1. Introduction
Probiotics are living microorganisms. When consumed in appropriate amounts, they can provide health benefits to the host, including protecting the intestines, anticancer effects, immune regulation, lowering blood lipids, controlling blood sugar, and promoting the absorption of nutrients. In the treatment of Ulcerative colitis (UC), potential mechanisms include inhibiting the growth of pathogenic bacteria, improving epithelial mucosal barrier function, regulating immunity, and reducing the secretion of pro-inflammatory factors [
1,
2,
3].
Lycium barbarum L. (L), belonging to the Solanaceae Lycium genus, is renowned for its antioxidant, anti-inflammatory, and anti-cancer properties [
4]. UC is a type of inflammatory bowel disease (IBD) that begins in the rectum and typically extends proximally to involve the entire colon. Its main pathological manifestations include intestinal barrier damage, loss of crypts, crypt abscesses, deformation of mucosal glands, infiltration of inflammatory cells, and loss of goblet cells. Currently, aminosalicylate drugs, including SASP and mesalazine, are commonly used for the treatment of UC. However, these drugs have potential adverse reactions such as drowsiness, gastrointestinal complications, and renal dysfunction. L-arginine plays a role in immune regulation and nitric oxide synthesis in UC [
5,
6,
7]. In this study, probiotics were combined with L to create a lyophilized bacterial powder through fermentation, which exhibits potential probiotic efficacy. Dextran sodium sulfate (DSS)-induced UC in mice was used as a model to investigate the effects of fermented L lyophilized powder (LP+
Ly) on the intestinal flora and metabolite profile. This study integrated immunohistochemistry, quantitative reverse-transcription PCR (qRT-PCR), macrogenomics, and metabolomics to explore the potential therapeutic value of LP+
Ly and provide new insights for the prevention and alleviation of UC.
2. Materials and Methods
2.1. Preparation of LP+Ly
The dried fruits of L were soaked and pulped in pure water at a mass ratio of 1:5 with 0.05% sodium D-isoascorbate for color protection. Allow it to soak at room temperature for 8–9 h. Then, add 0.15% pectinase and blend. Filter out the seeds using three layers of sterile gauze and a funnel. Next, add 5% white sugar and 10% citric acid. with a pH of 4.5, followed by pasteurization. The sterilized and cooled goji juice was inoculated with 5% (
v/
v) of
Lactiplantibacillus plantarum strain NXU0011 and fermented at 37 °C for 12 h. It was then subjected to vacuum freeze-drying for 30 h and ground to obtain the final product, denoted as LP+
Ly [
8].
2.2. Animal Experimental Design
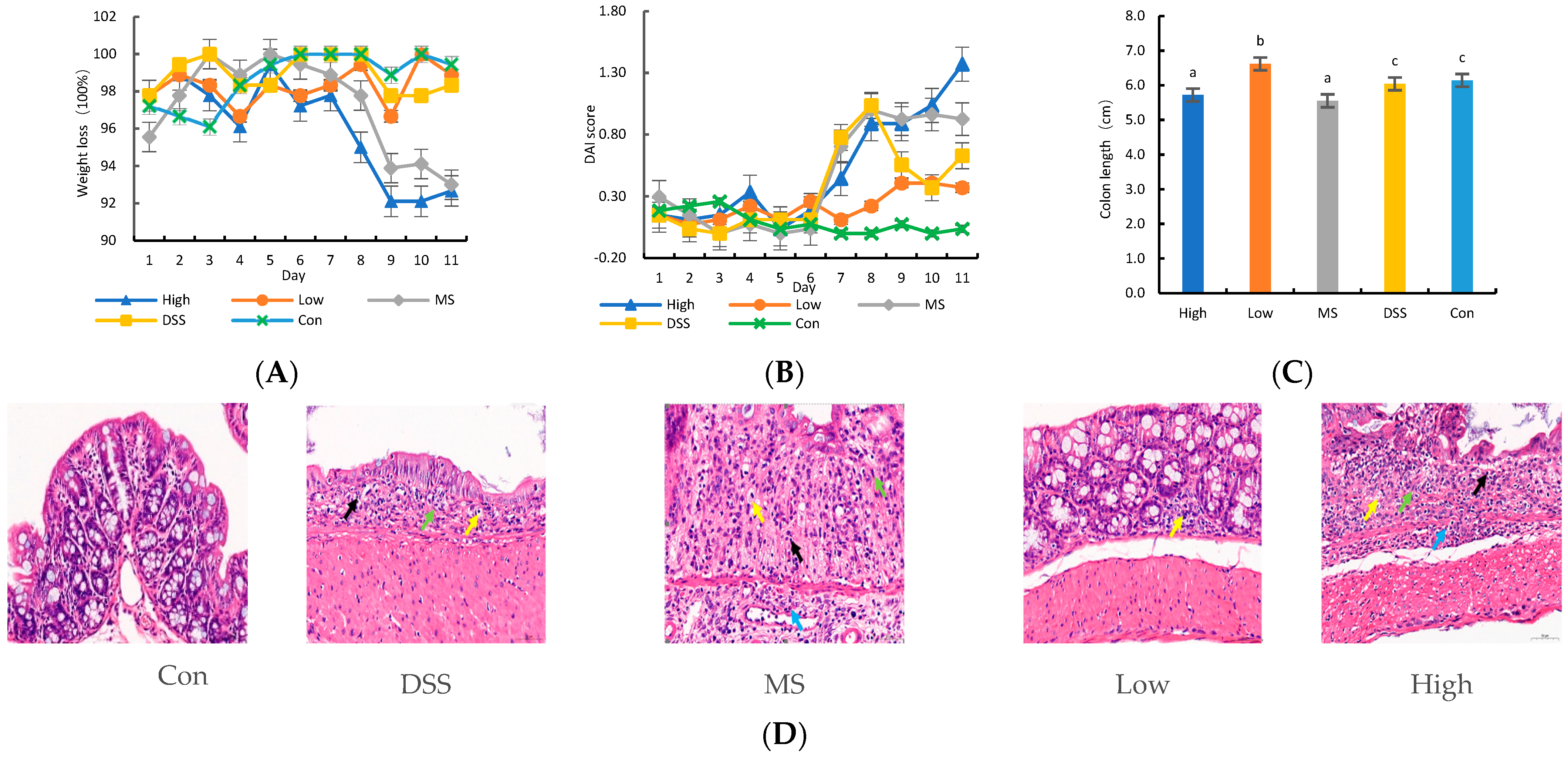
Thirty male SPF-grade C57BL/6J mice aged 6–8 weeks and free of specific pathogens were randomly divided into five groups: control (Con) group, model (DSS) group, positive drug (MS) group, LP+Ly high-dose (High) group, and LP+Ly low-dose (Low) group. The mice were acclimated and fed for 3 days. The Con group was given sterile water, while the other groups received sterile water containing 5% DSS (w/v) for 7 days to induce UC. Subsequently, the Con and DSS groups were administered 0.2 mL/day of phosphate-buffered saline (PBS) via gavage, the MS group received mesalazine dissolved in PBS at 40 mg/kg, and the High and Low groups were given 2 × 109 CFU/mL and 2 × 106 CFU/mL, respectively, of viable bacteria dissolved in PBS. On day 11, all mice were euthanized, and blood, colonic tissue, and feces were collected. The animal experiments were approved by the Experimental Animal Ethics Committee of Sichuan Lilaisinuo Biological Technology Co., Ltd. (Approval No.: LLSN-2022014).
2.3. Disease Activity Index (DAI)
During the modeling phase, the body weight, fecal characteristics, and fecal bleeding symptoms of each group of mice were observed and recorded daily. The DAI score was calculated by dividing the total score of the above three parameters by 3 [
9]. The scoring criteria are provided in
Table A1.
2.4. Histological Assessment
The colonic length was measured in all animals, followed by fixation in a 10% formalin solution for 48 h. The samples were dehydrated, embedded in paraffin, sectioned, and stained with hematoxylin and eosin (H&E). Microscopic observations were conducted to assess the pathological changes, and images were captured. The histological evaluation was based on the severity of inflammation, crypt damage, and the extent of pathological alterations [
10,
11].
2.5. RNA Isolation and qRT-PCR
RNA from colon tissue was extracted using TRIzol, and cDNA was synthesized through reverse transcription. qRT-PCR was performed using the Ultra SYBR Mixture reagent kit, with GAPDH as the internal reference gene. The primer sequences are provided in
Table 1.
The reaction system was prepared with a total volume of 20 μL, as described in
Table A2. Gene expression levels were determined using the relative quantification method, specifically the 2
−ΔΔCt method.
2.6. Immunohistochemistry Staining
Paraffin sections of the colon tissue from each experimental group were dried in a constant-temperature oven at 60 °C for 1 h. The sections were then washed three times with 0.01 mol·L−1 PBS solution for 5 min each, followed by washing with absolute ethanol for 10 min and two rinses with distilled water. Antigen retrieval was performed by heating the sections at 100 °C for 8 min, and the sections were then treated with 3% hydrogen peroxide for 30 min. The sections were incubated overnight at 4 °C with p-STAT3 and IL-6 antibodies, followed by incubation with a secondary antibody. DAB staining was performed, followed by counterstaining with hematoxylin. The sections were observed under a microscope, and micro-images were captured at 400× magnification.
