
Characterization of a Potential Probiotic Strain in Koumiss

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of LAB from Koumiss
2.2. Extraction of XM-38 Supernatant
2.3. In Vitro Assays of Probiotic Properties
2.3.1. Antibacterial Assay
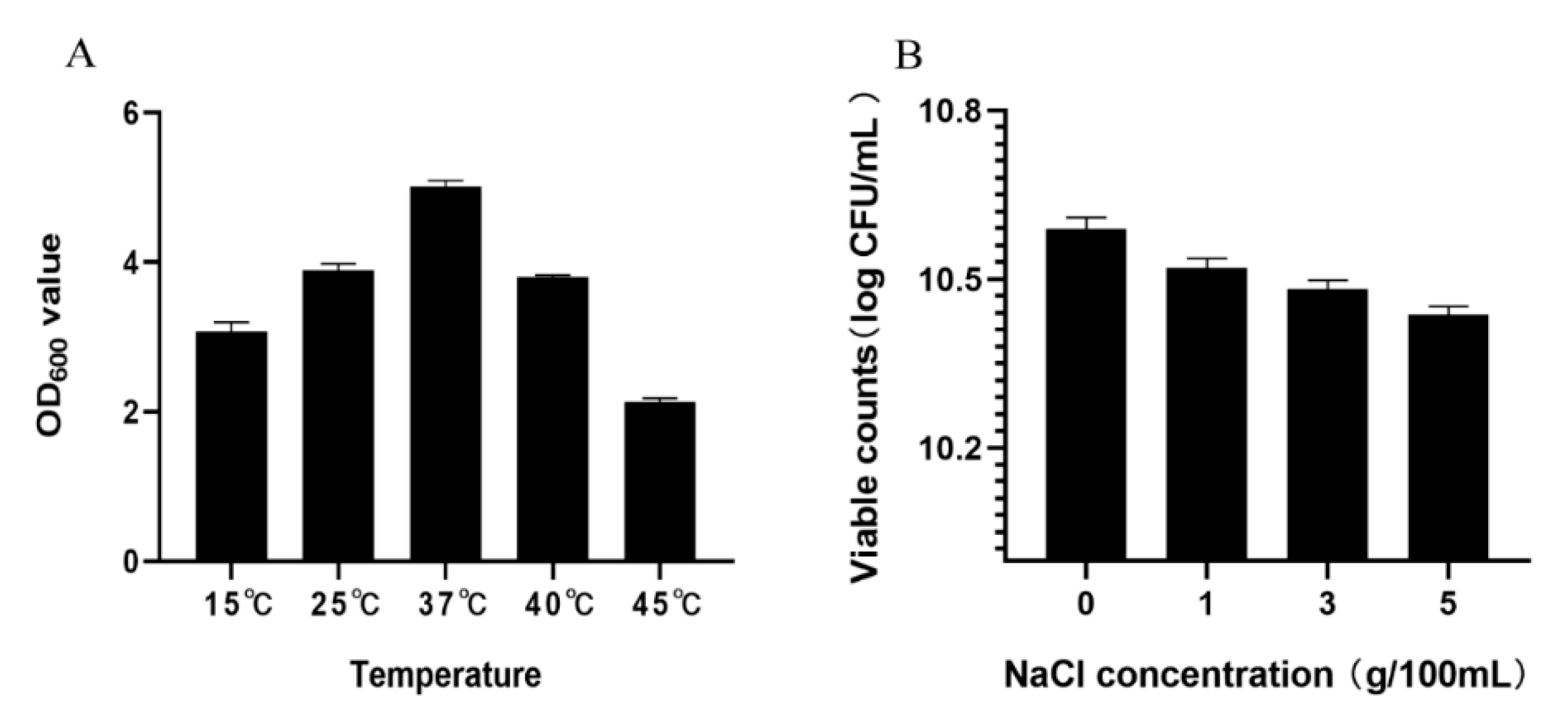
2.3.2. Toleration of Temperature
2.3.3. Toleration of NaCl
2.3.4. Toleration to Acid and Bile Salt
2.3.5. Auto-Aggregation Analysis
2.3.6. Hydrophobicity
2.3.7. Antioxidant Properties
2.3.8. Drug Susceptibility Analysis
2.3.9. Acute Oral Toxicity Test in Mice
2.4. Identification of XM-38
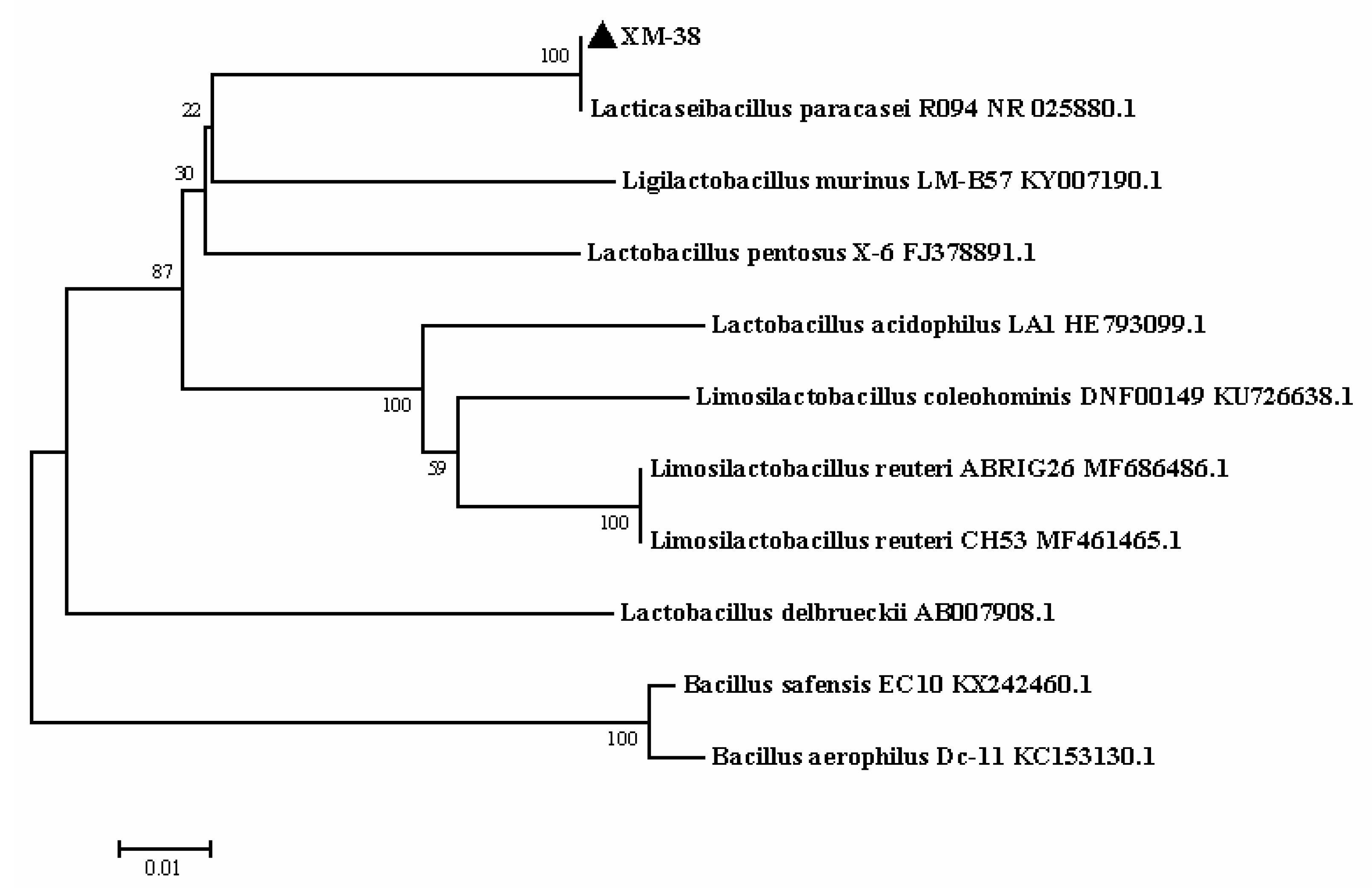
2.4.1. Whole-Genome Extraction, Sequencing, and Construction of Phylogenetic Trees
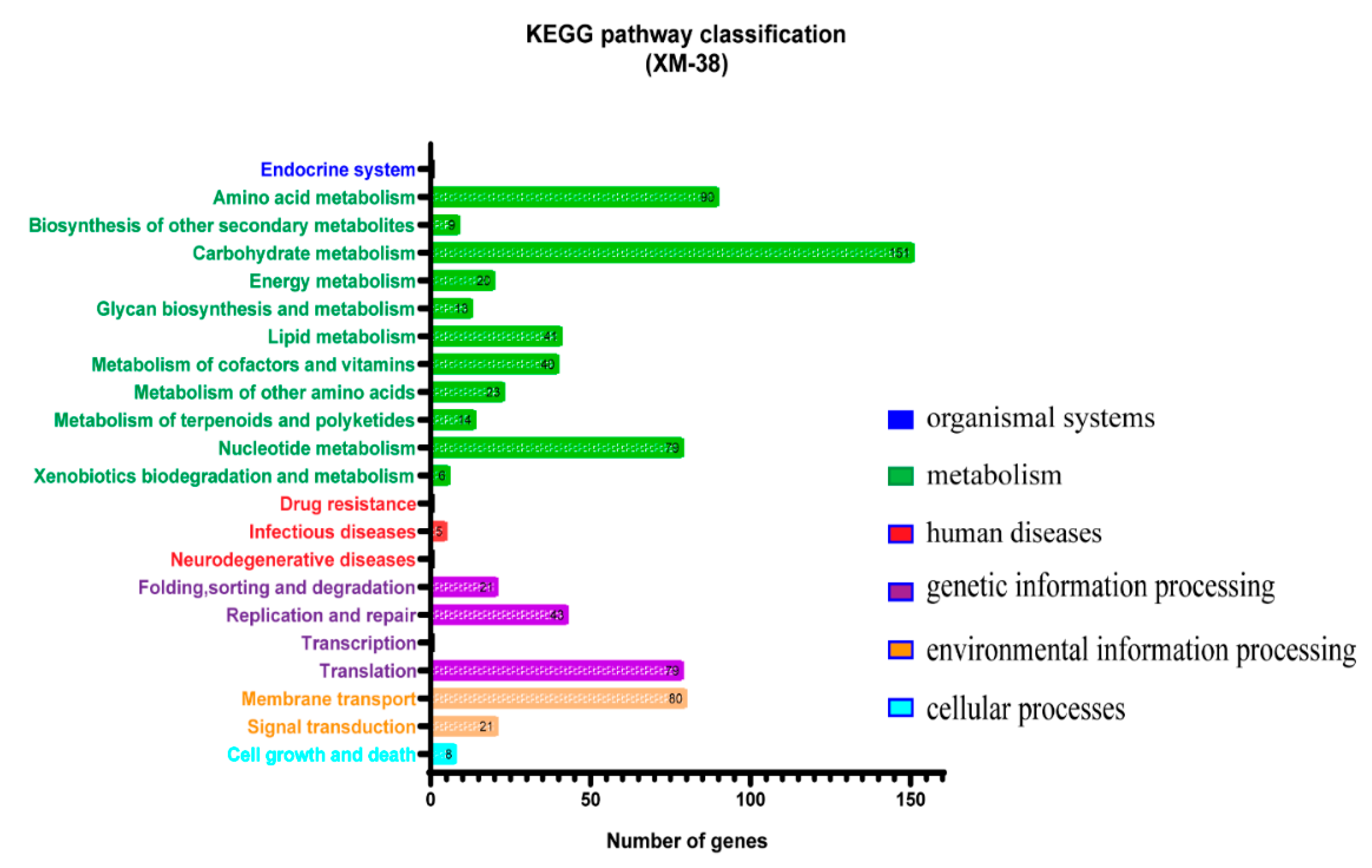
2.4.2. Genome Annotation
2.4.3. Prediction of Bacteriocin Genes
2.4.4. Antibiotic-Resistance Gene
2.4.5. Virulence Gene Prediction
2.4.6. Statistical Analysis
3. Results
3.1. Biological Identification of XM-38
3.1.1. Antibacterial Assay
3.1.2. Toleration to Temperature and NaCl
3.1.3. Toleration to Acid and Bile Salt
3.1.4. Hydrophobicity and Antioxidant Properties
3.1.5. Auto-Aggregation Analysis
3.1.6. Drug Susceptibility Analysis
3.1.7. Acute Oral Toxicity Test in Mice
3.2. Identification of XM-38
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sudun; Wulijideligen; Arakawa, K.; Miyamoto, M.; Miyamoto, T. Interaction between lactic acid bacteria and yeasts in airag, an alcoholic fermented milk. Anim. Sci. J. 2013, 84, 66–74. [Google Scholar] [CrossRef]
- Afzaal, M.; Saeed, F.; Anjum, F.; Waris, N.; Husaain, M.; Ikram, A.; Ateeq, H.; Muhammad Anjum, F.; Suleria, H. Nutritional and ethnomedicinal scenario of koumiss: A concurrent review. Food Sci. Nutr. 2021, 9, 6421–6428. [Google Scholar] [CrossRef] [PubMed]
- Lynch, K.M.; Zannini, E.; Coffey, A.; Arendt, E.K. Lactic Acid Bacteria Exopolysaccharides in Foods and Beverages: Isolation, Properties, Characterization, and Health Benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 155–176. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Huang, J.; Zhou, R. Genomics of lactic acid bacteria: Current status and potential applications. Crit. Rev. Microbiol. 2017, 43, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Park, G.S.; Hong, S.J.; Jung, B.K.; Park, S.; Jin, H.; Lee, S.J.; Shin, J.H.; Lee, H.S. Whole genome sequence of lactic acid bacterium Pediococcus acidilactici strain S1. Braz. J. Microbiol. 2017, 48, 395–396. [Google Scholar] [CrossRef]
