
Isolation of Lactococcus sp. X1 from Termite Gut, and Its Application in Lactic Acid Production

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Media Preparation
2.2. Isolation Procedures
2.3. Identification of Strain X1
2.4. Effects of Different Factors on LA Production
2.5. Simultaneous Saccharification and Fermentation (SSF) of Corncob
2.6. Chemical Analysis
3. Results
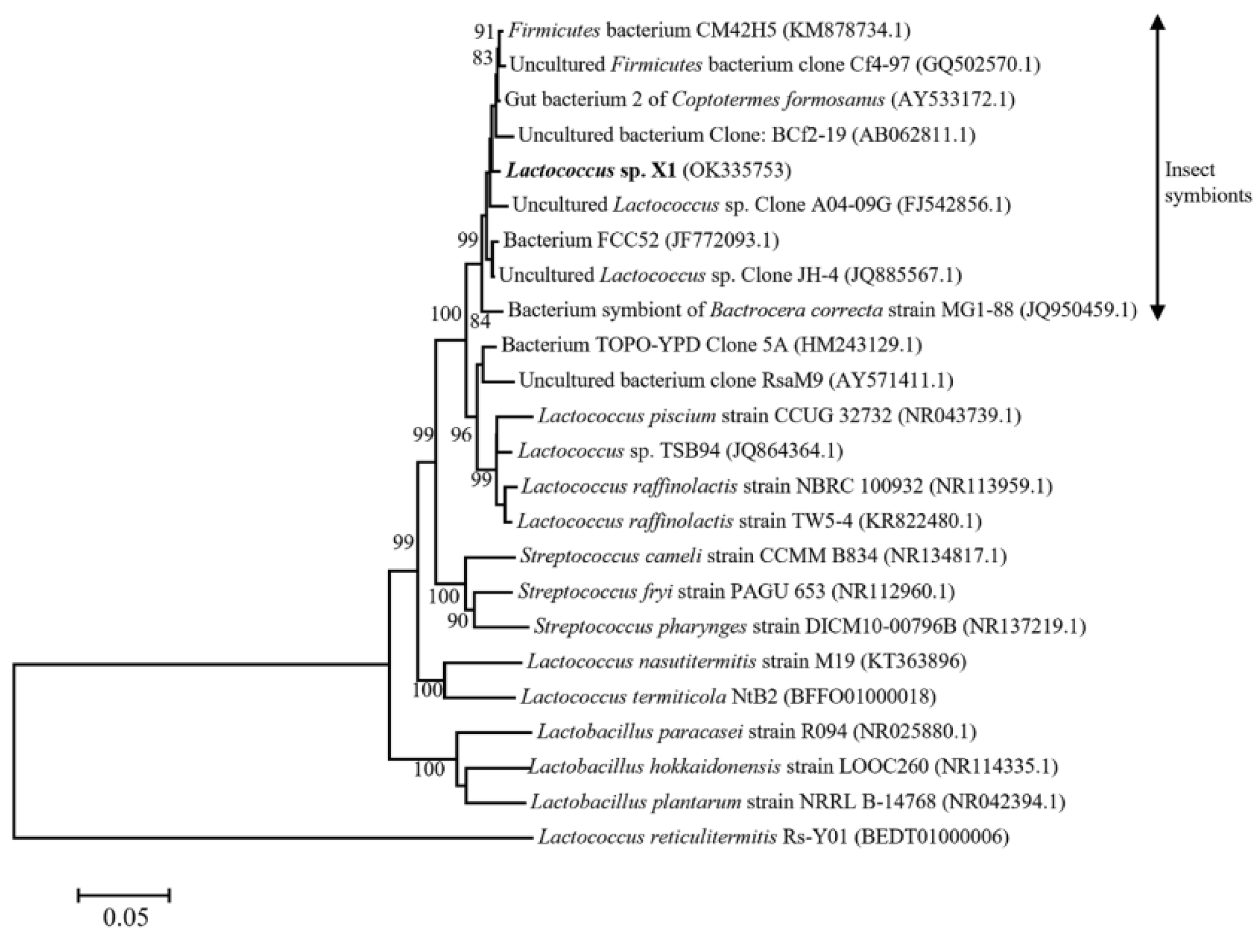
3.1. Identification of Strain X1
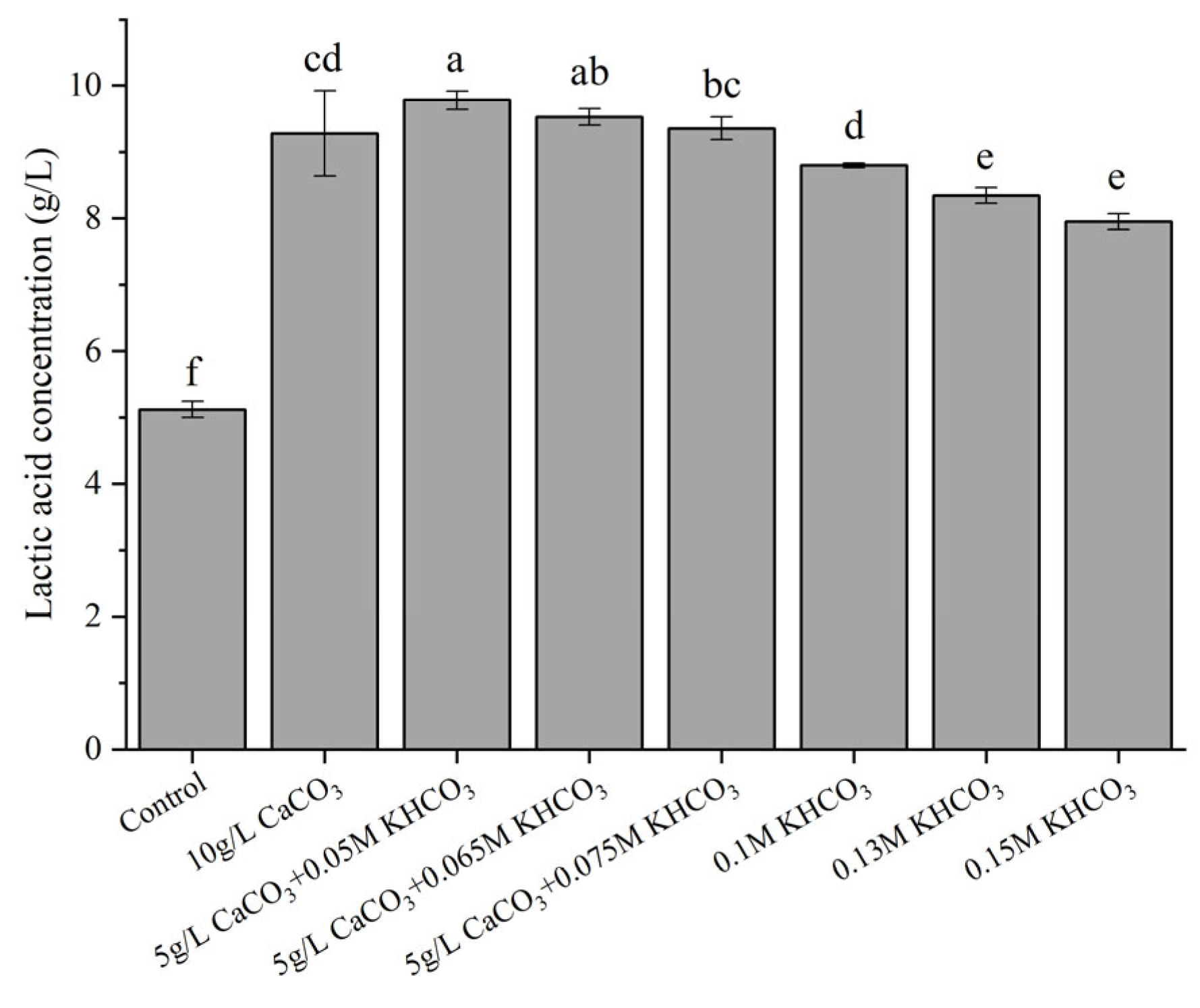
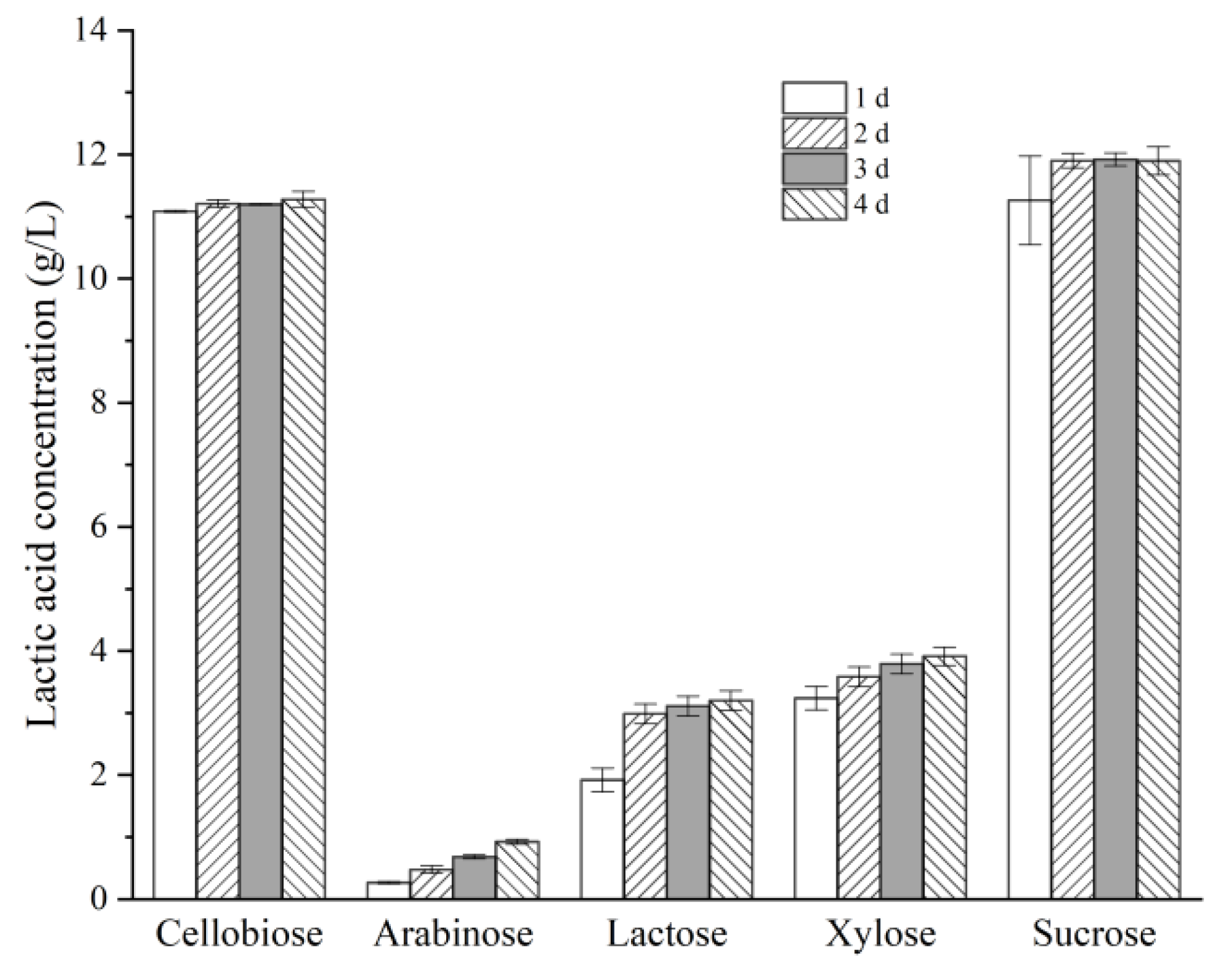
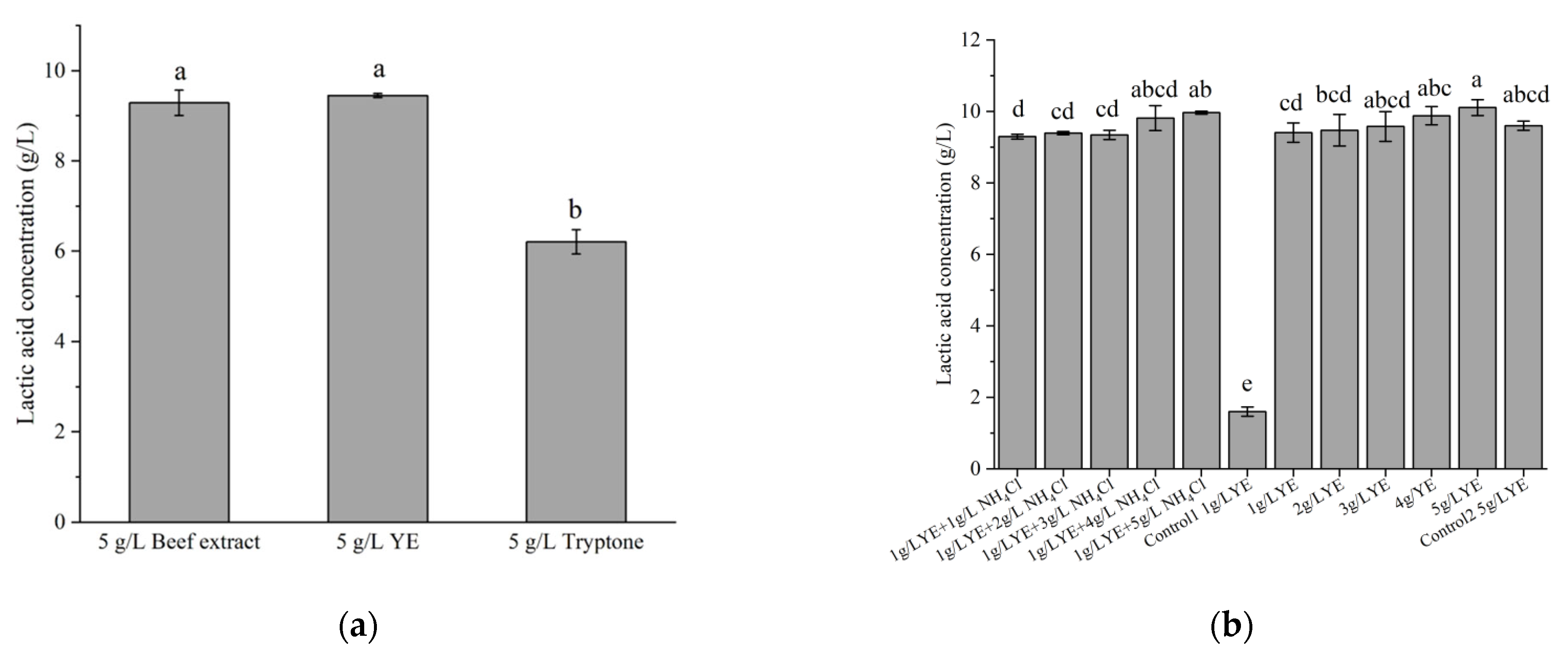
3.2. Effects of Different Factors on LA Production by Strain X1
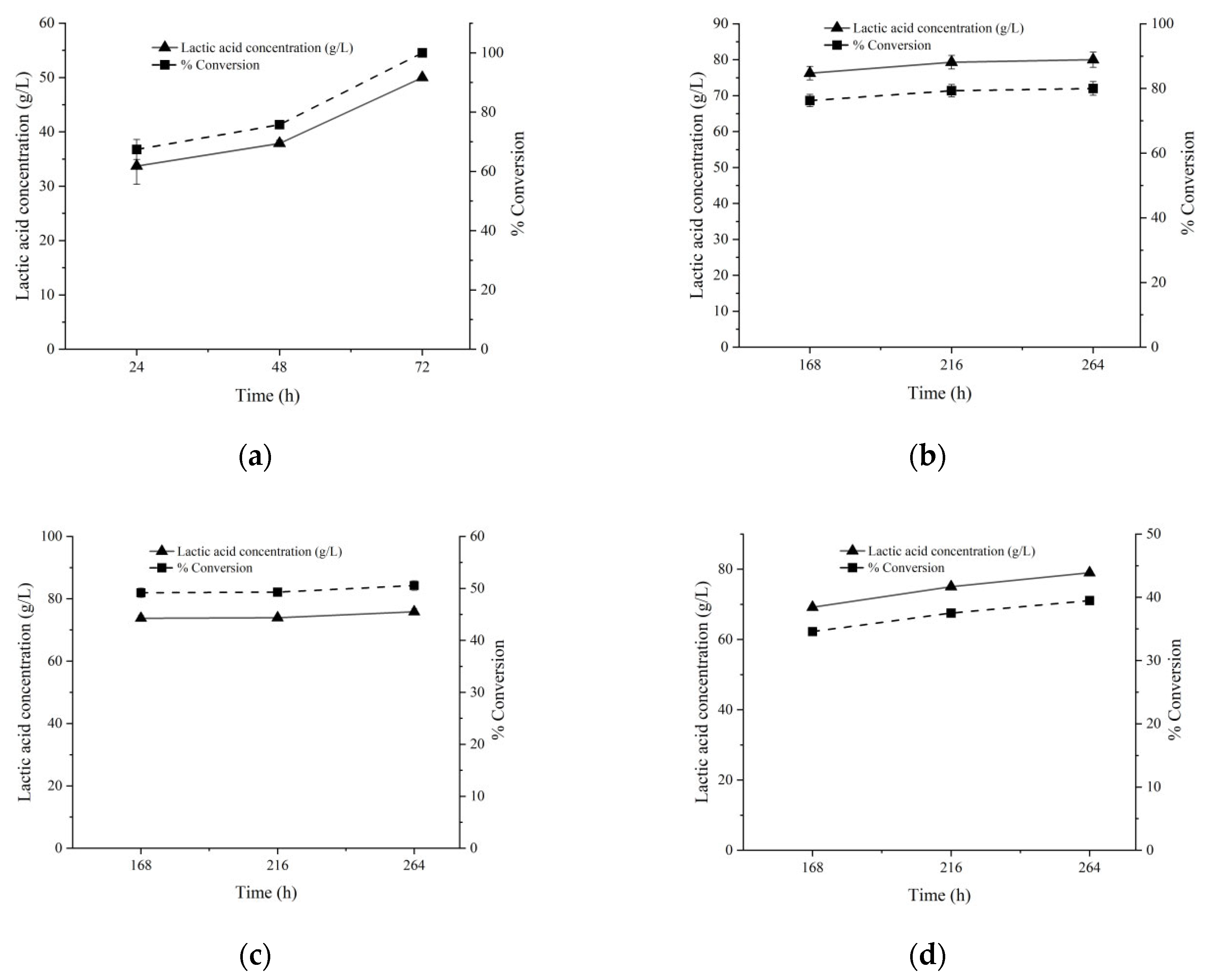
3.3. Fermentation of High Concentration Substrates
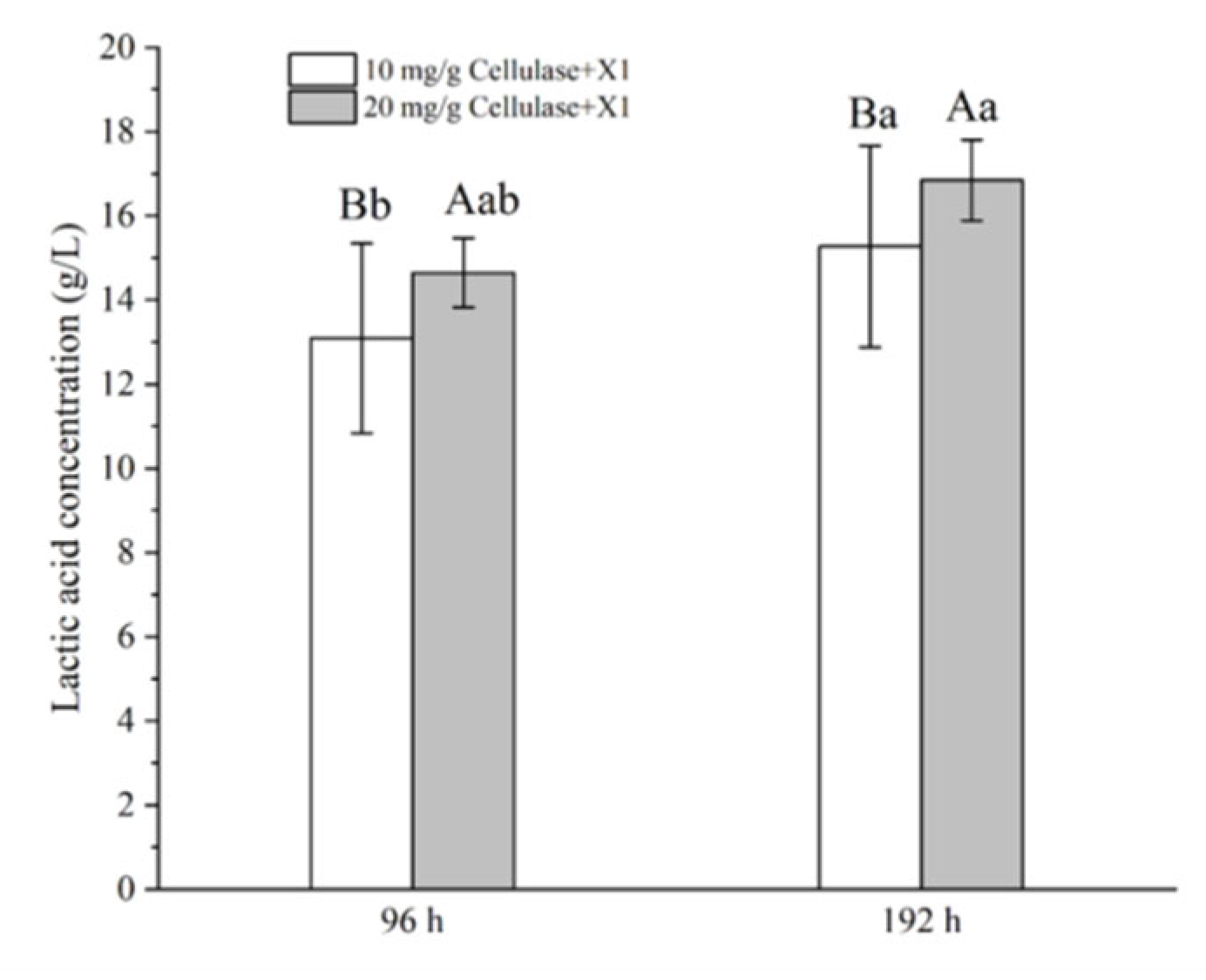
3.4. SSF of Corncob for LA Production
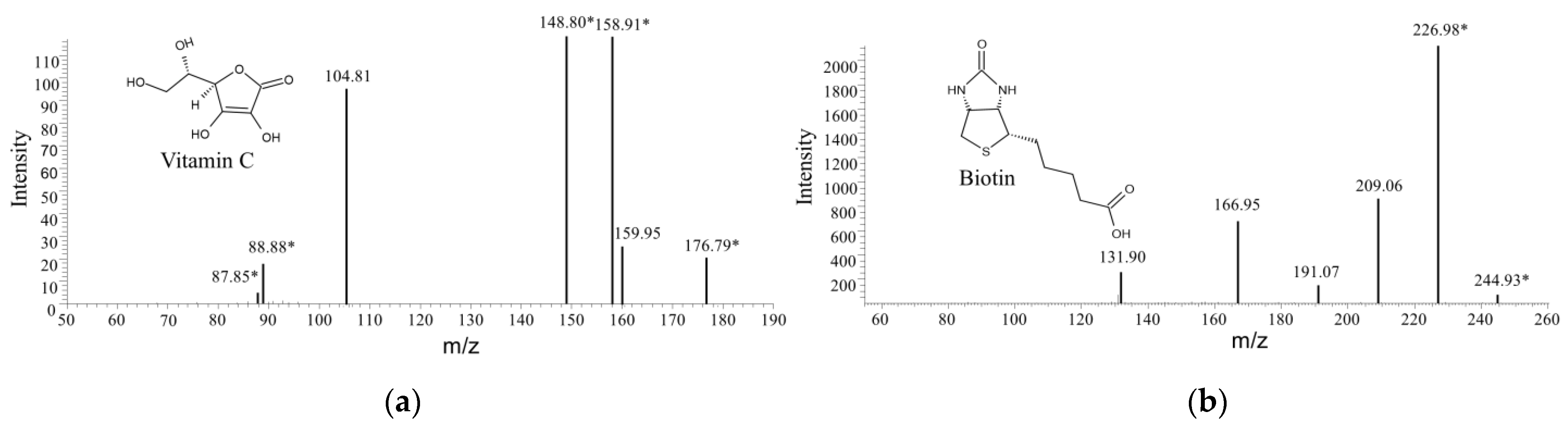
3.5. Vitamin Identification
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rawoof, S.A.A.; Kumar, P.S.; Vo, D.-V.N.; Devaraj, K.; Mani, Y.; Devaraj, T.; Subramanian, S. Production of optically pure lactic acid by microbial fermentation: A review. Environ. Chem. Lett. 2021, 19, 539–556. [Google Scholar] [CrossRef]
- Wang, Y.; Tashiro, Y.; Sonomoto, K. Fermentative production of lactic acid from renewable materials: Recent achievements, prospects, and limits. J. Biosci. Bioeng. 2015, 119, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Murakami, N.; Oba, M.; Iwamoto, M.; Tashiro, Y.; Noguchi, T.; Bonkohara, K.; Abdel-Rahman, M.A.; Zendo, T.; Shimoda, M.; Sakai, K.; et al. L-Lactic acid production from glycerol coupled with acetic acid metabolism by Enterococcus faecalis without carbon loss. J. Biosci. Bioeng. 