
Effect of Lactobacillus curvatus HY7602-Fermented Antler on Sarcopenia in Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Fermented Deer Antler
2.2. Animals, Diet, and Experimental Design
2.3. Treadmill Exercise Performance and Limb Grip Strength Tests
2.4. Histological Analysis
2.5. Serum Biochemistry
2.6. Real-Time Polymerase Chain Reaction (RT-PCR)
2.7. Measurement of SCFA Content
2.8. Statistical Analysis
3. Results
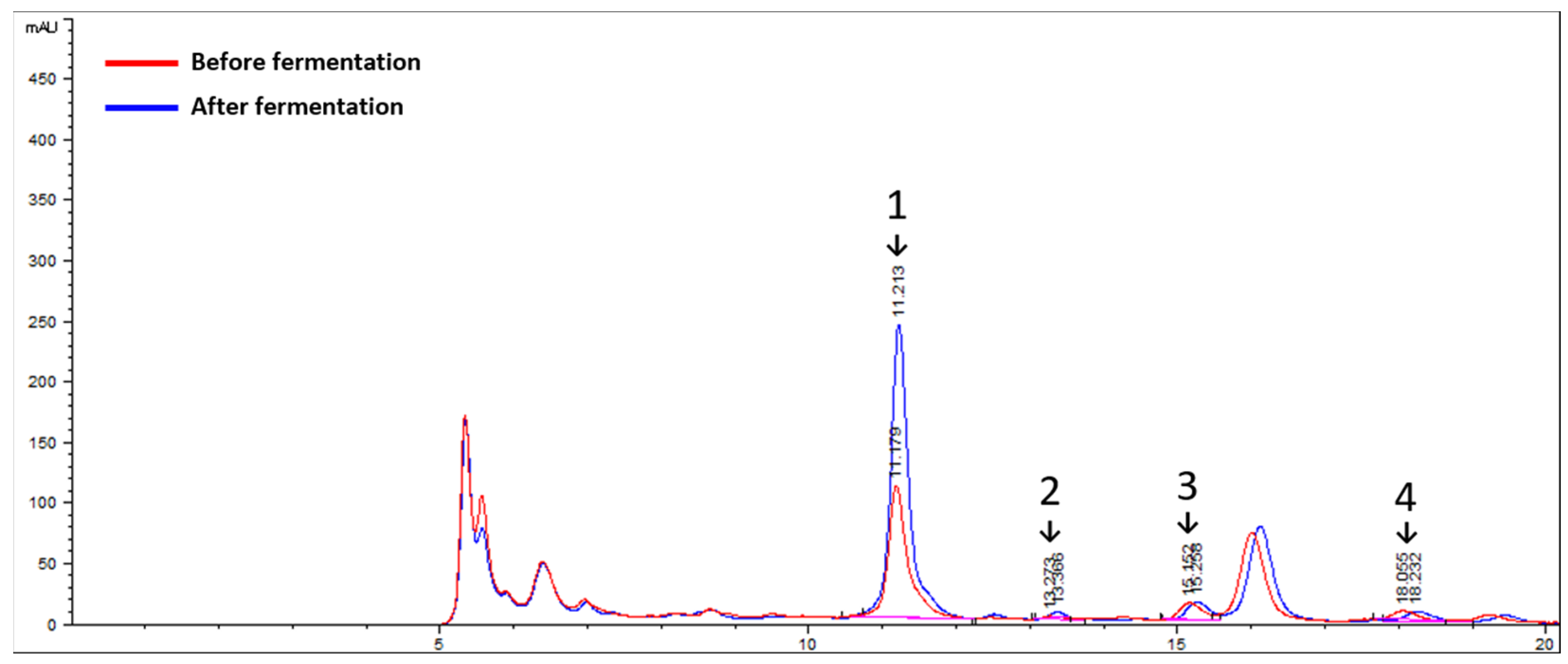
3.1. Characterization of L. curvatus HY7602 Fermentation in Antlers
3.1.1. Changes in pH Values
3.1.2. Changes in SCFA Content
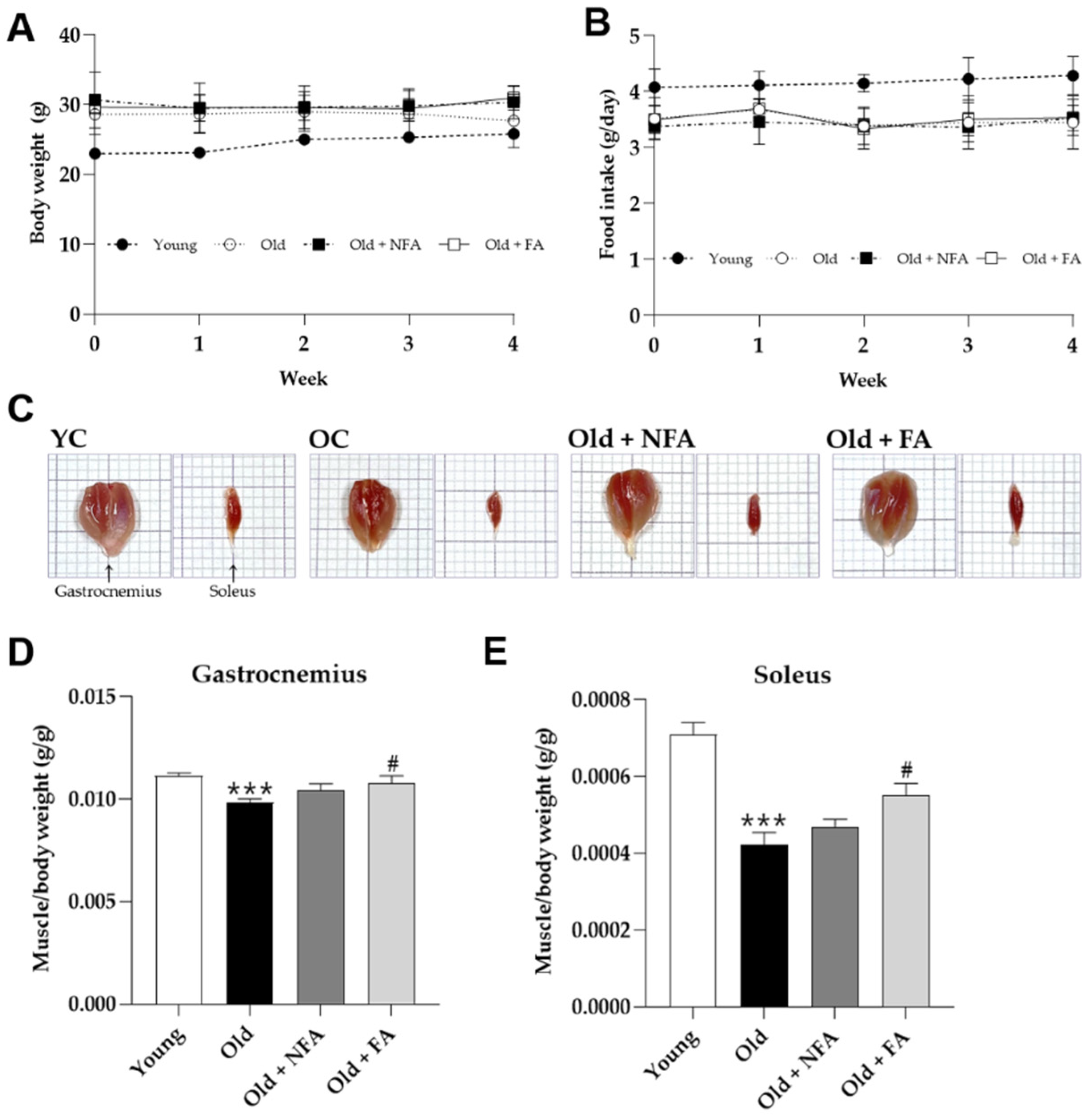
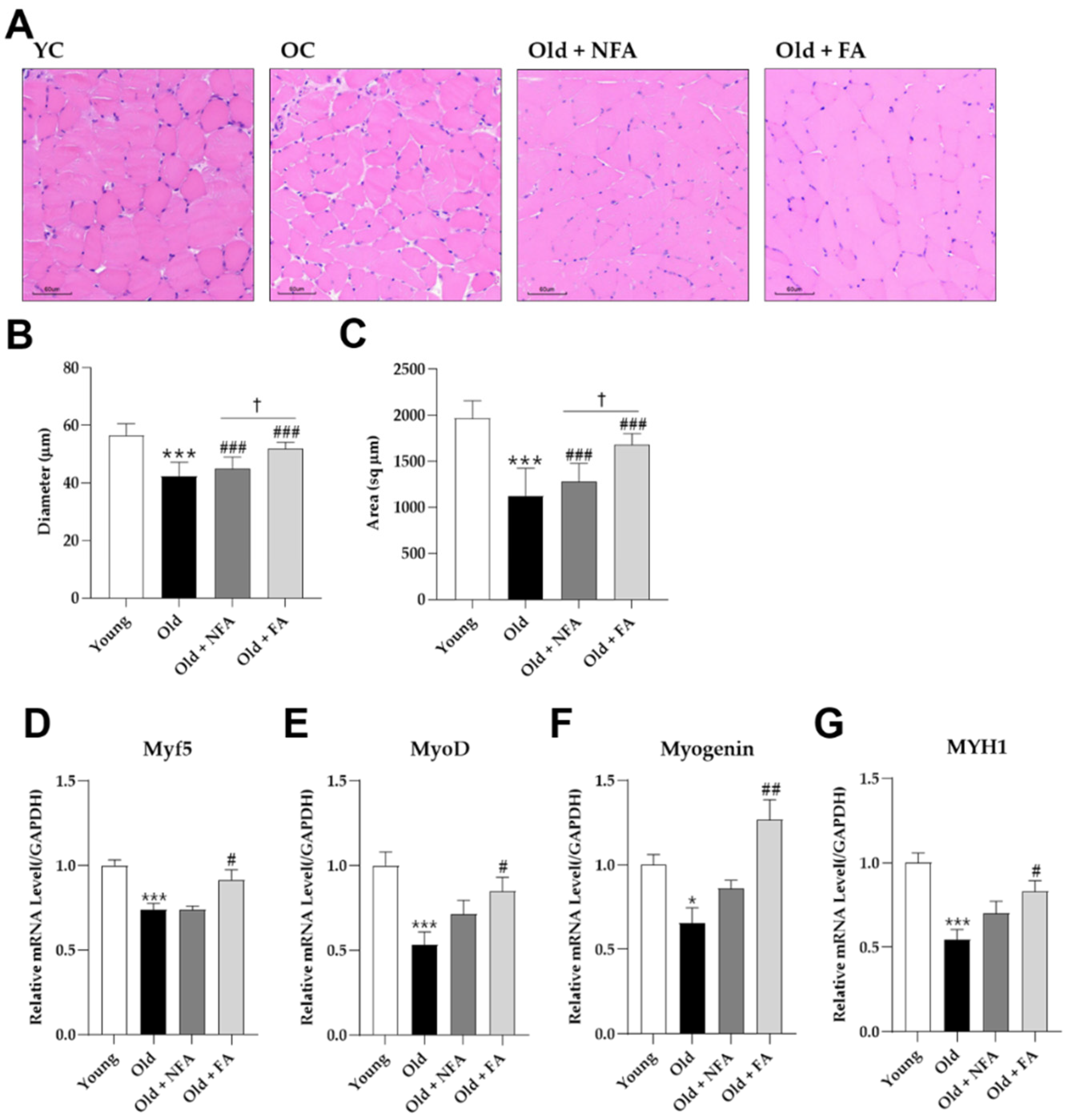
3.2. FA Recovers Muscle Loss in Aged Mice
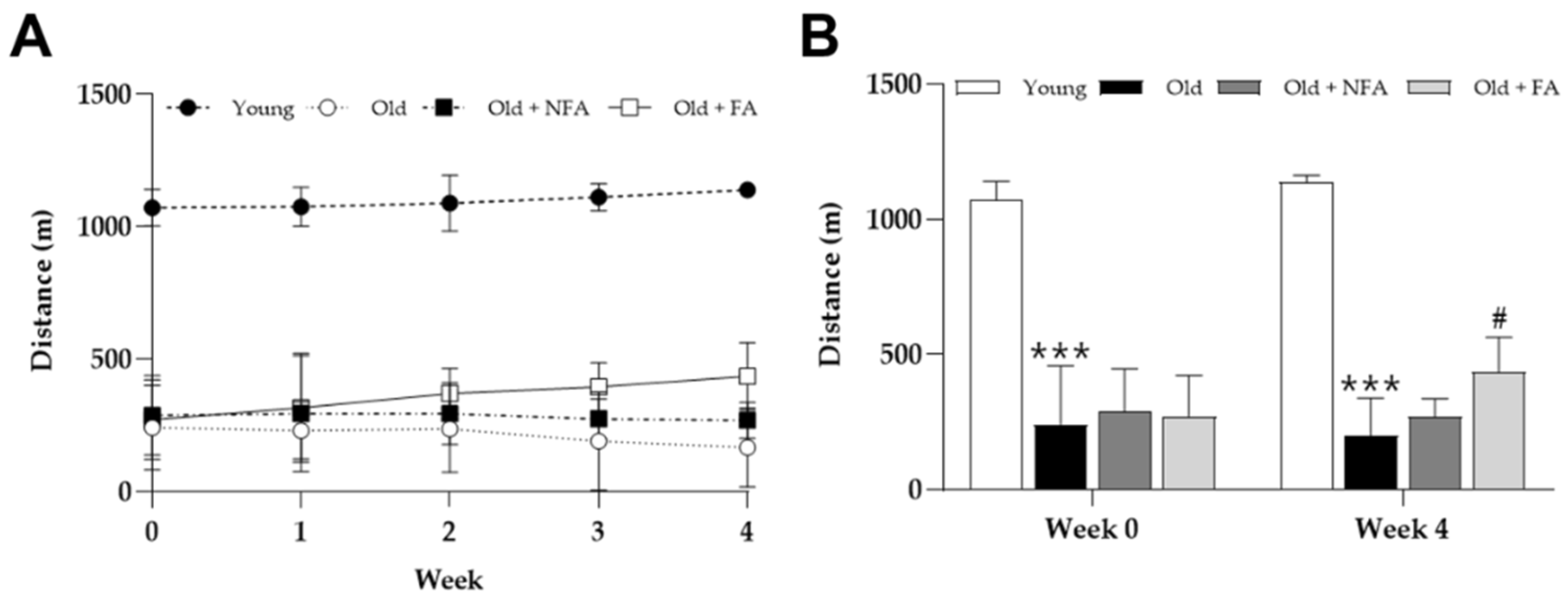
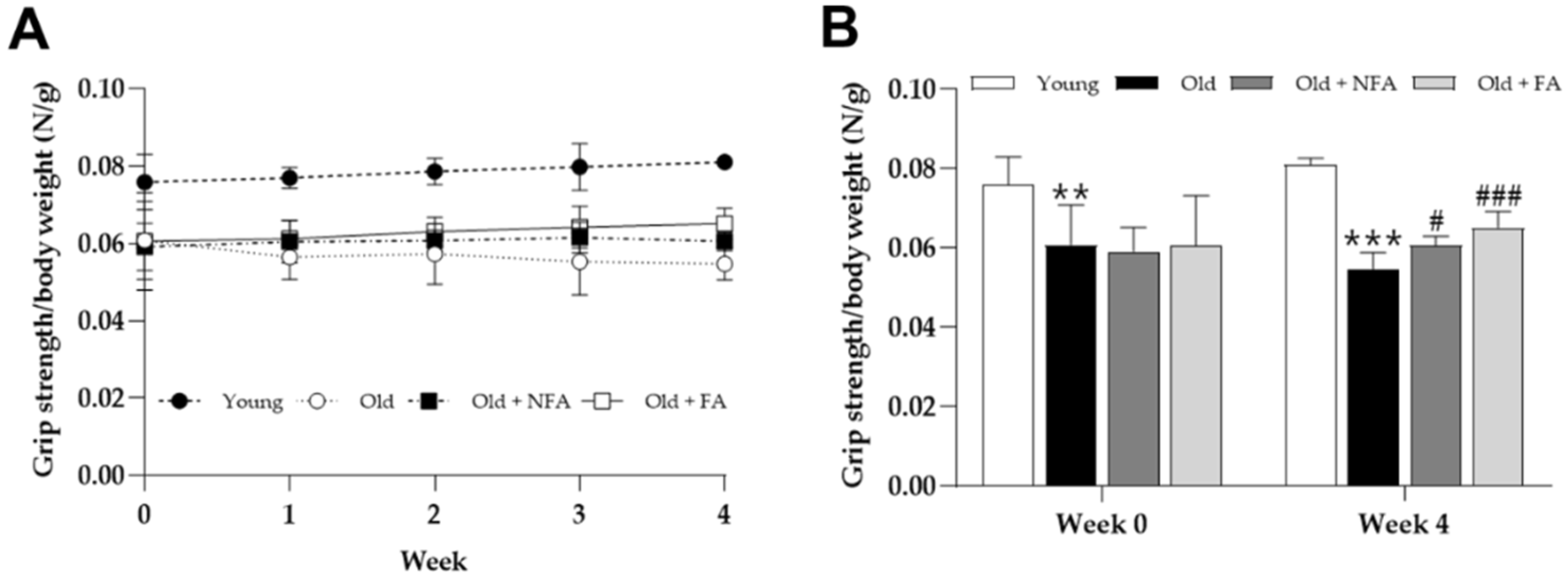
3.3. FA Ameliorates Limb Grip Strength and Exercise Performance Reduction in Aged Mice
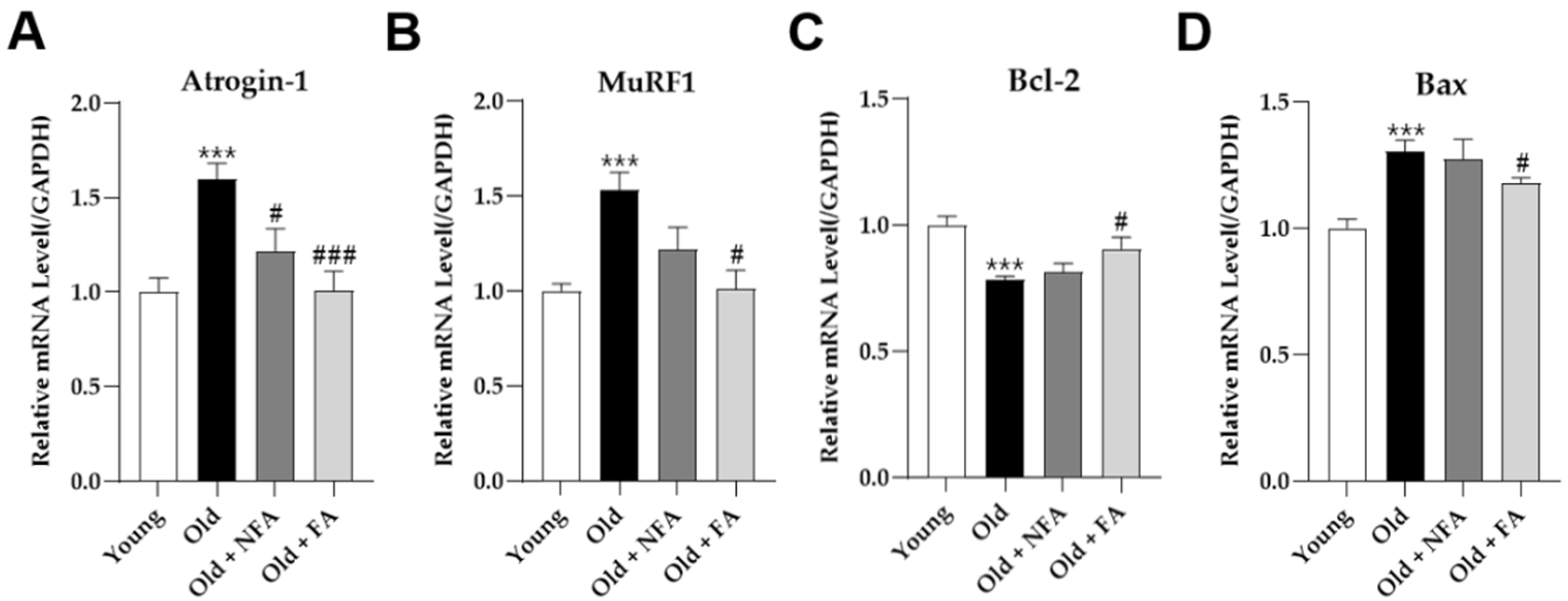
3.4. FA Inhibits Muscle Protein Degradation and Apoptosis and Promotes Muscle Fiber Regeneration
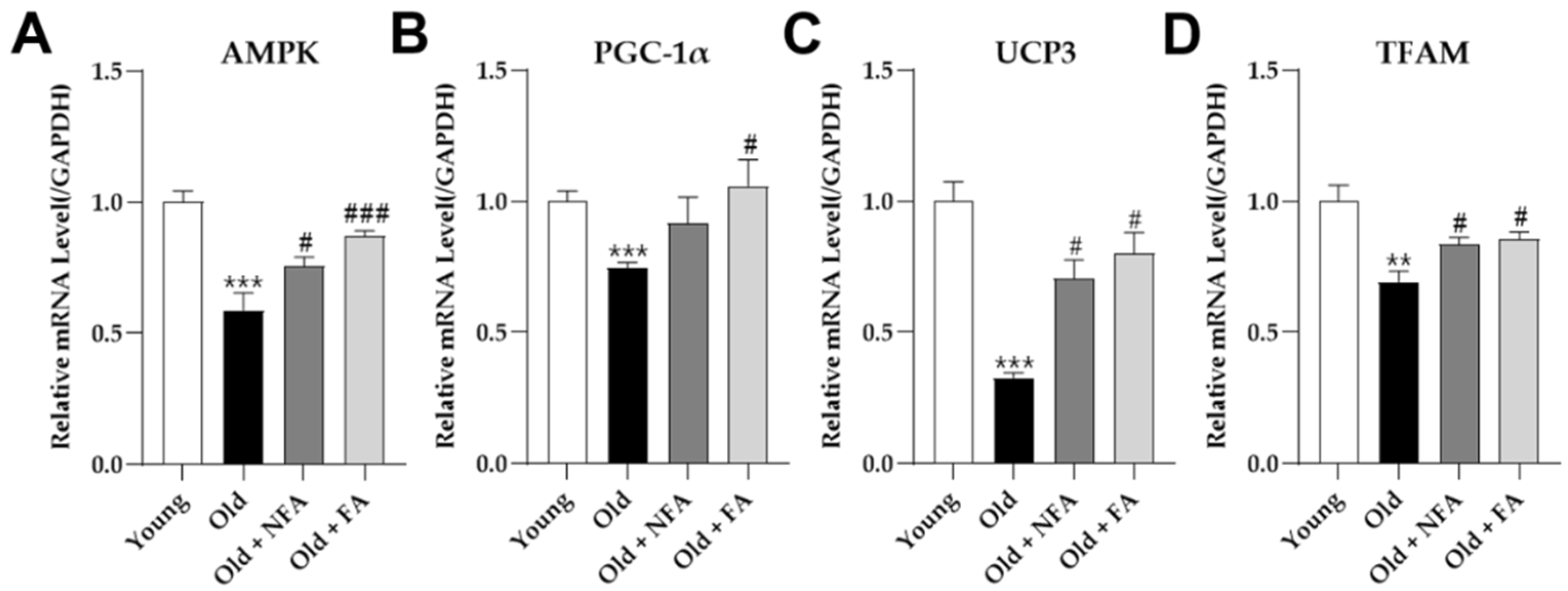
3.5. FA Promotes Mitochondrial Biogenesis in Aged Mice
3.6. Effects of the FA on Serum Biochemistry
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wilkinson, D.J.; Piasecki, M.; Atherton, P.J. The age-related loss of skeletal muscle mass and function: Measurement and physiology of muscle fibre atrophy and muscle fibre loss in humans. Ageing Res. Rev. 2018, 47, 123–132. [Google Scholar] [CrossRef]
- Mitchell, W.K.; Williams, J.; Atherton, P.; Larvin, M.; Lund, J.; Narici, M. Sarcopenia, dynapenia, and the impact of advancing age on human skeletal muscle size and strength; a quantitative review. Front. Physiol. 2012, 3, 260. [Google Scholar] [CrossRef] [PubMed]