2.7. DNA Sequencing and Gut Microbiota Analysis
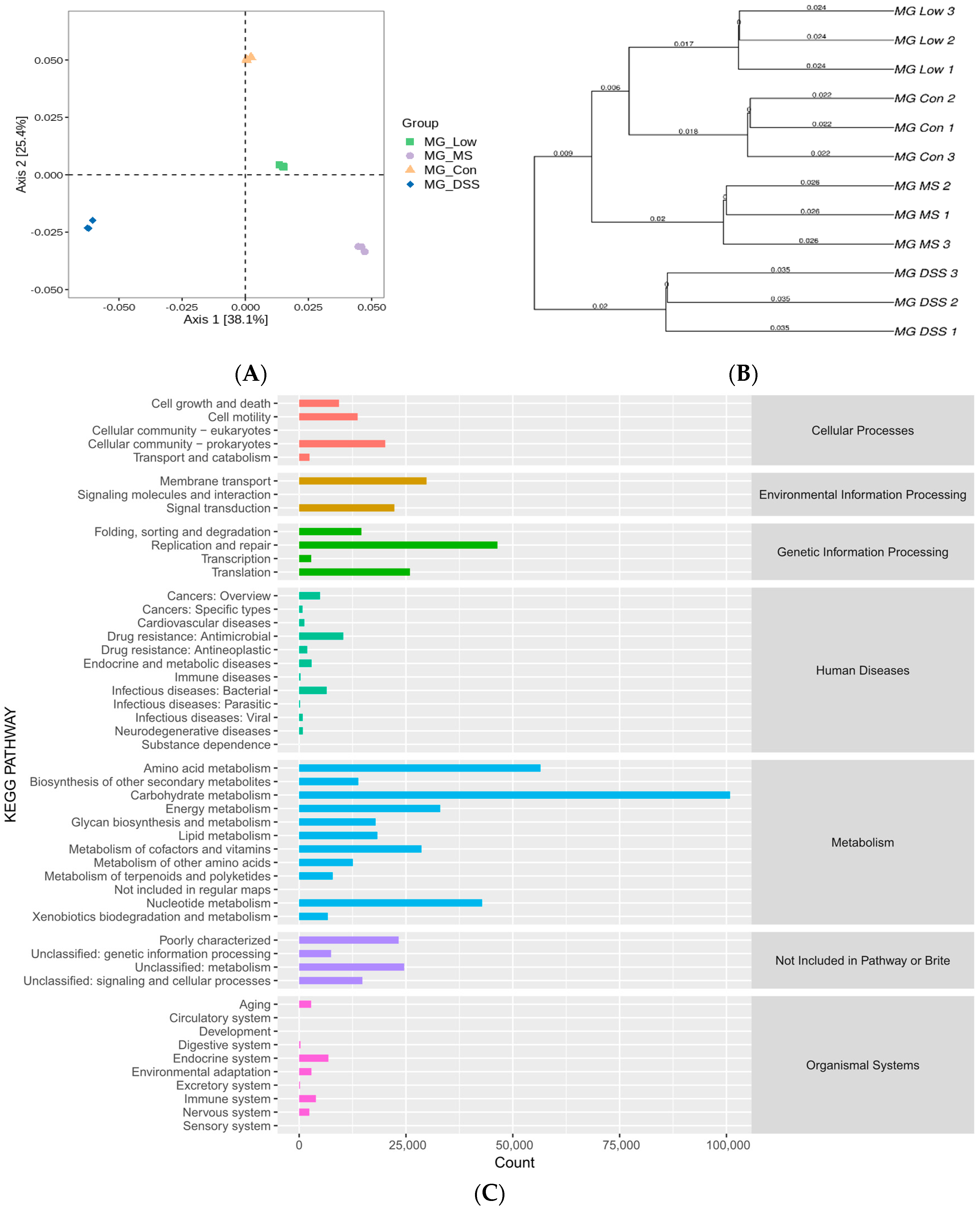
Total microbial genomic DNA samples were extracted using the OMEGA Mag-Bind Soil DNA Kit (M5635-02; Omega Bio-Tek, Norcross, GA, USA) according to the manufacturer’s instructions and stored at −20 °C for further analysis. The quantity and quality of the extracted DNA were assessed using a Qubit™ 4 Fluorometer (Invitrogen, Waltham, MA, USA) and agarose gel electrophoresis, respectively. The microbial DNA was then processed to construct metagenome shotgun sequencing libraries with 400-bp insert sizes using the Illumina TruSeq Nano DNA LT Library Preparation Kit. Each library was sequenced on the Illumina NovaSeq platform (Illumina, San Diego, California, USA), employing a PE150 strategy. The resulting sequencing data were analyzed to study the composition and functional changes of the gut microbiota among different groups.
2.8. Plasma Metabolites Analysis via Untargeted Metabolomics
Fasting blood samples were collected into 5-mL Vacutainer tubes containing the chelating agent EDTA and centrifuged at 1500× g and 4 °C for 15 min. Aliquots (150 μL) of plasma were stored at −80 °C until analysis. The plasma samples were thawed at 4 °C, and 100-μL aliquots were mixed with 400 μL of cold methanol/acetonitrile (1:1, v/v) for protein precipitation. The mixture was centrifuged at 14,000× g and 4 °C for 20 min. The supernatant was then dried in a vacuum centrifuge and re-dissolved in 100-μL acetonitrile/water (1:1, v/v) for UHPLC-Q-Exactive Orbitrap mass spectrometry (MS) analysis. The samples were finally centrifuged at 14,000× g and 4° C for 15 min before injection.
2.9. Statistical and Bioinformatics Analysis
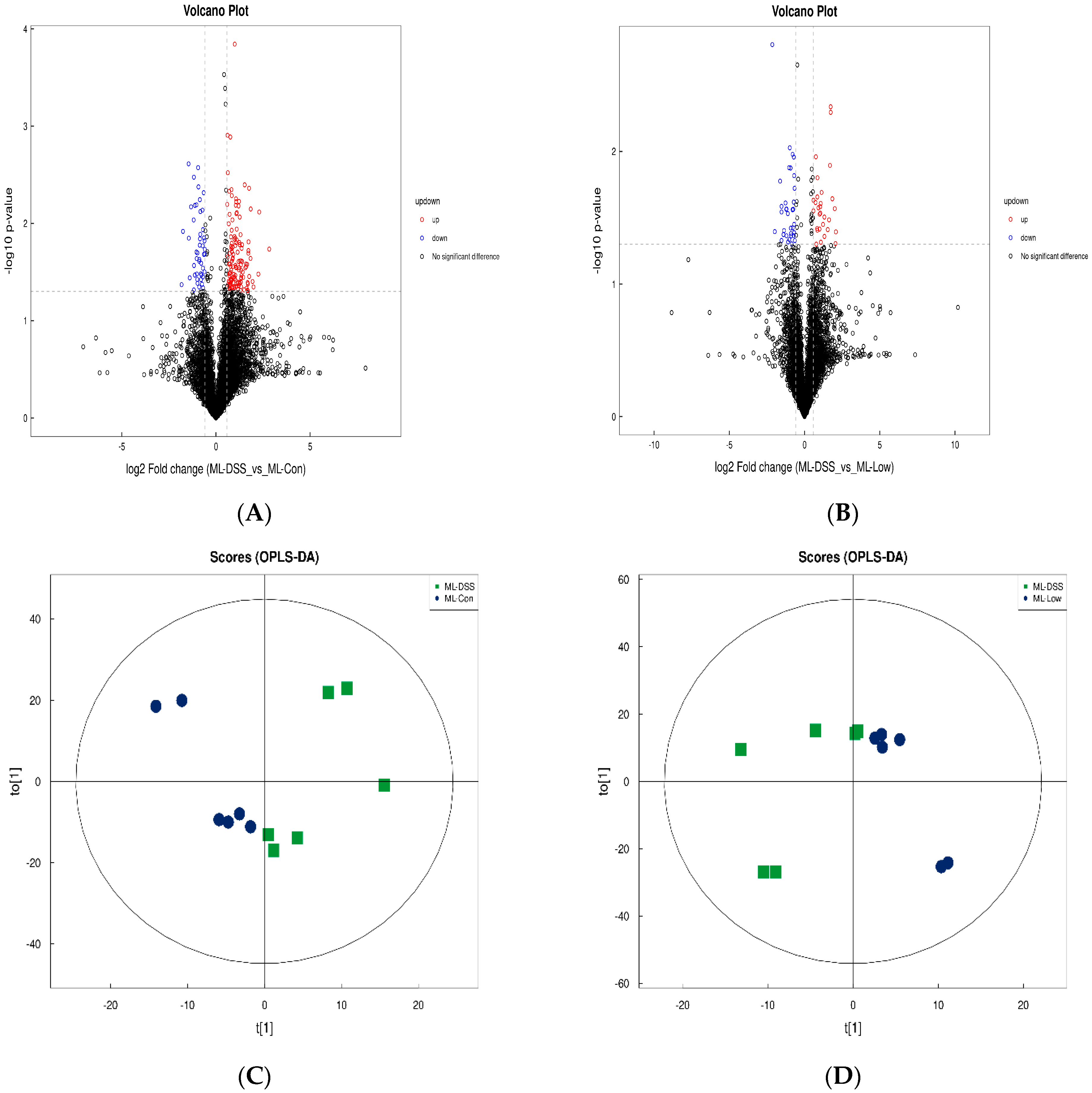
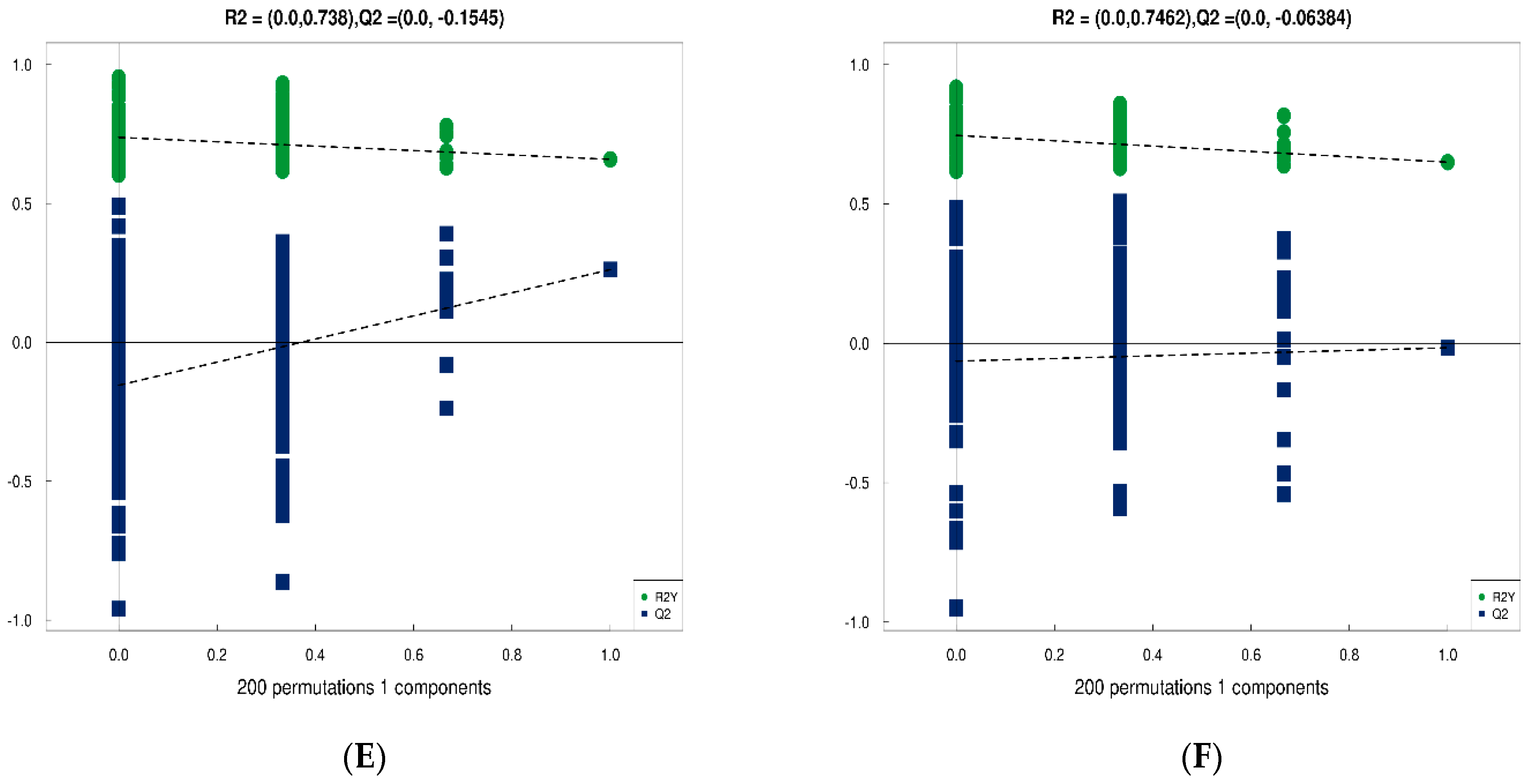
Normalized data were analyzed using GraphPad Prism software (ver. 8.0.2; GraphPad Software Inc., San Diego, CA, USA). Model stability was evaluated using 7-fold cross-validation. The significance of differences between two independent samples was determined using a t-test, with significance criteria of variable importance projection (VIP) > 1 and p < 0.05.
