
The Addition of Biochar and Hyper-Thermal Inoculum Can Regulate the Fate of Heavy Metals Resistant Bacterial Communities during the Livestock Manure Composting

Abstract
:1. Introduction
2. Materials and Methods
2.1. Compost Materials and Preparations
2.2. Physicochemical Analysis and Enzyme Activities
2.3. Bacterial Community Detection
2.4. Statistical Analyses
3. Results and Discussion
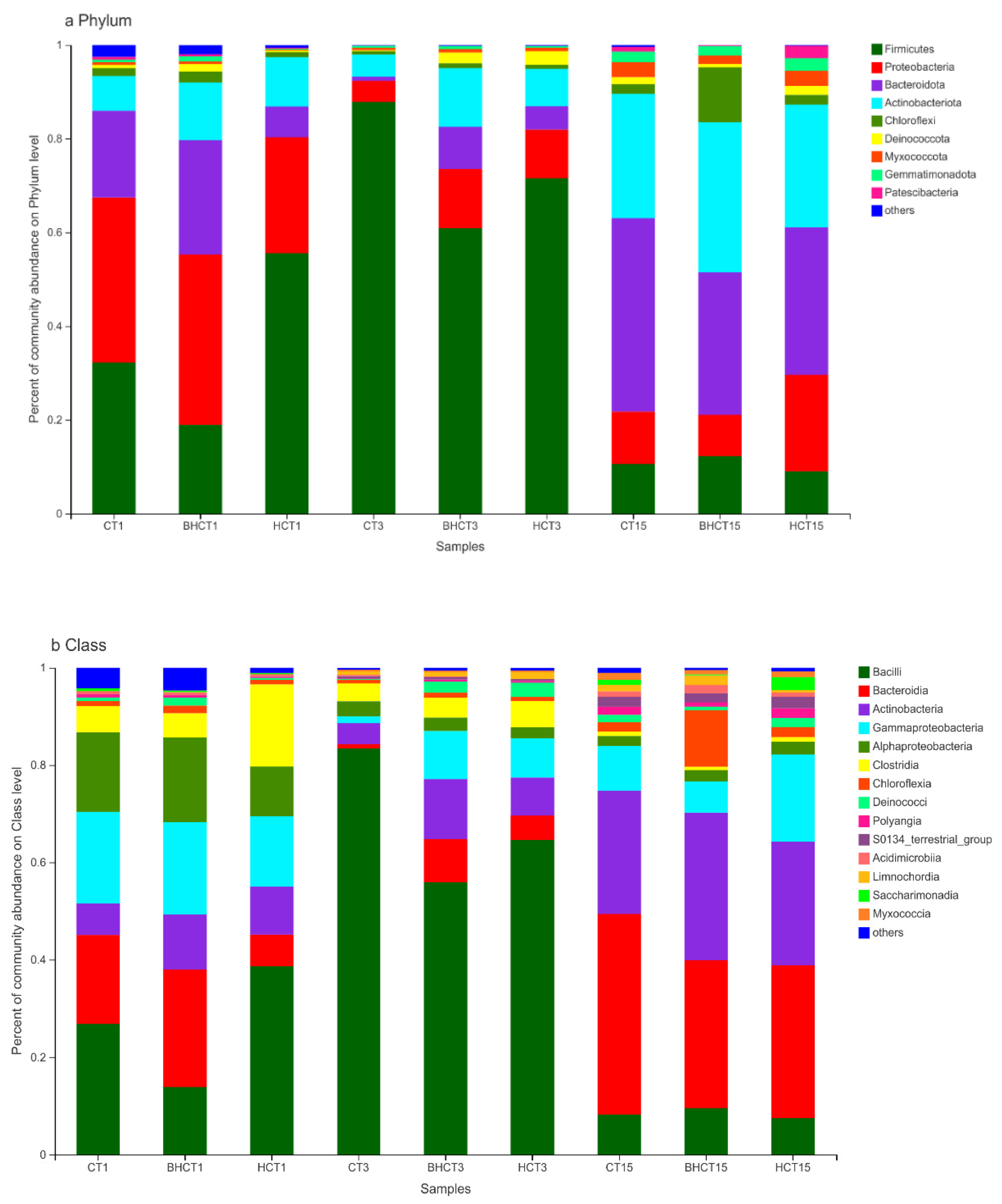
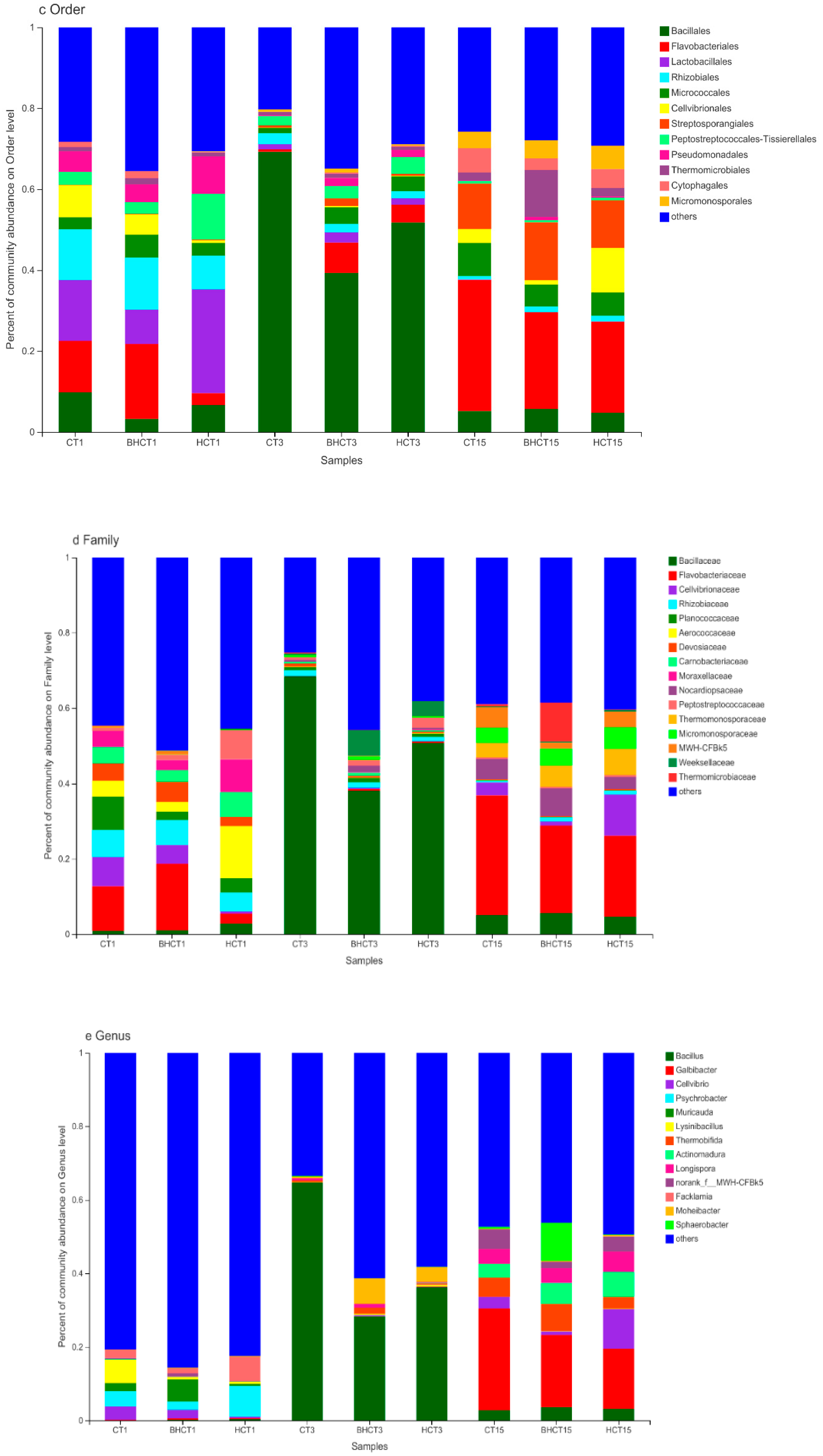
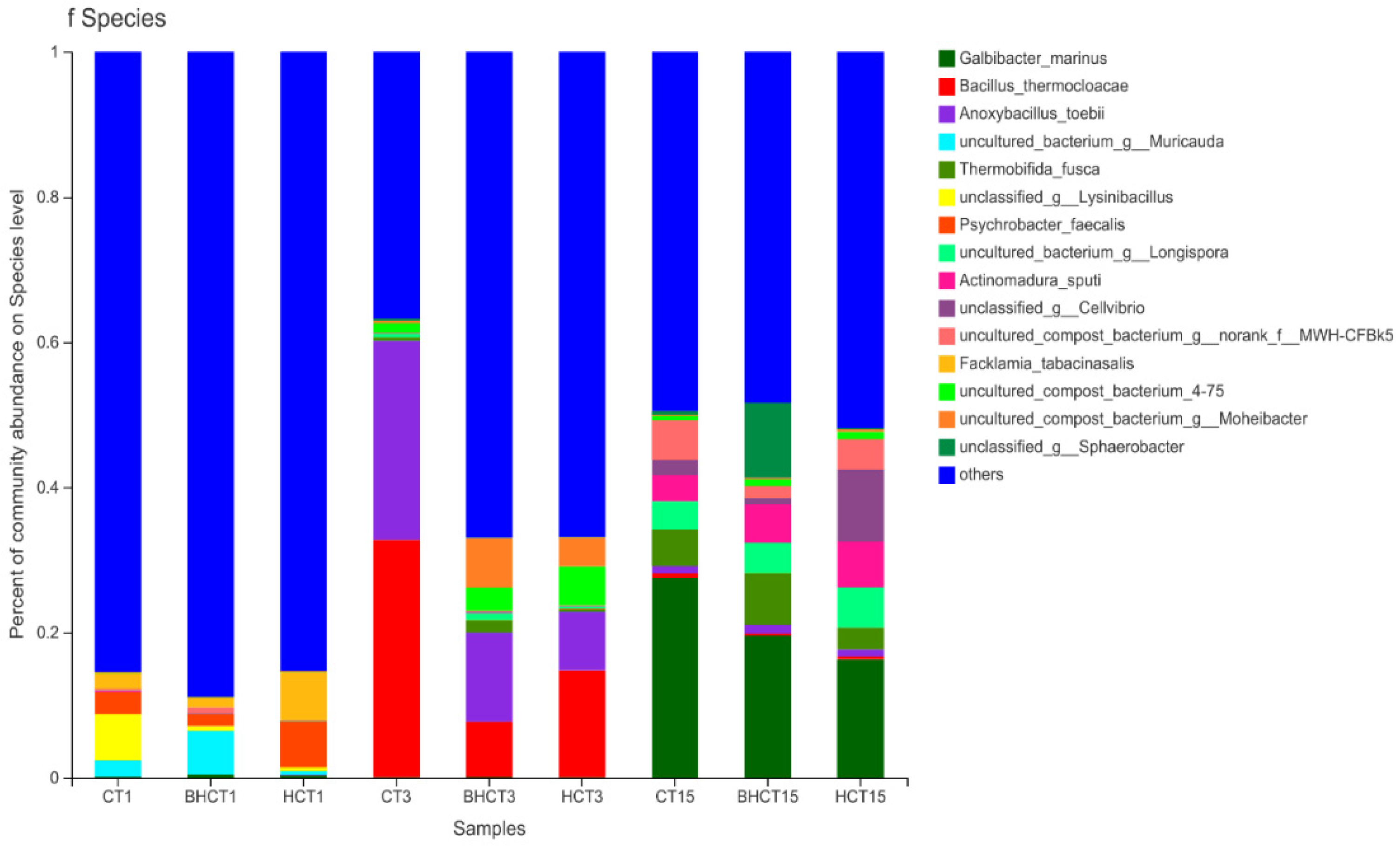
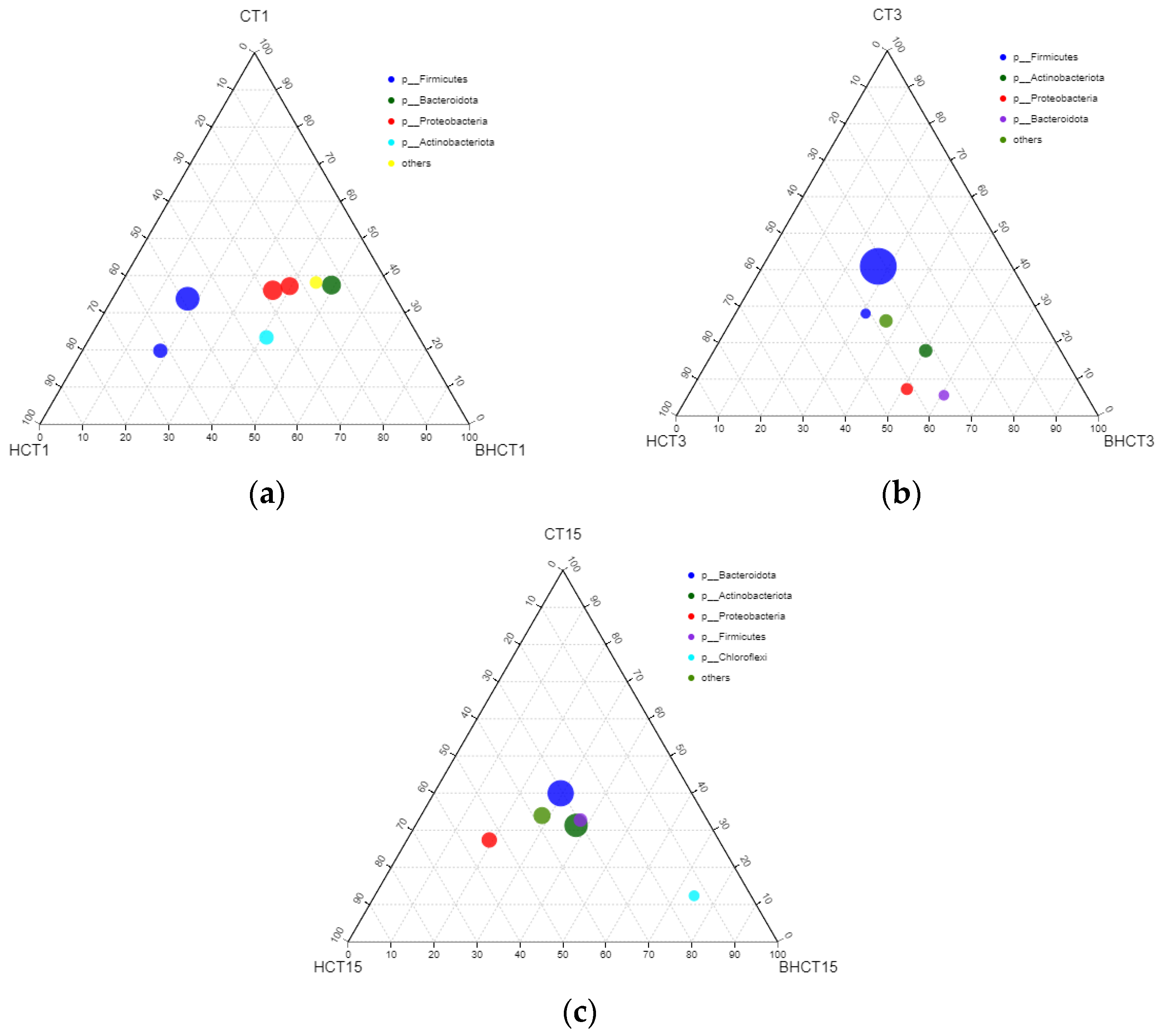
3.1. Species Assessment and Composition Analysis of Bacteria during Composting
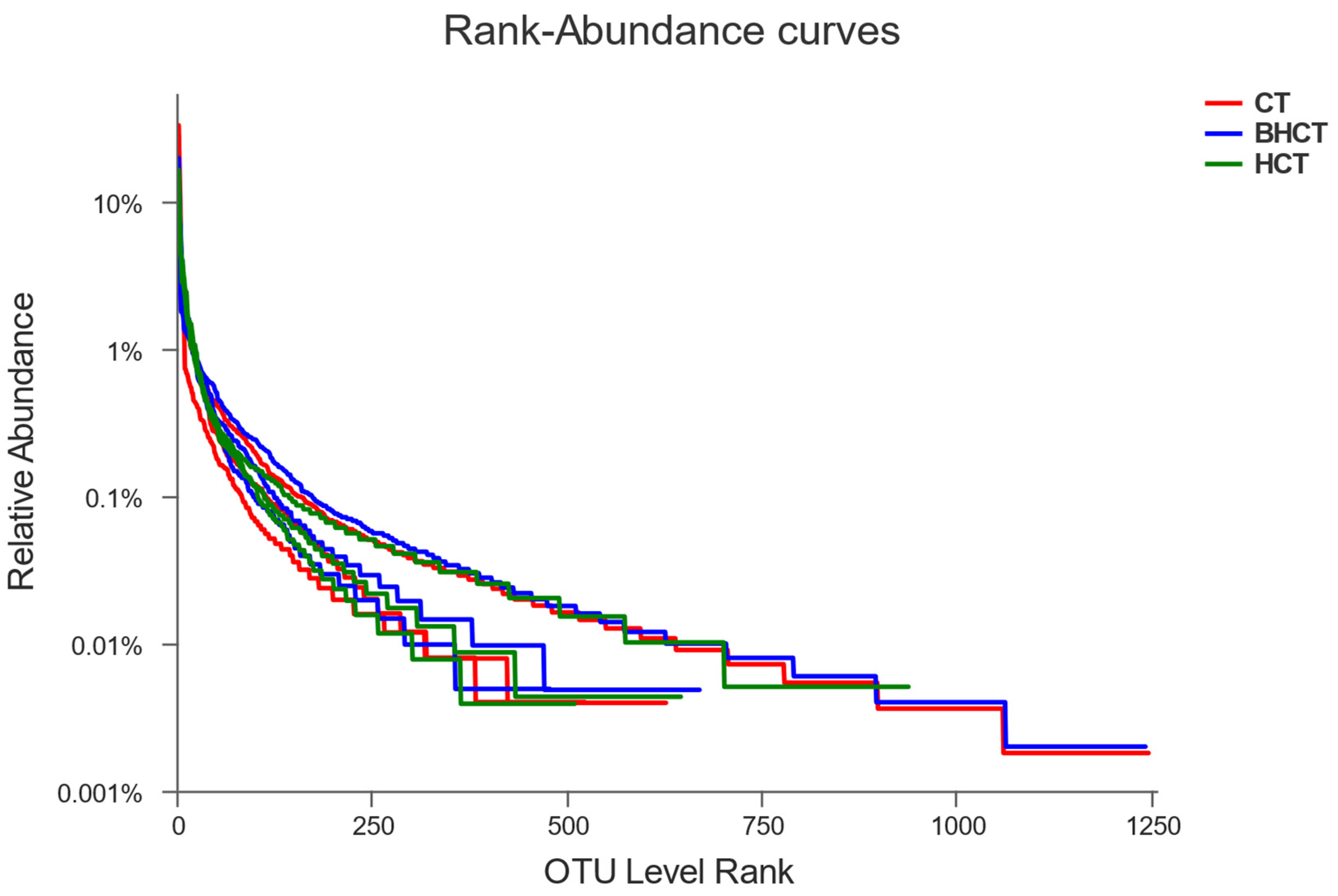
3.1.1. Annotation and Evaluation of Operational Taxonomic Units (OTUs)
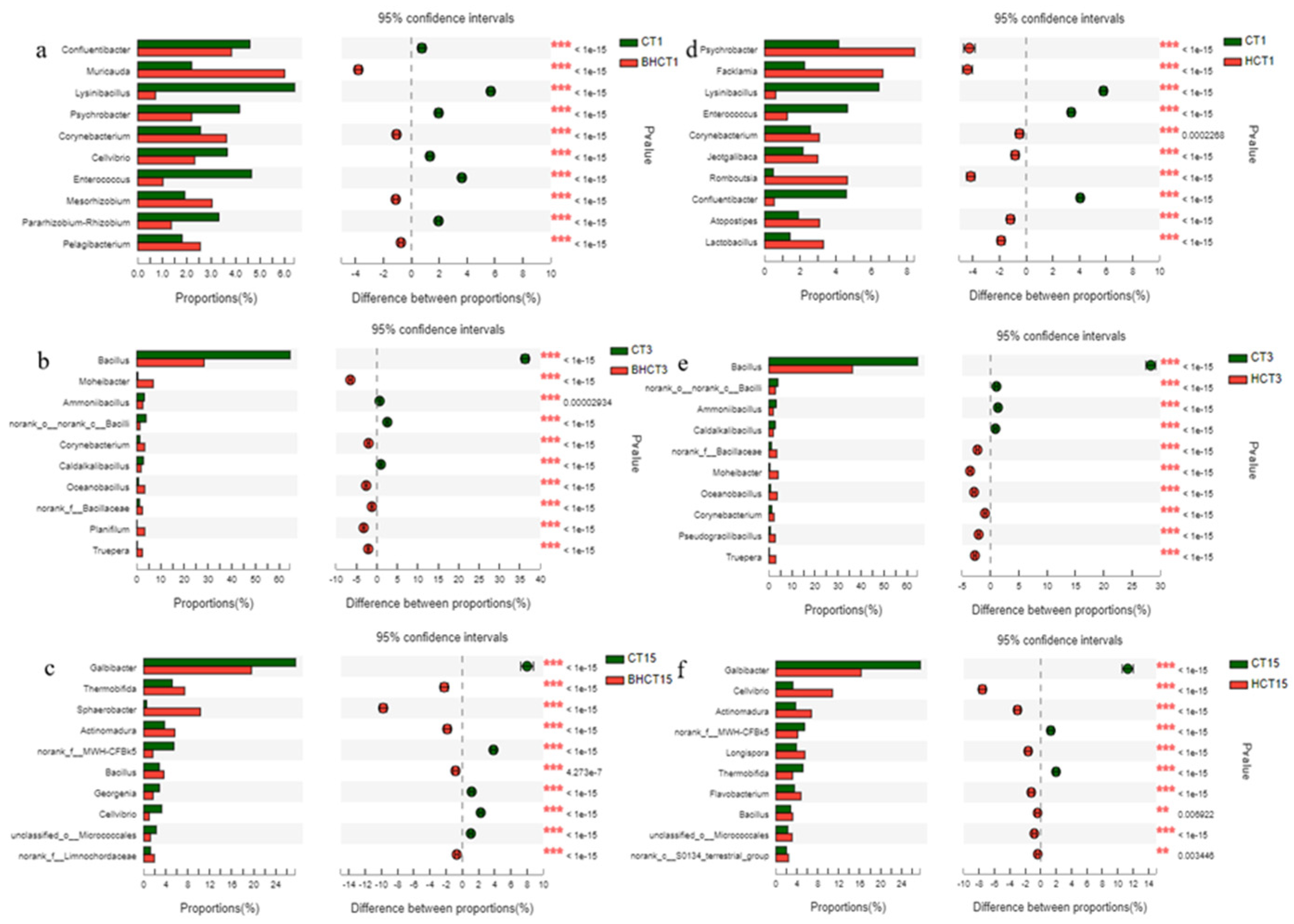
3.1.2. Relative Abundance of HMRB during Composting
3.2. Diversity Analysis of HMRB
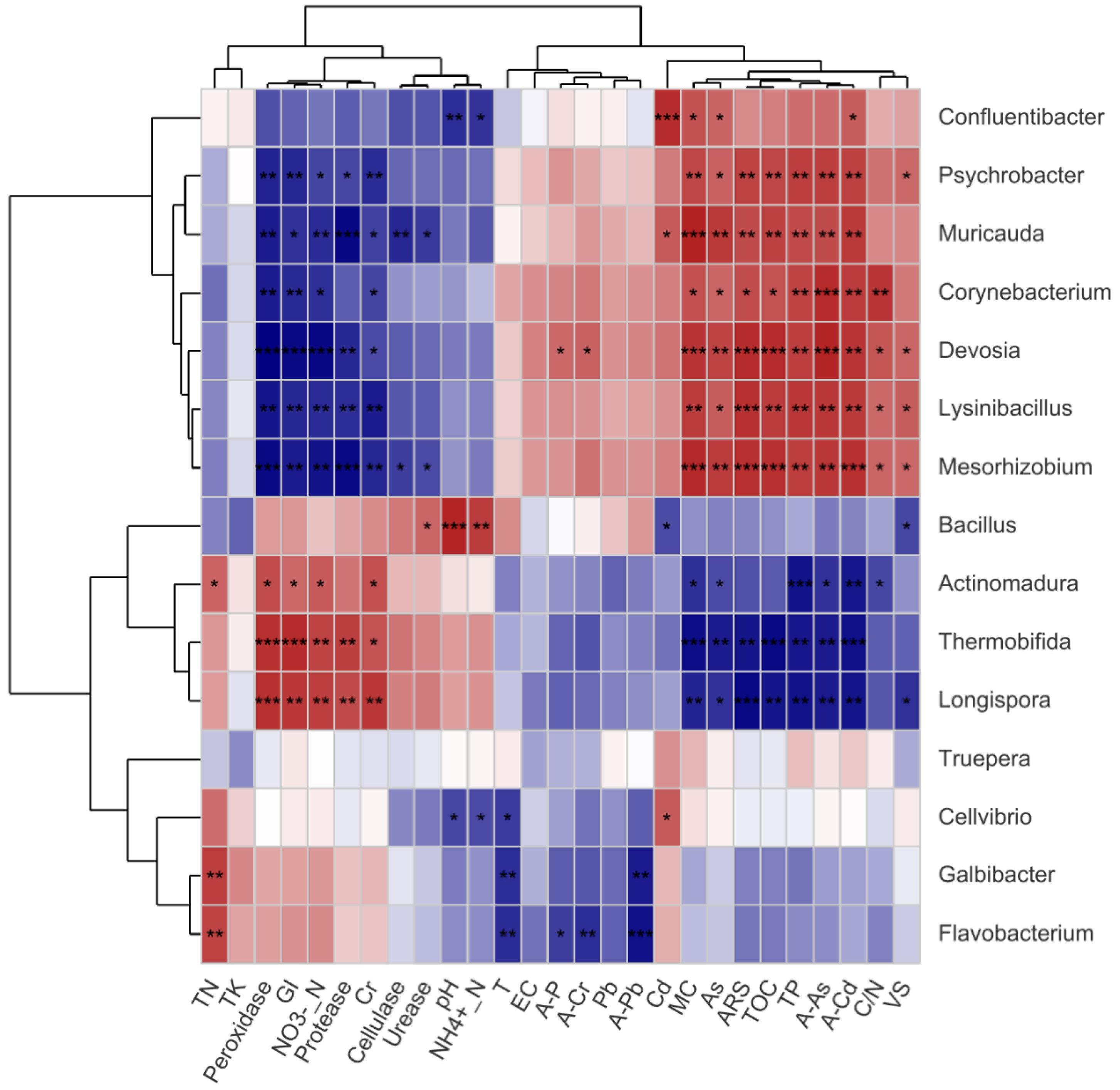
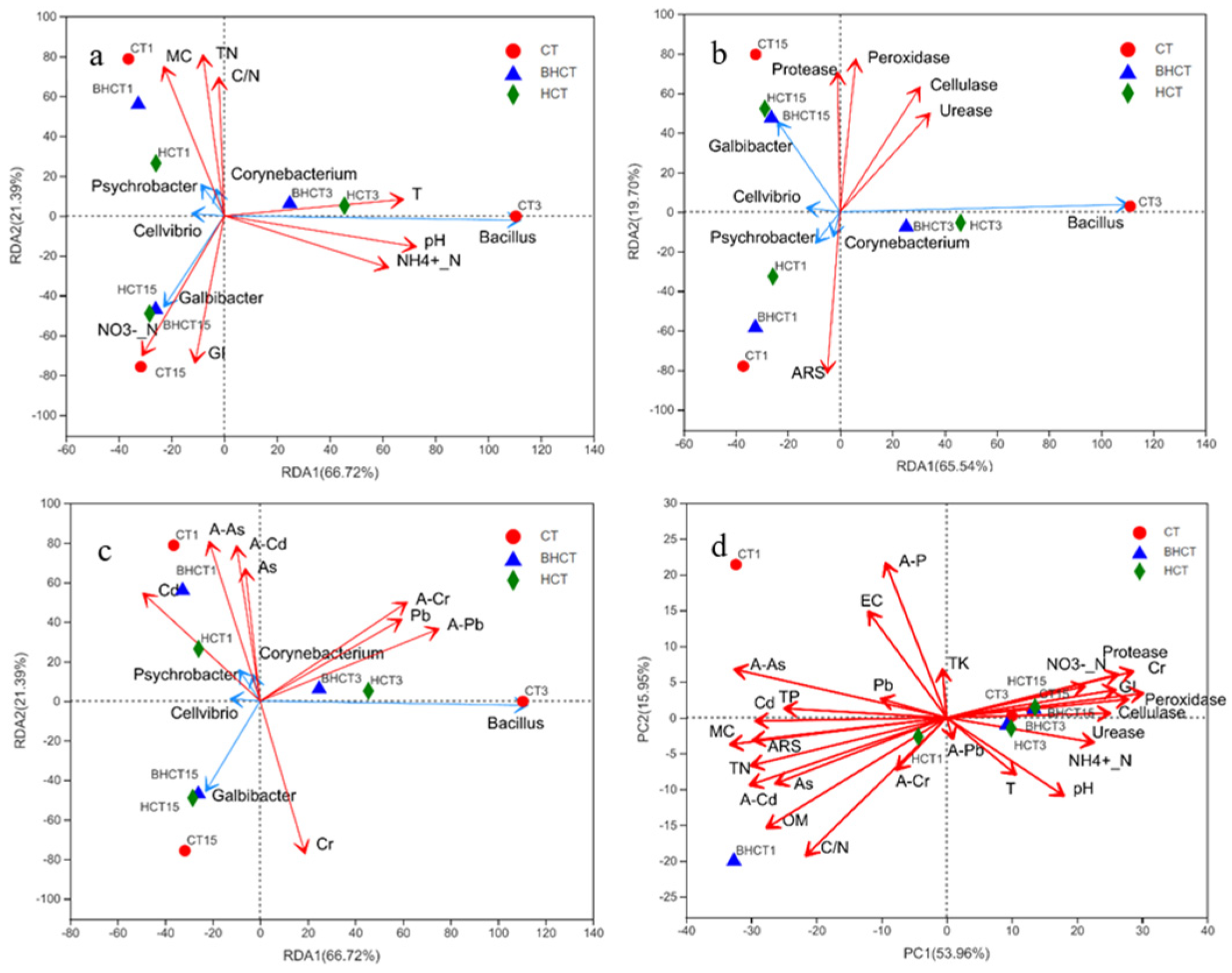
3.3. Correlation Analysis among HMRB Communities and Selected Factors
3.3.1. Correlation between the HMRB and Physicochemical Parameters
3.3.2. Correlation between the HMRB and Enzyme Activity
3.3.3. Correlation between HMRB Communities and Heavy Metals
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abd Elhafez, S.; Hamad, H.; Zaatout, A.; Malash, G. Management of agricultural waste for removal of heavy metals from aqueous solution: Adsorption behaviors, adsorption mechanisms, environmental protection and techno-economic analysis. Environ. Sci. Pollut. Res. 2017, 24, 1397–1415. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Xie, S.; Wang, Y.; Pan, X.; Yu, G.; Zhang, Y. Simultaneous heavy metal immobilization and antibiotics removal during synergetic treatment of sewage sludge and pig manure. Environ. Sci. Pollut. Res. 2020, 27, 30323–30332. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Li, X. Effects of Microbial Inoculation with Different Indigenous Bacillus Species on Physicochemical Characteristics and Bacterial Succession during Short-Term Composting. Fermentation 2022, 8, 152. [Google Scholar] [CrossRef]
- Awasthi, S.K.; Duan, Y.; Liu, T.; Zhang, Z.; Taherzadeh, M.J. Can biochar regulate the fate of heavy metals (Cu and Zn) resistant bacteria community during the poultry manure composting? J. Hazard. Mater. 2021, 406, 124593. [Google Scholar] [CrossRef]