- Phu, S.; Boersma, D.; Duque, G. Exercise and sarcopenia. J. Clin. Densitom. 2015, 18, 488–492. [Google Scholar] [CrossRef] [PubMed]
- Liguori, I.; Russo, G.; Aran, L.; Bulli, G.; Curcio, F.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Sarcopenia: Assessment of disease burden and strategies to improve outcomes. Clin. Interv. Aging 2018, 13, 913–927. [Google Scholar] [CrossRef]
- Larsson, L.; Degens, H.; Li, M.; Salviati, L.; Lee, Y.I.; Thompson, W.; Kirkland, J.L.; Sandri, M. Sarcopenia: Aging-related loss of muscle mass and function. Physiol. Rev. 2019, 99, 427–511. [Google Scholar] [CrossRef] [PubMed]
- Clavel, S.; Coldefy, A.S.; Kurkdjian, E.; Salles, J.; Margaritis, I.; Derijard, B. Atrophy-related ubiquitin ligases, atrogin-1 and MuRF1 are up-regulated in aged rat tibialis anterior muscle. Mech. Ageing Dev. 2006, 127, 794–801. [Google Scholar] [CrossRef]
- Marzetti, E.; Leeuwenburgh, C. Skeletal muscle apoptosis, sarcopenia and frailty at old age. Exp. Gerontol. 2006, 41, 1234–1238. [Google Scholar] [CrossRef]
- Leeuwenburgh, C. Role of apoptosis in sarcopenia. J. Gerontol. Ser. A 2003, 58, M999–M1001. [Google Scholar] [CrossRef]
- Alway, S.E.; Siu, P.M. Nuclear apoptosis contributes to sarcopenia. Exerc. Sport Sci. Rev. 2008, 36, 51–57. [Google Scholar] [CrossRef]
- Wagatsuma, A.; Sakuma, K. Mitochondria as a potential regulator of myogenesis. Sci. World J. 2013, 2013, 593267. [Google Scholar] [CrossRef]
