

The Profile of Exopolysaccharides Produced by Various Lactobacillus Species from Silage during Not-Fat Milk Fermentation
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Cultivation
2.2. EPS Isolation and Preparation of Extract
2.3. The Characterization of EPS Composition
2.4. Cytotoxicity Assay
2.5. Antioxidant Activity
2.5.1. Evaluation of Radical Scavenging Ability (RSA) by 2, 2-Di-phenyl-1-picrylhydrazyl (DPPH) Assay
2.5.2. Ferric Reducing Antioxidant Power Assay (FRAP)
2.5.3. HO Free Radical Scavenging Ability
2.5.4. Determination of Superoxide Anion Radical Scavenging Activity
2.5.5. Fe-Chelating Activity
2.6. Scanning Electron Microscopy
2.7. Statistical Analysis
3. Results
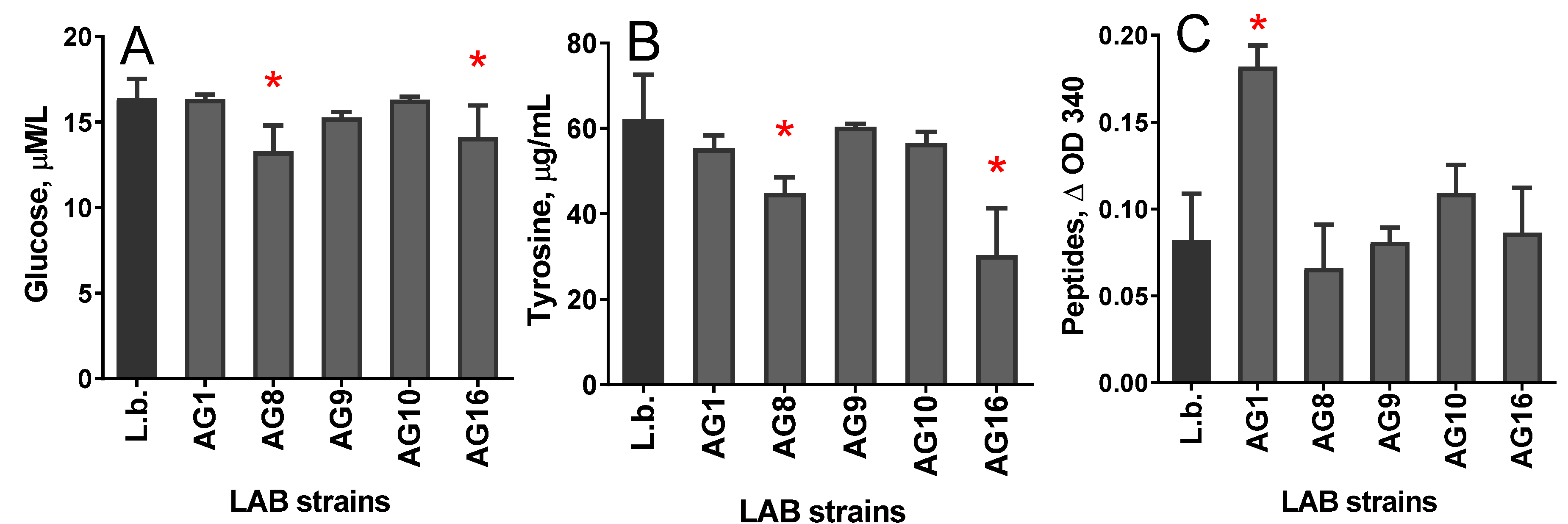
3.1. Chemical Composition of the Protein-Free Extract (PFE)
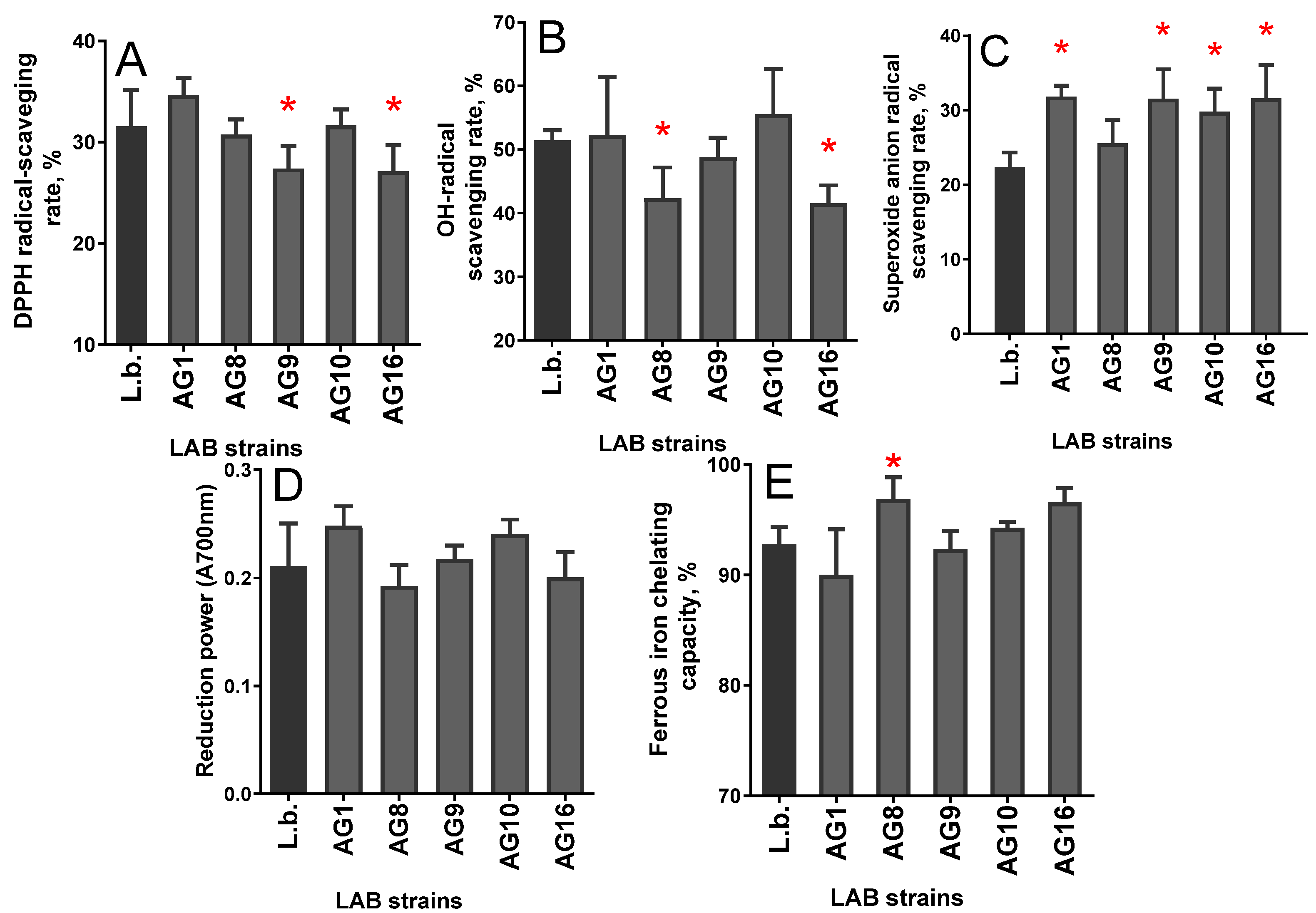
3.2. Antioxidant Properties of PFE
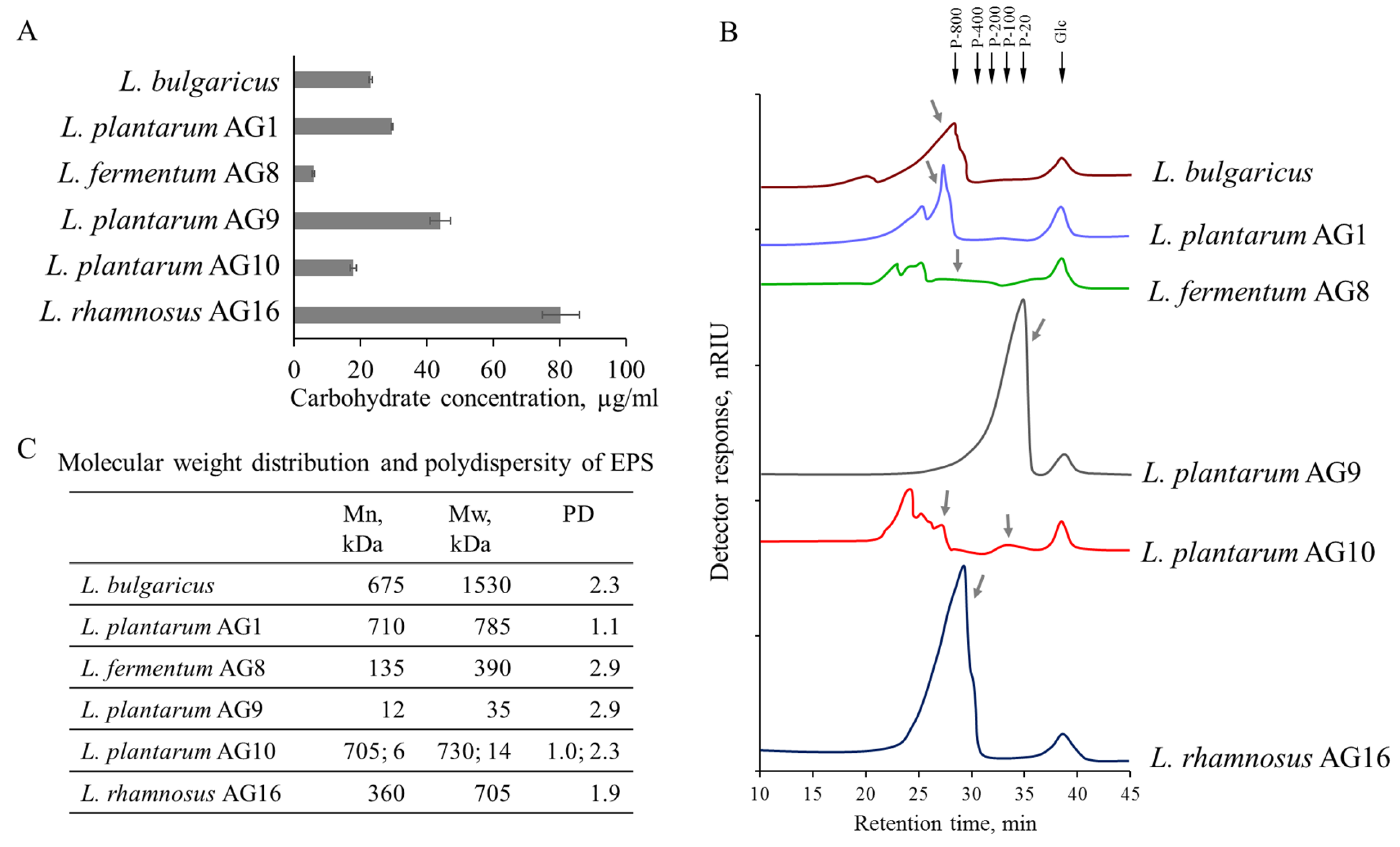
3.3. Yield and Composition of Exopolysaccharides (EPS)
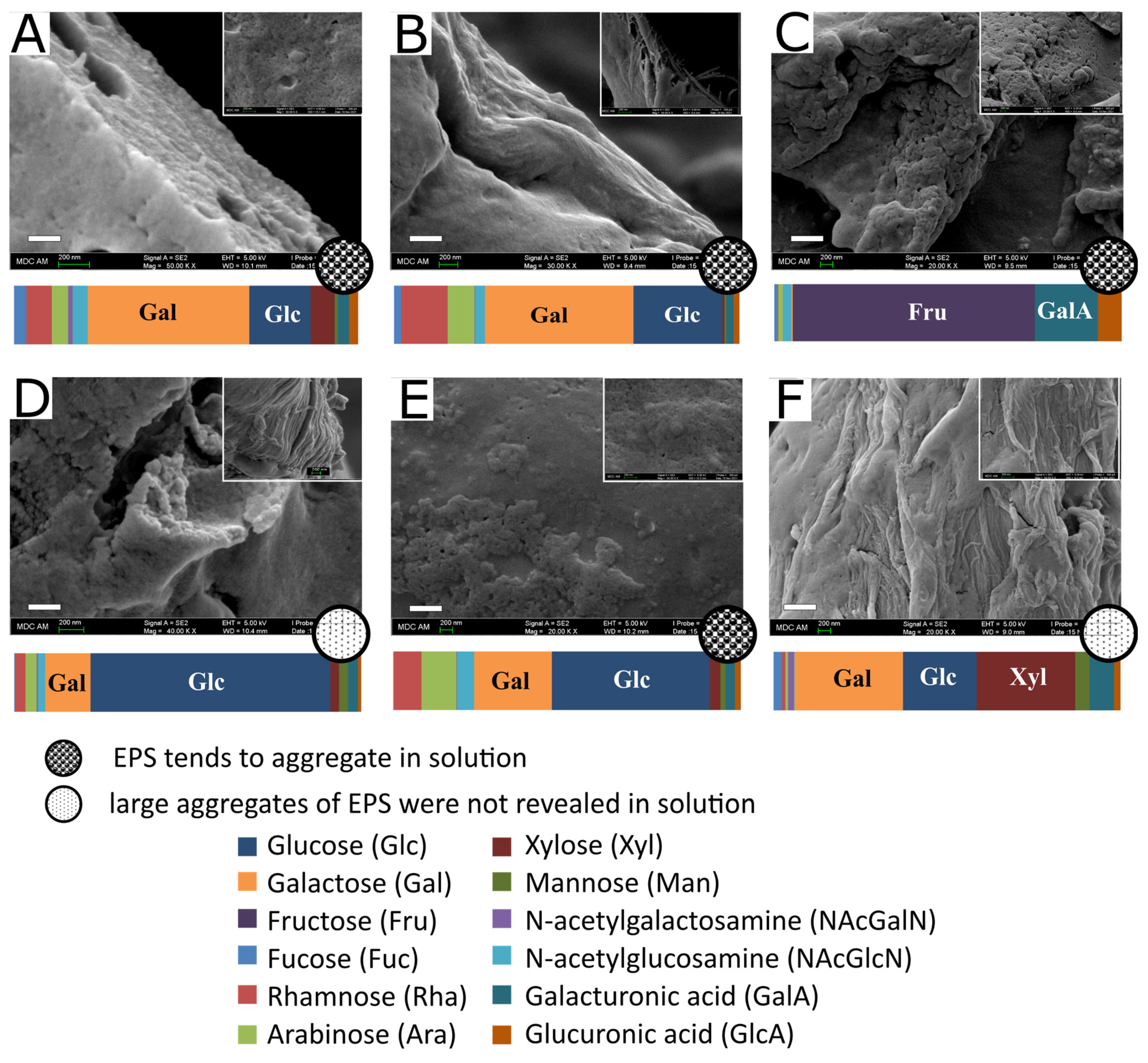
3.4. EPS Microstructureand Surface Morphology
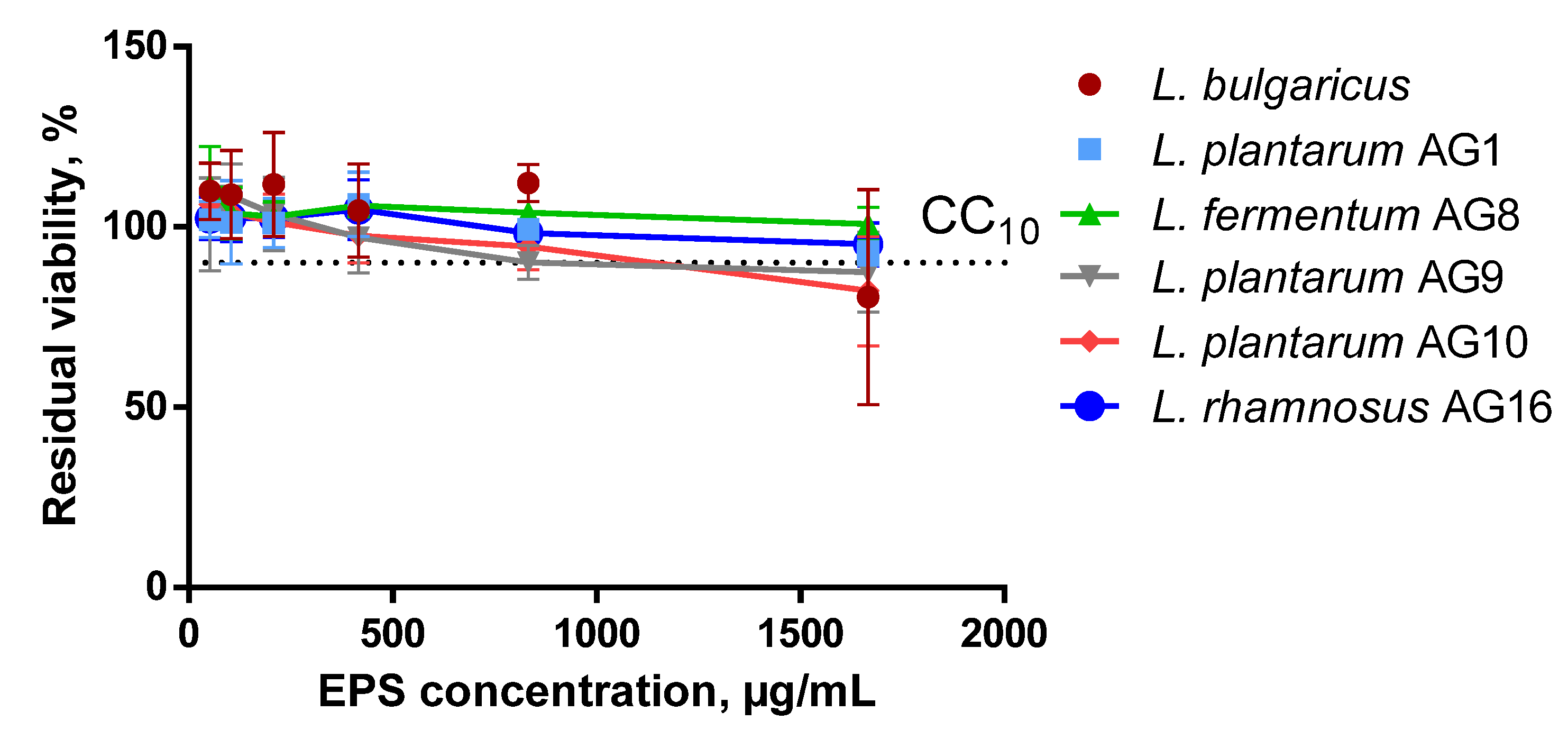
3.5. EPS Cytotoxicity
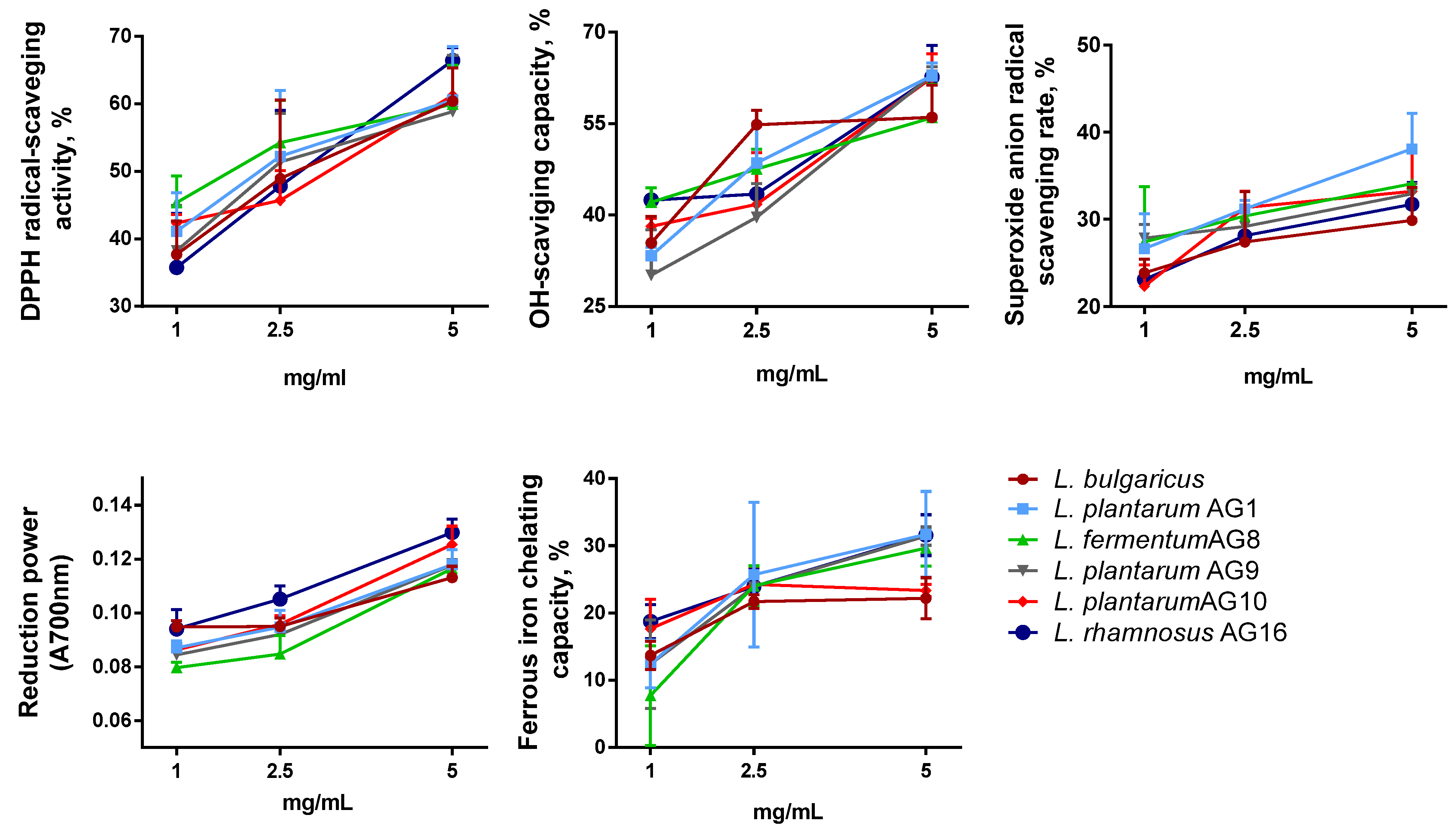
3.6. EPS Antioxidant Activity
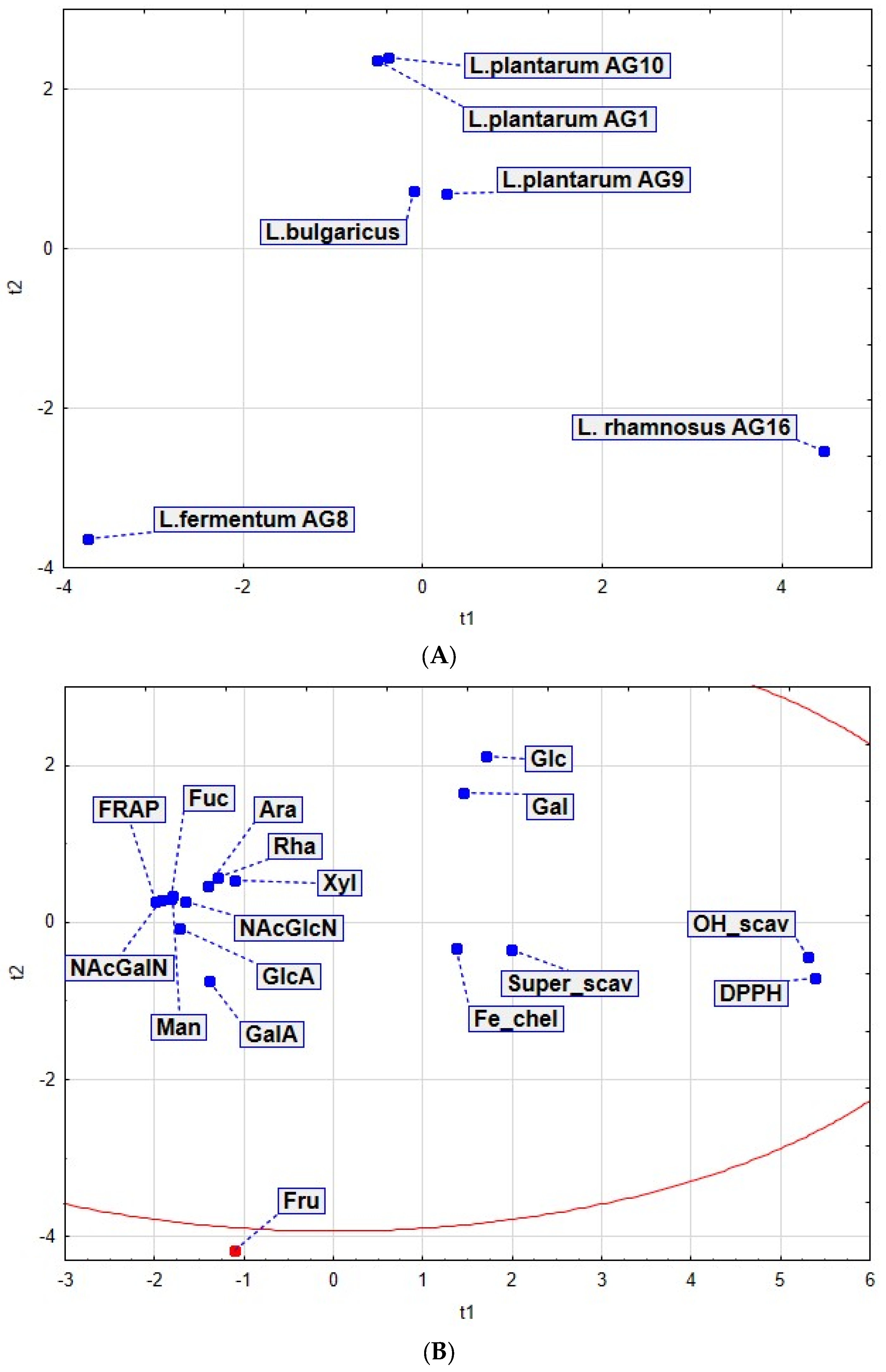
3.7. Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harapanahalli, A.K.; Younes, J.A.; Allan, E.; van der Mei, H.C.; Busscher, H.J. Chemical Signals and Mechanosensing in Bacterial Responses to Their Environment. PLoS Pathog. 2015, 11, e1005057. [Google Scholar] [CrossRef] [PubMed]
- Oleksy, M.; Klewicka, E. Exopolysaccharides produced by Lactobacillus sp.: Biosynthesis and applications. Crit. Rev. Food Sci. Nutr. 2018, 58, 450–462. [Google Scholar] [CrossRef] [PubMed]
- Daba, G.M.; Elnahas, M.O.; Elkhateeb, W.A. Contributions of exopolysaccharides from lactic acid bacteria as biotechnological tools in food, pharmaceutical, and medical applications. Int. J. Biol. Macromol. 2021, 173, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Jurášková, D.; Ribeiro, S.C.; Silva, C.C.G. Exopolysaccharides Produced by Lactic Acid Bacteria: From Biosynthesis to Health-Promoting Properties. Foods 2022, 11, 156. [Google Scholar] [CrossRef]
- Angelin, J.; Kavitha, M. Exopolysaccharides from probiotic bacteria and their health potential. Int. J. Biol. Macromol. 2020, 162, 853–865. [Google Scholar] [CrossRef]
- Nabot, M.; Guérin, M.; Sivakumar, D.; Remize, F.; Garcia, C. Variability of Bacterial Homopolysaccharide Production and Properties during Food Processing. Biology 2022, 11, 171. [Google Scholar] [CrossRef]