- Li, Z.; Fan, Y.; Huang, X.; Han, B. Microbial Diversity and Metabolites Dynamic of Light-Flavor Baijiu with Stacking Process. Fermentation 2022, 8, 67. [Google Scholar] [CrossRef]
- Cui, P.; Liao, H.; Bai, Y.; Li, X.; Zhao, Q.; Chen, Z.; Yu, Z.; Yi, Z.; Zhou, S. Hyperthermophilic composting reduces nitrogen loss via inhibiting ammonifiers and enhancing nitrogenous humic substance formation. Sci. Total Environ. 2019, 692, 98–106. [Google Scholar] [CrossRef]
- Ai, Y.-J.; Li, F.-P.; Gu, H.-H.; Chi, X.-J.; Yuan, X.-T.; Han, D.-Y. Combined effects of green manure returning and addition of sewage sludge compost on plant growth and microorganism communities in gold tailings. Environ. Sci. Pollut. Res. 2020, 27, 31686–31698. [Google Scholar] [CrossRef]
- Cao, Y.; Wang, L.; Qian, Y.; Xu, Y.; Wu, H.; Zhang, J.; Huang, H.; Chang, Z. Contributions of thermotolerant bacteria to organic matter degradation under a hyperthermophilic pretreatment process during chicken manure composting. BioResources 2019, 14, 6747–6766. [Google Scholar] [CrossRef]
- Guo, H.; Gu, J.; Wang, X.; Yu, J.; Nasir, M.; Peng, H.; Zhang, R.; Hu, T.; Wang, Q.; Ma, J. Responses of antibiotic and heavy metal resistance genes to bamboo charcoal and bamboo vinegar during aerobic composting. Environ. Pollut. 2019, 252, 1097–1105. [Google Scholar] [CrossRef]
- Liao, H.; Lu, X.; Rensing, C.; Friman, V.P.; Geisen, S.; Chen, Z.; Yu, Z.; Wei, Z.; Zhou, S.; Zhu, Y. Hyperthermophilic Composting Accelerates the Removal of Antibiotic Resistance Genes and Mobile Genetic Elements in Sewage Sludge. Environ Sci. Technol. 2018, 52, 266–276. [Google Scholar] [CrossRef] [Green Version]
- Awasthi, S.K.; Liu, T.; Awasthi, M.K.; Zhang, Z. Evaluation of biochar amendment on heavy metal resistant bacteria abundance in biosolids compost. Bioresour. Technol. 2020, 306, 123114. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Awasthi, S.K.; Liu, T.; Verma, S.; Zhang, Z.; Pandey, A.; Varjani, S.; Li, R.; Taherzadeh, M.J.; Awasthi, M.K. Patterns of heavy metal resistant bacterial community succession influenced by biochar amendment during poultry manure composting. J. Hazard. Mater. 2021, 420, 126562. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Awasthi, S.K.; Liu, T.; Verma, S.; Wang, Q.; Chen, H.; Ren, X.; Zhang, Z.; Awasthi, M.K. Positive impact of biochar alone and combined with bacterial consortium amendment on improvement of bacterial community during cow manure composting. Bioresour. Technol. 2019, 280, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Tang, J.; Liao, H.; Liu, X.; Zhou, P.; Chen, Z.; Rensing, C.; Zhou, S. The distinctive microbial community improves composting efficiency in a full-scale hyperthermophilic composting plant. Bioresour. Technol. 2018, 265, 146–154. [Google Scholar] [CrossRef]
- Liu, X.; Hou, Y.; Li, Z.; Yu, Z.; Tang, J.; Wang, Y.; Zhou, S. Hyperthermophilic composting of sewage sludge accelerates humic acid formation: Elemental and spectroscopic evidence. Waste Manag. 2020, 103, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Sun, K.; Yang, Y.; Wang, Z.; Han, L.; Wang, X.; Wu, F.; Xing, B. Comparison between soil-and biochar-derived humic acids: Composition, conformation, and phenanthrene sorption. Environ. Sci. Technol. 2018, 52, 1880–1888. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Zhao, Y.; Zhao, X.; Gao, X.; Zheng, Y.; Zuo, H.; Wei, Z. Roles of different humin and heavy-metal resistant bacteria from composting on heavy metal removal. Bioresour. Technol. 2020, 296, 122375. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, Y.; Zhang, Z.; Wei, Y.; Wang, H.; Lu, Q.; Li, Y.; Wei, Z. Effect of thermo-tolerant actinomycetes inoculation on cellulose degradation and the formation of humic substances during composting. Waste Manag. 2017, 68, 64–73. [Google Scholar] [CrossRef]
- Oustriere, N.; Marchand, L.; Rosette, G.; Friesl-Hanl, W.; Mench, M. Wood-derived-biochar combined with compost or iron grit for in situ stabilization of Cd, Pb, and Zn in a contaminated soil. Environ. Sci. Pollut. Res. 2017, 24, 7468–7481. [Google Scholar] [CrossRef]
- Handa, S.; Sharma, N.; Pathania, S. Multiple parameter optimization for maximization of pectinase production by Rhizopus sp. C4 under solid state fermentation. Fermentation 2016, 2, 10. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.-X.; Xiao, Q.; Shen, Y.-F.; Li, S.-Q. Effects of biochar and maize straw on the short-term carbon and nitrogen dynamics in a cultivated silty loam in China. Environ. Sci. Pollut. Res. 2017, 24, 1019–1029. [Google Scholar] [CrossRef] [PubMed]