2016, 121, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Singhvi, M.; Zendo, T.; Sonomoto, K. Free lactic acid production under acidic conditions by lactic acid bacteria strains: Challenges and future prospects. Appl. Microbiol. Biotechnol. 2018, 102, 5911–5924. [Google Scholar] [CrossRef] [PubMed]
- Raman, J.; Kim, J.-S.; Choi, K.R.; Eun, H.; Yang, D.; Ko, Y.-J.; Kim, S.-J. Application of lactic acid bacteria (LAB) in sustainable agriculture: Advantages and limitations. Int. J. Mol. Sci. 2022, 23, 7784. [Google Scholar] [CrossRef]
- Park, M.R.; Gauttam, R.; Fong, B.; Chen, Y.; Lim, H.G.; Feist, A.M.; Mukhopadhyay, A.; Petzold, C.J.; Simmons, B.A.; Singer, S.W. Revealing oxidative pentose metabolism in new Pseudomonas putida isolates. Environ. Microbiol. 2022. [Google Scholar] [CrossRef]
- Suman, S.K.; Dhawaria, M.; Tripathi, D.; Raturi, V.; Adhikari, D.K.; Kanaujia, P.K. Investigation of lignin biodegradation by Trabulsiella sp. isolated from termite gut. Int. Biodeterior. Biodegrad. 2016, 112, 12–17. [Google Scholar] [CrossRef]
- Tsegaye, B.; Balomajumder, C.; Roy, P. Isolation and characterization of novel lignolytic, cellulolytic, and hemicellulolytic bacteria from wood-feeding termite Cryptotermes brevis. Int. Microbiol. 2019, 22, 29–39. [Google Scholar] [CrossRef]
- Noda, S.; Sakamoto, M.; Aihara, C.; Yuki, M.; Katsuhara, M.; Ohkuma, M. Lactococcus termiticola sp. nov., isolated from the gut of the wood-feeding higher termite Nasutitermes takasagoensis. Int. J. Syst. Evol. Microbiol. 2018, 68, 3832–3836. [Google Scholar] [CrossRef]
