
Effect of Aspergillus niger Fermentation on the Metabolites in Corn Stalks
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Methods
2.2.1. Solid-State Fermentation
2.2.2. Biochemical Analysis
2.2.3. Sample Preparation for GC–MS
2.2.4. GC–MS Analysis
2.2.5. Data Processing, Statistical Analysis, and Identification of Metabolites
3. Results and Discussion
3.1. Biochemical Changes in Fermented Corn Stalks
3.2. Multivariate Statisitical Analysis of Detected Metabolites
3.3. Differential Metabolite Analysis Results and Visualization
3.3.1. Sugar and Glycoside Derivatives
3.3.2. Organic Acids and Their Derivatives
3.3.3. Alcohol Compounds
3.3.4. Benzene and Its Substituted Derivatives
3.3.5. Amino Acids
3.3.6. Phenolic Compounds
3.3.7. Flavonoids
3.3.8. Other Compounds
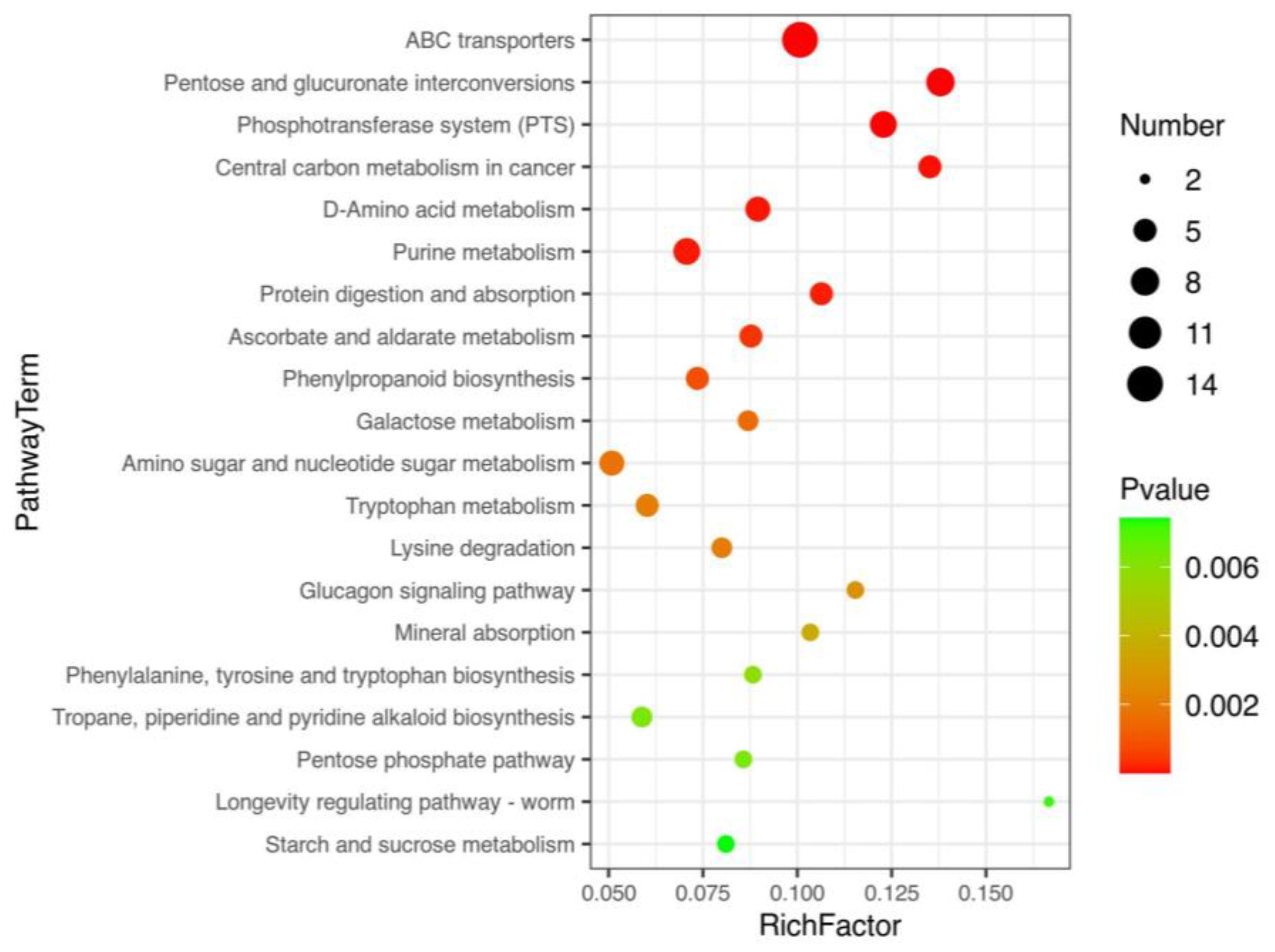
3.4. Metabolic Pahtway Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gado, H.M.; Elghandour, M.M.Y.; Cipriano, M.; Odongo, N.E.; Salem, A.Z.M. Rumen degradation and nutritive utilization of wheat straw, corn stalks and sugarcane bagasse ensiled with multienzymes. J. Appl. Anim. Res. 2017, 45, 485–489. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xia, K.; Wang, X.N.; Lin, X.; Liu, J.; Li, Y.J.; Liu, X.L.; Zhao, W.J.; Zhang, Y.G.; Guo, J.H. Improvement of feed intake, digestibility, plasma metabolites, and lactation performance of dairy cows fed mixed silage of sugar beet pulp and rice straw inoculated with lactic acid bacteria. J. Dairy Sci. 2022, 105, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Xiang, H.; Zhao, X.L.; Fang, Y.; Wang, F.; Liang, R.; Sun, X.Z.; Wang, S.P.; Zhong, R.Z. Feeding fungal-pretreated corn straw improves health and meat quality of lambs infected with gastrointestinal nematodes. Animals 2020, 10, 1659. [Google Scholar] [CrossRef]
- Zhao, X.L.; Wang, F.; Fang, Y.; Zhou, D.W.; Wang, S.P.; Wu, D.Q.; Wang, L.X.; Zhong, R.Z. High-potency white-rot fungal strains and duration of fermentation to optimize corn straw as ruminant feed. Bioresour. Technol. 2020, 312, 123512. [Google Scholar] [CrossRef]
- Liu, Y.; Yue, Z.; Jinglei, L.; Shan, L.; Shudong, H.; Hanju, S.; Shengfei, Y.; Shangying, X. Effect of enzymes addition on the fermentation of Chinese rice wine using defined fungal starter. LWT-Food Sci. Technol. 2021, 143, 111101. [Google Scholar]
- Jeong, E.; Seo, J.-A. Enzyme activity of Aspergillus section Nigri strains isolated from the Korean fermentation starter, nuruk. J. Microbiol. 2022, 60, 998–1006. [Google Scholar] [CrossRef] [PubMed]
- Di Francia, A.; Masucci, F.; De Rosa, G.; Vafficchio, M.L.; Proto, V. Effects of Aspergillus oryzae extract and a Saccharomyces cerevisiae fermentation product on intake, body weight gain and digestibility in buffalo calves. Anim. Feed. Sci. Technol. 2008, 140, 67–77. [Google Scholar] [CrossRef]
