
The Effect of Forage Source and Concentrated Liquid Feedstuff Supplementation on Improving the Synchronization of Ruminant Dietary Energy and Nitrogen Release In Vitro

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical Composition of M-CMS and Calculation of Synchrony Index
2.2. Preparation of Experimental Diets
2.2.1. Experiment 1: Effect of M-CMS Supplementation on Diets with or without Corn Silage
2.2.2. Experiment 2: Effect of M-CMS Supplementation on Corn Silage Level under High Synchrony Index
2.3. In Vitro Rumen Digestibility and Gas Production Assay
2.4. Sample Collection and Chemical Analysis
2.5. Statistical Analysis
3. Results
3.1. Effect of M-CMS Supplementation on Diets with or without Corn Silage
3.2. The M-CMS Supplementation Effect on Difference Corn Silage Levels under High Synchrony Index
4. Discussions
4.1. The Interaction of Forage Source and M-CMS Supplementation
4.2. The M-CMS Supplementation Effect on Different Corn Silage Levels under High Synchrony Index
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaswari, T.; Lebzien, P.; Flachowsky, G.; ter Meulen, U. Studies on the relationship between the synchronization index and the microbial protein synthesis in the rumen of dairy cows. Anim. Feed Sci. Technol. 2007, 139, 1–22. [Google Scholar] [CrossRef]
- Herrera-Saldana, R.; Gomez-Alarcon, R.; Torabi, M.; Huber, J.T. Influence of synchro- nizing protein and starch degradation in the rumen on nutrient utilization and microbial protein synthesis. J. Dairy Sci. 1990, 73, 142–148. [Google Scholar] [CrossRef]
- Henning, P.H.; Steyn, D.G.; Meissner, H.H. The effect of energy and nitrogen supply pattern on rumen bacterial growth in vitro. Anim. Sci. 1991, 53, 165–175. [Google Scholar] [CrossRef]
- Hall, M.B.; Huntington, G.B. Nutrient synchrony: Sound in theory, elusive in practice. J. Anim. Sci. 2008, 86, E287–E292. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, L.A.; Garnsworthy, P.C.; Newbold, J.R.; Buttery, P.J. Effect of synchronizing the rate of dietary energy and nitrogen release on rumen fermentation and microbial protein synthesis in the sheep. J. Agric. Sci. 1993, 120, 251–263. [Google Scholar] [CrossRef]
- Menke, H.H.; Steingass, H. Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim. Res. Dev. 1988, 28, 7–55. [Google Scholar]