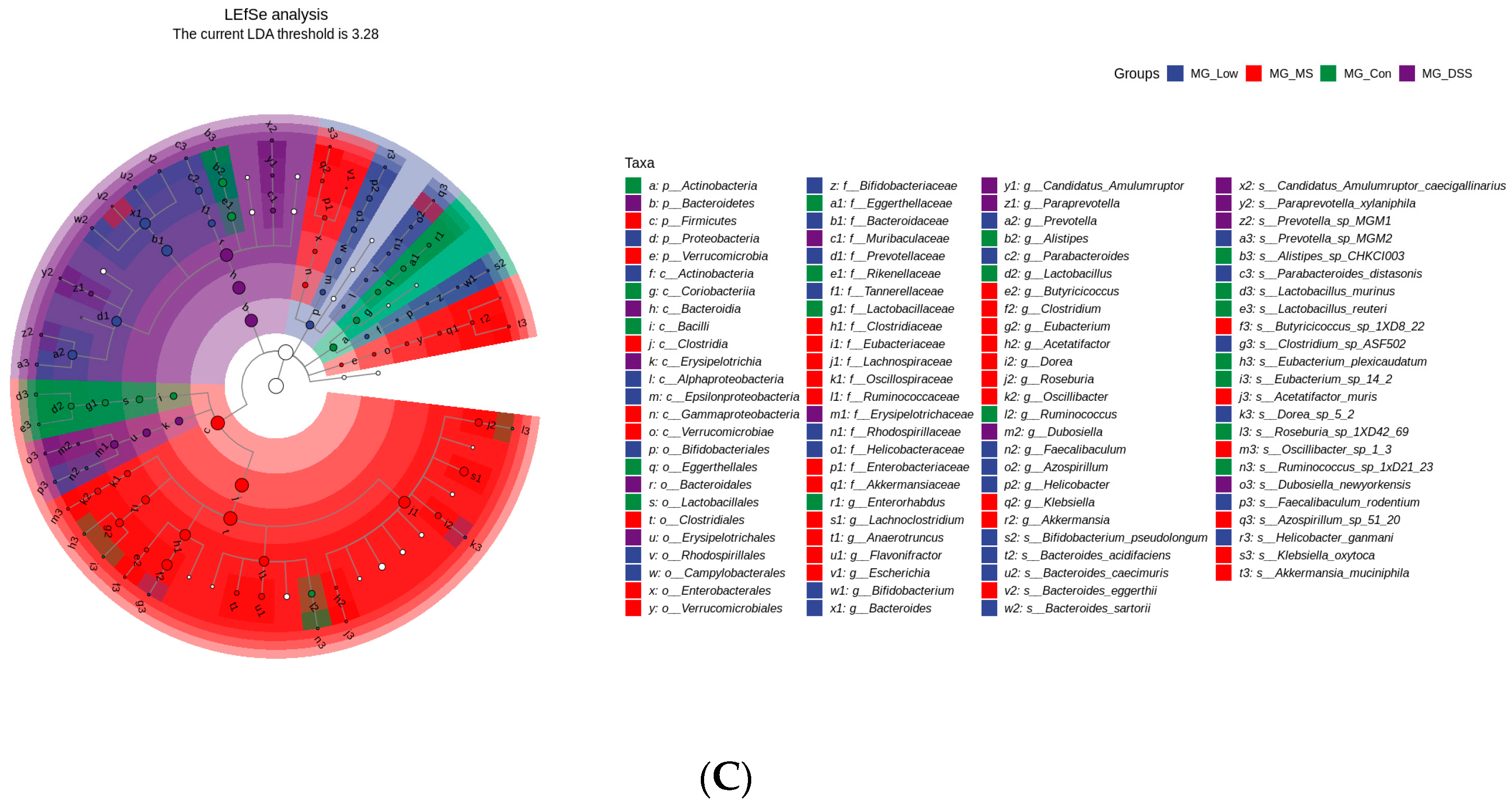
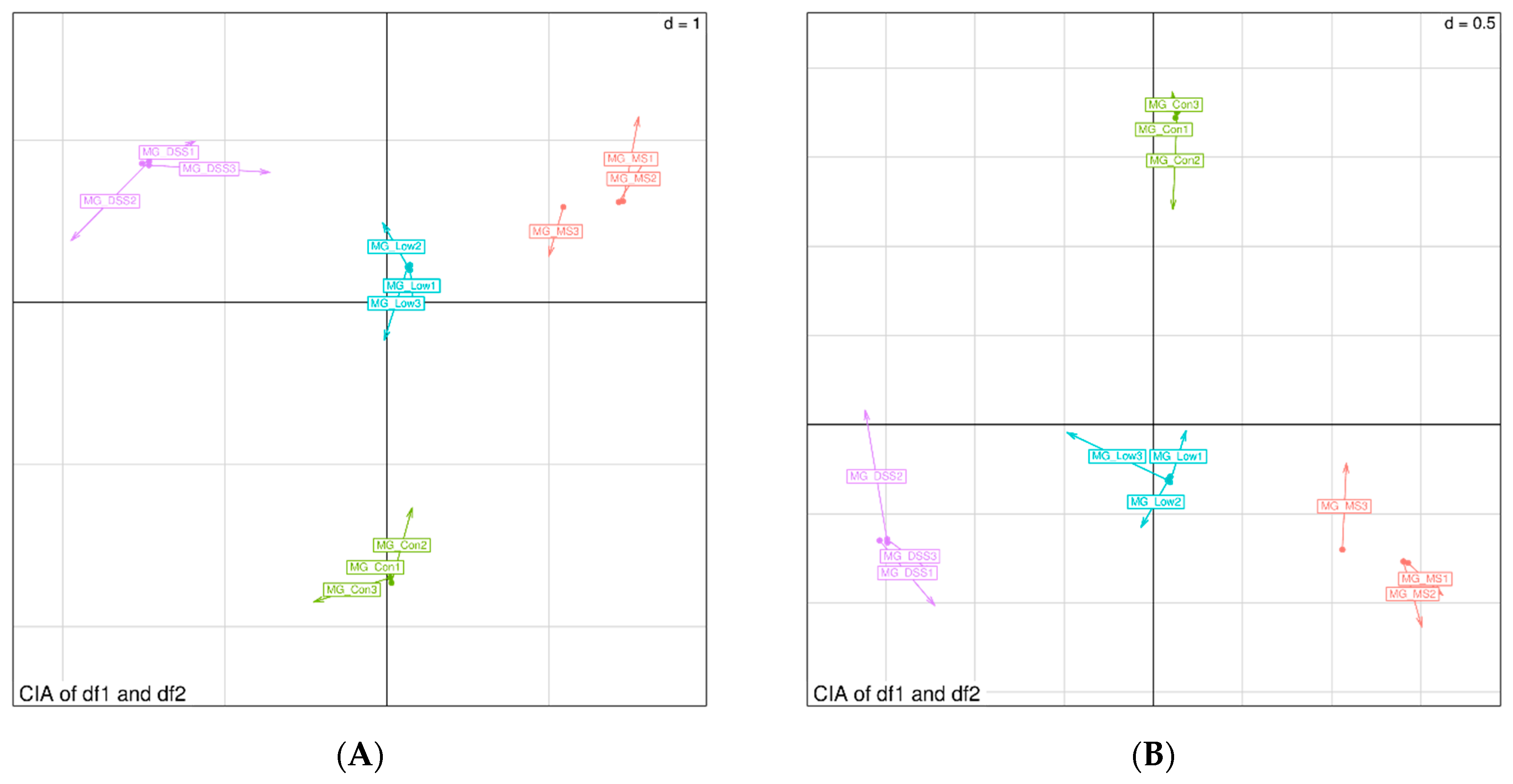
Quality-filtered raw sequencing reads from the metagenomic data were classified for each sample using Kraken2. CDS sequences from all samples were clustered with mmseqs2 at a protein sequence similarity threshold of 0.90. Linear discriminant analysis effect size (LEfSe) was employed to detect taxonomic and functional differences between groups and identify taxa and functions significantly enriched in different groups. Bray-Curtis distance was used to measure the dissimilarity in microbial community composition and function among samples. Principal coordinate analysis (PCoA) was employed for visualizing differences and similarities between samples based on microbial profiles.