- Higashiguchi, D.T.; Husseneder, C.; Grace, J.K.; Berestecky, J.M. Pilibacter termitis gen. nov., sp. nov., a lactic acid bacterium from the hindgut of the Formosan subterranean termite (Coptotermes formosanus). Int. J. Syst. Evol. Microbiol. 2006, 56, 15–20. [Google Scholar] [CrossRef]
- Yang, S.Y.; Zheng, Y.; Huang, Z.; Wang, X.M.; Yang, H. Lactococcus nasutitermitis sp. nov. isolated from a termite gut. Int. J. Syst. Evol. Microbiol. 2016, 66, 518–522. [Google Scholar] [CrossRef]
- Yuki, M.; Sakamoto, M.; Nishimura, Y.; Ohkuma, M. Lactococcus reticulitermitis sp. nov., isolated from the gut of the subterranean termite Reticulitermes speratus. Int. J. Syst. Evol. Microbiol. 2018, 68, 596–601. [Google Scholar] [CrossRef]
- Deng, H.; Zhu, J.; Tong, Y.; Kong, Y.; Tan, C.; Wang, M.; Wan, M.; Meng, X. Antibacterial characteristics and mechanisms of action of Aronia melanocarpa anthocyanins against Escherichia coli. LWT-Food Sci. Technol. 2021, 150, 112018. [Google Scholar] [CrossRef]
- Tajabadi, N.; Mardan, M.; Abdul Manap, M.Y.; Shuhaimi, M.; Meimandipour, A.; Nateghi, L.J.A. Detection and identification of Lactobacillus bacteria found in the honey stomach of the giant honeybee Apis dorsata. Apidologie 2011, 42, 642–649. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Li, X.; Yong, Q.; Yang, S.-T.; Ouyang, J.; Yu, S. Simultaneous saccharification and fermentation of xylo-oligosaccharides manufacturing waste residue for L-lactic acid production by Rhizopus oryzae. Biochem. Eng. J. 2015, 94, 92–99. [Google Scholar] [CrossRef]