- Yang, Y.; An, H.; Zhai, Z.; Wang, G.; Li, J.; Hao, Y. Complete genome sequence of Lactobacillus helveticus CAUH18, a potential probiotic strain originated from koumiss. J. Biotechnol. 2016, 224, 18–19. [Google Scholar] [CrossRef]
- Tarrah, A.; Pakroo, S.; Corich, V.; Giacomini, A. Whole-genome sequence and comparative genome analysis of Lactobacillus paracasei DTA93, a promising probiotic lactic acid bacterium. Arch. Microbiol. 2020, 202, 1997–2003. [Google Scholar] [CrossRef]
- Zhang, J.; Cui, X.; Zhang, M.; Bai, B.; Yang, Y.; Fan, S. The antibacterial mechanism of perilla rosmarinic acid. Biotechnol. Appl. Biochem. 2021, 69, 1757–1764. [Google Scholar] [CrossRef]
- Masuda, M.; Ide, M.; Utsumi, H.; Niiro, T.; Shimamura, Y.; Murata, M. Production potency of folate, vitamin B(12), and thiamine by lactic acid bacteria isolated from Japanese pickles. Biosci. Biotechnol. Biochem. 2012, 76, 2061–2067. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Tian, Y.; Cao, Y.; Li, J.; Guo, H.; Su, Y.; Tian, Y.; Wang, C.; Wang, T.; Zhang, L. Probiotic Properties of Lactobacillus paracasei subsp. paracasei L1 and Its Growth Performance-Promotion in Chicken by Improving the Intestinal Microflora. Front. Physiol. 2019, 10, 937. [Google Scholar] [CrossRef]
- De Albuquerque, T.M.R.; Garcia, E.F.; de Oliveira Araujo, A.; Magnani, M.; Saarela, M.; de Souza, E.L. In Vitro Characterization of Lactobacillus Strains Isolated from Fruit Processing By-Products as Potential Probiotics. Probiotics Antimicrob. Proteins 2018, 10, 704–716. [Google Scholar] [CrossRef]
- Kos, B.; Suskovic, J.; Vukovic, S.; Simpraga, M.; Frece, J.; Matosic, S. Adhesion and aggregation ability of probiotic strain Lactobacillus acidophilus M92. J. Appl. Microbiol. 2003, 94, 981–987. [Google Scholar] [CrossRef] [Green Version]
- Doyle, R.J.; Rosenberg, M. Measurement of microbial adhesion to hydrophobic substrata. Methods Enzymol. 1995, 253, 542–550. [Google Scholar]
- Son, S.; Lewis, B.A. Free radical scavenging and antioxidative activity of caffeic acid amide and ester analogues: Structure-activity relationship. J. Agric. Food Chem. 2002, 50, 468–472. [Google Scholar] [CrossRef]