- Kavitake, D.; Devi, P.B.; Singh, S.P.; Shetty, P.H. Characterization of a novel galactan produced by Weissella confusa KR780676 from an acidic fermented food. Int. J. Biol. Macromol. 2016, 86, 681–689. [Google Scholar] [CrossRef]
- Ale, E.C.; Perezlindo, M.J.; Pavón, Y.; Peralta, G.H.; Costa, S.; Sabb, A.G.N.; Bergamini, C.; Reinheimer, J.A.; Binetti, A.G. Technological, rheological and sensory characterizations of a yogurt containing an exopolysaccharide extract from Lactobacillus fermentum Lf2, a new food additive. Food Res. Int. 2016, 90, 259–267. [Google Scholar] [CrossRef]
- Badel, S.; Bernardi, T.; Michaud, P. New perspectives for Lactobacilli exopolysaccharides. Biotechnol. Adv. 2011, 29, 54–66. [Google Scholar] [CrossRef]
- Duboc, P.; Mollet, B. Applications of exopolysaccharides in the dairy industry. Int. Dairy J. 2001, 11, 759–768. [Google Scholar] [CrossRef]
- Doleyres, Y.; Schaub, L.; Lacroix, C. Comparison of the functionality of exopolysaccharides produced in situ or added as bioingredients on yogurt properties. J. Dairy Sci. 2005, 88, 4146–4156. [Google Scholar] [CrossRef] [Green Version]
- Perry, D.B.; McMahon, D.J.; Oberg, C.J. Effect of Exopolysaccharide-Producing Cultures on Moisture Retention in Low Fat Mozzarella Cheese1. J. Dairy Sci. 1997, 80, 799–805. [Google Scholar] [CrossRef]
- Korcz, E.; Kerényi, Z.; Varga, L. Dietary fibers, prebiotics, and exopolysaccharides produced by lactic acid bacteria: Potential health benefits with special regard to cholesterol-lowering effects. Food Funct. 2018, 9, 3057–3068. [Google Scholar] [CrossRef]
- Wegh, C.A.M.; Geerlings, S.Y.; Knol, J.; Roeselers, G.; Belzer, C. Postbiotics and Their Potential Applications in Early Life Nutrition and Beyond. Int. J. Mol. Sci. 2019, 20, 4673. [Google Scholar] [CrossRef] [Green Version]
- İspirli, H.; Demirbaş, F.; Dertli, E. Glucan type exopolysaccharide (EPS) shows prebiotic effect and reduces syneresis in chocolate pudding. J. Food Sci. Technol. 2018, 55, 3821–3826. [Google Scholar] [CrossRef]
- Bello, F.D.; Walter, J.; Hertel, C.; Hammes, W.P. In vitro study of prebiotic properties of levan-type exopolysaccharides from Lactobacilli and non-digestible carbohydrates using denaturing gradient gel electrophoresis. Syst. Appl. Microbiol. 2001, 24, 232–237. [Google Scholar] [CrossRef]