- Huhtanen, P.; Seppälä, A.; Ahvenjärvi, S.; Rinne, M. Prediction of in vivo neutral detergent fiber digestibility and digestion rate of potentially digestible neutral detergent fiber: Comparison of models. J. Anim. Sci. 2008, 86, 2657–2669. [Google Scholar] [CrossRef]
- Peltekova, V.D.; Broderick, G.A. In vitro ruminal degradation and synthesis of protein on fractions extracted from alfalfa hay and silage. J. Dairy Sci. 1996, 79, 612–619. [Google Scholar] [CrossRef]
- Bowman, J.G.P.; Sowell, B.F.; Patterson, J.A. Liquid supplementation for ruminants fed low-quality forage diets: A review. Anim. Feed Sci. Tech. 1995, 55, 105–138. [Google Scholar] [CrossRef]
- Scull, I.; Savon, L.; Gutierrez, O.; Valino, E.; Orta, I.; Mora, P.O.; Orta, H.; Yusmely, R.; Molineda, A.; Coto, G.; et al. Physio-Chemical composition of concentrated vinasse for their assessment in animal diets. Cuba. J. Agric. Sci. 2012, 46, 385–389. [Google Scholar]
- Ma, J.; Ma, C.; Fan, X.; Shah, A.; Mao, J. Use of condensed molasses fermentation solubles as an alternative source of concentrates in dairy cows. Anim. Biosci. 2021, 34, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Stemme, K.; Gerdes, B.; Harms, A.; Kamphues, J. Beet-vinasse (condensed molasses solubles) as an ingredient in diets for cattle and pigs-nutritive value and limitations. J. Anim. Physiol. Anim. Nutr. 2005, 89, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Henning, P.H.; Steyn, D.G.; Meissner, H.H. Effect of synchronization of energy and nitrogen supply on ruminal characteristics and microbial growth. J. Anim. Sci. 1993, 71, 2516–2528. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists. Official Methods of Analysis, 17th ed.; AOAC: Washington, DC, USA, 2000. [Google Scholar]
- Lei, Y.G.; Li, X.Y.; Wang, Y.Y.; Li, Z.Z.; Chen, Y.L.; Yang, Y.X. Determination of ruminal dry matter and crude protein degradability and degradation kinetics of several concentrate feed ingredients in cashmere goat. J. Appl. Anim. Res. 2018, 46, 134–140. [Google Scholar] [CrossRef]
- Seo, S.; Tedeschi, L.O.; Schwab, C.G.; Garthwaite, B.D. Fox, D.G. Evaluation of the passage rate equations in the 2001 dairy NRC model. J. Dairy Sci. 2006, 89, 2327–2342. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Dairy Cattle, 7th rev. ed.; National Academy of Sciences: Washington, DC, USA, 2001. [Google Scholar]
- Ankom Technology. In Vitro True Digestibility Using the DaisyII Incubator. Available online: https://www.ankom.com/sites/default/files/document-files/Method_3_Invitro_D200_D200I.pdf (accessed on 21 December 2018).
