
Metagenomic and Meta-Transcriptomic Analysis Reveal the Colonization and Expression Profile of Probiotic Strains in Humans and Animals

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Overall Profile of Probiotic Strains in Different Host Species
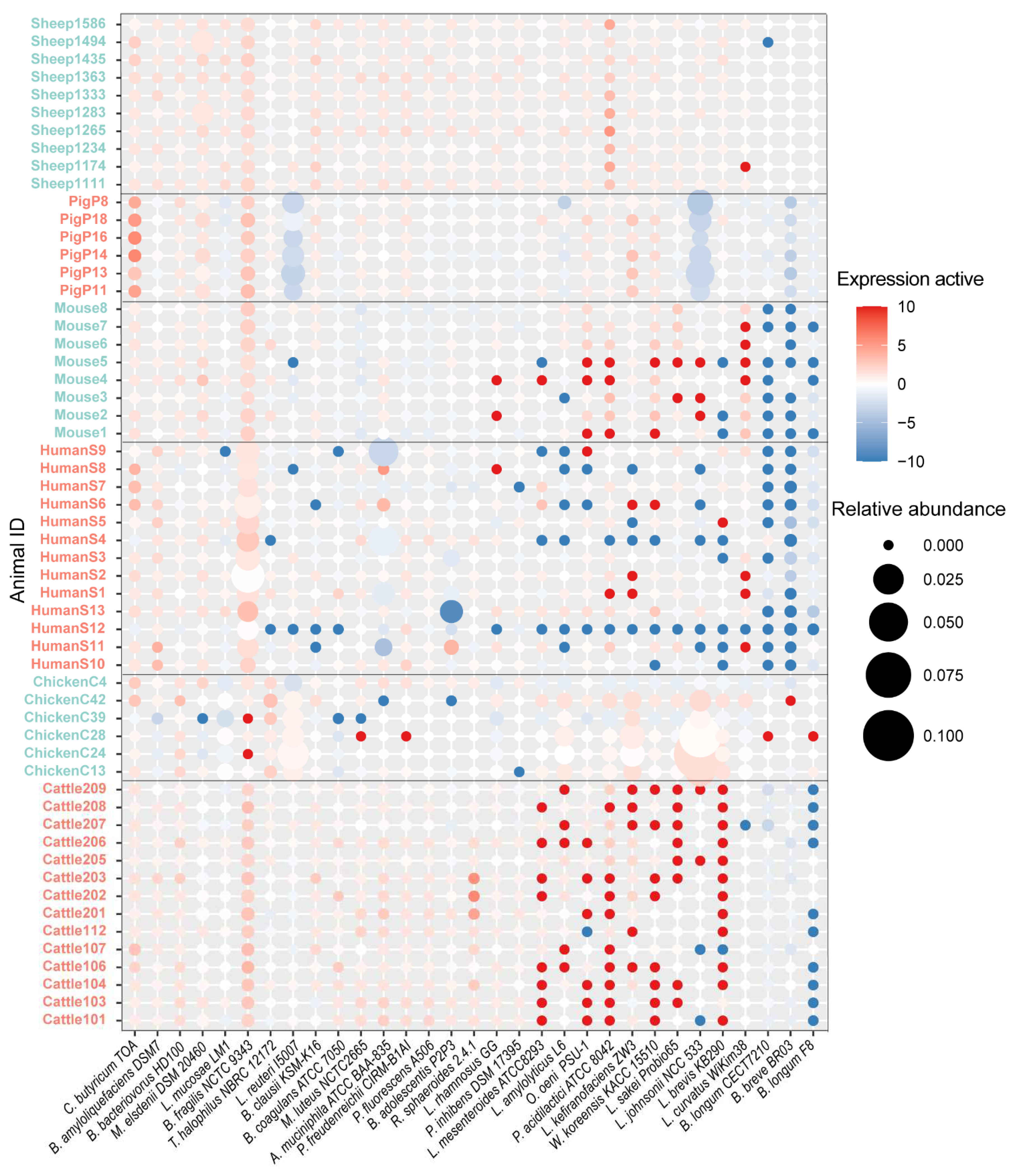
3.2. An Expression Profile of Probiotic Strains in Humans and Animals
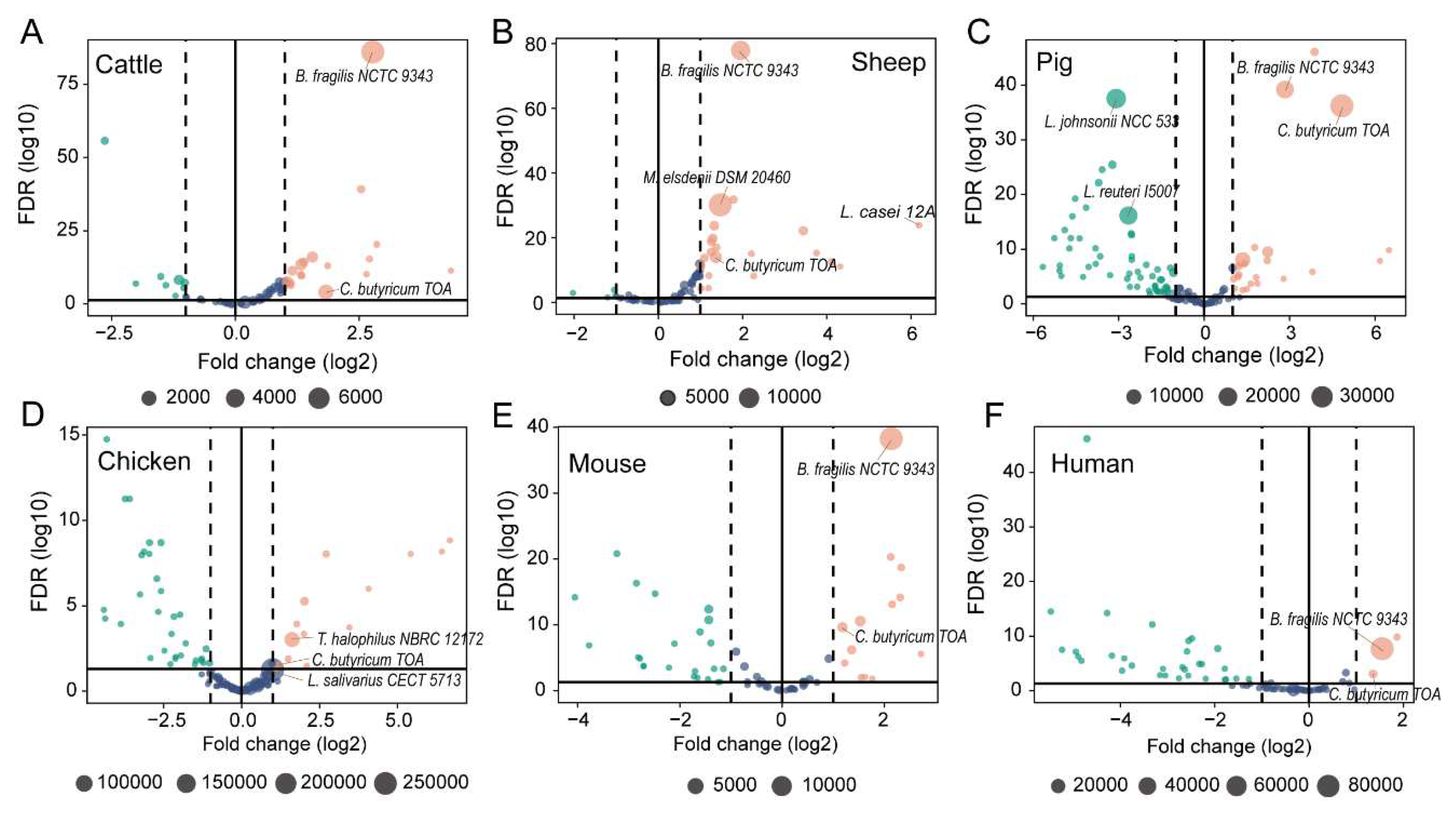
3.3. The Extremely Highly Expressed Probiotic Strains in the Gut or Rumen
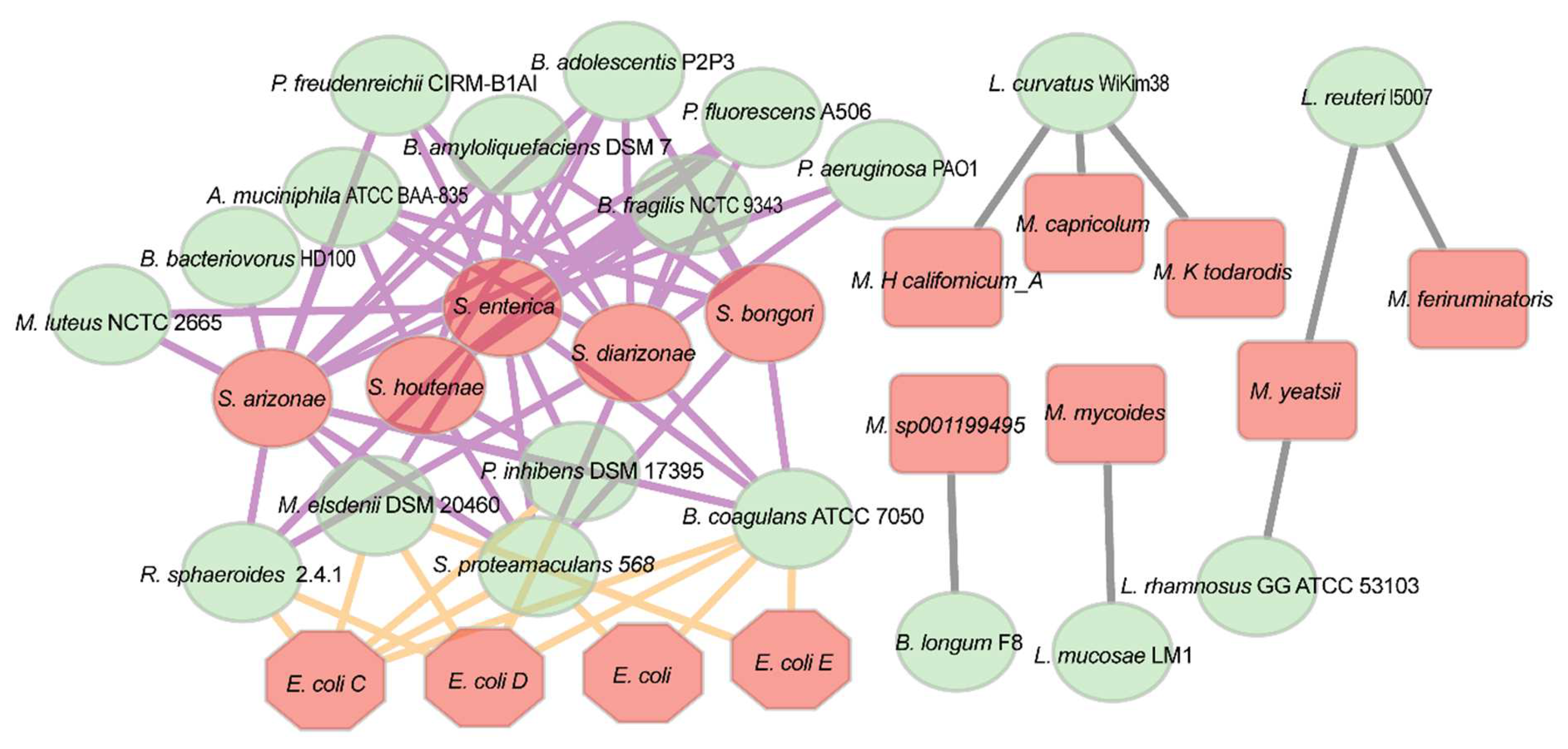
3.4. Network Analysis at Expression Level Revealed Associations between Probiotics with Pathogens
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rianda, D.; Agustina, R.; Setiawan, E.A.; Manikam, N.R.M. Effect of probiotic supplementation on cognitive function in children and adolescents: A systematic review of randomised trials. Benef. Microbes 2019, 10, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Galdeano, C.M.; Cazorla, S.I.; Dumit, J.M.L.; Vélez, E.; Perdigón, G. Beneficial effects of probiotic consumption on the immune system. Ann. Nutr. Metab. 2019, 74, 115–124. [Google Scholar]
- Jha, R.; Das, R.; Oak, S.; Mishra, P. Probiotics (direct-fed microbials) in poultry nutrition and their effects on nutrient utilization, growth and laying performance, and gut health: A systematic review. Animals 2020, 10, 1863. [Google Scholar] [CrossRef] [PubMed]
- Kambale, R.M.; Nancy, F.I.; Ngaboyeka, G.A.; Kasengi, J.B.; Bindels, L.B.; Van der Linden, D. Effects of probiotics and synbiotics on diarrhea in undernourished children: Systematic review with meta-analysis. Clin. Nutr. 2021, 40, 3158–3169. [Google Scholar] [CrossRef]
- Curciarello, R.; Canziani, K.E.; Salto, I.; Barbiera Romero, E.; Rocca, A.; Doldan, I.; Peton, E.; Brayer, S.; Sambuelli, A.M.; Goncalves, S.; et al. Probiotic Lactobacilli isolated from kefir promote down-regulation of inflammatory lamina propria t cells from patients with active IBD. Front. Pharmacol. 2021, 12, 658026. [Google Scholar] [CrossRef]
- Ebell, M.H. Probiotic Ineffective for Treatment of Acute Gastroenteritis in Young Children. Am. Fam. Physician 2019, 99, 717. [Google Scholar]
