
Improving the Functionality of Lentil–Casein Protein Complexes through Structural Interactions and Water Kefir-Assisted Fermentation
 , , , , ,
, , , , ,
Abstract
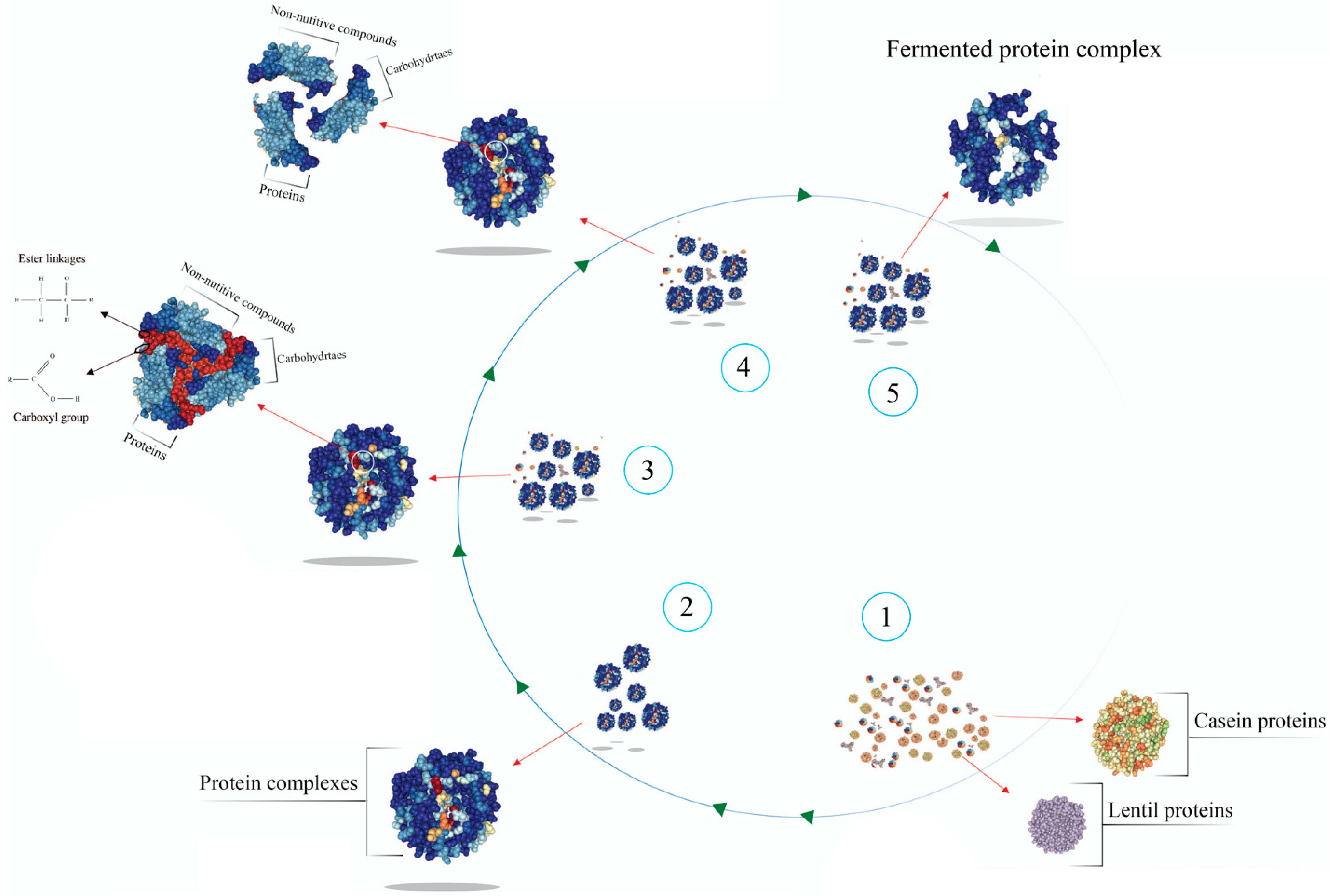
:1. Introduction
2. Materials and Methods
2.1. Extraction of LP
2.2. Protein Complexation
2.3. Water Solubility
2.4. Spectral Acquisition
2.4.1. Fourier-Transform Infrared Spectroscopy (FTIR)
2.4.2. Spectrofluorometry
2.4.3. Non-Covalent Forces
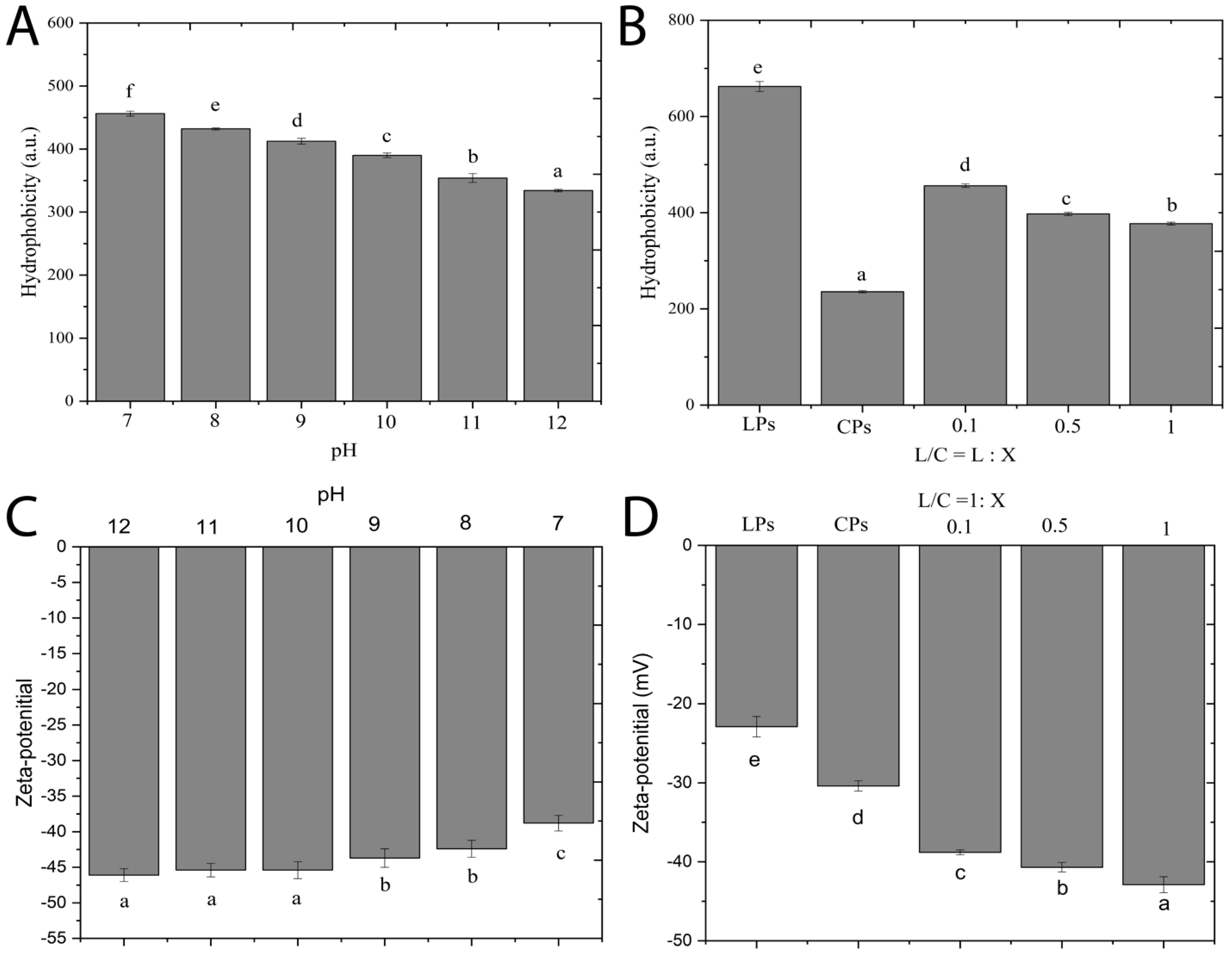
2.4.4. Surface Hydrophobicity (H0)
2.4.5. Ultraviolet-Visible Spectroscopy
2.5. Zeta Potential
2.6. Scanning Electron Microscopy (SEM)
2.7. Preparation of Water Kefir
2.8. Preparation of Fermented Lentil–Casein Protein Complex
2.9. pH and Total Soluble Solids (TSS)
2.10. Protein Digestibility
2.11. Determination of the Nutrients of Fermented Protein Complexes
2.11.1. Phenolic Compounds
2.11.2. Total Phenolic Content (TPC) and Total Saponin Content (TSC)
2.11.3. Determination of Sugars
2.12. Microbiological Quality
2.13. Statistical Analysis
3. Results and Discussion
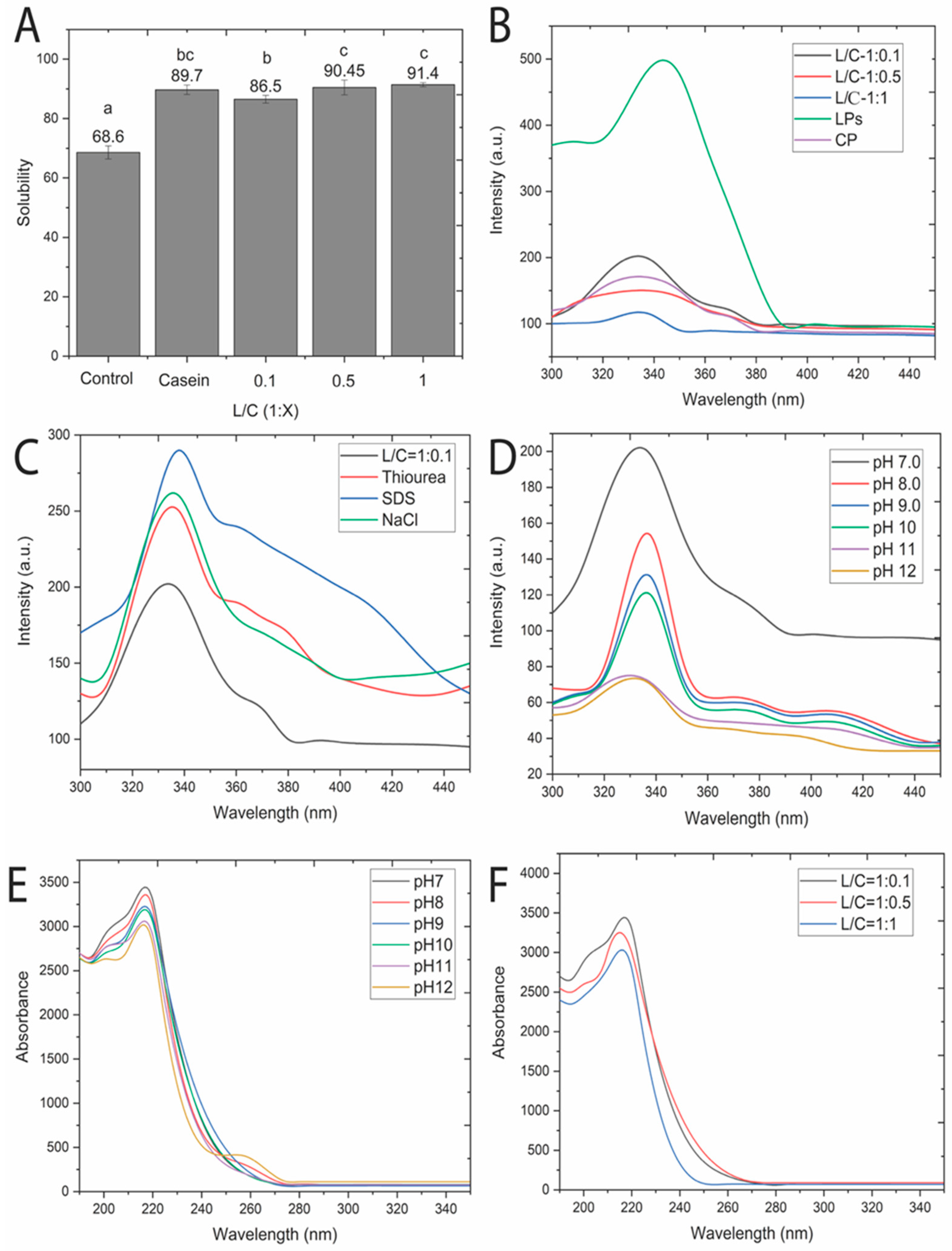
3.1. Solubility of Lentil–Casein Protein Complexes (LCPC)
3.2. LP and CP Interaction
3.3. Structural Folding of Protein Complexes
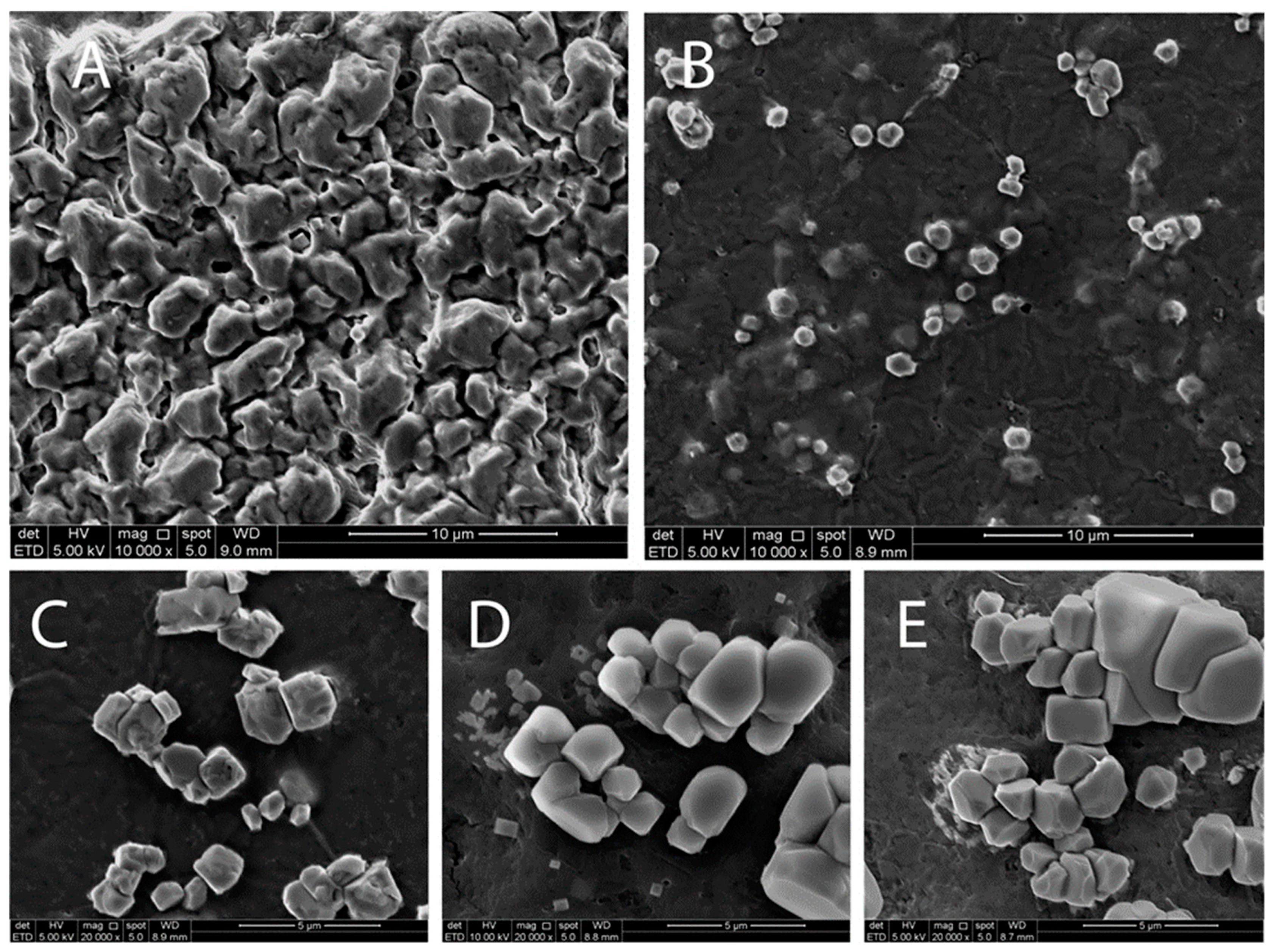
3.4. Surface Morphology of LCPC
3.5. Protein Interfacial Characteristics Resulting from Protein Complexation
3.6. Effects of Water Kefir-Assisted Fermentation on the pH and TSS of Fermented Lentil–Casein Protein Complex at a Ratio of 1.0:0.1 (LCPC 1.0:0.1)
3.7. Effects of Water Kefir-Assisted Fermentation on the Protein Quality of Fermented LCPC 1.0:0.1
3.7.1. Protein Digestibility
3.7.2. Secondary Protein Structure
3.8. Effects of Water Kefir-Assisted Fermentation on the Nutrients of Fermented LCPC 1.0:0.1
3.8.1. Phenolic Compounds
3.8.2. Total Saponin Content (TSC)
3.8.3. Sugars
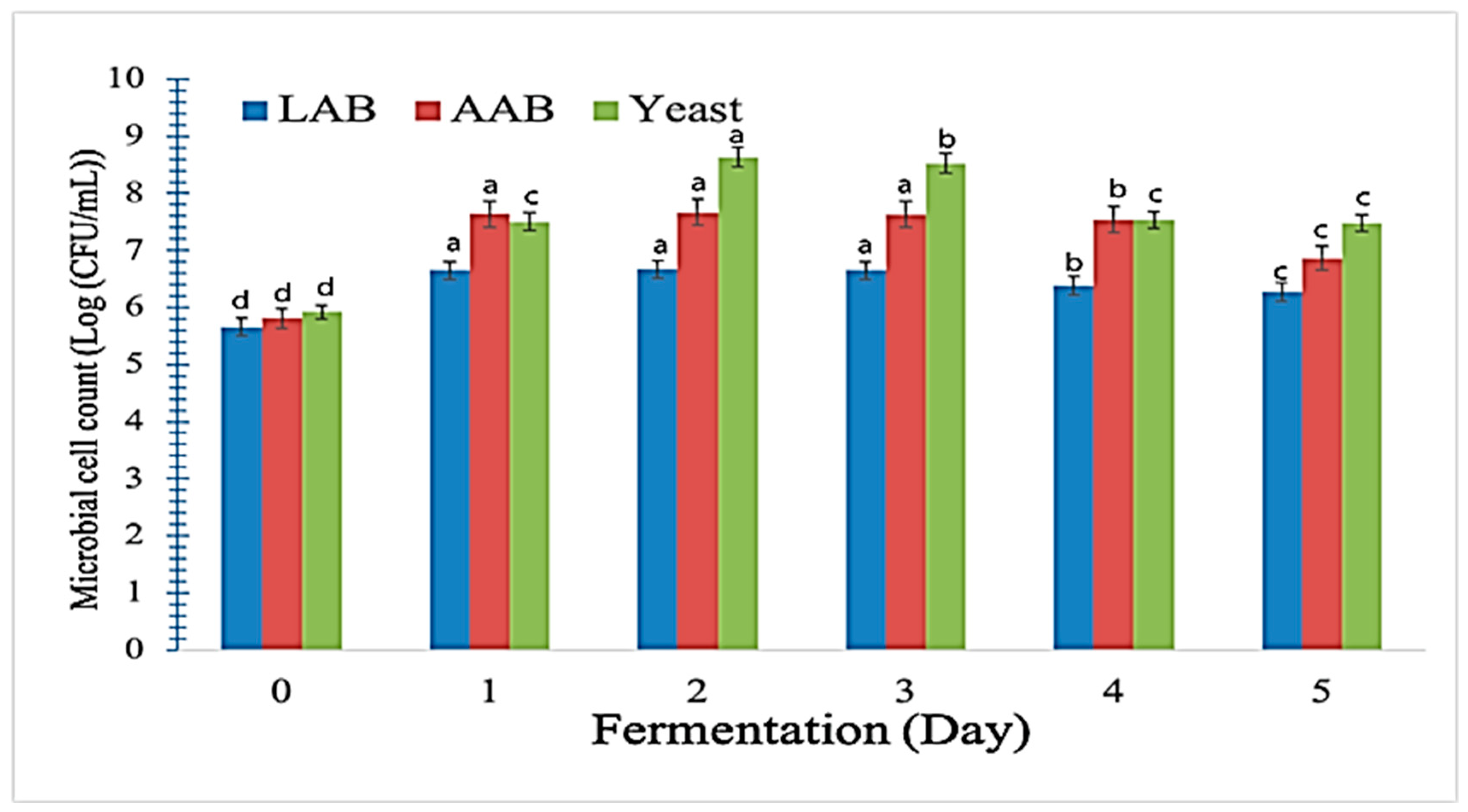
3.9. Effects of Water Kefir-Assisted Fermentation on the Microbiological Qualities of Fermented LCPC 1.0:0.1