- Spanghero, M.; Boccalon, S.; Gracco, L.; Gruber, L. NDF degradability of hays measured in situ and in vitro. Anim. Feed Sci. Technol. 2003, 104, 201–208. [Google Scholar] [CrossRef]
- Zinn, R.A.; Owens, F.N. A rapid procedure for purine measurement and its use for estimating net ruminal protein synthesis. Can. J. Anim. Sci. 1986, 66, 157–166. [Google Scholar] [CrossRef]
- Chaney, A.L.; Marbach, E.P. Modified reagents for determination of urea and ammonia. Clin. Chem. 1962, 8, 130–132. [Google Scholar] [CrossRef]
- Seo, J.K.; Kim, M.H.; Yang, J.Y.; Kim, H.J.; Lee, C.H.; Kim, K.H.; Ha, J.K. Effects of synchronicity of carbohydrate and protein degradation on rumen fermentation characteristics and microbial protein synthesis. Asian-Austral. J. Anim. Sci. 2013, 26, 358–365. [Google Scholar] [CrossRef]
- Cabrita, A.R.J.; Dewhurst, R.J.; Abreu, J.M.F.; Fonseca, A.J.M. Evaluation of the effects of synchronising the availability of N and energy on rumen function and production responses of dairy cows–a review. Anim. Res. 2006, 55, 1–24. [Google Scholar] [CrossRef]
- Kolver, E.; Muller, L.D.; Varga, G.A.; Cassidy, T.J. Synchronization of ruminal degradation of supplemental carbohydrate with pasture nitrogen in lactating dairy cows. J. Dairy. Sci. 1998, 81, 2017–2028. [Google Scholar] [CrossRef]
- Zali, A.; Eftekhari, M.; Fatehi, F.; Ganjkhanlou, M. Effect of vinasse (condensed molasses solubles) on performance and meat chemical composition of Holstein male calves. Ital. J. Anim. Sci. 2017, 16, 515–520. [Google Scholar] [CrossRef] [Green Version]
- Sannes, R.A.; Messman, M.A.; Vagnoni, D.B. Form of rumen-degradable carbohydrate and nitrogen on microbial protein synthesis and protein efficiency of dairy cows. J. Dairy Sci. 2002, 85, 900–908. [Google Scholar] [CrossRef]
- Broderick, G.A.; Radloff, W.J. Effect of molasses supplementation on the production of lactating dairy cows fed diets based on alfalfa and corn silage. J. Dairy Sci. 2004, 87, 2997–3009. [Google Scholar] [CrossRef]
- Hersom, M.J. Opportunities to enhance performance and efficiency through nutrient synchrony in forage-fed ruminants. J. Anim. Sci. 2007, 86, E306–E317. [Google Scholar] [CrossRef]
- Tedeschi, L.O.; Chalupa, W.; Janczewski, E.; Fox, D.G.; Sniffen, C.; Munson, R.; Kononoff, P.; Boston, R. Evaluation and application of the CPM dairy nutrition model. J. Agric. Sci. 2008, 146, 171–182. [Google Scholar] [CrossRef]