- Ismail, B.; Nampoothiri, K.M. Exposition of antitumour activity of a chemically characterized exopolysaccharide from a probiotic Lactobacillus plantarum MTCC 9510. Biologia 2013, 68, 1041–1047. [Google Scholar] [CrossRef] [Green Version]
- Sasikumar, K.; Kozhummal Vaikkath, D.; Devendra, L.; Nampoothiri, K.M. An exopolysaccharide (EPS) from a Lactobacillus plantarum BR2 with potential benefits for making functional foods. Bioresour. Technol. 2017, 241, 1152–1156. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, Z.; Qiu, L.; Zhang, F.; Xu, X.; Wei, H.; Tao, X. Characterization and bioactivities of the exopolysaccharide from a probiotic strain of Lactobacillus plantarum WLPL04. J. Dairy Sci. 2017, 100, 6895–6905. [Google Scholar] [CrossRef]
- Zhou, T.; Huo, R.; Kwok, L.-Y.; Li, C.; Ma, Y.; Mi, Z.; Chen, Y. Effects of applying Lactobacillus helveticus H9 as adjunct starter culture in yogurt fermentation and stor AGe. J. Dairy Sci. 2019, 102, 223–235. [Google Scholar] [CrossRef] [Green Version]
- Das, D.; Baruah, R.; Goyal, A. A food additive with prebiotic properties of an α-d-glucan from Lactobacillus plantarum DM5. Int. J. Biol. Macromol. 2014, 69, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Hongpattarakere, T.; Cherntong, N.; Wichienchot, S.; Kolida, S.; Rastall, R.A. In vitro prebiotic evaluation of exopolysaccharides produced by marine isolated lactic acid bacteria. Carbohydr. Polym. 2012, 87, 846–852. [Google Scholar] [CrossRef] [PubMed]
- Bomfim, V.B.; Neto, J.H.P.L.; Leite, K.S.; de Andrade Vieira, É.; Iacomini, M.; Silva, C.M.; dos Santos, K.M.O.; Cardarelli, H.R. Partial characterization and antioxidant activity of exopolysaccharides produced by Lactobacillus plantarum CNPC003. LWT 2020, 127, 109349. [Google Scholar] [CrossRef]
- Nehal, F.; Sahnoun, M.; Smaoui, S.; Jaouadi, B.; Bejar, S.; Mohammed, S. Characterization, high production and antimicrobial activity of exopolysaccharides from Lactococcus lactis F-mou. Microb. Pathog. 2019, 132, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Du, R.; Qiao, X.; Zhao, F.; Song, Q.; Zhou, Q.; Wang, Y.; Pan, L.; Han, Y.; Zhou, Z. Purification, characterization and antioxidant activity of dextran produced by Leuconostoc pseudomesenteroides from homemade wine. Carbohydr. Polym. 2018, 198, 529–536. [Google Scholar] [CrossRef]
- Pan, D.; Mei, X. Antioxidant activity of an exopolysaccharide purified from Lactococcus lactis subsp. lactis 12. Carbohydr. Polym. 2010, 80, 908–914. [Google Scholar] [CrossRef]