- Kassim, A.; Omuse, G.; Premji, Z.; Revathi, G. Comparison of Clinical Laboratory Standards Institute and European Committee on Antimicrobial Susceptibility Testing guidelines for the interpretation of antibiotic susceptibility at a University teaching hospital in Nairobi, Kenya: A cross-sectional study. Ann. Clin. Microbiol. Antimicrob. 2016, 15, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selt, F.; Hohloch, J.; Hielscher, T.; Sahm, F.; Capper, D.; Korshunov, A.; Usta, D.; Brabetz, S.; Ridinger, J.; Ecker, J.; et al. Establishment and application of a novel patient-derived KIAA1549:BRAF-driven pediatric pilocytic astrocytoma model for preclinical drug testing. Oncotarget 2017, 8, 11460–11479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OECD. OECD Guidelines for the Testing of Chemicals No. 423: Acute Oral Toxicity—Acute Toxic Class Method; OECD: Paris, France, 1996. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besemer, J.; Lomsadze, A.; Borodovsky, M. GeneMarkS: A self-training method for prediction of gene starts in microbial genomes. Implications for finding sequence motifs in regulatory regions. Nucleic Acids Res. 2001, 29, 2607–2618. [Google Scholar] [CrossRef] [Green Version]
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997, 25, 955–964. [Google Scholar] [CrossRef]
- Lagesen, K.; Hallin, P.; Rodland, E.A.; Staerfeldt, H.H.; Rognes, T.; Ussery, D.W. RNAmmer: Consistent and rapid annotation of ribosomal RNA genes. Nucleic Acids Res. 2007, 35, 3100–3108. [Google Scholar] [CrossRef]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Pop, M. ARDB—Antibiotic Resistance Genes Database. Nucleic Acids Res. 2009, 37, D443–D447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Chen, L.; Xiong, Z.; Sun, L.; Yang, J.; Jin, Q. VFDB 2012 update: Toward the genetic diversity and molecular evolution of bacterial virulence factors. Nucleic Acids Res. 2012, 40, D641–D645. [Google Scholar] [CrossRef]
- O’Shea, E.F.; Cotter, P.D.; Ross, R.P.; Hill, C. Strategies to improve the bacteriocin protection provided by lactic acid bacteria. Curr. Opin. Biotechnol. 2013, 24, 130–134. [Google Scholar] [CrossRef] [Green Version]
- Horgan, M.; O’Flynn, G.; Garry, J.; Cooney, J.; Coffey, A.; Fitzgerald, G.F.; Ross, R.P.; McAuliffe, O. Phage lysin LysK can be truncated to its CHAP domain and retain lytic activity against live antibiotic-resistant staphylococci. Appl. Environ. Microbiol. 2009, 75, 872–874. [Google Scholar] [CrossRef] [Green Version]