- Hood, D.A.; Memme, J.M.; Oliveira, A.N.; Triolo, M. Maintenance of skeletal muscle mitochondria in health, exercise, and aging. Annu. Rev. Physiol. 2019, 81, 19–41. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhao, H.; Liu, Z.; McMahon, C. Deer antler—A novel model for studying organ regeneration in mammals. Int. J. Biochem. Cell Biol. 2014, 56, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Gilbey, A.; Perezgonzalez, J.D. Health benefits of deer and elk velvet antler supplements: A systematic review of randomised controlled studies. N. Z. Med. J. 2012, 125, 80–86. [Google Scholar] [PubMed]
- Chen, J.C.; Hsiang, C.Y.; Lin, Y.C.; Ho, T.Y. Deer antler extract improves fatigue effect through altering the expression of genes related to muscle strength in skeletal muscle of mice. Evid. Based Complement. Altern. Med. 2014, 2014, 540580. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Choi, Y.H.; Kim, K.H.; Lee, Y.C.; Kim, C.H.; Moon, S.H.; Kang, S.G.; Park, Y.G. Protective and anti-arthritic effects of deer antler aqua-acupuncture (DAA), inhibiting dihydroorotate dehydrogenase, on phosphate ions-mediated chondrocyte apoptosis and rat collagen-induced arthritis. Int. Immunopharmacol. 2004, 4, 963–973. [Google Scholar] [CrossRef]
- Sui, Z.; Zhang, L.; Huo, Y.; Zhang, Y. Bioactive components of velvet antlers and their pharmacological properties. J. Pharm. Biomed. Anal. 2014, 87, 229–240. [Google Scholar] [CrossRef]
- Parvez, S.; Malik, K.A.; Ah Kang, S.; Kim, H.Y. Probiotics and their fermented food products are beneficial for health. J. Appl. Microbiol. 2006, 100, 1171–1185. [Google Scholar] [CrossRef]