- Yang, J.Y.; Seo, J.; Kim, H.J.; Seo, S.; Ha, J.K. Nutrient Synchrony: Is it a suitable strategy to improve nitrogen utilization and animal performance? Asian Australas. J. Anim. Sci. 2010, 23, 972–979. [Google Scholar] [CrossRef]
- Witt, M.W.; Sinclair, L.A.; Wilkinson, R.G.; Buttery, P.J. The effects of synchronizing the rate of dietary energy and nitrogen supply to the rumen on milk production and metabolism of ewes offered grass silage based diets. Anim. Sci. 2000, 71, 187–195. [Google Scholar] [CrossRef]
- El Khidir, O.A.; Thomsen, K.V. The effect of high levels of molasses in combinations with hay on digestibility of organic matter, microbial protein synthesis and volatile fatty acid production in vitro. Anim. Feed Sci. Technol. 1982, 7, 277–286. [Google Scholar] [CrossRef]
- Trevaskis, L.M.; Fulkerson, W.J.; Gooden, J. Provision of certain carbohydrate-based supplements to pasture-fed sheep, as well as time of harvesting of the pasture, influences pH, ammonia concentration and microbial protein synthesis in the rumen. Aust. J. Exp. Agric. 2001, 41, 21–27. [Google Scholar] [CrossRef]
- Valkeners, D.; Tahewis, A.; Piron, F.; Beekers, Y. Effect of imbalance between energy and nitrogen supplies on microbial protein synthesis and nitrogen metabolism in growing double muscled Belgian Blue bulls. J. Anim Sci. 2004, 82, 1818–1825. [Google Scholar] [CrossRef] [PubMed]
- Buchanan-Smith, J.G.; Yao, Y.T. Non-protein nitrogen in corn silage: A partial characterization, its utilization in the rumen and effect upon digestibility and retention of nitrogen in lambs. Can. J. Anim. Sci. 1978, 58, 681–690. [Google Scholar] [CrossRef]
- Wischer, G.; Boguhn, J.; Steingass, H.; Schollenberger, M.; Hartung, K.; Rodehutscord, M. Effect of monensin on in vitro fermentation of silages and microbial protein synthesis. Arch. Anim. Nutr. 2013, 67, 219–234. [Google Scholar] [CrossRef] [PubMed]
- Rotger, A.; Ferret, A.; Calsamiglia, S.; Manteca, X. Effects of nonstructural carbohydrates and protein sources on intake, apparent total tract digestibility, and ruminal metabolism in vivo and in vitro with high-concentrate beef cattle diets. J. Anim. Sci. 2006, 84, 1188–1196. [Google Scholar] [CrossRef]
- Whitelaw, F.G.; Eadie, J.M.; Bruce, L.A. Shand WJ. Methane formation in faunated and ciliate-free cattle and its relationship with rumen volatile fatty acid proportions. Br. J. Nutr. 1984, 52, 261–275. [Google Scholar] [CrossRef]