- Qureshi, N.; Gu, Q.; Li, P. Whole genome sequence analysis and in vitro probiotic characteristics of a Lactobacillus strain Lactobacillus paracasei ZFM54. J. Appl. Microbiol. 2020, 129, 422–433. [Google Scholar] [CrossRef]
- Koryszewska-Baginska, A.; Gawor, J.; Nowak, A.; Grynberg, M.; Aleksandrzak-Piekarczyk, T. Comparative genomics and functional analysis of a highly adhesive dairy Lactobacillus paracasei subsp. paracasei IBB3423 strain. Appl. Microbiol. Biotechnol. 2019, 103, 7617–7634. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wu, Y.; Wang, Y.; Xu, H.; Mei, X.; Yu, D.; Wang, Y.; Li, W. Antioxidant Properties of Probiotic Bacteria. Nutrients 2017, 9, 521. [Google Scholar] [CrossRef] [Green Version]
- Vlkova, E.; Rada, V.; Smehilova, M.; Killer, J. Auto-aggregation and co-aggregation ability in bifidobacteria and clostridia. Folia Microbiol. 2008, 53, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Valeriano, V.D.; Parungao-Balolong, M.M.; Kang, D.K. In vitro evaluation of the mucin-adhesion ability and probiotic potential of Lactobacillus mucosae LM1. J. Appl. Microbiol. 2014, 117, 485–497. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.M.; Im, D.S. Screening and characterization of probiotic lactic acid bacteria isolated from Korean fermented foods. J. Microbiol. Biotechnol. 2009, 19, 178–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, D.; Li, C.; Qin, Y.; Yin, R.; Du, S.; Ye, F.; Liu, C.; Liu, H.; Wang, M.; Li, Y.; et al. In vitro evaluation of the probiotic and functional potential of Lactobacillus strains isolated from fermented food and human intestine. Anaerobe 2014, 30, 1–10. [Google Scholar] [CrossRef]
- Vizoso Pinto, M.G.; Franz, C.M.; Schillinger, U.; Holzapfel, W.H. Lactobacillus spp. with in vitro probiotic properties from human faeces and traditional fermented products. Int. J. Food Microbiol. 2006, 109, 205–214. [Google Scholar] [CrossRef]
- EFSA Panel. Guidance on the assessment of bacterial susceptibility to antimicrobials of human and veterinary importance. EFSA J. 2012, 10, 2740. [Google Scholar]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277–D280. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Goto, S.; Hattori, M.; Aoki-Kinoshita, K.F.; Itoh, M.; Kawashima, S.; Katayama, T.; Araki, M.; Hirakawa, M. From genomics to chemical genomics: New developments in KEGG. Nucleic Acids Res. 2006, 34, D354–D357. [Google Scholar] [CrossRef]