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jarpa-Parra, M.; Bamdad, F.; Wang, Y.; Tian, Z.; Temelli, F.; Han, J.; Chen, L. Optimization of lentil protein extraction and the influence of process pH on protein structure and functionality. LWT 2014, 57, 461–469. [Google Scholar] [CrossRef]
- Alrosan, M.; Tan, T.-C.; Easa, A.M.; Gammoh, S.; Alu’datt, M.H. Mechanism of the structural interaction between whey and lentil proteins in the unique creation of a protein structure. J. Food Sci. 2021, 86, 5282–5294. [Google Scholar] [CrossRef] [PubMed]
- Alrosan, M.; Tan, T.-C.; Easa, A.M.; Gammoh, S.; Alu’datt, M.H. Effects of fermentation on the quality, structure, and nonnutritive contents of lentil (Lens culinaris) proteins. J. Food Qual. 2021, 2021, 5556450. [Google Scholar] [CrossRef]
- Alrosan, M.; Tan, T.-C.; Easa, A.M.; Gammoh, S.; Alu’datt, M.H. Molecular forces governing protein-protein interaction: Structure-function relationship of complexes protein in the food industry. Crit. Rev. Food Sci. Nutr. 2022, 62, 4036–4052. [Google Scholar] [CrossRef]
- Wang, R.; Xu, P.; Chen, Z.; Zhou, X.; Wang, T. Complexation of rice proteins and whey protein isolates by structural interactions to prepare soluble protein composites. LWT 2019, 101, 207–213. [Google Scholar] [CrossRef]
- Wang, T.; Chen, X.; Zhong, Q.; Chen, Z.; Wang, R.; Patel, A.R. Facile and efficient construction of water-soluble biomaterials with tunable mesoscopic structures using all-natural edible proteins. Adv. Funct. Mater. 2019, 29, 1901830. [Google Scholar] [CrossRef]
- Wang, Y.H.; Wang, J.M.; Wan, Z.L.; Yang, X.Q.; Chen, X.W. Corn protein hydrolysate as a new structural modifier for soybean protein isolate based O/W emulsions. LWT 2020, 118, 108763. [Google Scholar] [CrossRef]
- He, J.; Wang, R.; Feng, W.; Chen, Z.; Wang, T. Design of novel edible hydrocolloids by structural interplays between wheat gluten proteins and soy protein isolates. Food Hydrocoll. 2020, 100, 105395. [Google Scholar] [CrossRef]
- Azi, F.; Tu, C.; Rasheed, H.A.; Dong, M. Comparative study of the phenolics, antioxidant and metagenomic composition of novel soy whey-based beverages produced using three different water kefir microbiota. Int. J. Food Sci. Technol. 2020, 55, 1689–1697. [Google Scholar] [CrossRef]