- Nedelkov, K.; Raisanen, S.E.; Chen, X.; Harper, M.T.; Melgar, A.; Oh, J.; Compart, D.M.P.; Hristov, A.N. Effect of enzyme extracts from Aspergillus oryzae and Aspergillus niger on rumen bacterial and fungal diversity and fermentation in vitro. J. Dairy Sci. 2019, 102, 378–379. [Google Scholar]
- Zhang, J.Y.; Jin, W.; Jiang, Y.; Xie, F.; Mao, S.Y. Response of milk performance, rumen and hindgut microbiome to dietary supplementation with Aspergillus oryzae fermentation extracts in dairy cows. Curr. Microbiol. 2022, 79, 113. [Google Scholar] [CrossRef]
- Yin, Z.N.; Wu, W.J.; Sun, C.Z.; Lei, Z.G.; Chen, H.M.; Liu, H.F.; Chen, W.B.; Ma, J.J.; Min, T.; Zhang, M.M.; et al. Comparison of releasing bound phenolic acids from wheat bran by fermentation of three Aspergillus species. Int. J. Food Sci. Technol. 2018, 53, 1120–1130. [Google Scholar] [CrossRef]
- Yang, Y.A.; Wang, L.L.; Zhang, Y.F.; Li, L.B.; Shi, X.Y.; Liu, X.T.; Ren, X.D.; Dou, S. Transformation of corn stalk residue to humus-like substances during solid-state fermentation. Sustainability 2019, 11, 6771. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Xie, Y.; Li, X.M.; Wu, D.H.; Cai, G.L.; Lu, J. Key compounds and metabolic pathway responsible for the browning in Dangshan pear (Pyrus spp.) wine. J. Agric. Food Chem. 2021, 69, 10311–10320. [Google Scholar] [CrossRef]
- Wang, Y.B.; Lin, B.J.; Li, Z.X. Effect of Lactobacillus plantarum fermentation on metabolites in lotus leaf based on ultra-high-performance liquid chromatography-high-resolution mass spectrometry. Fermentation 2022, 8, 599. [Google Scholar] [CrossRef]
- Waksman, S.A.; Stevens, K.R. A system of proximate chemical analysis of plant materials. Ind. Eng. Chem. Anal. Ed. 1930, 2, 167–173. [Google Scholar] [CrossRef]
- Daly, P.; van Munster, J.M.; Blythe, M.J.; Ibbett, R.; Kokolski, M.; Gaddipati, S.; Lindquist, E.; Singan, V.R.; Barry, K.W.; Lipzen, A.; et al. Expression of Aspergillus niger CAZymes is determined by compositional changes in wheat straw generated by hydrothermal or ionic liquid pretreatments. Biotechnol. Biofuels Bioprod. 2017, 10, 35. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.E.; Lee, S.; Jang, E.S.; Shin, H.W.; Moon, B.S.; Lee, C.H. Metabolomic profiles of Aspergillus oryzae and Bacillus amyloliquefaciens during rice koji fermentation. Molecules 2016, 21, 773. [Google Scholar] [CrossRef] [Green Version]
- Berks, A.H. The chapman and hall dictionary of drugs on CD-ROM. J. Chem. Inf. Comput. Sci. 1995, 35, 332–333. [Google Scholar] [CrossRef]