- Ranadheera, C.S.; Vidanarachchi, J.; Rocha, R.; Cruz, A.; Ajlouni, S. Probiotic Delivery through Fermentation: Dairy vs. Non-Dairy Beverages. Fermentation 2017, 3, 67. [Google Scholar] [CrossRef]
- Verardo, V.; Gómez-Caravaca, A.M.; Tabanelli, G. Bioactive components in fermented foods and food by-products. Foods 2020, 9, 153. [Google Scholar] [CrossRef]
- Rhee, S.J.; Lee, J.E.; Lee, C.H. Importance of lactic acid bacteria in Asian fermented foods. Microb. Cell Fact. 2011, 10 (Suppl. S1), S5. [Google Scholar] [CrossRef]
- Denisa, L.; Zuzana, M.; Ľubomír, V. 12. Lactic acid bacteria and fermentation of cereals and pseudocereals. In Fermentation Processes; Faustino, J.A., Ed.; IntechOpen: Rijeka, Croatia, 2017. [Google Scholar]
- Wang, Y.; Wu, J.; Lv, M.; Shao, Z.; Hungwe, M.; Wang, J.; Bai, X.; Xie, J.; Wang, Y.; Geng, W. Metabolism characteristics of lactic acid bacteria and the expanding applications in food industry. Front. Bioeng. Biotechnol. 2021, 9, 612285. [Google Scholar] [CrossRef]
- Markowiak-Kopeć, P.; Śliżewska, K. The effect of probiotics on the production of short-chain fatty acids by human intestinal microbiome. Nutrients 2020, 12, 1107. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, J.G.; Chain, F.; Martín, R.; Bermúdez-Humarán, L.G.; Courau, S.; Langella, P. Beneficial effects on host energy metabolism of short-chain fatty acids and vitamins produced by commensal and probiotic bacteria. Microb. Cell Fact. 2017, 16, 79. [Google Scholar] [CrossRef] [PubMed]