- Alrosan, M.; Tan, T.-C.; Koh, W.Y.; Easa, A.M.; Gammoh, S.; Alu’datt, M.H. Overview of fermentation process: Structure-function relationship on protein quality and non-nutritive compounds of plant-based proteins and carbohydrates. Crit. Rev. Food Sci. Nutr. 2022, 1–15. [Google Scholar] [CrossRef]
- Chandra-Hioe, M.V.; Wong, C.H.M.; Arcot, J. The potential use of fermented chickpea and faba bean flour as food ingredients. Plant Foods Hum. Nutr. 2016, 71, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Official Methods of Analysis of AOAC. Method 930.29, 19th ed.; AOAC International: Rockville, USA, 2012. [Google Scholar]
- Morr, C.V.; German, B.; Kinsella, J.E.; Regenstein, J.M.; Van Buren, J.P.; Kilara, A.; Lewis, B.A.; Mangino, M.E. A collaborative study to develop a standardized food protein solubility procedure. J. Food Sci. 1985, 50, 1715–1718. [Google Scholar] [CrossRef]
- Johnston, S.P.; Nickerson, M.T.; Low, N.H. The physicochemical properties of legume protein isolates and their ability to stabilize oil-in-water emulsions with and without genipin. J. Food Sci. Technol. 2015, 52, 4135–4145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, C.C.; Monteiro, M.L.G.; da Costa-Lima, B.R.C.; Alvares, T.S.; Conte-Junior, C.A. In vitro digestibility of commercial whey protein supplements. LWT 2015, 61, 7–11. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Zhang, Z.-Q.; Liu, C.-M.; Xie, M.-Y.; Tu, Z.-C.; Liu, J.-H.; Liang, R.-H. The effect of dynamic high-pressure microfluidization on the activity, stability and conformation of trypsin. Food Chem. 2010, 123, 616–621. [Google Scholar] [CrossRef]
- Jarpa-Parra, M.; Bamdad, F.; Tian, Z.; Zeng, H.; Temelli, F.; Chen, L. Impact of pH on molecular structure and surface properties of lentil legumin-like protein and its application as foam stabilizer. Colloids Surf. B 2015, 132, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Tomczyńska-Mleko, M.; Kamysz, E.; Sikorska, E.; Puchalski, C.; Mleko, S.; Ozimek, L.; Kowaluk, G.; Gustaw, W.; Wesołowska-Trojanowska, M. Changes of secondary structure and surface tension of whey protein isolate dispersions upon pH and temperature. Czech J. Food Sci. 2014, 32, 82–89. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Zhou, C.; Fu, F.; Chen, Z.; Wu, Q. Effect of high-pressure homogenization on particle size and film properties of soy protein isolate. Ind. Crops Prod. 2013, 43, 538–544. [Google Scholar] [CrossRef]