- Münnich, M.; Klevenhusen, F.; Zebeli, Q. Feeding of molassed sugar beet pulp instead of maize enhances net food production of high-producing Simmental cows without impairing metabolic health. Anim. Feed Sci. Technol. 2018, 241, 75–83. [Google Scholar] [CrossRef]
- Weidmeier, R.D.; Tanner, B.H.; Bair, J.R.; Shenton, H.T.; Arambel, M.J.; Walters, J.L. Effects of a new molasses by-product, concentrated separator by-product, on nutrient digestibility and ruminal fermentation in cattle. J. Anim. Sci. 1992, 70, 1930–1940. [Google Scholar] [CrossRef]
- Van Amburgh, M.E.; Collao-Saenz, E.A.; Higgs, R.J.; Ross, D.A.; Recktenwald, E.B.; Raffrenato, E.; Chase, L.E.; Overton, T.R.; Mills, J.K.; Foskolos, A. The cornell net carbohydrate and protein system: Updates to the model and evaluation of version 6.5. J. Dairy Sci. 2015, 98, 6361–6380. [Google Scholar] [CrossRef]
- Morales, J.L.; Van Horn, H.H.; Moore, J.E. Dietary interaction of cane molasses with source of roughage: Intake and lactation effects. J. Dairy Sci. 1989, 72, 2331–2338. [Google Scholar] [CrossRef]
- Bach, A.; Calsamiglia, S.; Stern, M.D. Nitrogen metabolism in the rumen. J. Dairy Sci. 2005, 88, E9–E21. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Treatment 1 | ||||||
|---|---|---|---|---|---|---|
| T | S | |||||
| Item | N | L | H | N | L | H |
| Ingredient (% Dry matter, DM) | ||||||
| Oat hay | 27.07 | 26.80 | 26.16 | 12.53 | 12.53 | 12.53 |
| Pangolagrass hay | 0.00 | 0.00 | 0.00 | 3.08 | 3.08 | 3.08 |
| Alfalfa hay | 24.11 | 24.12 | 24.34 | 19.28 | 19.27 | 19.27 |
| Corn silage | 0.00 | 0.00 | 0.00 | 20.03 | 20.03 | 20.03 |
| Steam-flaked corn | 25.99 | 24.80 | 23.87 | 19.85 | 18.92 | 17.26 |
| Soybean meal | 14.51 | 14.08 | 13.62 | 16.77 | 15.76 | 15.58 |
| Soybean hull | 3.86 | 3.97 | 4.06 | 5.78 | 5.98 | 6.08 |
| Bypass lipid | 2.15 | 2.21 | 2.25 | 1.09 | 1.08 | 1.08 |
| Sodium bicarbonate | 0.80 | 0.80 | 0.80 | 0.86 | 0.86 | 0.86 |
| Calcium carbonate | 0.18 | 0.10 | 0.00 | 0.35 | 0.35 | 0.35 |
| Dicalcium phosphate | 0.43 | 0.44 | 0.45 | 0.18 | 0.18 | 0.18 |
| Bypass methionine | 0.17 | 0.18 | 0.18 | 0.00 | 0.00 | 0.00 |
| Magnesium oxide | 0.17 | 0.18 | 0.18 | 0.00 | 0.00 | 0.00 |
| Vitamin premix | 0.21 | 0.22 | 0.23 | 0.20 | 0.20 | 0.20 |
| Salt | 0.34 | 0.35 | 0.36 | 0.00 | 0.00 | 0.00 |