- Blaak, E.E.; Canfora, E.E.; Theis, S.; Frost, G.; Groen, A.K.; Mithieux, G.; Nauta, A.; Scott, K.; Stahl, B.; van Harsselaar, J.; et al. Short chain fatty acids in human gut and metabolic health. Benef. Microbes. 2020, 11, 411–455. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Kim, S.; Jeung, W.; Ra, J.; Heo, K.; Shim, J.; Lee, J. Fermented antler improves endurance during exercise performance by increasing mitochondrial biogenesis and muscle strength in mice. Appl. Sci. 2021, 11, 5386. [Google Scholar] [CrossRef]
- Kim, Y.-T.; Jeon, H.; Kim, S.; Heo, K.; Shim, J.; Lee, J.; Yang, D.; Kang, S.C. Fermented antler recovers stamina, muscle strength and muscle mass in middle-aged mice. Appl. Sci. 2022, 12, 106. [Google Scholar] [CrossRef]
- Jeon, H.; Kim, Y.; Jang, W.Y.; Kim, J.; Heo, K.; Shim, J.; Lee, J.; Yang, D.; Kang, S.C. Effects of Lactobacillus curvatus HY7602-fermented antlers in dexamethasone-induced muscle atrophy. Fermentation 2022, 8, 454. [Google Scholar] [CrossRef]
- Bintsis, T. Lactic acid bacteria as starter cultures: An update in their metabolism and genetics. AIMS Microbiol. 2018, 4, 665–684. [Google Scholar] [CrossRef]
- Keller, K.; Engelhardt, M. Strength and muscle mass loss with aging process. Age and strength loss. Muscle Ligaments Tendons J. 2013, 3, 346–350. [Google Scholar] [CrossRef]
- Song, W.; Kwak, H.B.; Lawler, J.M. Exercise training attenuates age-induced changes in apoptotic signaling in rat skeletal muscle. Antioxid. Redox Signal. 2006, 8, 517–528. [Google Scholar] [CrossRef]