- Tepari, E.A.; Nakhla, G.; Haroun, B.M.; Hafez, H. Co-fermentation of carbohydrates and proteins for biohydrogen production: Statistical optimization using Response Surface Methodology. Int. J. Hydrogen Energy 2020, 45, 2640–2654. [Google Scholar] [CrossRef]
- Çabuk, B.; Nosworthy, M.G.; Stone, A.K.; Korber, D.R.; Tanaka, T.; House, J.D.; Nickerson, M.T. Effect of fermentation on the protein digestibility and levels of non-nutritive compounds of pea protein concentrate. Food Technol. Biotechnol. 2018, 56, 257–264. [Google Scholar] [CrossRef]
- Jia, J.; Ji, B.; Tian, L.; Li, M.; Lu, M.; Ding, L.; Liu, X.; Duan, X. Mechanism study on enhanced foaming properties of individual albumen proteins by Lactobacillus fermentation. Food Hydrocoll. 2021, 111, 106218. [Google Scholar] [CrossRef]
- Tu, C.; Azi, F.; Huang, J.; Xu, X.; Xing, G.; Dong, M. Quality and metagenomic evaluation of a novel functional beverage produced from soy whey using water kefir grains. LWT 2019, 113, 108258. [Google Scholar] [CrossRef]
- dos Santos, D.C.; de Oliveira Filho, J.G.; Santana, A.C.A.; de Freitas, B.S.M.; Silva, F.G.; Takeuchi, K.P.; Egea, M.B. Optimization of soymilk fermentation with kefir and the addition of inulin: Physicochemical, sensory and technological characteristics. LWT 2019, 104, 30–37. [Google Scholar] [CrossRef]
- Pranoto, Y.; Anggrahini, S.; Efendi, Z. Effect of natural and Lactobacillus plantarum fermentation on in-vitro protein and starch digestibilities of sorghum flour. Food Biosci. 2013, 2, 46–52. [Google Scholar] [CrossRef]
- Klaenhammer, T.R.; Barrangou, R.; Buck, B.L.; Azcarate-Peril, M.A.; Altermann, E. Genomic features of lactic acid bacteria effecting bioprocessing and health. FEMS Microbiol. Rev. 2005, 29, 393–409. [Google Scholar] [CrossRef]