| M-CMS | 0.00 | 1.75 | 3.50 | 0.00 | 1.75 | 3.50 |
| Calculated parameter 2 | ||||||
| Forage % | 51.18 | 50.92 | 50.50 | 54.92 | 54.92 | 54.92 |
| SI | 0.81 | 0.81 | 0.81 | 0.79 | 0.80 | 0.80 |
| RDP (%CP) | 60.40 | 61.30 | 62.14 | 60.25 | 61.00 | 62.01 |
| RUP (%CP) | 39.60 | 38.70 | 37.86 | 39.75 | 39.00 | 37.99 |
| Chemical composition 3 (% DM) | ||||||
| OM | 93.44 | 93.15 | 92.88 | 93.43 | 93.10 | 92.74 |
| CP | 15.21 | 15.18 | 15.18 | 15.79 | 15.54 | 15.60 |
| EE | 4.32 | 4.34 | 4.35 | 3.52 | 3.48 | 3.43 |
| NDF | 35.49 | 35.21 | 34.83 | 38.30 | 38.18 | 38.01 |
| ADF | 20.99 | 20.91 | 20.78 | 22.74 | 22.73 | 22.73 |
| Ash | 6.56 | 6.74 | 6.89 | 6.57 | 6.79 | 7.04 |
| NSC | 38.42 | 38.42 | 38.53 | 35.81 | 35.90 | 35.70 |
| NEL(Mcal/kg) | 1.70 | 1.69 | 1.68 | 1.68 | 1.67 | 1.67 |
| Treatment 1 | ||||
|---|---|---|---|---|
| Item | HSN | HSV | LSN | LSV |
| Ingredient (% Dry matter, DM) | ||||
| Corn silage | 29.77 | 29.77 | 14.42 | 14.42 |
| Oat hay | 6.37 | 6.37 | 20.93 | 20.00 |
| Alfalfa hay | 14.33 | 14.33 | 14.88 | 15.81 |
| Corn | 13.74 | 17.74 | 14.02 | 19.14 |
| Soybean meal | 3.02 | 3.49 | 3.72 | 2.93 |
| Soybean hull | 17.24 | 8.64 | 16.31 | 8.03 |
| DDGS | 14.83 | 14.92 | 14.92 | 14.92 |
| Premix | 0.33 | 0.33 | 0.33 | 0.33 |
| Bypass lipid | 0.37 | 0.00 | 0.47 | 0.00 |
| M-CMS | 0.00 | 4.42 | 0.00 | 4.42 |
| Calculated parameter 2 | ||||
| Forage % | 50.47 | 50.47 | 50.23 | 50.23 |
| SI | 0.88 | 0.87 | 0.88 | 0.87 |
| RDP (%CP) | 59.34 | 61.27 | 58.86 | 60.81 |
| RUP (%CP) | 40.66 | 38.73 | 41.14 | 39.19 |
| Chemical composition 3 (% DM) | ||||
| OM | 94.82 | 94.16 | 94.05 | 94.43 |
| CP | 15.22 | 15.28 | 15.07 | 15.07 |
| EE | 3.98 | 3.95 | 3.86 | 3.85 |
| NDF | 44.81 | 44.26 | 42.38 | 41.64 |
| ADF | 26.60 | 22.32 | 26.57 | 22.42 |
| Ash | 5.18 | 5.84 | 5.95 | 5.57 |
| NSC | 30.81 | 30.67 | 32.74 | 33.87 |
| NEL (Mcal/kg) | 1.59 | 1.60 | 1.63 | 1.63 |
| Treatment 1 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Item | T | S | Effect (p-Value) 2 | |||||||
| N | L | H | N | L | H | SEM | F | C | F × C | |
| Total VFA (mM) | 60.55 | 61.52 | 62.04 | 59.18 | 63.17 | 60.74 | 0.35 | 0.773 | 0.241 | 0.492 |
| VFA (%) 3 | ||||||||||
| Ac | 52.71 bc | 51.63 c | 52.09 bc | 54.40 a | 53.31 ab | 53.63 ab | 0.37 | <0.001 | 0.029 | 0.973 |
| Pr | 25.66 a | 24.59 ab | 23.80 ab | 23.72 ab | 22.64 b | 22.66 b | 0.59 | 0.003 | 0.060 | 0.737 |