- Kothari, D.; Patel, S.; Kim, S.-K. Probiotic supplements might not be universally-effective and safe: A review. Biomed. Pharmacother. 2019, 111, 537–547. [Google Scholar] [CrossRef]
- Betancur, C.; Martínez, Y.; Tellez-Isaias, G.; Avellaneda, M.C.; Velázquez-Martí, B. In vitro characterization of indigenous probiotic strains isolated from Colombian creole pigs. Animals 2020, 10, 1204. [Google Scholar] [CrossRef]
- Chiang, M.-L.; Chen, H.-C.; Chen, K.-N.; Lin, Y.-C.; Lin, Y.-T.; Chen, M.-J. Optimizing production of two potential probiotic Lactobacilli strains isolated from piglet feces as feed additives for weaned piglets. Asian-Australas. J. Anim. Sci. 2015, 28, 1163. [Google Scholar] [CrossRef]
- Ouwehand, A.C.; Salminen, S.; Isolauri, E. Probiotics: An overview of beneficial effects. In Proceedings of the Lactic Acid Bacteria: Genetics, Metabolism and Applications: Proceedings of the Seventh Symposium on Lactic acid Bacteria: Genetics, Metabolism and Applications, Egmond aan Zee, The Netherlands, 1–5 September 2002; Springer: Dordrecht, The Netherlands, 2002; pp. 279–289. [Google Scholar]
- Wang, Y.; Hu, Y.; Liu, F.; Cao, J.; Lv, N.; Zhu, B.; Zhang, G.; Gao, G.F. Integrated metagenomic and metatranscriptomic profiling reveals differentially expressed resistomes in human, chicken, and pig gut microbiomes. Environ. Int. 2020, 138, 105649. [Google Scholar] [CrossRef]
- Li, F.; Hitch, T.C.; Chen, Y.; Creevey, C.J.; Guan, L.L. Comparative metagenomic and metatranscriptomic analyses reveal the breed effect on the rumen microbiome and its associations with feed efficiency in beef cattle. Microbiome 2019, 7, 6. [Google Scholar] [CrossRef]
- Kamke, J.; Kittelmann, S.; Soni, P.; Li, Y.; Tavendale, M.; Ganesh, S.; Janssen, P.H.; Shi, W.; Froula, J.; Rubin, E.M.; et al. Rumen metagenome and metatranscriptome analyses of low methane yield sheep reveals a Sharpea-enriched microbiome characterised by lactic acid formation and utilisation. Microbiome 2016, 4, 56. [Google Scholar] [CrossRef]