- Geng, A.; Wu, J.; Xie, R.; Wang, H.; Wu, Y.; Li, X.; Chang, F.; Sun, J. Highly thermostable GH51 α-arabinofuranosidase from Hungateiclostridium clariflavum DSM 19732. Appl. Microbiol. Biotechnol. 2019, 103, 3783–3793. [Google Scholar] [CrossRef]
- Zhang, C.; Pan, S.; Yu, X. Simultaneous determination of eight water-soluble vitamins in formula food for special medical purpose using ultra performance liquid chromatography-tandem mass spectrometry. Mod. Food 2021, 27, 164–167. [Google Scholar]
- Heo, J.; Cho, H.; Tamura, T.; Saitou, S.; Park, K.; Kim, J.S.; Hong, S.B.; Kwon, S.W.; Kim, S.J. Lactococcus allomyrinae sp. nov., isolated from gut of larvae of Allomyrina dichotoma. Int. J. Syst. Evol. Microbiol. 2019, 69, 3682–3688. [Google Scholar] [CrossRef]
- Tikhe, C.V.; Sethi, A.; Delatte, J.; Husseneder, C. Isolation and assessment of gut bacteria from the Formosan subterranean termite, Coptotermes formosanus (Isoptera: Rhinotermitidae), for paratransgenesis research and application. Insect Sci. 2017, 24, 93–102. [Google Scholar] [CrossRef]
- Burgess, C.; O’Connell-Motherway, M.; Sybesma, W.; Hugenholtz, J.; van Sinderen, D. Riboflavin production in Lactococcus lactis: Potential for in situ production of vitamin-enriched foods. Appl. Environ. Microbiol. 2004, 70, 5769–5777. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Van Bennekom, E.O.; Zhang, Y.; Abee, T.; Smid, E.J. Long-chain vitamin K2 production in Lactococcus lactis is influenced by temperature, carbon source, aeration and mode of energy metabolism. Microb. Cell Fact. 2019, 18, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gangadharan, D.; Nampoothiri, K.M. Folate production using Lactococcus lactis ssp cremoris with implications for fortification of skim milk and fruit juices. Lwt-Food Sci. Technol. 2011, 44, 1859–1864. [Google Scholar] [CrossRef]