- Xu, Y.; Cui, Y.; Wang, X.; Yue, F.; Shan, Y.; Liu, B.; Zhou, Y.; Yi, Y.; Lü, X. Purification, characterization and bioactivity of exopolysaccharides produced by Lactobacillus plantarum KX041. Int. J. Biol. Macromol. 2019, 128, 480–492. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, X.; Yang, Y.; Zhao, A.; Yang, Z. Characterization and bioactivities of an exopolysaccharide produced by Lactobacillus plantarum YW32. Int. J. Biol. Macromol. 2015, 74, 119–126. [Google Scholar] [CrossRef]
- Li, S.; Huang, R.; Shah, N.P.; Tao, X.; Xiong, Y.; Wei, H. Antioxidant and antibacterial activities of exopolysaccharides from Bifidobacterium bifidum WBIN03 and Lactobacillus plantarum R315. J. Dairy Sci. 2014, 97, 7334–7343. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Ji, J.; Rui, X.; Yu, J.; Tang, W.; Chen, X.; Jiang, M.; Dong, M. Production of exopolysaccharides by Lactobacillus helveticus MB2-1 and its functional characteristics in vitro. LWT Food Sci. Technol. 2014, 59, 732–739. [Google Scholar] [CrossRef]
- Lo, T.C.-T.; Chang, C.A.; Chiu, K.-H.; Tsay, P.-K.; Jen, J.-F. Correlation evaluation of antioxidant properties on the monosaccharide components and glycosyl link AGes of polysaccharide with different measuring methods. Carbohydr. Polym. 2011, 86, 320–327. [Google Scholar] [CrossRef]
- Jia, K.; Tao, X.; Liu, Z.; Zhan, H.; He, W.; Zhang, Z.; Zeng, Z.; Wei, H. Characterization of novel exopolysaccharide of Enterococcus faecium WEFA23 from infant and demonstration of its in vitro biological properties. Int. J. Biol. Macromol. 2019, 128, 710–717. [Google Scholar] [CrossRef]
- Trabelsi, I.; Slima, S.B.; Chaabane, H.; Riadh, B.S. Purification and characterization of a novel exopolysaccharides produced by Lactobacillus sp. Ca6. Int. J. Biol. Macromol. 2015, 74, 541–546. [Google Scholar] [CrossRef]
- Guo, Y.; Pan, D.; Sun, Y.; Xin, L.; Li, H.; Zeng, X. Antioxidant activity of phosphorylated exopolysaccharide produced by Lactococcus lactis subsp. lactis. Carbohydr. Polym. 2013, 97, 849–854. [Google Scholar] [CrossRef]
- Wang, K.; Li, W.; Rui, X.; Li, T.; Chen, X.; Jiang, M.; Dong, M. Chemical modification, characterization and bioactivity of a released exopolysaccharide (r-EPS1) from Lactobacillus plantarum 70810. Glycoconj. J. 2015, 32, 17–27. [Google Scholar] [CrossRef]
- Błaszczyk, K.; Wilczak, J.; Harasym, J.; Gudej, S.; Suchecka, D.; Królikowski, T.; Lange, E.; Gromadzka-Ostrowska, J. Impact of low and high molecular weight oat beta-glucan on oxidative stress and antioxidant defense in spleen of rats with LPS induced enteritis. Food Hydrocoll. 2015, 51, 272–280. [Google Scholar] [CrossRef]