- Chung, Y.W.; Gwak, H.-J.; Moon, S.; Rho, M.; Ryu, J.-H. Functional dynamics of bacterial species in the mouse gut microbiome revealed by metagenomic and metatranscriptomic analyses. PLoS ONE 2020, 15, e0227886. [Google Scholar] [CrossRef]
- Tisza, M.J.; Buck, C.B. A catalog of tens of thousands of viruses from human metagenomes reveals hidden associations with chronic diseases. Proc. Natl. Acad. Sci. USA 2021, 118, e2023202118. [Google Scholar] [CrossRef]
- Sun, Y.; Li, H.; Zheng, L.; Li, J.; Hong, Y.; Liang, P.; Kwok, L.-Y.; Zuo, Y.; Zhang, W.; Zhang, H. iProbiotics: A machine learning platform for rapid identification of probiotic properties from whole-genome primary sequences. Brief. Bioinform. 2021, 23, bbab477. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Rosen, B.D.; Bickhart, D.M.; Schnabel, R.D.; Koren, S.; Elsik, C.G.; Tseng, E.; Rowan, T.N.; Low, W.Y.; Zimin, A.; Couldrey, C.; et al. De novo assembly of the cattle reference genome with single-molecule sequencing. Gigascience 2020, 9, giaa021. [Google Scholar] [CrossRef]
- The International Sheep Genomics Consortium; Archibald, A.; Cockett, N.; Dalrymple, B.; Faraut, T.; Kijas, J.; Maddox, J.; McEwan, J.; Hutton Oddy, V.; Raadsma, H.; et al. The sheep genome reference sequence: A work in progress. Anim. Genet. 2010, 41, 449–453. [Google Scholar]
- Wallis, J.W.; Aerts, J.; Groenen, M.A.; Crooijmans, R.P.; Layman, D.; Graves, T.A.; Scheer, D.E.; Kremitzki, C.; Fedele, M.J.; Mudd, N.K.; et al. A physical map of the chicken genome. Nature 2004, 432, 761–764. [Google Scholar] [CrossRef]
- Groenen, M.A.; Archibald, A.L.; Uenishi, H.; Tuggle, C.K.; Takeuchi, Y.; Rothschild, M.F.; Rogel-Gaillard, C.; Park, C.; Milan, D.; Megens, H.-J.; et al. Analyses of pig genomes provide insight into porcine demography and evolution. Nature 2012, 491, 393–398. [Google Scholar] [CrossRef]