- Frontera, W.R.; Reid, K.F.; Phillips, E.M.; Krivickas, L.S.; Hughes, V.A.; Roubenoff, R.; Fielding, R.A. Muscle fiber size and function in elderly humans: A longitudinal study. J. Appl. Physiol. 2008, 105, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Tedesco, F.S.; Dellavalle, A.; Diaz-Manera, J.; Messina, G.; Cossu, G. Repairing skeletal muscle: Regenerative potential of skeletal muscle stem cells. J. Clin. Investig. 2010, 120, 11–19. [Google Scholar] [CrossRef]
- Domingues-Faria, C.; Vasson, M.P.; Goncalves-Mendes, N.; Boirie, Y.; Walrand, S. Skeletal muscle regeneration and impact of aging and nutrition. Ageing Res. Rev. 2016, 26, 22–36. [Google Scholar] [CrossRef]
- Peterson, C.M.; Johannsen, D.L.; Ravussin, E. Skeletal muscle mitochondria and aging: A review. J. Aging Res. 2012, 2012, 194821. [Google Scholar] [CrossRef] [PubMed]
- Gumucio, J.P.; Mendias, C.L. Atrogin-1, MuRF-1, and sarcopenia. Endocrine 2013, 43, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Bodine, S.C.; Baehr, L.M. Skeletal muscle atrophy and the E3 ubiquitin ligases MuRF1 and MAFbx/atrogin-1. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E469–E484. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Marzetti, E.; Calvani, R.; Bernabei, R.; Leeuwenburgh, C. Apoptosis in skeletal myocytes: A potential target for interventions against sarcopenia and physical frailty—A mini-review. Gerontology 2012, 58, 99–106. [Google Scholar] [CrossRef]
- Bentzinger, C.F.; Wang, Y.X.; Rudnicki, M.A. Building muscle: Molecular regulation of myogenesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008342. [Google Scholar] [CrossRef]
- Muñoz-Cánoves, P.; Neves, J.; Sousa-Victor, P. Understanding muscle regenerative decline with aging: New approaches to bring back youthfulness to aged stem cells. FEBS J. 2020, 287, 406–416. [Google Scholar] [CrossRef]