- Kopfmaier, P.; Sass, G. Antitumor-activity of treosulfan against human breast carcinomas. Cancer Chemother. Pharmacol. 1992, 31, 103–110. [Google Scholar] [CrossRef]
- Sommerdijk, N.; Feiters, M.C.; Nolte, R.J.M.; Zwanenburg, B. Aggregation behavior of a phospholipid based on D-(-)-threitol. J. R. Neth. Chem. Soc. 1994, 113, 194–200. [Google Scholar]
- Shen, Y.; Gu, L.; Zhang, J.; Chen, J.; Du, G. Effects of mixed carbon sources on glucose oxidase production by recombinant Pichia pastoris. Chin. J. Biotechnol. 2013, 29, 927–936. [Google Scholar]
- Hofmann, T.; Talos-Nebehaj, E.; Albert, L.; Nemeth, L. Antioxidant efficiency of Beech (Fagus sylvatica L.) bark polyphenols assessed by chemometric methods. Ind. Crops Prod. 2017, 108, 26–35. [Google Scholar] [CrossRef]
- Janek, T.; Dobrowolski, A.; Biegalska, A.; Mironczuk, A.M. Characterization of erythrose reductase from Yarrowia lipolytica and its influence on erythritol synthesis. Microb. Cell Factories 2017, 16, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flint, N.; Hamburg, N.M.; Holbrook, M.; Dorsey, P.G.; LeLeiko, R.M.; Berger, A.; de Cock, P.; Bosscher, D.; Vita, J.A. Effects of erythritol on endothelial function in patients with type 2 diabetes mellitus: A pilot study. Acta Diabetol. 2014, 51, 513–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahalak, K.K.; Firrman, J.; Tomasula, P.M.; Nunez, A.; Lee, J.J.; Bittinger, K.; Rinaldi, W.; Liu, L.S. Impact of steviol glycosides and erythritol on the human and cebus apella gut microbiome. J. Agric. Food Chem. 2020, 68, 13093–13101. [Google Scholar] [CrossRef] [PubMed]
- Bapu, D.; Runions, J.; Kadhim, M.; Brooks, S.A. N-acetylgalactosamine glycans function in cancer cell adhesion to endothelial cells: A role for truncated O-glycans in metastatic mechanisms. Cancer Lett. 2016, 375, 367–374. [Google Scholar] [CrossRef] [Green Version]
- Pearlin, B.V.; Muthuvel, S.; Govidasamy, P.; Villavan, M.; Alagawany, M.; Ragab Farag, M.; Dhama, K.; Gopi, M. Role of acidifiers in livestock nutrition and health: A review. J. Anim. Physiol. Anim. Nutr. 2020, 104, 558–569. [Google Scholar] [CrossRef] [Green Version]
- Khan, R.U.; Naz, S.; Raziq, F.; Qudratullah, Q.; Khan, N.A.; Laudadio, V.; Tufarelli, V.; Ragni, M. Prospects of organic acids as safe alternative to antibiotics in broiler chickens diet. Environ. Sci. Pollut. Res. 2022, 29, 32594–32604. [Google Scholar] [CrossRef]
- Tomaszewska, E.; Dobrowolski, P.; Puzio, I. Postnatal administration of 2-oxoglutaric acid improves the intestinal barrier affected by the prenatal action of dexamethasone in pigs. Nutrition 2012, 28, 190–196. [Google Scholar] [CrossRef]