- Keane, T.M.; Goodstadt, L.; Danecek, P.; White, M.A.; Wong, K.; Yalcin, B.; Heger, A.; Agam, A.; Slater, G.; Goodson, M.; et al. Mouse genomic variation and its effect on phenotypes and gene regulation. Nature 2011, 477, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Nurk, S.; Koren, S.; Rhie, A.; Rautiainen, M.; Bzikadze, A.V.; Mikheenko, A.; Vollger, M.R.; Altemose, N.; Uralsky, L.; Gershman, A.; et al. The complete sequence of a human genome. Science 2022, 376, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Rotmistrovsky, K.; Agarwala, R. BMTagger: Best Match Tagger for removing human reads from metagenomics datasets. 2011; preprint. [Google Scholar]
- Wood, D.E.; Lu, J.; Langmead, B. Improved metagenomic analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Chuvochina, M.; Chaumeil, P.-A.; Rinke, C.; Mussig, A.J.; Hugenholtz, P. A complete domain-to-species taxonomy for Bacteria and Archaea. Nat. Biotechnol. 2020, 38, 1079–1086. [Google Scholar] [CrossRef]
- Youngblut, N.D.; Ley, R.E. Struo2: Efficient metagenome profiling database construction for ever-expanding microbial genome datasets. PeerJ 2021, 9, e12198. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Watts, S.C.; Ritchie, S.C.; Inouye, M.; Holt, K.E. FastSpar: Rapid and scalable correlation estimation for compositional data. Bioinformatics 2019, 35, 1064–1066. [Google Scholar] [CrossRef]
- Smoot, M.E.; Ono, K.; Ruscheinski, J.; Wang, P.-L.; Ideker, T. Cytoscape 2.8: New features for data integration and network visualization. Bioinformatics 2011, 27, 431–432. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2. Wiley Interdiscip. Rev. Comput. Stat. 2011, 3, 180–185. [Google Scholar] [CrossRef]
- Grotkjær, T.; Bentzon-Tilia, M.; D’Alvise, P.; Dierckens, K.; Bossier, P.; Gram, L. Phaeobacter inhibens as probiotic bacteria in non-axenic Artemia and algae cultures. Aquaculture 2016, 462, 64–69. [Google Scholar] [CrossRef]
- García-Galán, A.; De la Fe, C.; Gomis, J.; Bataller, E.; Sánchez, A.; Quereda, J.; García-Roselló, E.; Gómez-Martín, A. The addition of Lactobacillus spp. negatively affects Mycoplasma bovis viability in bovine cervical mucus. BMC Vet. Res. 2020, 16, 251. [Google Scholar] [CrossRef]