- Lei, N.; Wang, M.; Zhang, L.; Xiao, S.; Fei, C.; Wang, X.; Zhang, K.; Zheng, W.; Wang, C.; Yang, R. Effects of low molecular weight yeast β-glucan on antioxidant and immunological activities in mice. Int. J. Mol. Sci. 2015, 16, 21575–21590. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Wang, C.; Shi, Q.; Ma, C. Preparation of different molecular weight polysaccharides from Porphyridium cruentum and their antioxidant activities. Int. J. Biol. Macromol. 2009, 45, 42–47. [Google Scholar] [CrossRef]
- Gavrilova, E.; Anisimova, E.; Gabdelkhadieva, A.; Nikitina, E.; Vafina, A.; Yarullina, D.; Bogachev, M.; Kayumov, A. Newly isolated lactic acid bacteria from silage targeting biofilms of foodborne pathogens during milk fermentation. BMC Microbiol. 2019, 19, 248. [Google Scholar] [CrossRef]
- Nikitina, E.; Petrova, T.; Adel, V.; Asya, E.; Monyr, N.Y.; Airat, K. Textural and Functional Properties of Skimmed and Whole Milk Fermented by Novel Lactiplantibacillus plantarum AG10 Strain Isolated from Silage. Fermentation 2022, 8, 290. [Google Scholar] [CrossRef]
- Cory, A.H.; Owen, T.C.; Barltrop, J.A.; Cory, J.G. Use of an aqueous soluble tetrazolium/formazan assay for cell growth assays in culture. Cancer Commun. 1991, 3, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Nikitina, E.V.; Yurtaeva, T.A.; Tsyganov, M.S.; Ezhkova, G.O. Physico-Chemical and Antioxidant Properties of Skimmed Varenets (Slavic Baked Milk Yogurt) Mixed with Enzyme-Modified Potato Starches. Curr. Res. Nutr. Food Sci. 2021, 9, 88–99. [Google Scholar] [CrossRef]
- Qin, S.; Huang, Z.; Wang, Y.; Pei, L.; Shen, Y. Probiotic potential of Lactobacillus isolated from horses and its therapeutic effect on DSS-induced colitis in mice. Microb. Pathog. 2022, 165, 105216. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.-M.; Zhou, J.-M.; Zhou, Q.-Q.; Li, P.; Xie, Y.-Y.; Zhou, T.; Gu, Q. Purification, characterization and biological activities of exopolysaccharides from Lactobacillus rhamnosus ZFM231 isolated from milk. LWT 2021, 147, 111561. [Google Scholar] [CrossRef]
- Muniandy, P.; Shori, A.B.; Baba, A.S. Influence of green, white and black tea addition on the antioxidant activity of probiotic yogurt during refrigerated stor AGe. Food Pack. Aging Shelf Life 2016, 8, 1–8. [Google Scholar] [CrossRef]
- Diez-Ozaeta, I.; Astiazaran, O.J. Fermented foods: An update on evidence-based health benefits and future perspectives. Food Res. Int. 2022, 156, 111133. [Google Scholar] [CrossRef]