- Sun, Y.Y.; Wang, C.H. Inhibitory effect of growth-inhibitor on Isochrysis galbana and inhibition of antioxidants on growth-inhibitor damage to its cells. Prog. Biochem. Biophys. 2007, 34, 1162–1168. [Google Scholar]
- Hu, X.; Wang, J. Preparation and application of chenodeoxycholic acid and its derivatives. Prog. Chem. 2016, 28, 814–828. [Google Scholar]
- Su, J.H.; Wang, H.X.; Ma, C.Y.; Liu, C.X.; Rahman, M.T.; Gao, C.Z.; Nie, R.J. Hypolipidemic mechanism of gypenosides via inhibition of pancreatic lipase and reduction in cholesterol micellar solubility. Eur. Food Res. Technol. 2016, 242, 305–312. [Google Scholar] [CrossRef]
- Broeders, E.P.M.; Nascimento, E.B.M.; Havekes, B.; Brans, B.; Roumans, K.H.M.; Tailleux, A.; Schaart, G.; Kouach, M.; Charton, J.; Deprez, B.; et al. The bile acid chenodeoxycholic acid increases human brown adipose tissue activity. Cell Metab. 2015, 22, 418–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onal, G.; Kutlu, O.; Gozuacik, D.; Emre, S.D. Lipid droplets in health and disease. Lipids Health Dis. 2017, 16, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, H.; Yang, C.; Yu, Y.; Yang, W.; Lu, N.; Wang, H.; Liu, F.; Wang, A.; Xu, X. Dietary cholesterol level affects growth, molting performance and ecdysteroid signal transduction in Procambarus clarkii. Aquaculture 2020, 523, 735198. [Google Scholar] [CrossRef]
- Xingyong, L.; Xinglian, C.; Lijuan, D.; Tao, L.; Benlin, Y.; Dongshun, Y.; Jinliang, S.; Luxiang, W. The interaction of various factors leads to rapid degradation of chlorogenic acid in roasted coffee beans during processing. Food Sci. China 2021, 42, 7–14. [Google Scholar]
- Josa, M.; Urizar, J.P.; Rapado, J.; Dios-Vieitez, C.; Castaneda-Hernandez, G.; Flores-Murrieta, F.; Renedo, M.J.; Troconiz, I.F. Pharmacokinetic/pharmacodynamic modeling of antipyretic and anti-inflammatory effects of naproxen in the rat. J. Pharmacol. Exp. Ther. 2001, 297, 198–205. [Google Scholar]
- De, A.; Singh, M.F.; Singh, V.; Ram, V.; Bisht, S. Treatment effect of L-Norvaline on the sexual performance of male rats with streptozotocin induced diabetes. Eur. J. Pharmacol. 2016, 771, 247–254. [Google Scholar] [CrossRef]
- Priefert, H.; Rabenhorst, J.; Steinbuchel, A. Biotechnological production of vanillin. Appl. Microbiol. Biotechnol. 2001, 56, 296–314. [Google Scholar] [CrossRef]
- Nauman, M.; Kale, R.K.; Singh, R.P. Polyphenols of Salix aegyptiaca modulate the activities of drug metabolizing and antioxidant enzymes, and level of lipid peroxidation. BMC Complement. Altern. Med. 2018, 18, 81. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.; Sun, J.; Sun, B.; Zhao, M.; Zheng, F.; Huang, M.; Sun, X.; Li, H. Intracellular antioxidant effect of vanillin, 4-methylguaiacol and 4-ethylguaiacol: Three components in Chinese Baijiu. RSC Adv. 2017, 7, 46395–46405. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez-Merino, C.; Lopez-Sanchez, C.; Lagoa, R.; Samhan-Arias, A.K.; Bueno, C.; Garcia-Martinez, V. Neuroprotective actions of flavonoids. Curr. Med. Chem. 2011, 18, 1195–1212. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.J.; Xiao, H.; Zeng, Z.; Sun, Z.W.; Lei, C.; Dong, J.Z.; Wang, Y. Composition and serum antioxidation of the main flavonoids from fermented vine tea (Ampelopsis grossedentata). J. Funct. Foods 2014, 9, 290–294. [Google Scholar] [CrossRef]
- Gandhi, G.R.; Neta, M.; Sathiyabama, R.G.; Quintans, J.D.S.; Silva, A.; Araujo, A.A.D.; Narain, N.; Quintans, L.J.; Gurgel, R.Q. Flavonoids as Th1/Th2 cytokines immunomodulators: A systematic review of studies on animal models. Phytomedicine 2018, 44, 74–84. [Google Scholar] [CrossRef]
- North, M.K.; Zotte, A.D.; Hoffman, L.C. The use of dietary flavonoids in meat production: A review. Anim. Feed. Sci. Technol. 2019, 257, 114291. [Google Scholar] [CrossRef]
- Hu, R.Z.; He, Z.Y.; Liu, M.; Tan, J.J.; Zhang, H.F.; Hou, D.X.; He, J.H.; Wu, S.S. Dietary protocatechuic acid ameliorates inflammation and up-regulates intestinal tight junction proteins by modulating gut microbiota in LPS-challenged piglets. J. Anim. Sci. Biotechnol. 2021, 12, 328–339. [Google Scholar] [CrossRef]
- Kakkar, S.; Bais, S. A review on protocatechuic acid and its pharmacological potential. ISRN Pharmacol. 2014, 2014, 952943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaewmool, C.; Kongtawelert, P.; Phitak, T.; Pothacharoen, P.; Udomruk, S. Protocatechuic acid inhibits inflammatory responses in LPS-activated BV2 microglia via regulating SIRT1/NF-kappa B pathway contributed to the suppression of microglial activation-induced PC12 cell apoptosis. J. Neuroimmunol. 2020, 341, 577164. [Google Scholar] [CrossRef]
- Abdelmageed, M.E.; Nader, M.A.; Zaghloul, M.S. Targeting HMGB1/TLR4/NF-kappa B signaling pathway by protocatechuic acid protects against 1-arginine induced acute pancreatitis and multiple organs injury in rats. Eur. J. Pharmacol. 2021, 906, 174279. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Lim, H.S.; Chung, S.K. A short and efficient stereoselective synthesis of all four diastereomers of sphingosine. Tetrahedron Asymmetry 2002, 13, 343–347. [Google Scholar] [CrossRef]
- Chen, J.F.; Eltzschig, H.K.; Fredholm, B.B. Adenosine receptors as drug targets—what are the challenges? Nat. Rev. Drug Discov. 2013, 12, 265–286. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fractions | Control | Fermented |
|---|---|---|
| Neutral detergent fiber | 66.98 ± 0.08 | 58.46 ± 0.28 |
| Acid detergent fiber | 38.12 ± 0.48 | 36.96 ± 0.64 |