- Jung, M.Y.; Lee, C.; Seo, M.J.; Roh, S.W.; Lee, S.H. Characterization of a potential probiotic bacterium Lactococcus raffinolactis WiKim0068 isolated from fermented vegetable using genomic and in vitro analyses. BMC Microbiol. 2020, 20, 136. [Google Scholar] [CrossRef] [PubMed]
- Wischral, D.; Arias, J.M.; Modesto, L.F.; de Franca Passos, D.; Pereira, N., Jr. Lactic acid production from sugarcane bagasse hydrolysates by Lactobacillus pentosus: Integrating xylose and glucose fermentation. Biotechnol. Prog. 2019, 35, e2718. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Yang, J.; Pang, H.; Kitahara, M. Lactococcus fujiensis sp. nov. A lactic acid bacterium isolated from vegetable matter. Int. J. Syst. Evol. Microbiol. 2011, 61, 1590–1594. [Google Scholar] [CrossRef] [Green Version]
- Passerini, D.; Coddeville, M.; Le Bourgeois, P.; Loubiere, P.; Ritzenthaler, P.; Fontagne-Faucher, C.; Daveran-Mingot, M.L.; Cocaign-Bousquet, M. The carbohydrate metabolism signature of Lactococcus lactis strain A12 reveals its sourdough ecosystem origin. Appl. Environ. Microbiol. 2013, 79, 5844–5852. [Google Scholar] [CrossRef] [Green Version]
- Heo, J.M.; Kim, H.J.; Lee, S.J. Efficient anaerobic consumption of D-xylose by E. coli BL21(DE3) via xylR adaptive mutation. BMC Microbiol. 2021, 21, 332. [Google Scholar] [CrossRef]