| Bu | 18.62 | 19.16 | 19.99 | 18.00 | 19.26 | 18.89 | 0.47 | 0.177 | 0.062 | 0.455 |
| BCVFA | 3.03 | 4.62 | 4.12 | 3.88 | 4.79 | 4.82 | 0.62 | 0.272 | 0.131 | 0.850 |
| A/P ratio | 2.05 c | 2.10 c | 2.19 bc | 2.29 ab | 2.35 ab | 2.37 a | 0.06 | <0.001 | 0.233 | 0.836 |
| NH3 (mM) | 25.75 d | 27.68 bc | 33.22 a | 26.64 cd | 27.57 bc | 28.77 b | 0.45 | 0.006 | <0.001 | <0.001 |
| In vitro digestibility (%) | ||||||||||
| DM | 63.59 abc | 61.89 cd | 61.57 d | 65.00 a | 62.30 bcd | 63.74 ab | 0.53 | 0.003 | 0.011 | 0.327 |
| OM | 68.07 abc | 66.47 cd | 66.39 d | 69.56 a | 66.89 bcd | 68.63 ab | 0.57 | 0.020 | 0.032 | 0.060 |
| NDF | 62.86 a | 61.61 a | 57.64 b | 61.66 a | 57.24 b | 58.66 b | 0.76 | 0.076 | 0.001 | 0.002 |
| MCP synthesis (mg/g OMD) 4 | 59.57 b | 75.37 a | 76.79 a | 56.88 b | 70.93 a | 74.75 a | 1.31 | 0.118 | <0.001 | 0.874 |
| Treatment 1 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Item 3 | T | S | Effect (p-Value) 2 | |||||||
| N | L | H | N | L | H | SEM | F | C | F × C | |
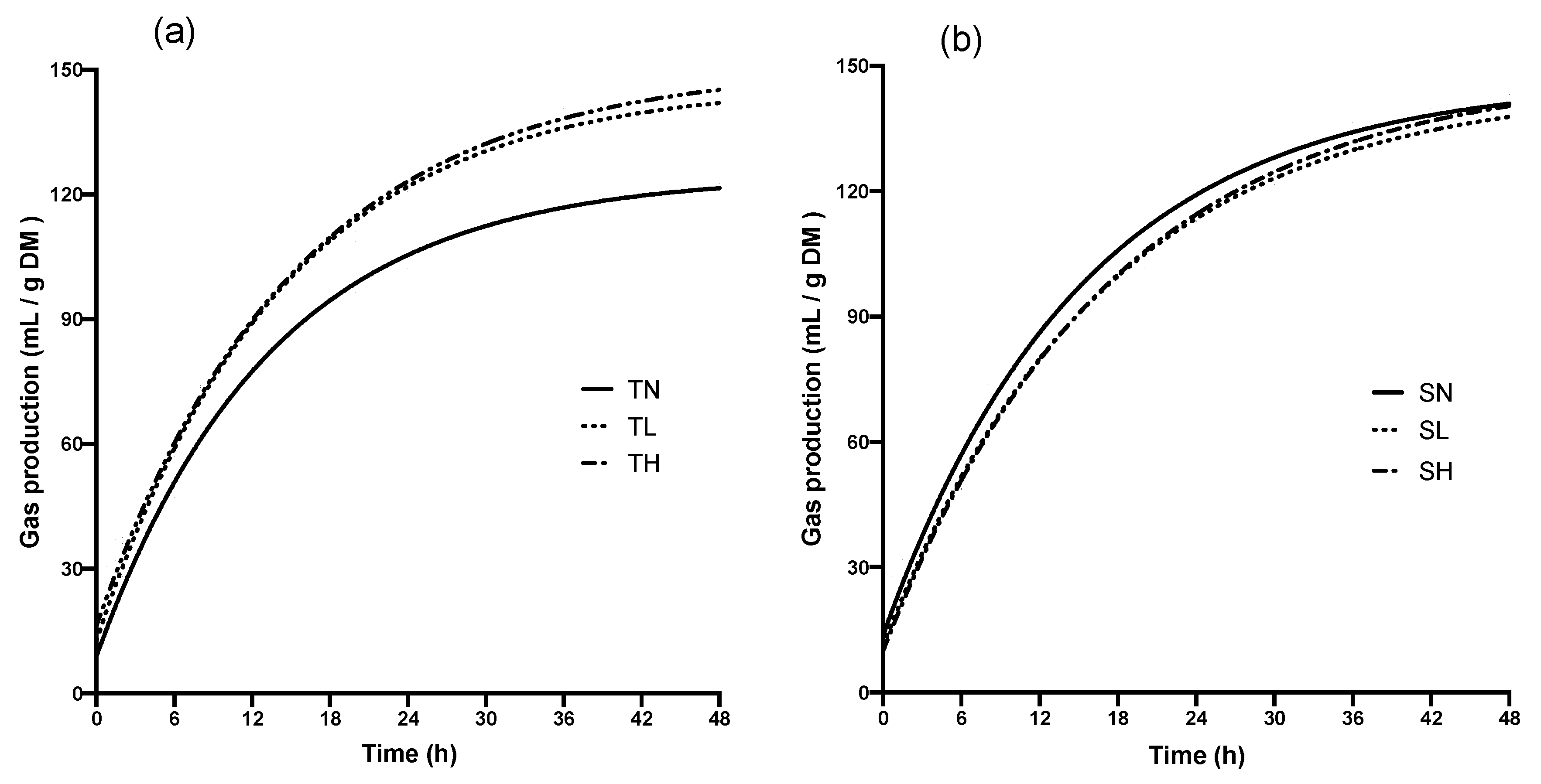
| GP (mL/g DM) | 166.56 b | 187.92 a | 193.94 a | 189.41 a | 190.23 a | 192.57 a | 0.94 | 0.005 | <0.001 | <0.001 |
| Fast % | 10.40 c | 13.29 b | 16.49 a | 15.97 a | 12.38 bc | 11.18 bc | 0.75 | 0.735 | 0.423 | <0.001 |
| K-fast (h−1) | 3.60 b | 3.33 bc | 4.39 a | 2.51 d | 2.97 c | 3.06 c | 0.13 | <0.001 | 0.000 | 0.003 |
| K-slow (h−1) | 0.066 a | 0.058 b | 0.056 b | 0.051 c | 0.050 c | 0.051 c | 0.001 | <0.001 | 0.005 | 0.003 |
| t1/2 -fast (h) | 0.19 c | 0.23 b | 0.16 d | 0.27 a | 0.23 bc | 0.23 b | 0.01 | <0.001 | <0.001 | <0.001 |
| t1/2 -slow (h) | 11.07 d | 11.92 cd | 12.45 bc | 14.14 a | 13.85 a | 13.55 ab | 0.38 | <0.001 | 0.576 | 0.057 |
| Span-fast (mL) | 15.21 d | 25.02 b | 32.19 a | 30.80 a | 23.53 bc | 20.70 c | 1.15 | 0.954 | 0.110 | <0.001 |
| Span-slow (mL) | 146.32 d | 163.28 bc | 163.11 bc | 158.89 c | 166.85 bc | 173.79 a | 2.08 | <0.001 | <0.001 | 0.126 |
| Treatment 1 | Effect (p-Value) 2 | |||||||
|---|---|---|---|---|---|---|---|---|
| Item 3 | HSN | HSV | LSN | LSV | SEM | SL | C | SL × C |
| pH | 5.62 b | 5.57 c | 5.68 a | 5.66 ab | 0.01 | <0.001 | 0.005 | 0.001 |
| Total VFAs (mM) | 89.04 | 88.48 | 89.00 | 85.60 | 2.85 | 0.620 | 0.501 | 0.625 |
| VFAs (%) | ||||||||
| Ac | 57.67 a | 56.71 b | 57.04 b | 55.10 b | 0.95 | 0.260 | 0.154 | 0.614 |
| Pr | 26.02 | 29.68 ab | 30.06 ab | 31.64 a | 1.27 | 0.035 | 0.061 | 0.428 |