- Ma, C.; Zhang, C.; Chen, D.; Jiang, S.; Shen, S.; Huo, D.; Huang, S.; Zhai, Q.; Zhang, J. Probiotic consumption influences universal adaptive mutations in indigenous human and mouse gut microbiota. Commun. Biol. 2021, 4, 1198. [Google Scholar] [CrossRef]
- Aziz, G.; Tariq, M.; Zaidi, A.H. Mining indigenous honeybee gut microbiota for Lactobacillus with probiotic potential. Microbiology 2021, 167, 001032. [Google Scholar] [CrossRef]
- Lecocq, A.; Natsopoulou, M.; Berggreen, I.; Eilenberg, J.; Heckmann, L.-H.L.; Nielsen, H.; Stensvold, C.; Jensen, A. Probiotic properties of an indigenous Pediococcus pentosaceus strain on Tenebrio molitor larval growth and survival. J. Insects Food Feed 2021, 7, 975–986. [Google Scholar] [CrossRef]
- Rebollar, E.A.; Antwis, R.E.; Becker, M.H.; Belden, L.K.; Bletz, M.C.; Brucker, R.M.; Harrison, X.A.; Hughey, M.C.; Kueneman, J.G.; Loudon, A.H.; et al. Using “omics” and integrated multi-omics approaches to guide probiotic selection to mitigate chytridiomycosis and other emerging infectious diseases. Front. Microbiol. 2016, 7, 68. [Google Scholar] [CrossRef]
- Yan, W.; Sun, C.; Yuan, J.; Yang, N. Gut metagenomic analysis reveals prominent roles of Lactobacillus and cecal microbiota in chicken feed efficiency. Sci. Rep. 2017, 7, 45308. [Google Scholar] [CrossRef]
- Fesseha, H.; Demlie, T.; Mathewos, M.; Eshetu, E. Effect of Lactobacillus species probiotics on growth performance of dual-purpose chicken. Vet. Med. Res. Rep. 2021, 12, 75–83. [Google Scholar] [CrossRef]
- Forte, C.; Manuali, E.; Abbate, Y.; Papa, P.; Vieceli, L.; Tentellini, M.; Trabalza-Marinucci, M.; Moscati, L. Dietary Lactobacillus acidophilus positively influences growth performance, gut morphology, and gut microbiology in rurally reared chickens. Poult. Sci. 2018, 97, 930–936. [Google Scholar] [CrossRef]
- Peng, Q.; Zeng, X.; Zhu, J.; Wang, S.; Liu, X.; Hou, C.; Thacker, P.; Qiao, S. Effects of dietary Lactobacillus plantarum B1 on growth performance, intestinal microbiota, and short chain fatty acid profiles in broiler chickens. Poult. Sci. 2016, 95, 893–900. [Google Scholar] [CrossRef]