- Esperón, F.; Albero, B.; Ugarte-Ruíz, M.; Domínguez, L.; Carballo, M.; Tadeo, J.L.; del Mar Delgado, M.; Moreno, M.Á.; de la Torre, A. Assessing the benefits of composting poultry manure in reducing antimicrobial residues, pathogenic bacteria, and antimicrobial resistance genes: A field-scale study. Environ. Sci. Pollut. Res. 2020, 27, 27738–27749. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.-Y.; Zhang, J.; Zhong, X.-Z.; Tan, L.; Tang, Y.-Q.; Kida, K. Production of nitrate-rich compost from the solid fraction of dairy manure by a lab-scale composting system. Waste Manag. 2016, 51, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Chen, Z.; Li, Y.; Ding, K.; Liu, W.; Liu, Y.; Yuan, Y.; Zhang, M.; Baker, A.J.; Yang, W.; et al. Factors influencing heavy metal availability and risk assessment of soils at typical metal mines in Eastern China. J. Hazard. Mater. 2020, 400, 123289. [Google Scholar] [CrossRef]
- Cardelli, V.; De Feudis, M.; Fornasier, F.; Massaccesi, L.; Cocco, S.; Agnelli, A.; Weindorf, D.C.; Corti, G. Changes of topsoil under Fagus sylvatica along a small latitudinal-altitudinal gradient. Geoderma 2019, 344, 164–178. [Google Scholar] [CrossRef]
- Wei, H.; Wang, L.; Hassan, M.; Xie, B. Succession of the functional microbial communities and the metabolic functions in maize straw composting process. Bioresour. Technol. 2018, 256, 333–341. [Google Scholar] [CrossRef]
- Awasthi, M.K.; Duan, Y.; Awasthi, S.K.; Liu, T.; Chen, H.; Pandey, A.; Zhang, Z.; Taherzadeh, M.J. Emerging applications of biochar: Improving pig manure composting and attenuation of heavy metal mobility in mature compost. J. Hazard. Mater. 2020, 389, 122116. [Google Scholar] [CrossRef]
- Zhang, R.; Gu, J.; Wang, X.; Yang, L.; Zhang, K.; Yin, Y.; Zhang, X. Contributions of the microbial community and environmental variables to antibiotic resistance genes during co-composting with swine manure and cotton stalks. J. Hazard. Mater. 2018, 358, 82–91. [Google Scholar] [CrossRef]
- Forsberg, K.J.; Patel, S.; Gibson, M.K.; Lauber, C.L.; Knight, R.; Fierer, N.; Dantas, G. Bacterial phylogeny structures soil resistomes across habitats. Nature 2014, 509, 612. [Google Scholar] [CrossRef] [Green Version]