- Ayala-Niño, A.; Rodríguez-Serrano, G.M.; Jiménez-Alvarado, R.; Bautista-Avila, M.; Sánchez-Franco, J.A.; González-Olivares, L.G.; Cepeda-Saez, A. Bioactivity of peptides released during lactic fermentation of amaranth proteins with potential cardiovascular protective effect: An in vitro study. J. Med. Food 2019, 22, 976–981. [Google Scholar] [CrossRef]
- Carbonaro, M.; Maselli, P.; Nucara, A. Relationship between digestibility and secondary structure of raw and thermally treated legume proteins: A Fourier transform infrared (FT-IR) spectroscopic study. Amino Acids 2012, 43, 911–921. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Y.; Jiang, L.; Qi, B.; Zhou, L. Relationship between secondary structure and surface hydrophobicity of soybean protein isolate subjected to heat treatment. J. Chem. 2014, 2014, 475389. [Google Scholar] [CrossRef] [Green Version]
- Salazar-Villanea, S.; Hendriks, W.H.; Bruininx, E.; Gruppen, H.; Van Der Poel, A.F.B. Protein structural changes during processing of vegetable feed ingredients used in swine diets: Implications for nutritional value. Nutr. Res. Rev. 2016, 29, 126–141. [Google Scholar] [CrossRef] [Green Version]
- Liyana-Pathirana, C.M.; Shahidi, F. Importance of insoluble-bound phenolics to antioxidant properties of wheat. J. Agric. Food Chem. 2006, 54, 1256–1264. [Google Scholar] [CrossRef]
- Adebo, O.A.; Gabriela Medina-Meza, I. Impact of fermentation on the phenolic compounds and antioxidant activity of whole cereal grains: A mini review. Molecules 2020, 25, 927. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Wang, C.; Guo, G.; Huo, W.J.; Zhang, Y.L.; Pei, C.X.; Zhang, S.L.; Wang, H. Effects of branched-chain volatile fatty acids supplementation on growth performance, ruminal fermentation, nutrient digestibility, hepatic lipid content and gene expression of dairy calves. Anim. Feed Sci. Technol. 2018, 237, 27–34. [Google Scholar] [CrossRef]