- Grobben, G.J.; Sikkema, J.; Smith, M.R.; de Bont, J.A. Production of extracellular polysaccharides by Lactobacillus delbrueckii ssp. bulgaricus NCFB 2772 grown in a chemically defined medium. J. Appl. Bacteriol. 1995, 79, 103–107. [Google Scholar] [CrossRef]
- Grobben, G.J.; Smith, M.R.; Sikkema, J.; de Bont, J.A. Influence of fructose and glucose on the production of exopolysaccharides and the activities of enzymes involved in the sugar metabolism and the synthesis of sugar nucleotides in Lactobacillus delbrueckii subsp. bulgaricus NCFB 2772. Appl. Microbiol. Biotechnol. 1996, 46, 279–284. [Google Scholar] [CrossRef]
- Gamar, L.; Blondeau, K.; Simonet, J.M. Physiological approach to extracellular polysaccharide production by Lactobacillus rhamnosus strain C83. J. Appl. Microbiol. 1997, 83, 281–287. [Google Scholar] [CrossRef]
- Goh, K.K.; Haisman, D.R.; Singh, H. Development of an improved procedure for isolation and purification of exopolysaccharides produced by Lactobacillus delbrueckii subsp. bulgaricus NCFB 2483. Appl. Microbiol. Biotechnol. 2005, 67, 202–208. [Google Scholar] [CrossRef]
- Kimmel, S.A.; Roberts, R.F. Development of a growth medium suitable for exopolysaccharide production by Lactobacillus delbrueckii ssp. bulgaricus RR. Int. J. Food Microbiol. 1998, 40, 87–92. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, X.; Tian, Z.; He, C.; Yang, Y.; Yang, Z. Isolation and characterization of exopolysaccharide-producing Lactobacillus plantarum SKT109 from Tibet Kefir. Pol. J. Food Nutr. Sci. 2015, 65, 269–279. [Google Scholar] [CrossRef] [Green Version]
- Imran, M.Y.; Reehana, N.; Jayaraj, K.A.; Ahamed, A.A.; Dhanasekaran, D.; Thajuddin, N.; Alharbi, N.S.; Muralitharan, G. Statistical optimization of exopolysaccharide production by Lactobacillus plantarum NTMI05 and NTMI20. Int. J. Biol. Macromol. 2016, 93, 731–745. [Google Scholar] [CrossRef]
- Berecka, M.P.; Waśko, A.; Szwajgier, D.; Choma, A. Bifidogenic and antioxidant activity of exopolysaccharides produced by Lactobacillus rhamnosus E/N cultivated on different carbon sources. Pol. J. Microbiol. 2013, 62, 181–189. [Google Scholar] [CrossRef]
- Fukuda, K.; Shi, T.; Nagami, K.; Leo, F.; Nakamura, T.; Yasuda, K.; Senda, A.; Motoshima, H.; Urashima, T. Effects of carbohydrate source on physicochemical properties of the exopolysaccharide produced by Lactobacillus fermentum TDS030603 in a chemically defined medium. Carbohydr. Polym. 2010, 79, 1040–1045. [Google Scholar] [CrossRef]