- McGovern, P.E.; Zhang, J.; Tang, J.; Zhang, Z.; Hall, G.R.; Moreau, R.A.; Nunez, A.; Butrym, E.D.; Richards, M.P.; Wang, C.S.; et al. Fermented beverages of pre- and proto-historic China. Proc. Natl. Acad. Sci. USA 2004, 101, 17593–17598. [Google Scholar] [CrossRef] [Green Version]
- Tamang, J.P.; Shin, D.H.; Jung, S.J.; Chae, S.W. Functional Properties of Microorganisms in Fermented Foods. Front. Microbiol. 2016, 7, 578. [Google Scholar] [CrossRef] [Green Version]
- Cuamatzin-Garcia, L.; Rodriguez-Rugarcia, P.; El-Kassis, E.G.; Galicia, G.; Meza-Jimenez, M.L.; Banos-Lara, M.D.R.; Zaragoza-Maldonado, D.S.; Perez-Armendariz, B. Traditional Fermented Foods and Beverages from around the World and Their Health Benefits. Microorganisms 2022, 10, 1151. [Google Scholar] [CrossRef] [PubMed]
- Mathur, H.; Beresford, T.P.; Cotter, P.D. Health Benefits of Lactic Acid Bacteria (LAB) Fermentates. Nutrients 2020, 12, 1679. [Google Scholar] [CrossRef] [PubMed]
- Mokoena, M.P. Lactic Acid Bacteria and Their Bacteriocins: Classification, Biosynthesis and Applications against Uropathogens: A Mini-Review. Molecules 2017, 22, 1255. [Google Scholar] [CrossRef] [Green Version]
- LeBlanc, J.G.; Levit, R.; Savoy de Giori, G.; de Moreno de LeBlanc, A. Application of vitamin-producing lactic acid bacteria to treat intestinal inflammatory diseases. Appl. Microbiol. Biotechnol. 2020, 104, 3331–3337. [Google Scholar] [CrossRef] [PubMed]
- De Boeck, I.; Spacova, I.; Vanderveken, O.M.; Lebeer, S. Lactic acid bacteria as probiotics for the nose? Microb. Biotechnol. 2021, 14, 859–869. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, F.M.; Silva, H.L.A.; Poso, S.M.V.; Barroso, M.V.; Lanzetti, M.; Rocha, R.S.; Graca, J.S.; Esmerino, E.A.; Freitas, M.Q.; Silva, M.C.; et al. Probiotic Prato cheese attenuates cigarette smoke-induced injuries in mice. Food Res. Int. 2019, 123, 697–703. [Google Scholar] [CrossRef]
- Moradi, M.; Kousheh, S.A.; Almasi, H.; Alizadeh, A.; Guimaraes, J.T.; Yilmaz, N.; Lotfi, A. Postbiotics produced by lactic acid bacteria: The next frontier in food safety. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3390–3415. [Google Scholar] [CrossRef]