- Heiniö, R.-L.; Noort, M.W.J.; Katina, K.; Alam, S.A.; Sozer, N.; De Kock, H.L.; Hersleth, M.; Poutanen, K. Sensory characteristics of wholegrain and bran-rich cereal foods–A review. Trends Food Sci. Technol. 2016, 47, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Del Hierro, J.N.; Herrera, T.; García-Risco, M.R.; Fornari, T.; Reglero, G.; Martin, D. Ultrasound-assisted extraction and bioaccessibility of saponins from edible seeds: Quinoa, lentil, fenugreek, soybean and lupin. Food Res. Int. 2018, 109, 440–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, K.W.; Iqbal, S.; Khong, N.M.H.; Ooi, D.-J.; Ismail, M. Antioxidant activity of phenolics–saponins rich fraction prepared from defatted kenaf seed meal. LWT 2014, 56, 181–186. [Google Scholar] [CrossRef]
- Lai, L.-R.; Hsieh, S.-C.; Huang, H.-Y.; Chou, C.-C. Effect of lactic fermentation on the total phenolic, saponin and phytic acid contents as well as anti-colon cancer cell proliferation activity of soymilk. J. Biosci. Bioeng. 2013, 115, 552–556. [Google Scholar] [CrossRef]
- Gulitz, A.; Stadie, J.; Wenning, M.; Ehrmann, M.A.; Vogel, R.F. The microbial diversity of water kefir. Int. J. Food Microbiol. 2011, 151, 284–288. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Secondary Protein Components | Peak (cm−1) | Protein Samples | p-Value | ||||
|---|---|---|---|---|---|---|---|
| LP | CP | LCPC 1.0:0.1 | LCPC 1.0:0.5 | LCPC 1:1 | |||
| β-Sheet | |||||||
| 1614.42 | 9.07 | 13.23 | 11.30 | 12.09 | 12.28 | ||
| 1622.13 | 7.65 | 8.00 | 8.54 | 8.81 | 9.06 | ||
| 1633.71 | 11.04 | 10.75 | 11.60 | 11.82 | 12.07 | ||
| β-Sheet (Ʃ) | 27.76 a | 31.97 a | 31.44 a | 32.72 a | 33.40 a | >0.05 | |
| Random coils | 1645.28 | 15.78 d | 16.00 e | 15.04 b | 14.88 a | 15.16 c | <0.05 |
| α-Helix | 1654.07 | 18.48 d | 13.48 c | 9.76 b | 9.48 a | 9.30 a | <0.05 |
| β-Turn | |||||||
| 1668.43 | 10.42 | 8.18 | 12.31 | 11.61 | 11.55 | ||
| 1681.93 | 11.11 | 13.82 | 11.10 | 10.79 | 10.74 | ||
| 1693.50 | 16.45 | 16.55 | 20.34 | 20.52 | 19.84 | ||
| β-Turn (Ʃ) | 37.98 a | 38.55 a | 43.75 a | 42.91 a | 42.13 a | >0.05 | |
| Fermentation Period | p-Value | ||||||
|---|---|---|---|---|---|---|---|
| Day 0 | Day 1 | Day 2 | Day 3 | Day 4 | Day 5 | ||
| pH | 7.09 ± 0.03 a | 4.53 ± 0.03 b | 4.04 ± 0.05 c | 3.94 ± 0.01 d | 3.89 ± 0.01 e | 3.83 ± 0.01 f | <0.05 |
| TSS | 2.13 ± 0.05 a | 1.40 ± 0.00 b | 1.30 ± 0.00 c | 1.20 ± 0.00 d | 1.20 ± 0.00 d | 1.10 ± 0.00 e | <0.05 |
