SF | 0.0958 | 0.2913 | 0.0714 | 0.3739 | 0.2343 | 0.0264 | ||
| GAA | 0.1565 | 0.5228 | 0.3000 | 0.0399 | 0.2650 | 0.0048 | ||
| SF × GAA | 0.0748 | 0.5242 | 0.5525 | 0.3739 | 0.2818 | 0.0430 | ||
| Pooled SEM 2 | 0.0700 | 0.3441 | 1.96 | 0.0000 | 0.0013 | 0.0030 | ||
| p value | ||||||||
| Maize genotype | 0.2186 | 0.4336 | 0.7079 | 0.1138 | 0.0026 | <0.0001 | ||
| State of the forage | 0.4128 | 0.3142 | 0.0230 | 0.1473 | 0.1077 | <0.0001 | ||
| Guanidinoacetic acid | 0.1994 | 0.3665 | 0.5312 | 0.0034 | 0.5732 | 0.0716 | ||
| Maize genotype × State of the forage | 0.0788 | 0.4591 | 0.2350 | 0.0751 | 0.1115 | <0.0001 | ||
| Maize genotype × Guanidinoacetic acid | 0.4496 | 0.2123 | 0.6985 | 0.0411 | 0.0004 | <0.0001 | ||
| State of the forage × Guanidinoacetic acid | 0.6044 | 0.2061 | 0.1862 | 0.1473 | 0.4167 | 0.0561 | ||
| Maize genotype × State of the forage × Guanidinoacetic acid | 0.1569 | 0.5650 | 0.6160 | 0.0751 | 0.1102 | <0.0001 | ||
| Maize Genotype | State of the Forage (SF) | Guanidinoacetic Acid (GAA) | Fermentation Characteristics 1 | CH4 Conversion Efficiency 2 | |||||
|---|---|---|---|---|---|---|---|---|---|
| pH | DMD | SCFA | ME | CH4:SCFA | CH4:ME | CH4:OM | |||
| Amarillo | Fresh | − | 7.04 | 44.09 | 3.54 | 5.50 | 72.20 | 7.54 | 9.75 |
| + | 7.09 | 37.94 | 3.73 | 5.59 | 33.63 | 3.61 | 4.74 | ||
| Ensiled | − | 6.88 | 50.07 | 5.05 | 5.93 | 111.39 | 15.10 | 20.68 | |
| + | 6.93 | 51.55 | 5.34 | 6.08 | 69.61 | 9.60 | 13.41 | ||
| SEM 3 | 0.09 | 1.38 | 0.38 | 0.19 | 21.24 | 2.48 | 3.12 | ||
| SF | 0.1619 | 0.0021 | 0.0141 | 0.0739 | 0.1515 | 0.0521 | 0.0347 | ||
| GAA | 0.6100 | 0.1650 | 0.5582 | 0.5576 | 0.1316 | 0.1296 | 0.1201 | ||
| SF × GAA | 1.0000 | 0.0502 | 0.8950 | 0.8952 | 0.9434 | 0.7671 | 0.7352 | ||
| Montesa | Fresh | − | 7.14 | 34.12 | 4.11 | 5.75 | 86.90 | 10.01 | 13.44 |
| + | 7.15 | 33.88 | 3.56 | 5.47 | 79.59 | 8.23 | 10.40 | ||
| Ensiled | − | 6.95 | 50.88 | 5.51 | 6.17 | 37.66 | 5.40 | 7.69 | |
| + | 6.97 | 51.30 | 4.67 | 5.74 | 40.15 | 5.25 | 6.98 | ||
| SEM 3 | 0.06 | 1.11 | 0.30 | 0.16 | 12.18 | 1.38 | 1.93 | ||
| SF | 0.0365 | 0.0001 | 0.0144 | 0.0946 | 0.0220 | 0.0511 | 0.0758 | ||
| GAA | 0.8426 | 0.9393 | 0.0837 | 0.0839 | 0.8528 | 0.5227 | 0.3865 | ||
| SF × GAA | 0.9050 | 0.7813 | 0.6526 | 0.6532 | 0.7083 | 0.5867 | 0.5796 | ||
| Olotillo | Fresh | − | 7.38 | 33.60 | 3.76 | 5.54 | 34.45 | 3.75 | 4.82 |
| + | 7.25 | 35.91 | 2.68 | 4.98 | 60.01 | 5.18 | 6.00 | ||
| Ensiled | − | 7.10 | 40.03 | 3.63 | 5.21 | 64.80 | 7.39 | 9.03 | |
| + | 7.09 | 44.53 | 4.21 | 5.51 | 148.36 | 18.26 | 23.31 | ||
| SEM 3 | 0.03 | 0.91 | 0.19 | 0.10 | 20.69 | 2.53 | 3.31 | ||
| SF | 0.0030 | 0.0011 | 0.0221 | 0.3828 | 0.0455 | 0.0299 | 0.0314 | ||
| GAA | 0.1167 | 0.0198 | 0.2665 | 0.2647 | 0.0577 | 0.0719 | 0.0800 | ||
| SF × GAA | 0.1383 | 0.2931 | 0.0128 | 0.0128 | 0.2336 | 0.1357 | 0.1193 | ||
| Tampiqueño | Fresh | − | 7.28 | 39.98 | 4.08 | 5.73 | 121.30 | 14.47 | 19.95 |
| + | 7.20 | 35.66 | 3.70 | 5.54 | 72.18 | 7.82 | 10.17 | ||
| Ensiled | − | 7.06 | 44.24 | 4.43 | 5.65 | 45.07 | 5.68 | 7.39 | |
| + | 7.08 | 41.22 | 3.62 | 5.23 | 35.28 | 3.92 | 4.71 | ||