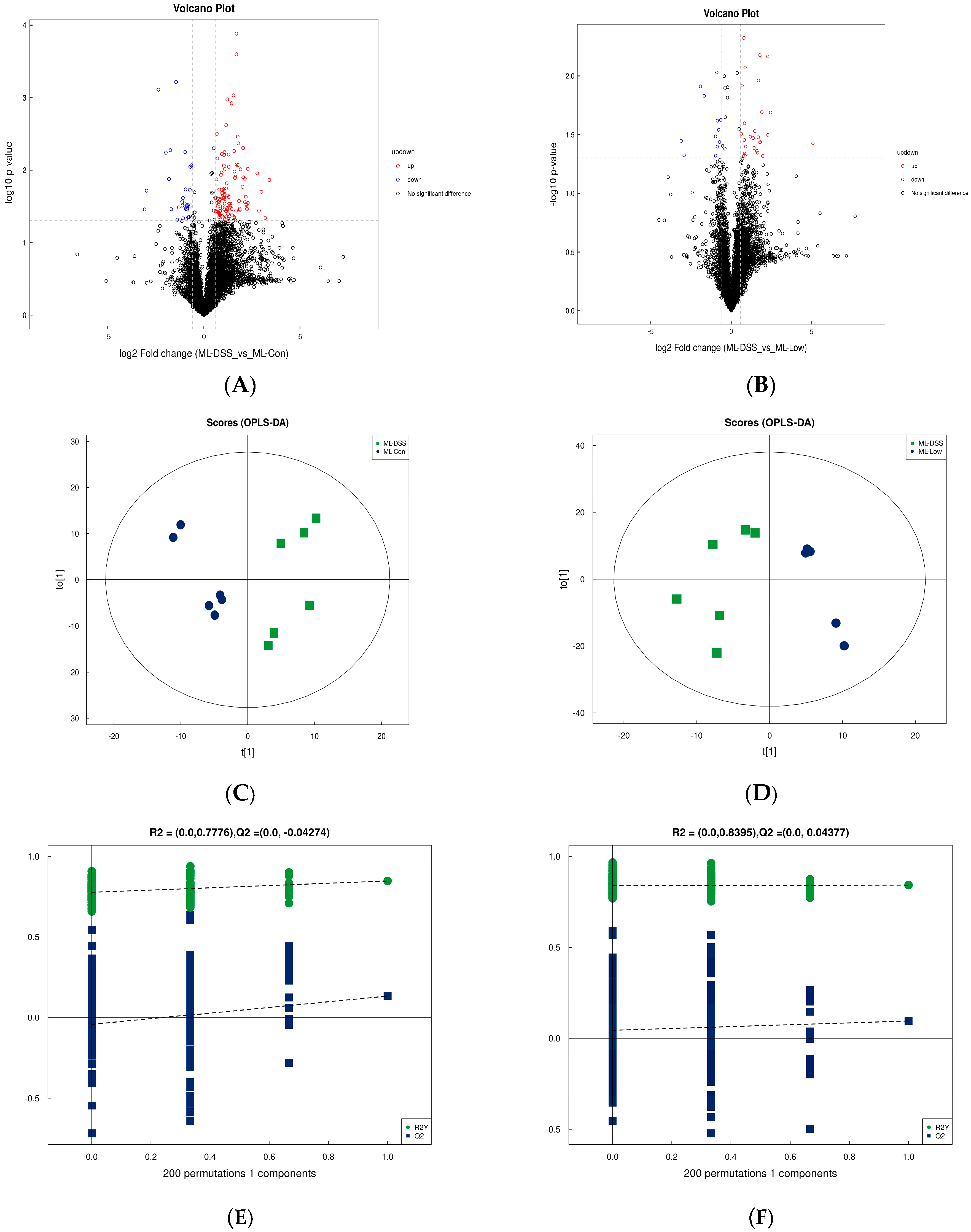
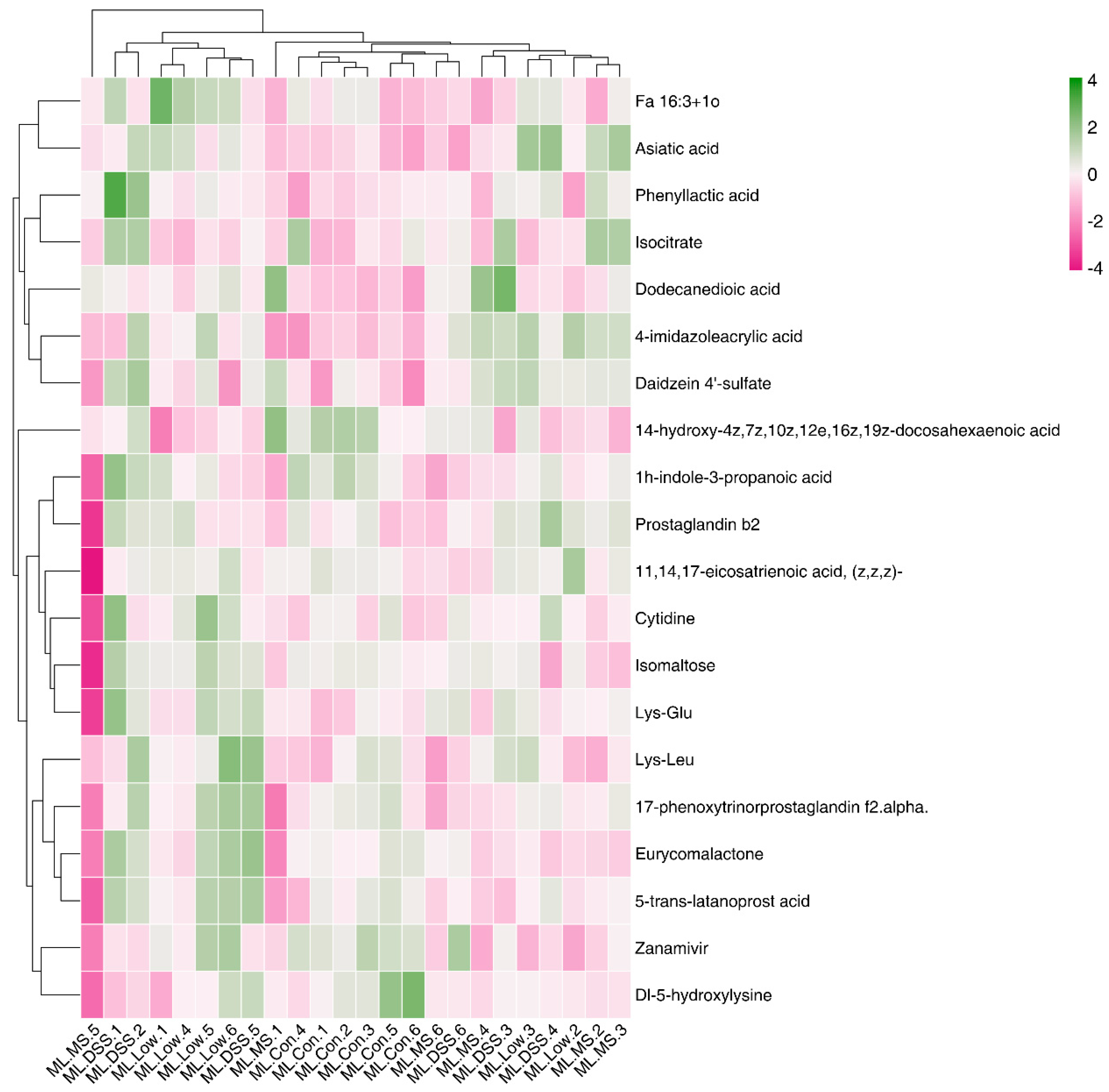
The raw MS data were converted to mzXML format using ProteoWizard MSConvert. The converted data were processed with XCMS software for data extraction. Metabolite structure identification was conducted using a standard compound database. Both univariate and multivariate statistical analyses were used for intergroup differences and differential metabolite analyses. Significantly different metabolites were subjected to cluster analysis, Spearman correlation analysis, and bioinformatics analysis involving Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways.
4. Analysis and Discussion
Plant-based diets have been associated with protective effects against various diseases by reducing oxidative stress, modulating cytokine secretion, and regulating gut microbiota to fortify the intestinal barrier [
17]. In this experiment, we employed microbial fermentation to ferment goji berries with the goal of producing beneficial microorganisms and secondary metabolites. This process was expected to enhance the efficacy of LP+
Ly in alleviating UC.
One of the inflammatory factors, P-STAT3, is indicative of inflammation, where a higher degree of phosphorylation suggests increased inflammation. IL-6, a pro-inflammatory cytokine, can stimulate the phosphorylation of P-STAT3 [
18]
. Additionally, dysbiosis of the gut microbiota due to defects in immune system-related genes can cause an imbalance in the microbial community, consistent with existing studies [
19]. Farraye et al. [
20] also established a significant positive correlation between the expression of the miR-214 gene and the severity of UC in patients.
In our study, through immunohistochemical analysis, we measured the levels of P-STAT3, IL-6, and miR-214 in colonic tissues. Our findings showed that LP+Ly effectively suppressed the mRNA expression of pro-inflammatory cytokines, such as P-STAT3 and IL-6. Consequently, the suppression of pro-inflammatory cytokines by LP+Ly may be essential for mitigating DSS-induced intestinal inflammation and damage.
DSS-induced UC in mice is typified by symptoms such as weight loss, bloody diarrhea, colon shortening, and intestinal damage [
21,
22]. Moreover, UC patients often exhibit a decline in the number of goblet cells in the colon, leading to increased mucosal barrier permeability [
23]. Our study found that LP+
Ly indeed mitigated these pathological symptoms and ameliorated histological damage in UC mice. Nonetheless, LP+
Ly comprises a multifarious mixture of active components. It is therefore plausible that its effectiveness against UC is attributable to the synergistic action of multiple constituents. Further studies are necessary to determine how these components interact to alleviate UC and to identify the primary active components.