| Protein digestibility | 79.53 ± 0.43 b | 80.60 ± 0.79 b | 84.42 ± 1.47 a | 85.33 ± 0.88 a | 86.39 ± 1.80 a | 86.79 ± 0.43 a | <0.05 |
| TSC | 46.20 ± 0.05 a | 45.30 ± 1.11 a | 41.23 ± 0.60 b | 38.50 ± 0.51 c | 37.93 ± 0.64 cd | 37.13 ± 0.92 d | <0.05 |
| Sugars | |||||||
| Fructose | 0.59 ± 0.02 c | 2.43 ± 0.09 a | 1.11 ± 0.04 b | 0.50 ± 0.02 d | 0.24 ± 0.01 e | 0.07 ± 0.01 f | <0.05 |
| Glucose | ND | 1.24 ± 0.02 b | 2.72 ± 0.04 a | 1.05 ± 0.02 c | 0.42 ± 0.01 b | 0.20 ± 0.00 a | <0.05 |
| Sucrose | 5.41 ± 0.14 a | 1.06 ± 0.02 b | 0.14 ± 0.01 c | 0.07 ± 0.00 cd | ND | ND | <0.05 |
| Secondary Protein Components | Peak (cm−1) | Fermentation Period | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| Day 0 | Day 1 | Day 2 | Day 3 | Day 4 | Day 5 | |||
| β-Sheet | ||||||||
| 1614.42 | 8.51 | 11.30 | 11.48 | 11.50 | 11.28 | 11.25 | ||
| 1622.13 | 11.58 | 8.91 | 8.98 | 9.06 | 8.88 | 8.83 | ||
| 1633.71 | 15.32 | 11.76 | 11.85 | 11.80 | 11.51 | 11.58 | ||
| β-Sheet (Ʃ) | 30.90 a | 31.97 a | 32.31 a | 32.34 a | 31.67 a | 31.66 a | >0.05 | |
| RC (Ʃ) | 1645.28 | 15.32 f | 15.83 e | 17.65 d | 18.85 c | 20.43 b | 21.67 a | <0.05 |
| α-Helix (Ʃ) | 1654.07 | 9.90 a | 9.71 b | 7.71 c | 7.63 d | 7.25 e | 6.58 f | <0.05 |
| β-Turn | ||||||||
| 1668.43 | 12.29 | 12.41 | 13.23 | 13.38 | 13.03 | 13.51 | ||
| 1681.93 | 11.35 | 11.73 | 12.69 | 12.85 | 12.70 | 11.21 | ||
| 1693.50 | 20.24 | 18.33 | 16.40 | 14.94 | 14.90 | 15.3 | ||
| β-Turn (Ʃ) | 43.87 a | 42.47 a | 42.32 a | 41.17 a | 40.63 a | 40.07 a | >0.05 | |
| Ratio (α-helix:β-sheet) | 32.04 | 30.38 | 23.87 | 23.58 | 22.90 | 20.81 | ||
| Fermentation Period [Day] | p-Value | ||||||
|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | 5 | ||
| TPC | 394.46 ± 3.72 d | 516.83 ± 8.95 b | 624.88 ± 7.23 a | 458.33 ± 11.51 c | 411.17 ± 11.74 d | 405.80 ± 14.21 d | <0.05 |
| Phenolic Compounds | |||||||
| Catechin | 34.03 ± 0.48 f | 61.90 ± 0.54 b | 63.08 ± 0.77 a | 59.53 ± 0.21 c | 56.49 ± 0.14 d | 51.74 ± 0.39 e | <0.05 |
| Chlorogenic | 84.93 ± 0.98 e | 89.61 ± 0.76 bc | 105.03 ± 0.42 a | 90.62 ± 0.29 b | 88.45 ± 1.11 cd | 87.34 ± 0.76 d | <0.05 |