| SEM 3 | 0.09 | 0.91 | 0.32 | 0.17 | 41.82 | 5.24 | 7.41 | ||
| SF | 0.1232 | 0.0057 | 0.6963 | 0.3103 | 0.2476 | 0.2922 | 0.2908 | ||
| GAA | 0.7483 | 0.0158 | 0.1406 | 0.1410 | 0.5200 | 0.4672 | 0.4476 | ||
| SF × GAA | 0.5627 | 0.5153 | 0.5396 | 0.5409 | 0.6627 | 0.6649 | 0.6567 | ||
| Tuxpeño | Fresh | − | 7.09 | 41.14 | 5.08 | 6.21 | 91.74 | 11.84 | 17.00 |
| + | 7.10 | 34.13 | 4.23 | 5.78 | 37.71 | 4.39 | 5.91 | ||
| Ensiled | − | 7.02 | 39.36 | 4.34 | 5.74 | 40.16 | 4.84 | 6.51 | |
| + | 7.10 | 42.61 | 3.69 | 5.41 | 101.72 | 11.10 | 14.07 | ||
| SEM 3 | 0.05 | 1.23 | 0.25 | 0.13 | 30.73 | 3.81 | 5.33 | ||
| SF | 0.4839 | 0.0526 | 0.0627 | 0.0306 | 0.8496 | 0.9713 | 0.8375 | ||
| GAA | 0.3778 | 0.2007 | 0.0401 | 0.0402 | 0.9084 | 0.8827 | 0.7572 | ||
| SF × GAA | 0.4282 | 0.0140 | 0.7168 | 0.7172 | 0.1332 | 0.1462 | 0.1553 | ||
| Pooled SEM 3 | 0.07 | 1.12 | 0.30 | 0.15 | 27.28 | 3.36 | 4.64 | ||
| p value | |||||||||
| Maize genotype | 0.0015 | <0.0001 | 0.0015 | 0.0006 | 0.9460 | 0.9558 | 0.9604 | ||
| State of the forage | 0.0301 | <0.0001 | 0.0002 | 0.4089 | 0.9709 | 0.5263 | 0.5823 | ||
| Guanidinoacetic acid | 0.9475 | 0.0954 | 0.0057 | 0.0057 | 0.8244 | 0.5705 | 0.4344 | ||
| Maize genotype × State of the forage | 0.0022 | <0.0001 | 0.0003 | 0.0028 | 0.0280 | 0.0212 | 0.0233 | ||
| Maize genotype × Guanidinoacetic acid | 0.6926 | 0.0024 | 0.1436 | 0.1439 | 0.1630 | 0.1965 | 0.2400 | ||
| State of the forage × Guanidinoacetic acid | 0.8693 | 0.0003 | 0.3626 | 0.3626 | 0.0871 | 0.0761 | 0.0756 | ||
| Maize genotype × State of the forage × Guanidinoacetic acid | 0.7568 | 0.0220 | 0.1358 | 0.1364 | 0.5794 | 0.5269 | 0.5363 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarado-Ramírez, E.R.; Andrade-Yucailla, V.; Elghandour, M.M.M.Y.; Acosta-Lozano, N.; Rivas-Jacobo, M.A.; López-Aguirre, D.; Garay-Martínez, J.R.; Vazquez-Mendoza, P.; Barros-Rodríguez, M.; Salem, A.Z.M. Effects of Guanidinoacetic Acid on Ruminal Fermentation and Greenhouse Gas Production Using Fresh Forage and Silage from Different Maize (Zea mays L.) Genotypes. Fermentation 2023, 9, 437. https://doi.org/10.3390/fermentation9050437
Alvarado-Ramírez ER, Andrade-Yucailla V, Elghandour MMMY, Acosta-Lozano N, Rivas-Jacobo MA, López-Aguirre D, Garay-Martínez JR, Vazquez-Mendoza P, Barros-Rodríguez M, Salem AZM. Effects of Guanidinoacetic Acid on Ruminal Fermentation and Greenhouse Gas Production Using Fresh Forage and Silage from Different Maize (Zea mays L.) Genotypes. Fermentation. 2023; 9(5):437. https://doi.org/10.3390/fermentation9050437
Chicago/Turabian StyleAlvarado-Ramírez, Edwin Rafael, Verónica Andrade-Yucailla, Mona Mohamed Mohamed Yasseen Elghandour, Néstor Acosta-Lozano, Marco Antonio Rivas-Jacobo, Daniel López-Aguirre, Jonathan Raúl Garay-Martínez, Paulina Vazquez-Mendoza, Marcos Barros-Rodríguez, and Abdelfattah Zeidan Mohamed Salem. 2023. "Effects of Guanidinoacetic Acid on Ruminal Fermentation and Greenhouse Gas Production Using Fresh Forage and Silage from Different Maize (Zea mays L.) Genotypes" Fermentation 9, no. 5: 437. https://doi.org/10.3390/fermentation9050437