- Dowarah, R.; Verma, A.K.; Agarwal, N.; Singh, P.; Singh, B.R. Selection and characterization of probiotic lactic acid bacteria and its impact on growth, nutrient digestibility, health and antioxidant status in weaned piglets. PLoS ONE 2018, 13, e0192978. [Google Scholar] [CrossRef] [Green Version]
- Ndahetuye, J.B.; Koo, O.K.; O’Bryan, C.A.; Ricke, S.C.; Crandall, P.G. Role of lactic acid bacteria as a biosanitizer to prevent attachment of Listeria monocytogenes F6900 on deli slicer contact surfaces. J. Food Prot. 2012, 75, 1429–1436. [Google Scholar] [CrossRef]
- Liu, Z.; Xu, C.; Tian, R.; Wang, W.; Ma, J.; Gu, L.; Liu, F.; Jiang, Z.; Hou, J. Screening beneficial bacteriostatic lactic acid bacteria in the intestine and studies of bacteriostatic substances. J. Zhejiang Univ. Sci. B 2021, 22, 533–547. [Google Scholar] [CrossRef]
- Adetoye, A.; Pinloche, E.; Adeniyi, B.A.; Ayeni, F.A. Characterization and anti-salmonella activities of lactic acid bacteria isolated from cattle faeces. BMC Microbiol. 2018, 18, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bateman, A.; Rawlings, N.D. The CHAP domain: A large family of amidases including GSP amidase and peptidoglycan hydrolases. Trends Biochem. Sci. 2003, 28, 234–237. [Google Scholar] [CrossRef] [PubMed]
- Becker, S.C.; Swift, S.; Korobova, O.; Schischkova, N.; Kopylov, P.; Donovan, D.M.; Abaev, I. Lytic activity of the staphylolytic Twort phage endolysin CHAP domain is enhanced by the SH3b cell wall binding domain. FEMS Microbiol. Lett. 2015, 362, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibbs, G.M.; Davidson, B.E.; Hillier, A.J. Novel expression system for large-scale production and purification of recombinant class IIa bacteriocins and its application to piscicolin 126. Appl. Environ. Microbiol. 2004, 70, 3292–3297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarate, G.; Chaia, A.P.; Gonzalez, S.; Oliver, G. Viability and beta-galactosidase activity of dairy propionibacteria subjected to digestion by artificial gastric and intestinal fluids. J. Food Prot. 2000, 63, 1214–1221. [Google Scholar] [CrossRef] [PubMed]
- Ku, S.; Park, M.S.; Ji, G.E.; You, H.J. Review on Bifidobacterium bifidum BGN4: Functionality and Nutraceutical Applications as a Probiotic Microorganism. Int. J. Mol. Sci. 2016, 17, 1544. [Google Scholar] [CrossRef] [Green Version]