- Sui, Q.; Zhang, J.; Chen, M.; Wang, R.; Wang, Y.; Wei, Y. Fate of microbial pollutants and evolution of antibiotic resistance in three types of soil amended with swine slurry. Environ. Pollut. 2019, 245, 353–362. [Google Scholar] [CrossRef]
- Huang, W.; Li, Y.; Liu, X.; Wang, W.; Zhou, S. Linking the electron transfer capacity with the compositional characteristics of dissolved organic matter during hyperthermophilic composting. Sci. Total Environ. 2021, 755, 142687. [Google Scholar] [CrossRef] [PubMed]
- Riaz, L.; Wang, Q.; Yang, Q.; Li, X.; Lixunan, L. Potential of industrial composting and anaerobic digestion for the removal of antibiotics, antibiotic resistance genes and heavy metals from chicken manure. Sci. Total Environ. 2020, 718, 137414. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Zhou, D.; Wei, H.; Wu, S.; Xie, B. Alleviating soil degradation caused by watermelon continuous cropping obstacle: Application of urban waste compost. Chemosphere 2021, 262, 128387. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Nasir, M.; Lv, J.; Dai, Y.; Gao, J. Understanding the variation of microbial community in heavy metals contaminated soil using high throughput sequencing. Ecotoxicol. Environ. Saf. 2017, 144, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Zhai, W.; Wong, M.T.; Luo, F.; Hashmi, M.Z.; Liu, X.; Edwards, E.A.; Tang, X.; Xu, J. Arsenic Methylation and its Relationship to Abundance and Diversity of arsM Genes in Composting Manure. Sci. Rep. 2017, 7, srep42198. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.C., IV; Oladeinde, A.; Kieran, T.J.; Finger, J.W., Jr.; Bayona-Vásquez, N.J.; Cartee, J.C.; Beasley, J.C.; Seaman, J.C.; McArthur, J.V.; Rhodes, O.E., Jr. Co-occurrence of antibiotic, biocide, and heavy metal resistance genes in bacteria from metal and radionuclide contaminated soils at the Savannah River Site. Microb. Biotechnol. 2020, 13, 1179–1200. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Rajkumar, M.; Vicente, J.; Freitas, H. Inoculation of Ni-resistant plant growth promoting bacterium Psychrobacter sp. strain, S.R.; S8 for the improvement of nickel phytoextraction by energy crops. Int. J. Phytoremed. 2010, 13, 126–139. [Google Scholar] [CrossRef]