- Åkerberg, C.; Hofvendahl, K.; Zacchi, G.; Hahn-Hägerdal, B. Modelling the influence of pH, temperature, glucose and lactic acid concentrations on the kinetics of lactic acid production by Lactococcus lactis ssp. lactis ATCC 19435 in whole-wheat flour. Appl. Microbiol. Biotechnol. 1998, 49, 682–690. [Google Scholar] [CrossRef]
- Okano, K.; Zhang, Q.; Shinkawa, S.; Yoshida, S.; Tanaka, T.; Fukuda, H.; Kondo, A. Efficient production of optically pure D-lactic acid from raw corn starch by using a genetically modified L-lactate dehydrogenase gene-deficient and α-amylase-secreting Lactobacillus plantarum strain. Appl. Environ. Microbiol. 2009, 75, 462–467. [Google Scholar] [CrossRef] [Green Version]
- Bustos, G.; Moldes, A.B.; Alonso, J.L.; Vázquez, M. Optimization of D-lactic acid production by Lactobacillus coryniformis using response surface methodology. Food Microbiol. 2004, 21, 143–148. [Google Scholar] [CrossRef]
- Adsul, M.; Khire, J.; Bastawde, K.; Gokhale, D. Production of lactic acid from cellobiose and cellotriose by Lactobacillus delbrueckii mutant Uc-3. Appl. Environ. Microbiol. 2007, 73, 5055–5057. [Google Scholar] [CrossRef] [Green Version]
- Singhvi, M.; Joshi, D.; Adsul, M.; Varma, A.; Gokhale, D. D-(−)-Lactic acid production from cellobiose and cellulose by Lactobacillus lactis mutant RM2-2 4. Green Chem. 2010, 12, 1106–1109. [Google Scholar] [CrossRef]
- Bischoff, K.M.; Liu, S.; Hughes, S.R.; Rich, J.O. Fermentation of corn fiber hydrolysate to lactic acid by the moderate thermophile Bacillus coagulans. Biotechnol. Lett. 2010, 32, 823–828. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, X.; Luo, J.; Qi, B.; Wan, Y. An efficient process for lactic acid production from wheat straw by a newly isolated Bacillus coagulans strain IPE22. Bioresour. Technol. 2014, 158, 396–399. [Google Scholar] [CrossRef]