- LaFramboise, W.A.; Guthrie, R.D.; Scalise, D.; Elborne, V.; Bombach, K.L.; Armanious, C.S.; Magovern, J.A. Effect of muscle origin and phenotype on satellite cell muscle-specific gene expression. J. Mol. Cell. Cardiol. 2003, 35, 1307–1318. [Google Scholar] [CrossRef]
- Schiaffino, S.; Rossi, A.C.; Smerdu, V.; Leinwand, L.A.; Reggiani, C. Developmental myosins: Expression patterns and functional significance. Skelet. Muscle 2015, 5, 22. [Google Scholar] [CrossRef]
- Gan, Z.; Fu, T.; Kelly, D.P.; Vega, R.B. Skeletal muscle mitochondrial remodeling in exercise and diseases. Cell Res. 2018, 28, 969–980. [Google Scholar] [CrossRef] [PubMed]
- Jornayvaz, F.R.; Shulman, G.I. Regulation of mitochondrial biogenesis. Essays Biochem. 2010, 47, 69–84. [Google Scholar] [CrossRef]
- Le Couteur, D.G.; Blyth, F.M.; Creasey, H.M.; Handelsman, D.J.; Naganathan, V.; Sambrook, P.N.; Seibel, M.J.; Waite, L.M.; Cumming, R.G. The association of alanine transaminase with aging, frailty, and mortality. J. Gerontol. A. Biol. Sci. Med. Sci. 2010, 65, 712–717. [Google Scholar] [CrossRef] [PubMed]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The role of short-chain fatty acids from gut microbiota in gut-brain communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef]
- Frampton, J.; Murphy, K.G.; Frost, G.; Chambers, E.S. Short-chain fatty acids as potential regulators of skeletal muscle metabolism and function. Nat. Metab. 2020, 2, 840–848. [Google Scholar] [CrossRef]