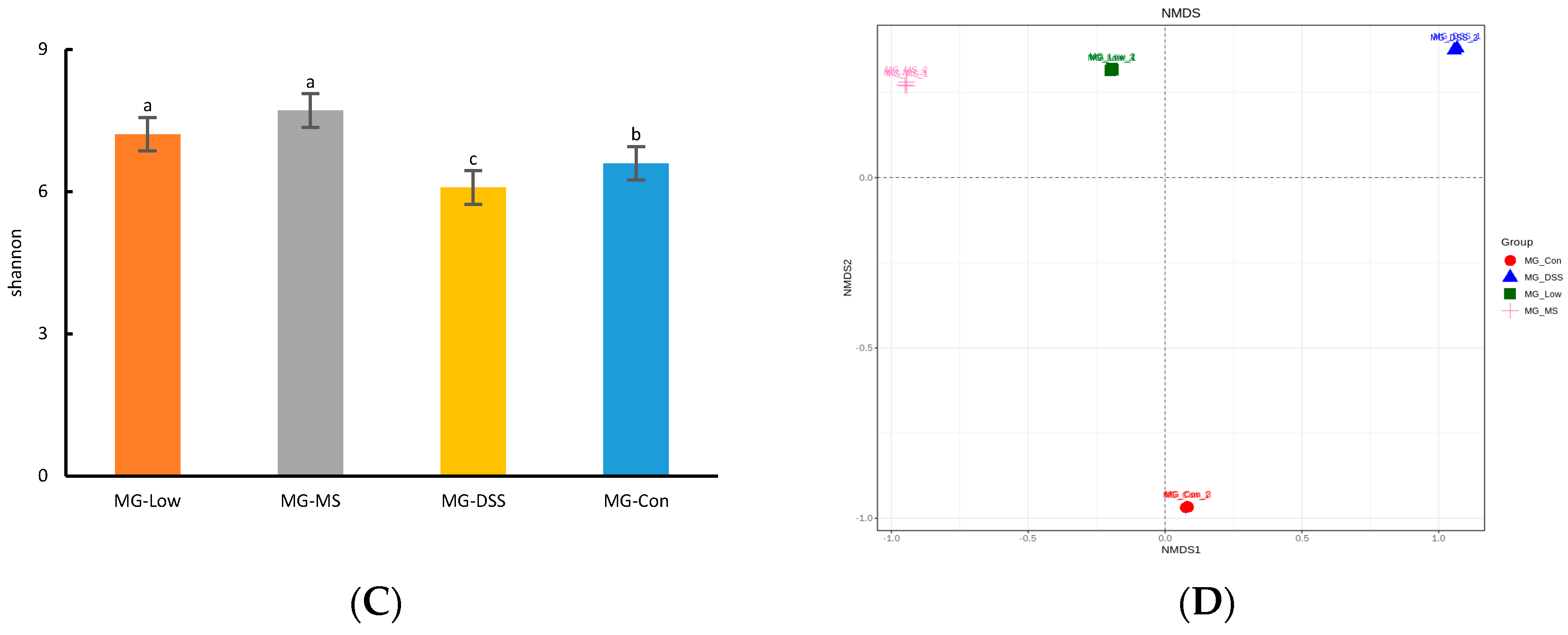
The gut microbiota is implicated in various physiological functions, such as regulating inflammatory responses, controlling appetite, and maintaining intestinal integrity [
24]. Disruption of the gut microbiota, termed dysbiosis, can serve as an important marker of the pathological and physiological state of the host. Our study demonstrated that gut microbiota diversity and richness diminished in the DSS-induced group of mice, as evidenced by a decrease in the species richness, Chao-1 index, Shannon index, and Simpson index. However, the administration of medication and LP+
Ly reversed these trends, with LP+
Ly nearly restoring these indicators to levels observed in healthy mice.
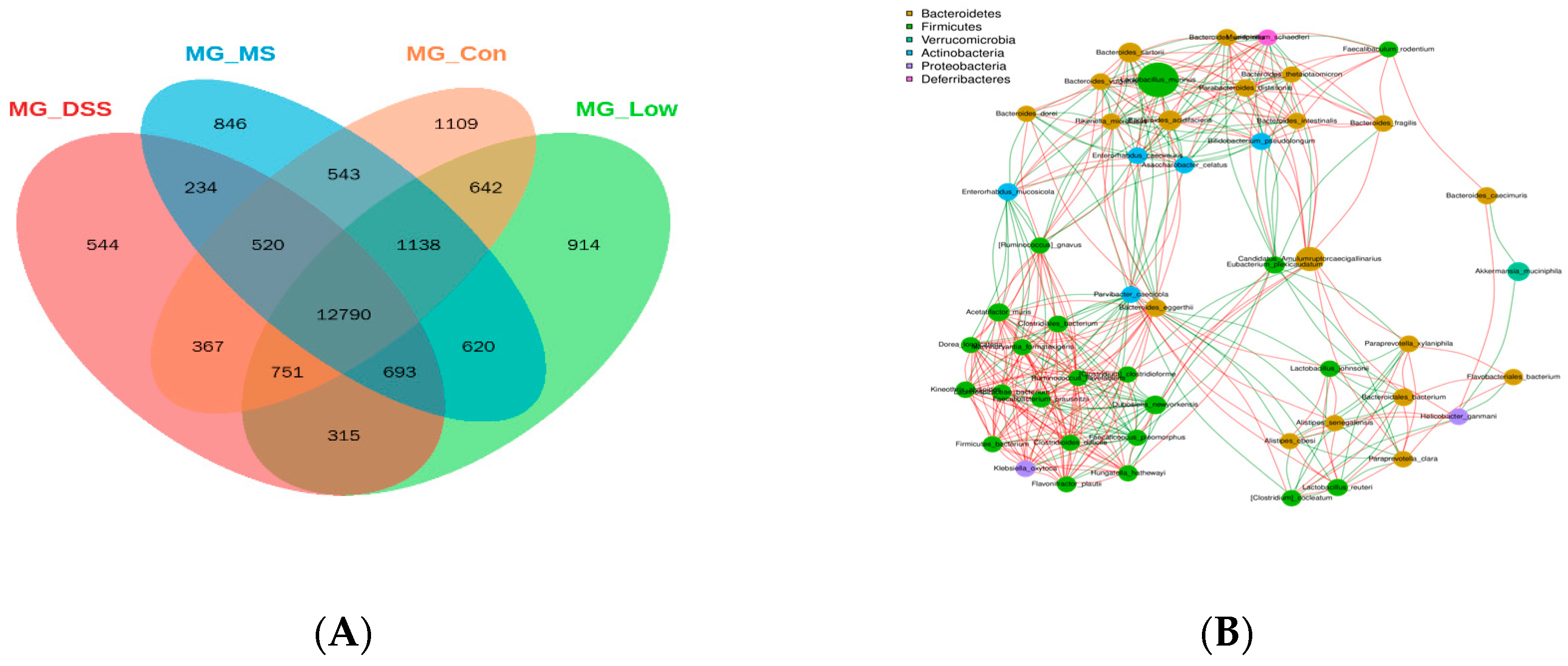
Additionally, LP+
Ly altered the composition of the gut microbiota in UC mice, including changes in the abundance of specific bacterial phyla such as
Bacteroidetes and
Firmicutes. This could be attributed to the polysaccharides present in LP+
Ly, as polysaccharides are known to be effective in modulating the gut microbiota. In this study, the occurrence of UC significantly reduced the relative abundance of
Lactiplantibacillus. Unfortunately, intervention with LP+
Ly did not reverse this decreasing trend. Additionally, LP+
Ly increased the relative abundance of
Helicobacter. It is worth noting that
Helicobacter has been negatively associated with IBD [
25], and its depletion can disrupt the balance of the gut microbiota [
26]. This is consistent with the significant decrease in relative abundance of
Helicobacter seen in the DSS group. At the genus level, the gut microbiota structure was disrupted in the DSS group, with a decreased relative abundance of beneficial bacteria (such as
Lactiplantibacillus,
Clostridium, and
Akkermansia), while administration of LP+
Ly reduced the relative abundance of harmful bacteria (such as
Candidatus Amulumruptor,
Odoribacter, and
Alloprevotella), indicating a broad-spectrum antibacterial effect of LP+
Ly. Moreover, in this experiment, the relative abundance of short-chain fatty acid (SCFA)-producing bacteria, such as
Alistipes,
Clostridium, and
Roseburia, was decreased in the DSS group. Conversely, the relative abundance of SCFA-producing bacteria, including
Bacteroides and
Parabacteroides, was higher in the groups treated with the drug and LP+
Ly compared to the Con group. This indicates that the alleviating effect of LP+
Ly on UC mice partially originates from its modulation of the gut microbiota.
LP+
Ly also influenced the relative abundance of SCFA-producing bacteria, which are crucial for the proliferation of intestinal epithelial cells and the maintenance of epithelial barrier function [
27,
28,
29]. The abundance of such bacteria is reduced in IBD patients, whereas probiotic bacteria that produce SCFAs have been found to alleviate clinical symptoms in these patients [
30,
31].