| Lignin | 5.24 ± 0.26 | 5.14 ± 0.32 |
| Cellulose | 33.92 ± 0.42 | 31.36 ± 0.62 |
| Hemicellulose | 23.86 ± 0.54 | 20.06 ± 0.22 |
| Crude protein | 8.20 ± 0.16 | 11.24 ± 0.18 |
| Species | Metabolite Name | VIP | log2(FC) | Increase/Decrease |
|---|---|---|---|---|
| Sugar and glycoside derivatives | D-Threitol | 2.45 | 11.53 | Increase |
| D-Ribulose-5-phosphate | 2.13 | 8.62 | Increase | |
| Mannitol-1-phosphate | 1.73 | 5.74 | Increase | |
| Coniferin | 1.64 | 5.08 | Increase | |
| Erythritol | 1.61 | 4.99 | Increase | |
| N-Acetyl-D-mannosamine | 1.52 | 4.41 | Increase | |
| N-Acetylgalactosamine | 1.52 | −4.41 | Decrease | |
| Melibiose | 1.70 | −5.59 | Decrease | |
| Melezitose | 1.96 | −7.34 | Decrease | |
| Sucrose | 2.06 | −8.81 | Decrease | |
| Organic acids and derivatives | Citrazinic | 1.92 | 7.06 | Increase |
| 3-Hydroxypalmitic acid | 1.77 | 5.97 | Increase | |
| Oxoglutaric acid | 1.63 | 5.06 | Increase | |
| Chenodeoxycholic acid | 1.60 | 4.87 | Increase | |
| Cholic acid | 1.52 | 4.39 | Increase | |
| Xanthurenic acid | 1.72 | 1.87 | Increase | |
| Azelaic acid | 1.61 | −4.96 | Decrease | |
| Heneicosanoic acid | 2.48 | −11.8 | Decrease | |
| Alcohol compounds | D-myo-Inositol-4-phosphate | 1.57 | 4.72 | Increase |
| 1-Heptanol | 1.54 | −4.54 | Decrease | |
| Cholesterol | 1.65 | −5.26 | Decrease | |
| Isochlorogenic acid | 1.82 | −6.38 | Decrease | |
| Chlorogenic acid | 2.21 | −9.39 | Decrease | |
| Benzene and substituted derivatives | (r)-Amphetamine | 1.55 | 4.62 | Increase |
| Oxymetazoline Naproxen | 1.60 1.83 | −4.93 6.41 | Decrease Increase | |
| Amino acids | 5-Aminovaleric acid | 1.57 | 4.74 | Increase |
| Phenolic compounds | Vanillin | 1.81 | 6.24 | Increase |
| Flavonoids | Catechin | 1.87 | 6.63 | Increase |
| Other compounds | UDP-Glucuronic acid | 2.36 | 10.66 | Increase |
| D-erythro-Sphingosine | 1.85 | 7.77 | Increase | |
| Adenosine | 1.81 | −6.27 | Decrease |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, Z.; Chen, T.; Cai, G.; Huang, X.; Zhong, S.; Li, X.; Zhang, E. Effect of Aspergillus niger Fermentation on the Metabolites in Corn Stalks. Fermentation 2023, 9, 50. https://doi.org/10.3390/fermentation9010050
Fan Z, Chen T, Cai G, Huang X, Zhong S, Li X, Zhang E. Effect of Aspergillus niger Fermentation on the Metabolites in Corn Stalks. Fermentation. 2023; 9(1):50. https://doi.org/10.3390/fermentation9010050
Chicago/Turabian StyleFan, Zhen, Tianming Chen, Guolin Cai, Xiaoyu Huang, Suchuan Zhong, Xiaoming Li, and Enping Zhang. 2023. "Effect of Aspergillus niger Fermentation on the Metabolites in Corn Stalks" Fermentation 9, no. 1: 50. https://doi.org/10.3390/fermentation9010050