| Epicatechin | 106.85 ± 0.54 f | 127.87 ± 0.57 b | 157.55 ± 0.87 a | 122.05 ± 0.67 a | 116.31 ± 0.25 d | 112.33 ± 0.28 e | <0.05 |
| Quercetin | 14.05 ± 0.68 c | 14.16 ± 0.44 c | 15.07 ± 0.18 c | 14.55 ± 0.56 bc | 16.05 ± 0.34 a | 3.04 ± 0.10 d | <0.05 |
| Rutin | 2.43 ± 0.05 c | 1.90 ± 0.08 d | 2.27 ± 0.04 cd | 4.50 ± 0.23 a | 4.37 ± 0.07 a | 3.69 ± 0.49 b | <0.05 |
| Caffeic acid | ND | ND | ND | ND | 4.10 ± 0.05 b | 5.29 ± 0.04 a | <0.05 |
| Ferulic acid | 3.77 ± 0.05 b | 3.86 ± 0.07 a | 4.14 ± 0.04 a | 4.04 ± 0.04 a | 3.83 ± 0.11 b | 3.81 ± 0.08 b | <0.05 |
| Gallic acid | 11.76 ± 0.90 e | 19.44 ± 0.43 b | 20.51 ± 0.39 b | 27.67 ± 0.59 a | 15.85 ± 0.84 c | 14.02 ± 0.44 d | <0.05 |
| Sinapic acid | 0.00 | 1.61 ± 0.04 ab | 1.41 ± 0.11 a | 1.15 ± 0.03 ab | 1.00 ± 0.03 ab | 0.90 ± 0.02 b | <0.05 |
| Synergic acid | 46.90 ± 0.97 b | 46.96 ± 0.51 b | 50.27 ± 0.48 a | 47.60 ± 0.87 b | 47.45 ± 1.16 b | 47.28 ± 2.50 b | <0.05 |
| Phenolic Compounds (Ʃ) | 304.71 f | 367.30 c | 419.33 a | 371.72 b | 353.88 d | 329.46 e | <0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alrosan, M.; Tan, T.-C.; Easa, A.M.; Alu’datt, M.H.; Tranchant, C.C.; Almajwal, A.M.; Gammoh, S.; Maghaydah, S.; Dheyab, M.A.; Jameel, M.S.; et al. Improving the Functionality of Lentil–Casein Protein Complexes through Structural Interactions and Water Kefir-Assisted Fermentation. Fermentation 2023, 9, 194. https://doi.org/10.3390/fermentation9020194
Alrosan M, Tan T-C, Easa AM, Alu’datt MH, Tranchant CC, Almajwal AM, Gammoh S, Maghaydah S, Dheyab MA, Jameel MS, et al. Improving the Functionality of Lentil–Casein Protein Complexes through Structural Interactions and Water Kefir-Assisted Fermentation. Fermentation. 2023; 9(2):194. https://doi.org/10.3390/fermentation9020194
Chicago/Turabian StyleAlrosan, Mohammad, Thuan-Chew Tan, Azhar Mat Easa, Muhammad H. Alu’datt, Carole C. Tranchant, Ali Madi Almajwal, Sana Gammoh, Sofyan Maghaydah, Mohammed Ali Dheyab, Mahmood S. Jameel, and et al. 2023. "Improving the Functionality of Lentil–Casein Protein Complexes through Structural Interactions and Water Kefir-Assisted Fermentation" Fermentation 9, no. 2: 194. https://doi.org/10.3390/fermentation9020194