- Fang, Y.; Jia, X.; Chen, L.; Lin, C.; Zhang, H.; Chen, J. Effect of thermotolerant bacterial inoculation on the microbial community during sludge composting. Can. J. Microbiol. 2019, 65, 750–761. [Google Scholar] [CrossRef]
- Mei, J.; Wu, Y.; Qian, F.; Chen, C.; Shen, Y.; Zhao, Y. Methane-oxidizing microorganism properties in landfills. Pol. J. Environ. Stud. 2019, 28, 3809–3818. [Google Scholar]
- Chen, X.; Zhao, Y.; Zhang, C.; Zhang, D.; Yao, C.; Meng, Q.; Zhao, R.; Wei, Z. Speciation, toxicity mechanism and remediation ways of heavy metals during composting: A novel theoretical microbial remediation method is proposed. J. Environ. Manag. 2020, 272, 111109. [Google Scholar] [CrossRef]
- Zhu, L.; Zhao, Y.; Zhang, W.; Zhou, H.; Chen, X.; Li, Y.; Wei, D.; Wei, Z. Roles of bacterial community in the transformation of organic nitrogen toward enhanced bioavailability during composting with different wastes. Bioresour. Technol. 2019, 285, 121326. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Zhang, Y.; Qu, M.; Yin, Y.; Fan, K.; Hu, B.; Zhang, H.; Wei, M.; Ma, C. Effects of biochar on the microbial activity and community structure during sewage sludge composting. Bioresour. Technol. 2019, 272, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhou, Y.; Qin, S.; Awasth, S.K.; Liu, T.; Liu, H.; Zhang, Z.; Awasthi, M.K. Distribution of heavy metal resistant bacterial community succession in cow manure biochar amended sheep manure compost. Bioresour. Technol. 2021, 335, 125282. [Google Scholar] [CrossRef]
- Harindintwali, J.D.; Zhou, J.; Yu, X. Lignocellulosic crop residue composting by cellulolytic nitrogen-fixing bacteria: A novel tool for environmental sustainability. Sci. Total Environ. 2020, 715, 136912. [Google Scholar] [CrossRef]
- Rajapaksha, A.U.; Chen, S.S.; Tsang, D.C.; Zhang, M.; Vithanage, M.; Mandal, S.; Gao, B.; Bolan, N.S.; Ok, Y.S. Engineered/designer biochar for contaminant removal/immobilization from soil and water: Potential and implication of biochar modification. Chemosphere 2016, 148, 276–291. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Gu, J.; Wang, X.; Wang, Q.; Sun, W.; Hu, T.; Guo, H.; Ma, J.; Bao, J. Insight into the fate of antibiotic resistance genes and bacterial community in co-composting green tea residues with swine manure. J. Environ. Manag. 