- Han, Q.; Huang, X.; Yan, F.; Yin, J.; Xiao, Y. The role of gut microbiota in the skeletal muscle development and fat deposition in pigs. Antibiotics 2022, 11, 793. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 0 h | 2 h | 4 h | 6 h | 8 h | 24 h | |
|---|---|---|---|---|---|---|
| pH | 6.20 ± 0.10 | 4.96 ± 0.05 | 4.51± 0.03 | 4.27 ± 0.12 | 4.11 ± 0.08 | 4.09 ± 0.01 |
| Contents (µg/mL) | |||
|---|---|---|---|
| Acetate | Propionate | Butyrate | |
| Before Fermentation | 14.17 ± 2.41 | 7.73 ± 1.05 | 1.31 ± 0.56 |
| After Fermentation | 24.29 ± 5.70 * | 6.87 ± 0.69 | 2.01 ± 0.81 |
| ALT (U/L) | AST (U/L) | ALP (U/L) | CK (U/L) | Crea (mg/dL) | |
|---|---|---|---|---|---|
| Young | 31.2 ± 3.3 | 54.3 ± 5.5 | 409 ± 24 | 132 ± 18 | 0.38 ± 0.01 |
| Old | 36.7 ± 3.9 * | 70.9 ± 11.0 * | 316 ± 60 * | 158 ± 36 | 0.35 ± 0.04 |
| Old + NFA | 32.6 ± 5.4 | 62.5 ± 8.8 | 272 ± 22 | 139 ± 26 | 0.31 ± 0.02 # |
| Old + FA | 31.3 ± 2.6 # | 60.2 ± 4.9 | 298 ± 35 | 99 ± 26 # | 0.31 ± 0.01 # |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeon, H.; Lee, K.; Kim, J.-Y.; Shim, J.-J.; Lee, J.-L. Effect of Lactobacillus curvatus HY7602-Fermented Antler on Sarcopenia in Mice. Fermentation 2023, 9, 429. https://doi.org/10.3390/fermentation9050429
Jeon H, Lee K, Kim J-Y, Shim J-J, Lee J-L. Effect of Lactobacillus curvatus HY7602-Fermented Antler on Sarcopenia in Mice. Fermentation. 2023; 9(5):429. https://doi.org/10.3390/fermentation9050429
Chicago/Turabian StyleJeon, Hyejin, Kippeum Lee, Joo-Yun Kim, Jae-Jung Shim, and Jung-Lyoul Lee. 2023. "Effect of Lactobacillus curvatus HY7602-Fermented Antler on Sarcopenia in Mice" Fermentation 9, no. 5: 429. https://doi.org/10.3390/fermentation9050429