- Nakphaichit, M.; Sobanbua, S.; Siemuang, S.; Vongsangnak, W.; Nakayama, J.; Nitisinprasert, S. Protective effect of Lactobacillus reuteri KUB-AC5 against Salmonella Enteritidis challenge in chickens. Benef. Microbes 2019, 10, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Tian, Y.; Cao, Y.; Li, J.; Guo, H.; Su, Y.; Tian, Y.; Wang, C.; Wang, T.; Zhang, L. Probiotic properties of Lactobacillus paracasei subsp. paracasei L1 and its growth performance-promotion in chicken by improving the intestinal microflora. Front. Physiol. 2019, 10, 937. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Ni, X.; Zeng, D.; Wang, H.; Jing, B.; Yin, Z.; Pan, K. Effect of a dietary probiotic, Lactobacillus johnsonii BS15, on growth performance, quality traits, antioxidant ability, and nutritional and flavour substances of chicken meat. Anim. Prod. Sci. 2016, 57, 920–926. [Google Scholar] [CrossRef]
- Olnood, C.G.; Beski, S.S.; Choct, M.; Iji, P.A. Novel probiotics: Their effects on growth performance, gut development, microbial community and activity of broiler chickens. Anim. Nutr. 2015, 1, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Fatufe, A.; Matanmi, O. The effect of probiotics supplementation on the growth performance of two strains of cockerels. J. Cent. Eur. Agric. 2008, 9. [Google Scholar] [CrossRef]
- Yasueda, A.; Mizushima, T.; Nezu, R.; Sumi, R.; Tanaka, M.; Nishimura, J.; Kai, Y.; Hirota, M.; Osawa, H.; Nakajima, K.; et al. The effect of Clostridium butyricum MIYAIRI on the prevention of pouchitis and alteration of the microbiota profile in patients with ulcerative colitis. Surg. Today 2016, 46, 939–949. [Google Scholar] [CrossRef]
- Chen, L.; Li, S.; Zheng, J.; Li, W.; Jiang, X.; Zhao, X.; Li, J.; Che, L.; Lin, Y.; Xu, S.; et al. Effects of dietary Clostridium butyricum supplementation on growth performance, intestinal development, and immune response of weaned piglets challenged with lipopolysaccharide. J. Anim. Sci. Biotechnol. 2018, 9, 62. [Google Scholar] [CrossRef]
- Yang, C.; Cao, G.; Ferket, P.; Liu, T.; Zhou, L.; Zhang, L.; Xiao, Y.; Chen, A. Effects of probiotic, Clostridium butyricum, on growth performance, immune function, and cecal microflora in broiler chickens. Poult. Sci. 2012, 91, 2121–2129. [Google Scholar] [CrossRef]