- Dunne, C.; O’Mahony, L.; Murphy, L.; Thornton, G.; Morrissey, D.; O’Halloran, S.; Feeney, M.; Flynn, S.; Fitzgerald, G.; Daly, C.; et al. In vitro selection criteria for probiotic bacteria of human origin: Correlation with in vivo findings. Am. J. Clin. Nutr. 2001, 73 (Suppl. 2), 386S–392S. [Google Scholar] [CrossRef] [Green Version]
- Pan, W.H.; Li, P.L.; Liu, Z. The correlation between surface hydrophobicity and adherence of Bifidobacterium strains from centenarians’ faeces. Anaerobe 2006, 12, 148–152. [Google Scholar] [CrossRef]
- Al Kassaa, I.; Hamze, M.; Hober, D.; Chihib, N.E.; Drider, D. Identification of vaginal lactobacilli with potential probiotic properties isolated from women in North Lebanon. Microb. Ecol. 2014, 67, 722–734. [Google Scholar] [CrossRef]
- Romero-Luna, H.E.; Peredo-Lovillo, A.; Hernandez-Mendoza, A.; Hernandez-Sanchez, H.; Cauich-Sanchez, P.I.; Ribas-Aparicio, R.M.; Davila-Ortiz, G. Probiotic Potential of Lactobacillus paracasei CT12 Isolated from Water Kefir Grains (Tibicos). Curr. Microbiol. 2020, 77, 2584–2592. [Google Scholar] [CrossRef]
- Kerry, R.G.; Patra, J.K.; Gouda, S.; Park, Y.; Shin, H.S.; Das, G. Benefaction of probiotics for human health: A review. J. Food Drug Anal. 2018, 26, 927–939. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Indicator Strains | G+/G− | Inhibition Zone (mm) |
|---|---|---|
| Listeria monocytogenes | G+ | 23.30 ± 0.75 |
| Staphylococcus aureus | G+ | 16.20 ± 0.02 |
| Salmonella typhimurium | G− | 20.11 ± 0.75 |
| Escherichia coli | G− | 21.00 ± 0.02 |
| Shigella | G− | 17.00 ± 0.02 |
| pH | Growth of Strain XM-38 (log CFU·mL−1) | |||
|---|---|---|---|---|
| 0 h | 1 h | 2 h | 3 h | |
| 6.2 (control) | 6.80 ± 0.50 a | 7.62 ± 0.41 b | 8.07 ± 0.58 b | 8.23 ± 0.29 b |
| 5.5 | 6.79 ± 0.36 a | 6.87 ± 0.47 a | 6.92 ± 0.30 a | 6.98 ± 0.35 a |
| 4.5 | 6.75 ± 0.43 a | 6.74 ± 0.49 a | 6.75 ± 0.22 a | 6.77 ± 0.27 a |
| 3.5 | 6.67 ± 0.39 a | 6.72 ± 0.34 a | 6.72 ± 0.09 a | 6.73 ± 0.29 a |
| 2.5 | 6.65 ± 0.47 a | 6.68 ± 0.57 a | 6.69 ± 0.49 a | 6.69 ± 0.15 a |
| Bile Salt Concentration (%) | Growth of Strain XM-38 (log CFU·mL−1) | ||||
|---|---|---|---|---|---|
| 0 h | 1 h | 2 h | 3 h | 4 h | |
| 0 (control) | 6.75 ± 0.21 a | 7.57 ± 0.14 b | 8.13 ± 0.32 b | 8.25 ± 0.30 b | 8.64 ± 0.08 b |
| 0.1 | 6.72 ± 0.35 a | 6.69 ± 0.40 a | 6.78 ± 0.27 a | 6.82 ± 0.52 a | 6.76 ± 0.54 a |
| 0.2 | 6.71 ± 0.24 a | 6.72 ± 0.20 a | 6.74 ± 0.33 a | 6.77 ± 0.38 a | 6.76 ± 0.10 a |