2020, 266, 110581. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Ou, Y.; Wang, L.; Yan, B.; Li, Y.; Bao, M. Critical passivation mechanisms on heavy metals during aerobic composting with different grain-size zeolite. J. Hazard. Mater. 2021, 406, 124313. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Y.; Li, Y.; Wu, Y.; Zeng, Z.; Xu, R.; Wang, S.; Li, H.; Zhang, J. Changes of heavy metal fractions during co-composting of agricultural waste and river sediment with inoculation of Phanerochaete chrysosporium. J. Hazard. Mater. 2019, 378, 120757. [Google Scholar] [CrossRef]
- Wu, D.; Wei, Z.; Qu, F.; Mohamed, T.A.; Zhu, L.; Zhao, Y.; Jia, L.; Zhao, R.; Liu, L.; Li, P. Effect of Fenton pretreatment combined with bacteria inoculation on humic substances formation during lignocellulosic biomass composting derived from rice straw. Bioresour. Technol. 2020, 303, 122849. [Google Scholar] [CrossRef]
- Aponte, H.; Meli, P.; Butler, B.; Paolini, J.; Matus, F.; Merino, C.; Cornejo, P.; Kuzyakov, Y. Meta-analysis of heavy metal effects on soil enzyme activities. Sci. Total Environ. 2020, 737, 139744. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, Y.; Li, Y.; Wu, Y.; Chen, Y.; Zeng, G.; Zhang, J.; Li, H. Influence of biochar on heavy metals and microbial community during composting of river sediment with agricultural wastes. Bioresour. Technol. 2017, 243, 347–355. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Composting Raw Materials | Initial Mixture | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Chicken Manure | Cow Dung | Wheat Straw | Biochar | Saw Dust | Hyper-Thermal Inoculum | CT | HCT | BHCT | |
| Weight (kg) | 105 ± 1.51 | 255 ± 1.63 | 60 ± 1.01 | 7.5 ± 0.23 | 22.5 ± 0.32 | 15.0 ± 0.23 | 153.21 ± 1.34 | 155.19 ± 1.69 | 156.6 ± 1.76 |
| Moisture (%) | 67.85 ± 0.80 | 65.21 ± 0.93 | 7.64 ± 0.12 | 1.53 ± 0.08 | 8.63 ± 0.09 | 10.32 ± 0.53 | 56.61 ± 0.29 | 52.07 ± 0.68 | 53.72 ± 0.76 |
| pH | 6.79 ± 0.07 | 6.83 ± 0.05 | 7.23 ± 0.03 | 8.69 ± 0.05 | 8.45 ± 0.03 | 8.15 ± 0.06 | 7.69 ± 0.09 | 8.02 ± 0.11 | 7.92 ± 0.06 |
| TOC (%) | 55.31 ± 1.03 | 51.81 ± 0.87 | 62.35 ± 0.86 | 78.03 ± 0.56 | 58.76 ± 0.56 | 63.78 ± 0.91 | 52.72 ± 0.87 | 53.48 ± 0.76 | 64.39 ± 0.92 |
| TN (%) | 2.13 ± 0.02 | 1.62 ± 0.06 | 5.32 ± 0.06 | 0.59 ± 0.05 | 0.49 ± 0.02 | 0.79 ± 0.04 | 1.74 ± 0.02 | 1.65 ± 0.07 | 1.80 ± 0.09 |