- Kohiruimaki, M.; Ohtsuka, H.; Tanami, E.; Kitagawa, M.; Masui, M.; Ando, T.; Kawamura, S. Effects of active egg white product/Clostridium butyricum Miyairi 588 additive on peripheral leukocyte populations in periparturient dairy cows. J. Vet. Med. Sci. 2008, 70, 321–323. [Google Scholar] [CrossRef]
- Cai, L.; Yu, J.; Hartanto, R.; Qi, D. Dietary supplementation with Saccharomyces cerevisiae, Clostridium butyricum and their combination ameliorate rumen fermentation and growth performance of heat-stressed goats. Animals 2021, 11, 2116. [Google Scholar] [CrossRef]
- Ariyoshi, T.; Hagihara, M.; Takahashi, M.; Mikamo, H. Effect of Clostridium butyricum on gastrointestinal infections. Biomedicines 2022, 10, 483. [Google Scholar] [CrossRef]
- Tran, N.T.; Li, Z.; Ma, H.; Zhang, Y.; Zheng, H.; Gong, Y.; Li, S. Clostridium butyricum: A promising probiotic confers positive health benefits in aquatic animals. Rev. Aquac. 2020, 12, 2573–2589. [Google Scholar] [CrossRef]
- Sato, Y.; Kuroki, Y.; Oka, K.; Takahashi, M.; Rao, S.; Sukegawa, S.; Fujimura, T. Effects of dietary supplementation with Enterococcus faecium and Clostridium butyricum, either alone or in combination, on growth and fecal microbiota composition of post-weaning pigs at a commercial farm. Front. Vet. 2019, 6, 26. [Google Scholar] [CrossRef]
- Wang, J.; Ishfaq, M.; Guo, Y.; Chen, C.; Li, J. Assessment of probiotic properties of Lactobacillus salivarius isolated from chickens as feed additives. Front. Vet. Sci. 2020, 7, 415. [Google Scholar] [CrossRef]
- Waites, K.B.; Talkington, D.F. Mycoplasma pneumoniae and its role as a human pathogen. Clin. Microbiol. Rev. 2004, 17, 697–728. [Google Scholar] [CrossRef]
- Mackie, D.; Ball, H.; Logan, E. Isolation of Mycoplasma californicum from an outbreak of bovine mastitis and the experimental reproduction of the disease. Vet. Rec. 1982, 110, 578–580. [Google Scholar] [CrossRef]