In recent times, research has highlighted the significant role of dietary amino acids in preventing and treating intestinal inflammation. Amino acids are implicated in protecting the intestine through mechanisms such as inhibiting intestinal epithelial cell apoptosis, alleviating intestinal inflammation, reducing oxidative stress reactions, and restoring intestinal barrier function [
32,
33]. Our study employed metabolomic analysis to investigate the plasma metabolites of UC mice post-intervention with medication and LP+
Ly. The goal was to elucidate the metabolic mechanisms through which LP+
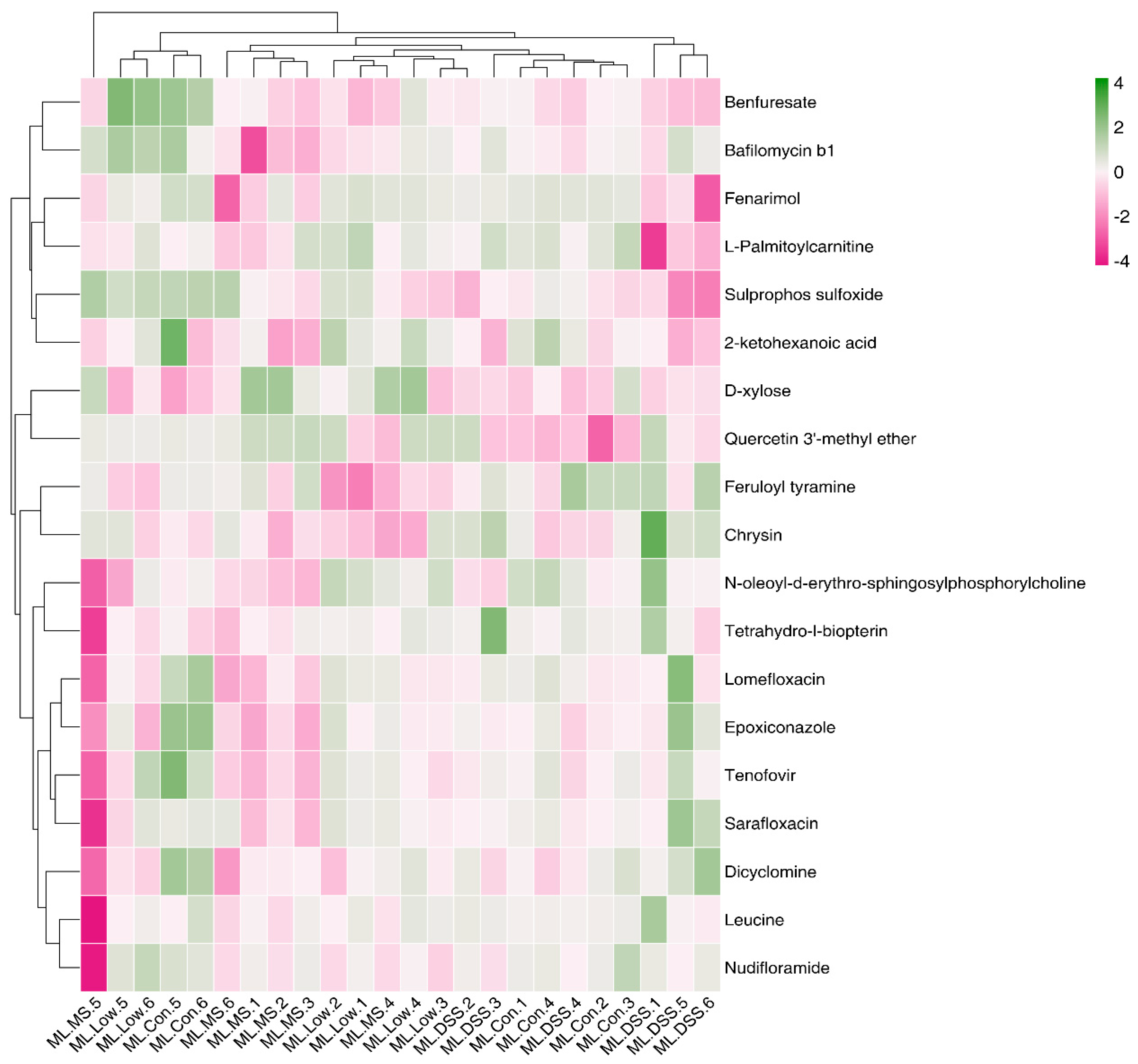
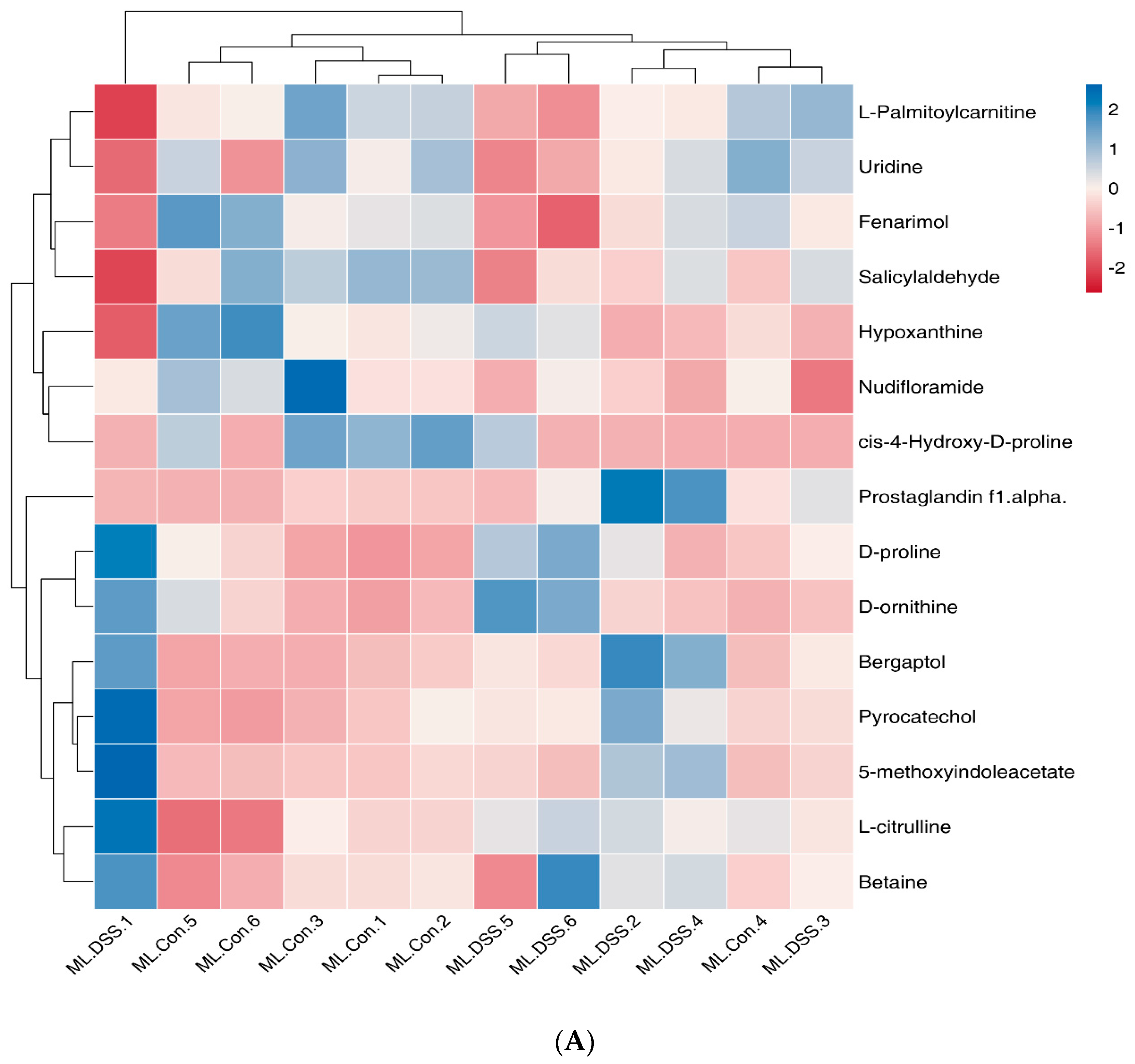
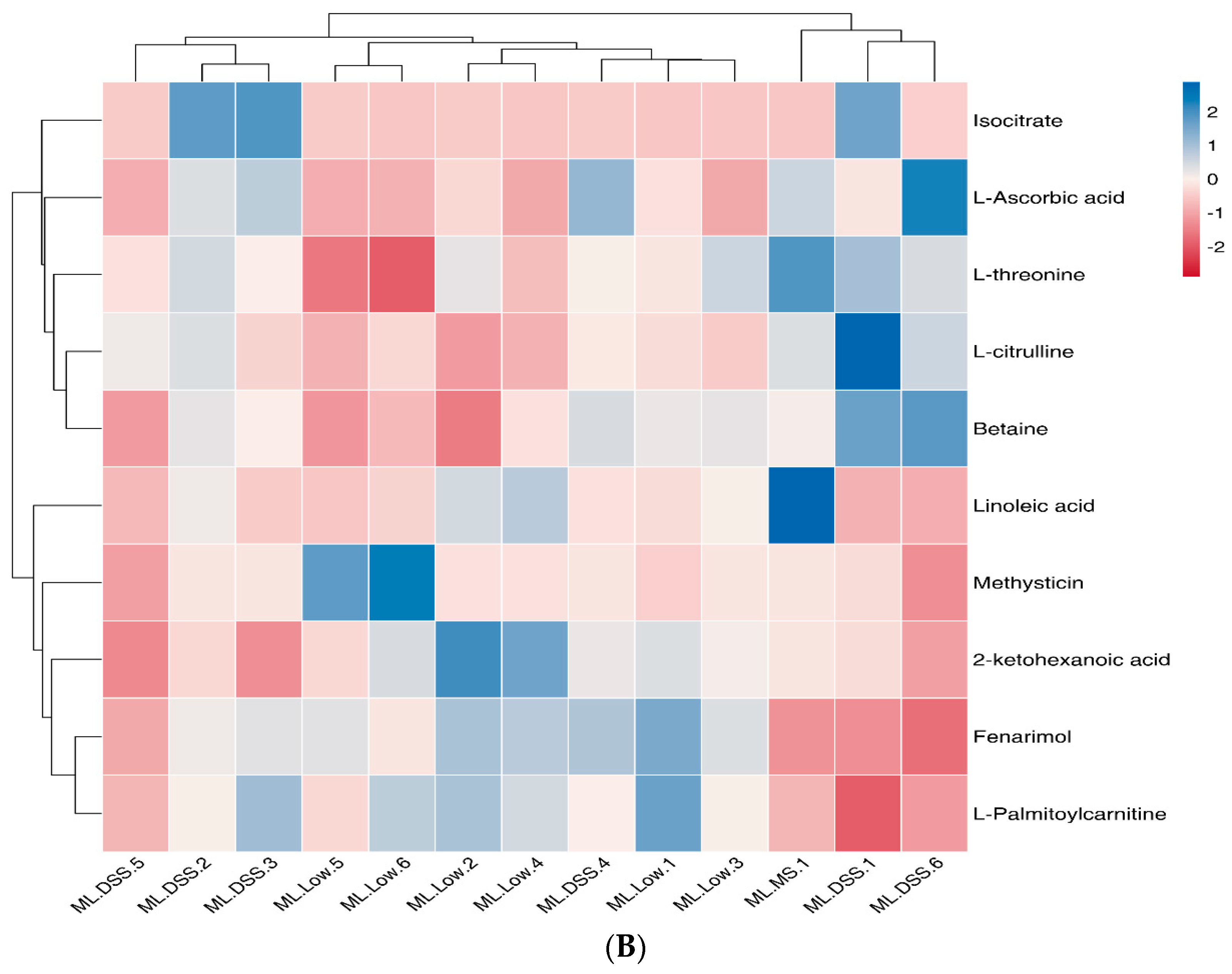
Ly alleviates UC. Analysis of selected metabolites revealed distinct trends in small molecule metabolites such as fenarimol, L-citrulline, and 2-ketohexanoic acid during DSS treatment. These trends indicated significant differences in the relative abundance of metabolites among the groups. UC mice displayed significant metabolic disturbances, which LP+
Ly was able to partially normalize.