| 0.3 | 6.67 ± 0.30 a | 6.67 ± 0.46 a | 6.62 ± 0.34 a | 6.58 ± 0.45 a | 6.62 ± 0.43 a |
| Gene id | Identity | Min Identity | Antibiotic Resistance |
|---|---|---|---|
| XM38_GM000138 | 42.4 | 80 | vancomycin |
| XM38_GM000679 | 46.3 | 80 | deoxycholate, fosfomycin |
| XM38_GM000778 | 44.4 | 80 | lincomycin |
| XM38_GM002177 | 40 | 80 | vancomycin, teicoplanin |
| XM38_GM002210 | 62.3 | 30 | bacitracin |
| XM38_GM002581 | 53.3 | 80 | lincosamide, streptogramin B, macrolide |
| XM38_GM002903 | 45.6 | 80 | vancomycin |
| XM38_GM003005 | 41.6 | 80 | bacitracin |
| Gene Id | VFDB Name | Identity (%) | Length (bp) | Location |
|---|---|---|---|---|
| XM38_GM000099 | Trehalose-recycling ABC transporter | 40.8 | 741 | 96,094–96,834 |
| XM38_GM000229 | Trehalose-recycling ABC transporter | 40.1 | 960 | 219,268–220,227 |
| XM38_GM000668 | Mg2+ transport | 45.3 | 2118 | 613,904–616,021 |
| XM38_GM000821 | FbpABC | 40.9 | 1224 | 769,042–770,265 |
| XM38_GM000857 | LPS | 47.7 | 441 | 801,052–801,492 |
| XM38_GM000900 | Oligopeptide-binding protein | 41.2 | 1512 | 844,024–845,535 |
| XM38_GM000925 | Capsular polysaccharide | 48.1 | 573 | 865,883–866,455 |
| XM38_GM000936 | ClpE | 48.5 | 2151 | 874,820–876,970 |
| XM38_GM001159 | Iron-cofactored SOD | 41.7 | 618 | 1,060,994–1,061,611 |
| XM38_GM001180 | Ferrous iron transport | 40 | 264 | 1,085,394–1,085,657 |
| Genome Size (bp) | Gene Number | Gene Length (bp) | GC Content (%) | tRNA | rRNA |
|---|---|---|---|---|---|
| 2,910,589 | 3118 | 2,495,028 | 47.11 | 59 | 15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, H.-Z.; Song, Q.-J.; Guo, H.; Liu, C.-Y.; Yang, C.; Li, X.; Wang, Y.-X.; Ma, Z.-P.; Wang, F.-X.; Wen, Y.-J. Characterization of a Potential Probiotic Strain in Koumiss. Fermentation 2023, 9, 87. https://doi.org/10.3390/fermentation9020087
Zhao H-Z, Song Q-J, Guo H, Liu C-Y, Yang C, Li X, Wang Y-X, Ma Z-P, Wang F-X, Wen Y-J. Characterization of a Potential Probiotic Strain in Koumiss. Fermentation. 2023; 9(2):87. https://doi.org/10.3390/fermentation9020087
Chicago/Turabian StyleZhao, Hong-Zhe, Qian-Jin Song, Hao Guo, Chun-Yu Liu, Chao Yang, Xin Li, Ya-Xin Wang, Zi-Peng Ma, Feng-Xue Wang, and Yong-Jun Wen. 2023. "Characterization of a Potential Probiotic Strain in Koumiss" Fermentation 9, no. 2: 87. https://doi.org/10.3390/fermentation9020087