| Bu | 13.71 | 13.61 | 12.90 | 13.26 | 0.34 | 0.111 | 0.712 | 0.508 |
| BCVFA | 2.61 | 2.69 | 2.62 | 2.52 | 0.13 | 0.561 | 0.956 | 0.488 |
| Ac/Pr ratio | 2.23 a | 1.96 ab | 1.90 ab | 1.75 b | 0.13 | 0.058 | 0.128 | 0.671 |
| NH3 (mM) | 34.47 | 35.50 | 35.47 | 34.19 | 0.46 | 0.852 | 0.880 | 0.163 |
| In vitro digestibility | ||||||||
| DM% | 66.89 a | 64.19 b | 63.08 bc | 62.04 c | 0.76 | <0.001 | 0.010 | 0.207 |
| OM% | 71.70 a | 68.80 a | 67.61 b | 66.49 ab | 0.82 | 0.001 | 0.013 | 0.257 |
| NDF% | 62.86 | 59.64 | 62.35 | 63.19 | 0.74 | 0.607 | 0.647 | 0.913 |
| MCP synthesis (mg/g OMD) 4 | 75.81 c | 78.23 c | 85.80 b | 92.19 a | 0.92 | <0.001 | 0.001 | 0.053 |
| Treatment 1 | Effect (p-Value) 2 | |||||||
|---|---|---|---|---|---|---|---|---|
| Item 3 | HSN | HSV | LSN | LSV | SEM | SL | M-CMS | SL × C |
| GP (mL/g DM) | 176.60 a | 172.96 ab | 170.20 b | 167.55 b | 0.72 | 0.001 | 0.049 | 0.736 |
| Fast % | 5.32 ab | 6.20 a | 6.13 a | 5.07 b | 0.27 | 0.583 | 0.737 | 0.004 |
| K-fast (h−1) | 2.080 b | 2.695 a | 2.134 b | 2.155 b | 0.16 | 0.151 | 0.068 | 0.087 |
| K-slow (h−1) | 0.097 c | 0.107 b | 0.094 c | 0.113 a | 0.001 | 0.321 | <0.0001 | 0.005 |
| t1/2 -fast (h) | 0.34 a | 0.26 b | 0.33 a | 0.32 a | 0.02 | 0.263 | 0.084 | 0.132 |
| t1/2 -slow (h) | 7.12 a | 6.47 b | 7.36 a | 6.14 b | 0.09 | 0.631 | <0.0001 | 0.008 |
| Span-fast (mL) | 9.43 b | 10.71 a | 10.44 a | 8.51 b | 0.45 | 0.219 | 0.482 | 0.004 |
| Span-slow (mL) | 177.39 a | 173.36 b | 170.46 bc | 167.86 c | 1.44 | 0.001 | 0.040 | 0.628 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-H.; Chen, C.-Y.; Wang, H.-T. The Effect of Forage Source and Concentrated Liquid Feedstuff Supplementation on Improving the Synchronization of Ruminant Dietary Energy and Nitrogen Release In Vitro. Fermentation 2022, 8, 443. https://doi.org/10.3390/fermentation8090443
Chen Y-H, Chen C-Y, Wang H-T. The Effect of Forage Source and Concentrated Liquid Feedstuff Supplementation on Improving the Synchronization of Ruminant Dietary Energy and Nitrogen Release In Vitro. Fermentation. 2022; 8(9):443. https://doi.org/10.3390/fermentation8090443
Chicago/Turabian StyleChen, Yi-Hsuan, Ching-Yi Chen, and Han-Tsung Wang. 2022. "The Effect of Forage Source and Concentrated Liquid Feedstuff Supplementation on Improving the Synchronization of Ruminant Dietary Energy and Nitrogen Release In Vitro" Fermentation 8, no. 9: 443. https://doi.org/10.3390/fermentation8090443