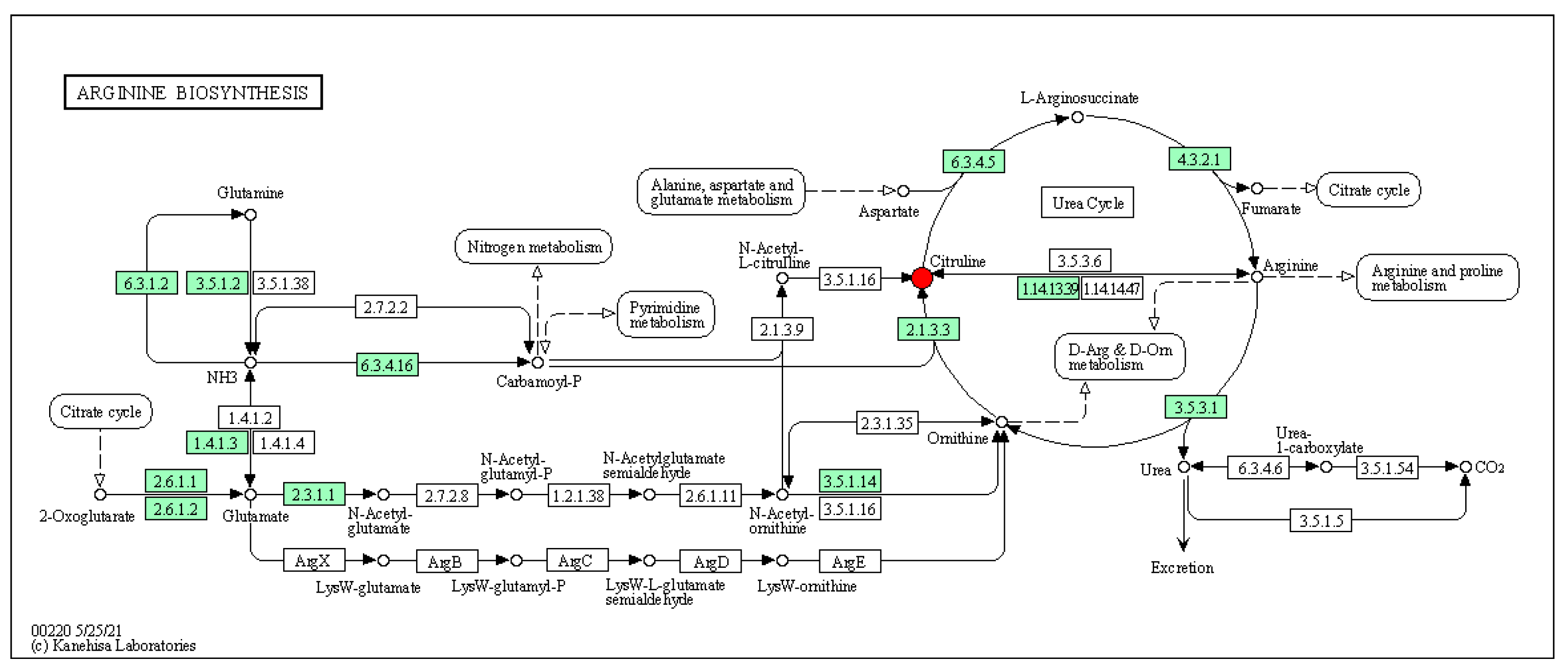
Notably, the NO in the inflamed bowel is generated mostly by an inducible nitric oxide synthase (NOS2) from L-arginine (further referred to as arginine). L-citrulline, an intermediate metabolic amino acid produced by small intestinal cells, was modulated by LP+
Ly. L-citrulline is involved in the urea cycle, arginine synthesis, and nitric oxide production. Previous research has shown that certain amino acids, such as glutamine and arginine, can influence the pathogenesis of IBD [
34]. In DSS-treated mice, L-arginine supplementation reduced colonic inflammation [
35,
36,
37]. Additionally, L-arginine treatment of DSS-treated mice resulted in increased ex vivo migration of colonic epithelial cells, suggesting an increased capacity for wound repair [
38]. Moreover, plasma guanidinoacetate levels are considered promising quantitative biomarker for diagnosing gastrointestinal complications [
39]. Our study observed that plasma guanidinoacetate levels in UC mice were significantly higher compared to the Con group. Changes in guanidinoacetate levels are associated with arginine biosynthesis in the body. LP+
Ly intervention resulted in changes in L-citrulline levels, which in turn regulated arginine biosynthesis, contributing to the alleviating effects of LP+
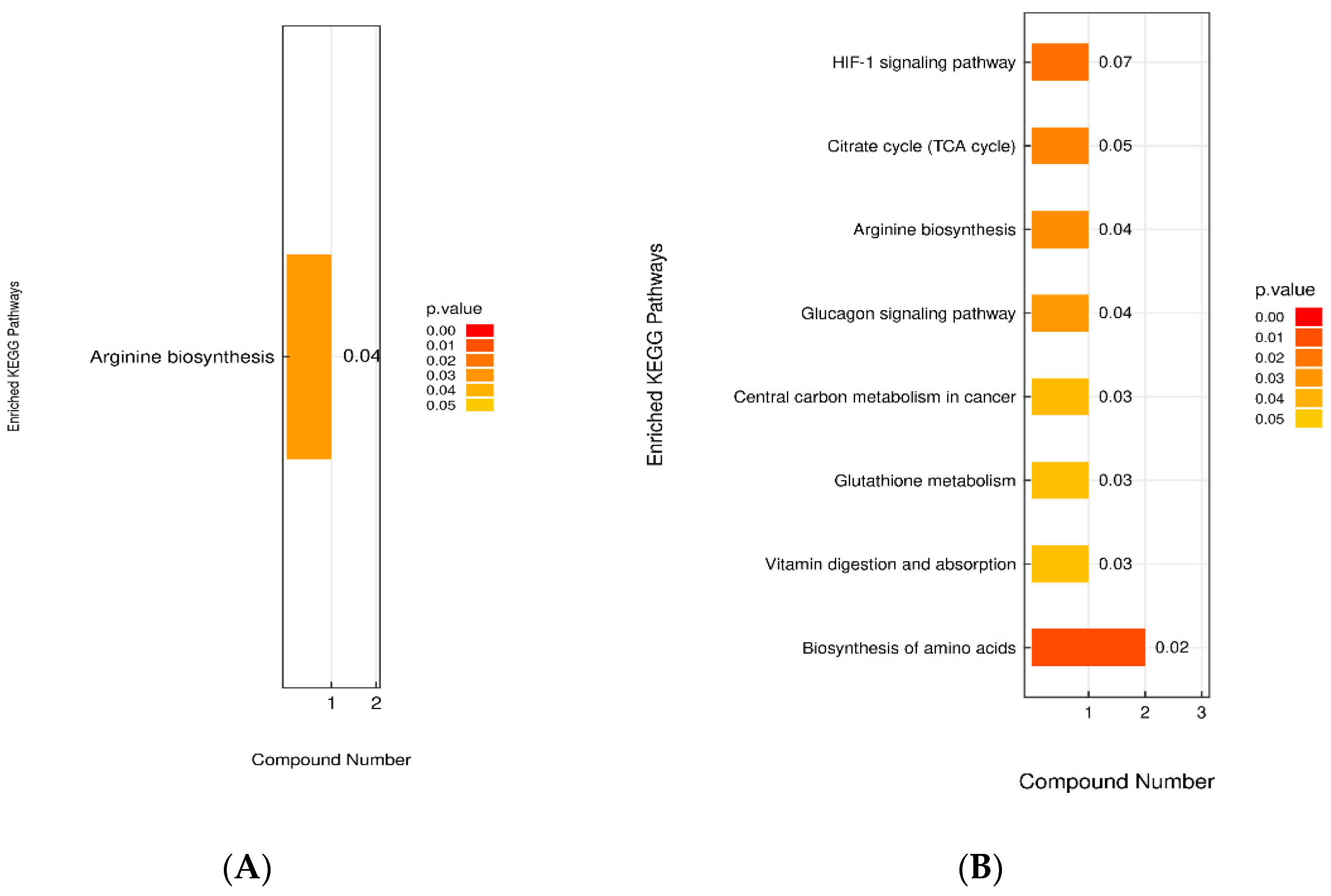
Ly. KEGG enrichment analysis suggested that the arginine biosynthesis pathway, in which L-citrulline participates, may be a potential therapeutic target for the action of LP+
Ly.