- Ismail, B.; Nampoothiri, K.M. Molecular characterization of an exopolysaccharide from a probiotic Lactobacillus plantarum MTCC 9510 and its efficacy to improve the texture of starchy food. J. Food Sci. Technol. 2014, 51, 4012–4018. [Google Scholar] [CrossRef] [Green Version]
- Vijayendra, S.; Palanivel, G.; Mahadevamma, S.; Tharanathan, R. Physico-chemical characterization of a new heteropolysaccharide produced by a native isolate of heterofermentative Lactobacillus sp. CFR-2182. Arch. Microbiol. 2009, 191, 303–310. [Google Scholar] [CrossRef]
- Faber, E.J.; Zoon, P.; Kamerling, J.P.; Vliegenthart, J.F. The exopolysaccharides produced by Streptococcus thermophilus Rs and Sts have the same repeating unit but differ in viscosity of their milk cultures. Carbohydr. Res. 1998, 310, 269–276. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, Y.; Ye, L.; Wang, C. The anti-cancer effects and mechanisms of lactic acid bacteria exopolysaccharides in vitro: A review. Carbohydr. Polym. 2021, 253, 117308. [Google Scholar] [CrossRef]
- Tallon, R.; Bressollier, P.; Urdaci, M.C. Isolation and characterization of two exopolysaccharides produced by Lactobacillus plantarum EP56. Res. Microbiol. 2003, 154, 705–712. [Google Scholar] [CrossRef]
- Ramsey, I.; Corsini, N.; Peters, M.D.J.; Eckert, M. A rapid review of consumer health information needs and preferences. Patient Educ. Couns. 2017, 100, 1634–1642. [Google Scholar] [CrossRef] [PubMed]
- Arakawa, K.; Matsunaga, K.; Takihiro, S.; Moritoki, A.; Ryuto, S.; Kawai, Y.; Masuda, T.; Miyamoto, T. Lactobacillus gasseri requires peptides, not proteins or free amino acids, for growth in milk. J. Dairy Sci. 2015, 98, 1593–1603. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, M.W.; Tellez, A.M. Lactobacillus helveticus: The proteolytic system. Front. Microbiol. 2013, 4, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trimigno, A.; Bøge Lyndgaard, C.; Atladóttir, G.A.; Aru, V.; Balling Engelsen, S.; Harder Clemmensen, L.K. An NMR Metabolomics Approach to Investigate Factors Affecting the Yoghurt Fermentation Process and Quality. Metabolites 2020, 10, 293. [Google Scholar] [CrossRef]
- Power, O.; Jakeman, P.; FitzGerald, R.J. Antioxidative peptides: Enzymatic production, in vitro and in vivo antioxidant activity and potential applications of milk-derived antioxidative peptides. Amino Acids 2013, 44, 797–820. [Google Scholar] [CrossRef] [PubMed]