- Krasteva, I.; Liljander, A.; Fischer, A.; Smith, D.G.; Inglis, N.F.; Scacchia, M.; Pini, A.; Jores, J.; Sacchini, F. Characterization of the in vitro core surface proteome of Mycoplasma mycoides subsp. mycoides, the causative agent of contagious bovine pleuropneumonia. Vet. Microbiol. 2014, 168, 116–123. [Google Scholar] [CrossRef]
- Anyansi, C.; Straub, T.J.; Manson, A.L.; Earl, A.M.; Abeel, T. Computational methods for strain-level microbial detection in colony and metagenome sequencing data. Front. Microbiol. 2020, 11, 1925. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host Species | Sample Size | Sequencing Platform | Sample Type | Country | Data Accession | Citation |
|---|---|---|---|---|---|---|
| Cattle | 14 | Illumina HiSeq 2000 | Rumen digesta | Canada | PRJNA448333 | [12] |
| Sheep | 10 | Illumina HiSeq 2000 | Rumen digesta | New Zealand | PRJNA202380 | [13] |
| Human | 13 | Illumina HiSeq 2000 | Feces | USA | PRJNA354235 | [15] |
| Pig | 6 | Illumina NovaSeq | Feces | China | NA | [11] |
| Mice | 8 | Illumina HiSeq 4000 | Gut context | Canada | PRJEB33889 | [14] |
| Chicken | 6 | Illumina NovaSeq | Feces | China | NA | [11] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, Y.; Chen, R.; Zhang, Z.; Jin, R.; Xie, T.; Liu, X.; Chai, J.; Howe, S.; Zhao, J.; Li, Y.; et al. Metagenomic and Meta-Transcriptomic Analysis Reveal the Colonization and Expression Profile of Probiotic Strains in Humans and Animals. Fermentation 2023, 9, 417. https://doi.org/10.3390/fermentation9050417
Peng Y, Chen R, Zhang Z, Jin R, Xie T, Liu X, Chai J, Howe S, Zhao J, Li Y, et al. Metagenomic and Meta-Transcriptomic Analysis Reveal the Colonization and Expression Profile of Probiotic Strains in Humans and Animals. Fermentation. 2023; 9(5):417. https://doi.org/10.3390/fermentation9050417
Chicago/Turabian StylePeng, Yunjuan, Routing Chen, Zhihao Zhang, Rui Jin, Ting Xie, Xinting Liu, Jianmin Chai, Samantha Howe, Jiangchao Zhao, Ying Li, and et al. 2023. "Metagenomic and Meta-Transcriptomic Analysis Reveal the Colonization and Expression Profile of Probiotic Strains in Humans and Animals" Fermentation 9, no. 5: 417. https://doi.org/10.3390/fermentation9050417