- Hu, J.; Zhang, Z.; Lin, Y.; Zhao, S.; Mei, Y.; Liang, Y.; Peng, N. High-titer lactic acid production from NaOH-pretreated corn stover by Bacillus coagulans LA204 using fed-batch simultaneous saccharification and fermentation under non-sterile condition. Bioresour. Technol. 2015, 182, 251–257. [Google Scholar] [CrossRef]
- Tanaka, T.; Hoshina, M.; Tanabe, S.; Sakai, K.; Ohtsubo, S.; Taniguchi, M. Production of D-lactic acid from defatted rice bran by simultaneous saccharification and fermentation. Bioresour. Technol. 2006, 97, 211–217. [Google Scholar] [CrossRef]
- Laopaiboon, P.; Thani, A.; Leelavatcharamas, V.; Laopaiboon, L. Acid hydrolysis of sugarcane bagasse for lactic acid production. Bioresour. Technol. 2010, 101, 1036–1043. [Google Scholar] [CrossRef]
- Karnaouri, A.; Asimakopoulou, G.; Kalogiannis, K.G.; Lappas, A.; Topakas, E. Efficient D-lactic acid production by Lactobacillus delbrueckii subsp. bulgaricus through conversion of organosolv pretreated lignocellulosic biomass. Biomass Bioenerg. 2020, 140, 105672. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Carbon Sources (g/L) | Yields (g/g) | Maximum Theoretical Yield (g/g) | D, L Type | Optical Purity (%) | References |
|---|---|---|---|---|---|---|
| L. plantarum NCIMB 8826 | Glucose (100) | 0.89 | 1.00 | L | 99.6 | [30] |
| L. pentosus ATCC 8041 | Xylose (20) | 0.94 | 1.00 | N.A. a | N.A. a | [25] |
| L. lactis ssp. lactis ATCC 19435 | Glucose (182) | 0.47 | 1.00 | L | 99.0 | [29] |
| L. coryniformis ATCC 25600 | Glucose (100) | 0.59 | 1.00 | D | 99.0 | [31] |