- Díaz, M.; Decker, E.A. Antioxidant mechanisms of caseinophosphopeptides and casein hydrolysates and their application in ground beef. J. Agric. Food Chem. 2004, 52, 8208–8213. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample of EPS | Monosaccharide Composition, mol% | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fuc | Rha | Ara | NAcGalN | NAcGlcN | Gal | Glc | Xyl | Man | Fru | GalA | GlcA | |
| L. bulgaricus | 3.6 ± 0.1 | 7.3 ± 0.2 | 4.9 ± 0.1 | 1.3 ± 0.0 | 4.2 ± 0.1 | 46.9 ± 1.6 | 17.9 ± 0.7 | 7.1 ± 0.2 | 1.0 ± 0.0 | 0.0 ± 0.0 | 3.2 ± 0.1 | 2.6 ± 0.1 |
| L. plantarum AG1 | 2.2 ± 0.1 | 13.4 ± 0.4 | 7.8 ± 0.2 | 0.1 ± 0.0 | 2.9 ± 0.1 | 42.9 ± 1.3 | 25.7 ± 0.8 | 0.6 ± 0.0 | 0.6 ± 0.0 | 0.0 ± 0.0 | 2.1 ± 0.1 | 1.7 ± 0.1 |
| L. fermentum AG8 | 0.7 ± 0.0 | 0.3 ± 0.0 | 1.4 ± 0.0 | 0.0 ± 0.0 | 2.4 ± 0.1 | 0.3 ± 0.0 | 0.1 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 69.9 ± 2.1 | 18.2 ± 0.5 | 6.7 ± 0.2 |
| L. plantarum AG9 | 0.3 ± 0.0 | 2.9 ± 0.1 | 3.3 ± 0.1 | 0.2 ± 0.0 | 2.1 ± 0.1 | 13.1 ± 0.6 | 69.1 ± 2.8 | 2.6 ± 0.1 | 2.6 ± 0.1 | 0.0 ± 0.0 | 2.9 ± 0.1 | 0.9 ± 0.0 |
| L. plantarum AG10 | 0.0 ± 0.0 | 8.1 ± 0.2 | 10.2 ± 0.3 | 0.1 ± 0.0 | 5.0 ± 0.2 | 22.3 ± 0.9 | 45.5 ± 1.6 | 3.0 ± 0.1 | 1.4 ± 0.0 | 0.0 ± 0.0 | 2.7 ± 0.1 | 1.7 ± 0.1 |
| L. rhamnosus AG16 | 2.5 ± 0.1 | 0.8 ± 0.0 | 0.8 ± 0.0 | 1.7 ± 0.1 | 0.2 ± 0.0 | 31.3 ± 1.1 | 21.1 ± 0.8 | 28.5 ± 0.9 | 4.2 ± 0.1 | 0.0 ± 0.0 | 6.9 ± 0.2 | 2.0 ± 0.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikitina, E.; Petrova, T.; Sungatullina, A.; Bondar, O.; Kharina, M.; Mikshina, P.; Gavrilova, E.; Kayumov, A. The Profile of Exopolysaccharides Produced by Various Lactobacillus Species from Silage during Not-Fat Milk Fermentation. Fermentation 2023, 9, 197. https://doi.org/10.3390/fermentation9020197
Nikitina E, Petrova T, Sungatullina A, Bondar O, Kharina M, Mikshina P, Gavrilova E, Kayumov A. The Profile of Exopolysaccharides Produced by Various Lactobacillus Species from Silage during Not-Fat Milk Fermentation. Fermentation. 2023; 9(2):197. https://doi.org/10.3390/fermentation9020197
Chicago/Turabian StyleNikitina, Elena, Tatyana Petrova, Alya Sungatullina, Oxana Bondar, Maria Kharina, Polina Mikshina, Elizaveta Gavrilova, and Airat Kayumov. 2023. "The Profile of Exopolysaccharides Produced by Various Lactobacillus Species from Silage during Not-Fat Milk Fermentation" Fermentation 9, no. 2: 197. https://doi.org/10.3390/fermentation9020197