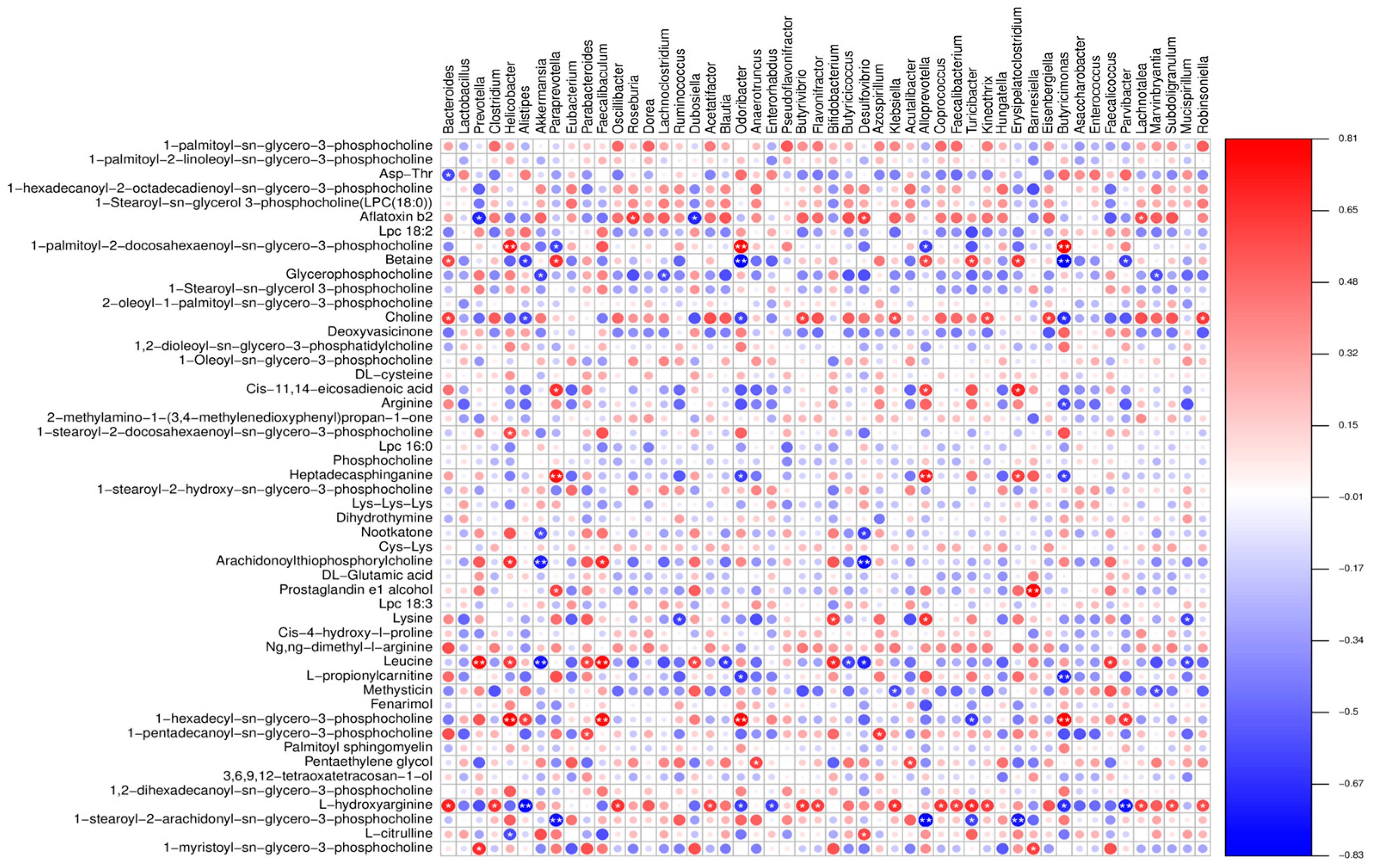
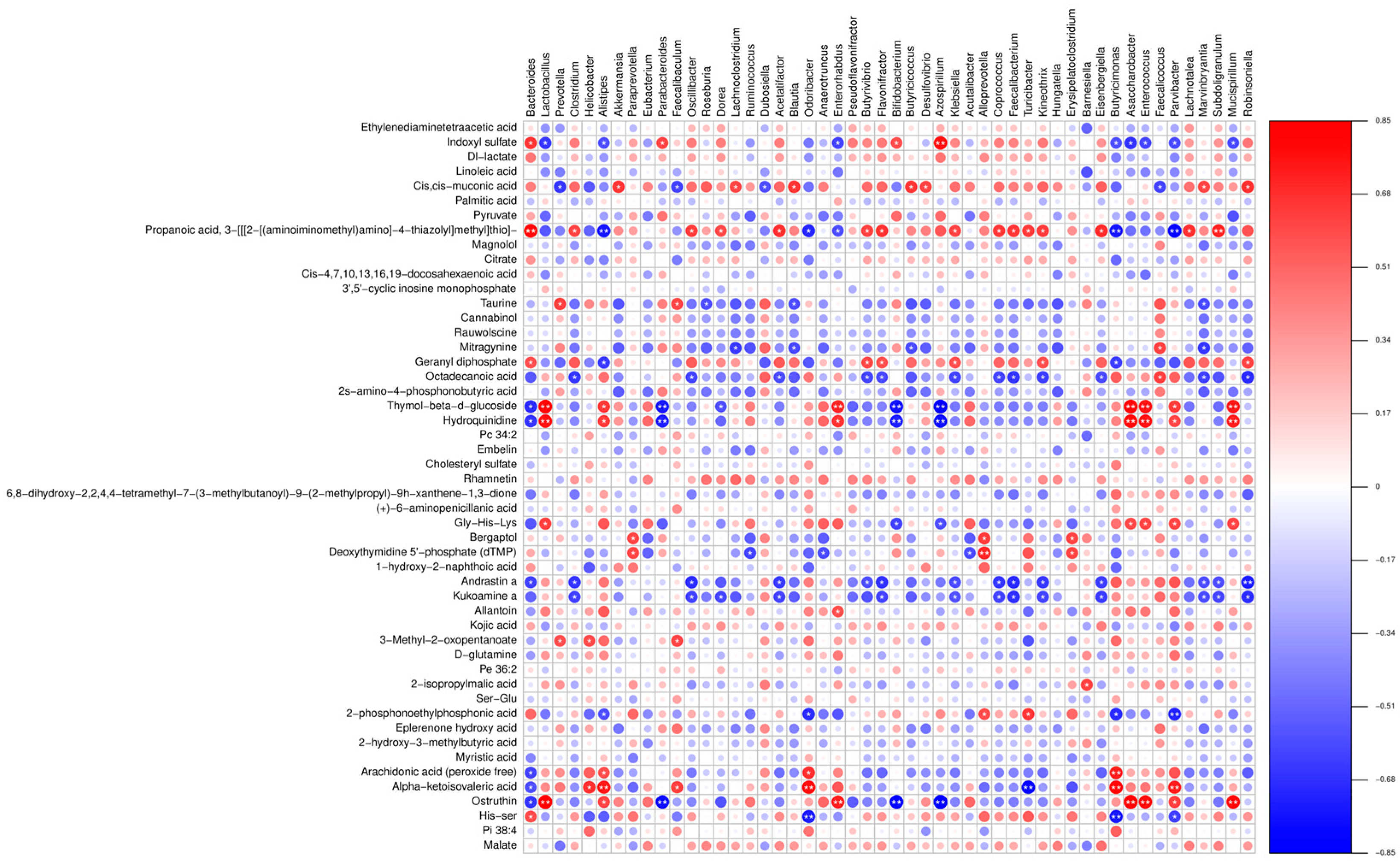
Furthermore, our correlation analysis showed a strong association between numerous gut microbiota and differential metabolites. Previous studies have shown that supplementation with arginine can restore gut microbiota diversity [
36]. Therefore, we postulate that LP+
Ly exerts its beneficial effects by modulating the gut microbiota and influencing amino acid metabolism.
In summary, LP+Ly exhibited promising anti-inflammatory effects in DSS-induced UC in mice. This study provides scientific evidence supporting the potential clinical application of LP+Ly in the prevention and treatment of UC and may also contribute to the development of novel therapeutic approaches for UC.




{kind=link}
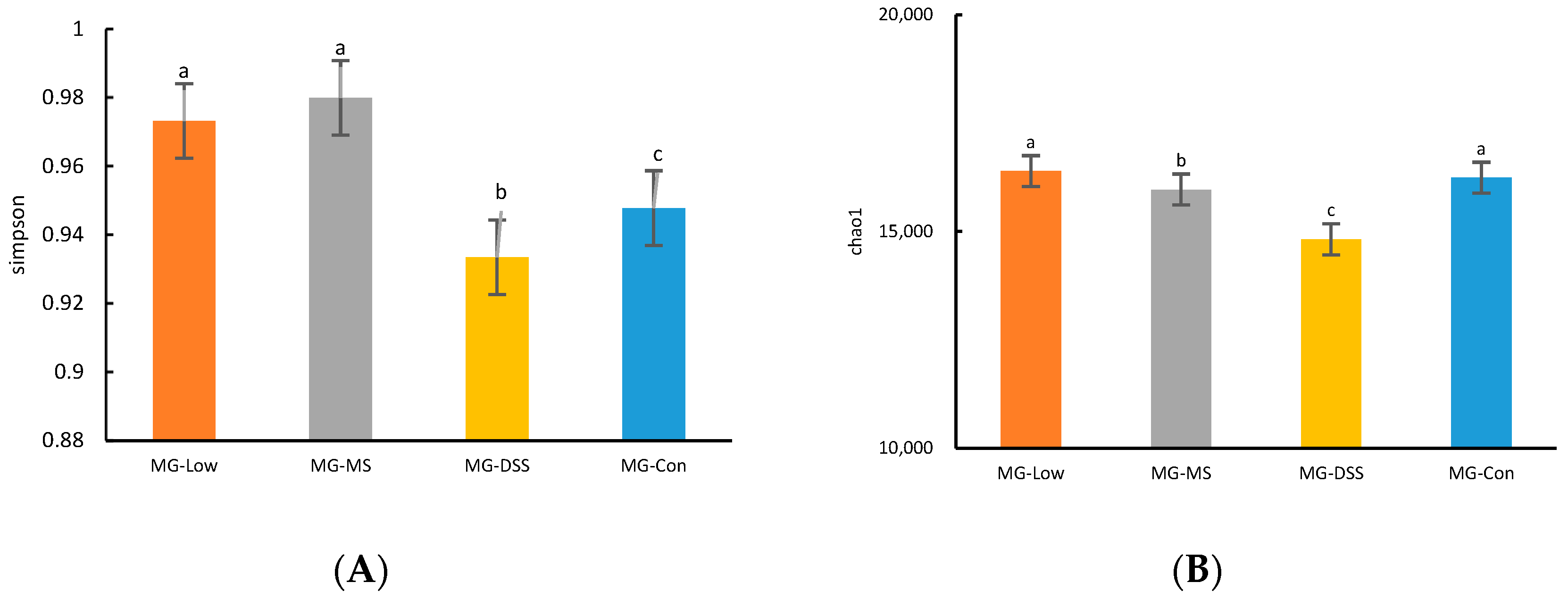{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




