| Lactococcus sp. X1 | Glucose (50) Sucrose (100) Xylose (10) | 0.99 0.80 0.39 | 1.00 1.05 0.60 b | L | 95.1 | This work |
| Strains | Substrates | Pretreatment | Fermentation | Yields (g/g) | References |
|---|---|---|---|---|---|
| B. coagulans strain IPE22 | Wheat straw | Sulfuric acid | SSF | 0.46 | [35] |
| B. coagulans LA204 | Corn stover | NaOH | Fed-batch SSF | 0.68 | [36] |
| B. coagulans MXL-9 | Corn fiber hydrolysates | Sulfuric acid or alkaline H2O2 | SHF | 0.39 | [34] |
| Lb. delbrueckii subsp. delbrueckii IFO 3202 | Defatted rice bran | / | SSF | 0.28 | [37] |
| Lc. lactis IO-1 | Sugar cane baggase | HCl | Batch | 0.36 | [38] |
| Lb. delbrueckii subsp. bulgaricus | Beechwood | Mild oxidative organosolv | SSF | 0.69 | [39] |
| Pine | 0.40 | ||||
| Lactococcus sp. X1 | Corncob | NaOH | SSF | 0.34 | This work |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, N.; Geng, A.; Tu, Z.; Fan, Y.; Xie, R.; Li, X.; Sun, J. Isolation of Lactococcus sp. X1 from Termite Gut, and Its Application in Lactic Acid Production. Fermentation 2023, 9, 85. https://doi.org/10.3390/fermentation9020085
Li N, Geng A, Tu Z, Fan Y, Xie R, Li X, Sun J. Isolation of Lactococcus sp. X1 from Termite Gut, and Its Application in Lactic Acid Production. Fermentation. 2023; 9(2):85. https://doi.org/10.3390/fermentation9020085
Chicago/Turabian StyleLi, Nana, Alei Geng, Zhuowei Tu, Yanan Fan, Rongrong Xie, Xia Li, and Jianzhong Sun. 2023. "Isolation of Lactococcus sp. X1 from Termite Gut, and Its Application in Lactic Acid Production" Fermentation 9, no. 2: 85. https://doi.org/10.3390/fermentation9020085