| C/N | 25.97 ± 1.02 | 31.98 ± 1.08 | 11.72 ± 0.96 | 132.25 ± 1.09 | 121.21 ± 1.06 | 80.25 ± 1.32 | 30.03 ± 0.76 | 32.41 ± 0.84 | 35.77 ± 0.93 |
| Parameter | CT (Initial Value) | BHCT (Initial Value) | HCT (Initial Value) | CT (Final Value) | BHCT (Final Value) | HCT (Final Value) |
|---|---|---|---|---|---|---|
| pH | 6.79 ± 0.07 | 6.83 ± 0.05 | 7.23 ± 0.03 | 8.15 ± 0.06 | 8.02 ± 0.11 | 7.92 ± 0.06 |
| TN | 1.74 ± 0.02 | 1.65 ± 0.07 | 1.80 ± 0.09 | 0.95 ± 0.03 | 1.19 ± 0.05 | 0.98 ± 0.01 |
| C/N | 30.03 ± 0.76 | 32.41 ± 0.84 | 35.77 ± 0.93 | 24.37± | 20.73± | 21.41± |
| As (mg kg −1) | 6.07 ± 0.04 | 6.68 ± 0.01 | 6.37 ± 0.01 | 4.33 ± 0.03 | 2.49 ± 0.04 | 3.61 ± 0.01 |
| A-As (mg kg −1) | 0.02 | 0.02 | 0.02 | 0.01 | 0.01 | 0.01 |
| Pb (mg kg −1) | 22.25 ± 0.05 | 21.92 ± 0.07 | 20.25 ± 0.05 | 21.03 ± 0.04 | 19.07 ± 0.02 | 20.11 ± 0.07 |
| A-Pb (mg kg −1) | 3.45 ± 0.01 | 3.35 ± 0.01 | 3.35 ± 0.02 | 1.25 ± 0.02 | 1.06 ± 0.03 | 1.27 ± 0.01 |
| Cd (mg kg −1) | 0.48 ± 0.01 | 0.48 ± 0.02 | 0.43 ± 0.01 | 0.40 ± 0.01 | 0.35 ± 0.01 | 0.44 ± 0.03 |
| A-Cd (mg kg −1) | 0.14 | 0.15 | 0.13 | 0.10 | 0.08 | 0.09 |
| Cr (mg kg −1) | 38.17 ± 0.27 | 37.11 ± 0.34 | 37.95 ± 0.25 | 51.41 ± 0.16 | 47.37 ± 0.1 | 52.69 ± 0.15 |
| A-Cr (mg kg −1) | 5.11 ± 0.05 | 5.41 ± 0.07 | 5.57 ± 0.03 | 4.19 ± 0.06 | 3.30 ± 0.05 | 3.98 ± 0.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.; Zhu, T.; Xiao, Q.; An, N. The Addition of Biochar and Hyper-Thermal Inoculum Can Regulate the Fate of Heavy Metals Resistant Bacterial Communities during the Livestock Manure Composting. Fermentation 2022, 8, 207. https://doi.org/10.3390/fermentation8050207
Zhang Q, Zhu T, Xiao Q, An N. The Addition of Biochar and Hyper-Thermal Inoculum Can Regulate the Fate of Heavy Metals Resistant Bacterial Communities during the Livestock Manure Composting. Fermentation. 2022; 8(5):207. https://doi.org/10.3390/fermentation8050207
Chicago/Turabian StyleZhang, Qingjun, Tong Zhu, Qingxiang Xiao, and Ning An. 2022. "The Addition of Biochar and Hyper-Thermal Inoculum Can Regulate the Fate of Heavy Metals Resistant Bacterial Communities during the Livestock Manure Composting" Fermentation 8, no. 5: 207. https://doi.org/10.3390/fermentation8050207