
Microbial Quality and Growth Dynamics in Shameta: A Traditional Ethiopian Cereal-Based Fermented Porridge

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
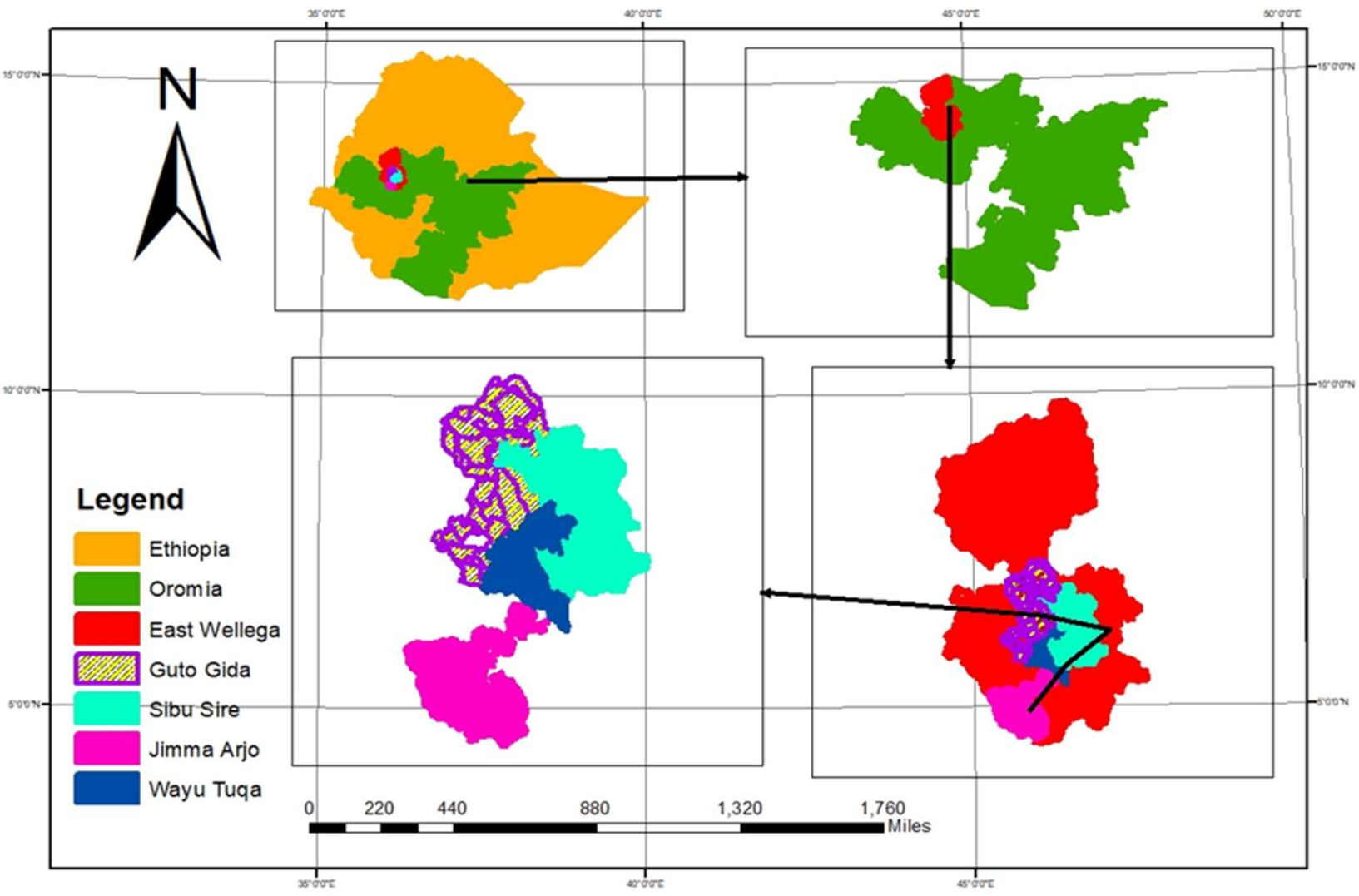
2.1. Description of the Study Area
2.2. Shameta Sample Collection and Laboratory-Scale Fermentation of Shameta
2.2.1. Study Population and Household Shameta Sampling
2.2.2. Collection of Shameta Samples
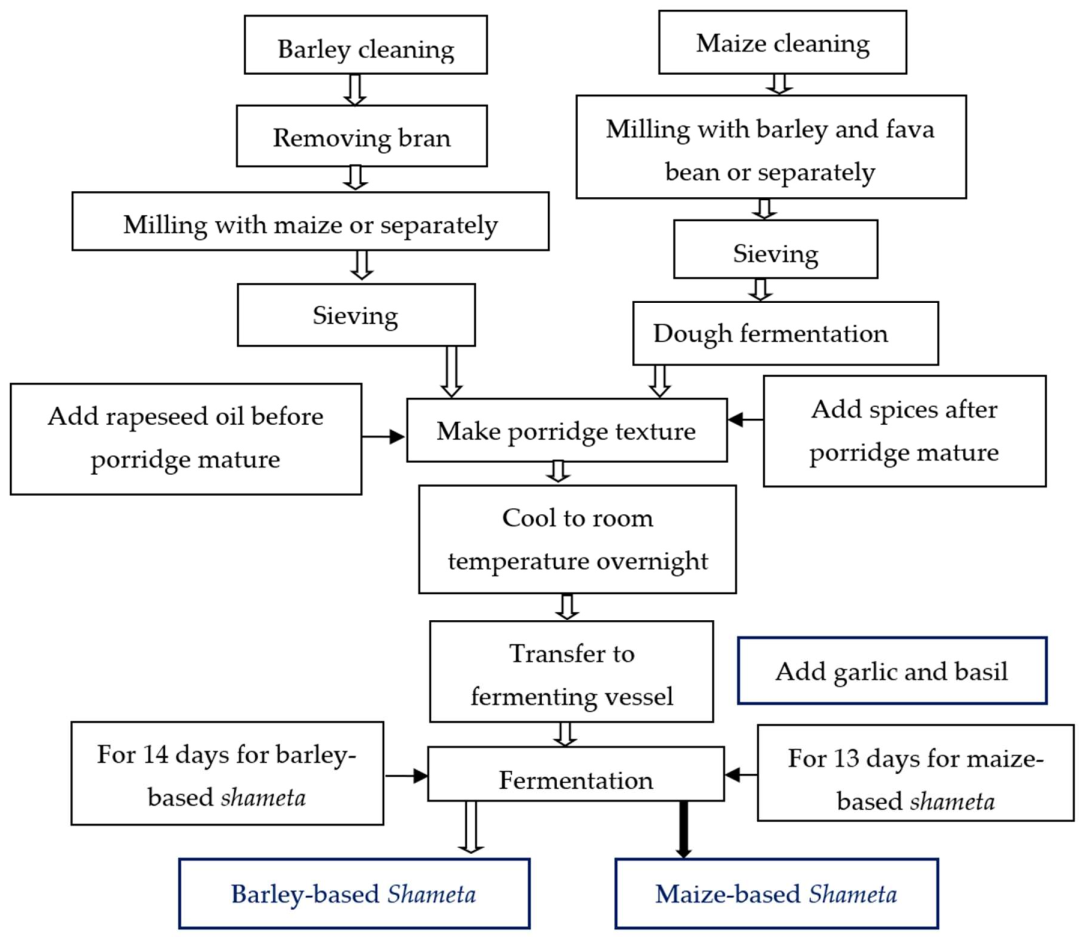
2.2.3. Laboratory Sample Preparation
2.3. Determination of pH and Titratable Acidity (TA)
2.4. Moisture Content Determination
2.5. Enumeration of Microbial Groups
2.5.1. Aerobic Mesophilic Bacteria (AMB) Counts
2.5.2. Enterobacteriaceae Counts
2.5.3. Total Coliform Counts
2.5.4. Aerobic Spore-Forming Bacteria Count (ASFB)
2.5.5. Lactic Acid Bacteria Counts
2.5.6. Staphylococci Counts
2.5.7. Yeast and Mold Counts
2.5.8. Detection of Salmonella spp.
2.6. Characterization of LAB
2.7. Cell Shape, Cell Arrangement and Motility Test
2.8. Gram Reaction
2.9. Catalase Test
2.10. Acid and Gas Production from Glucose
2.11. Test for Tolerance to Different Salt Concentrations
2.12. Test for Tolerance to Different pH Levels
Data Analysis
3. Results and Discussion
3.1. Physicochemical and Microbial Quality of Shameta
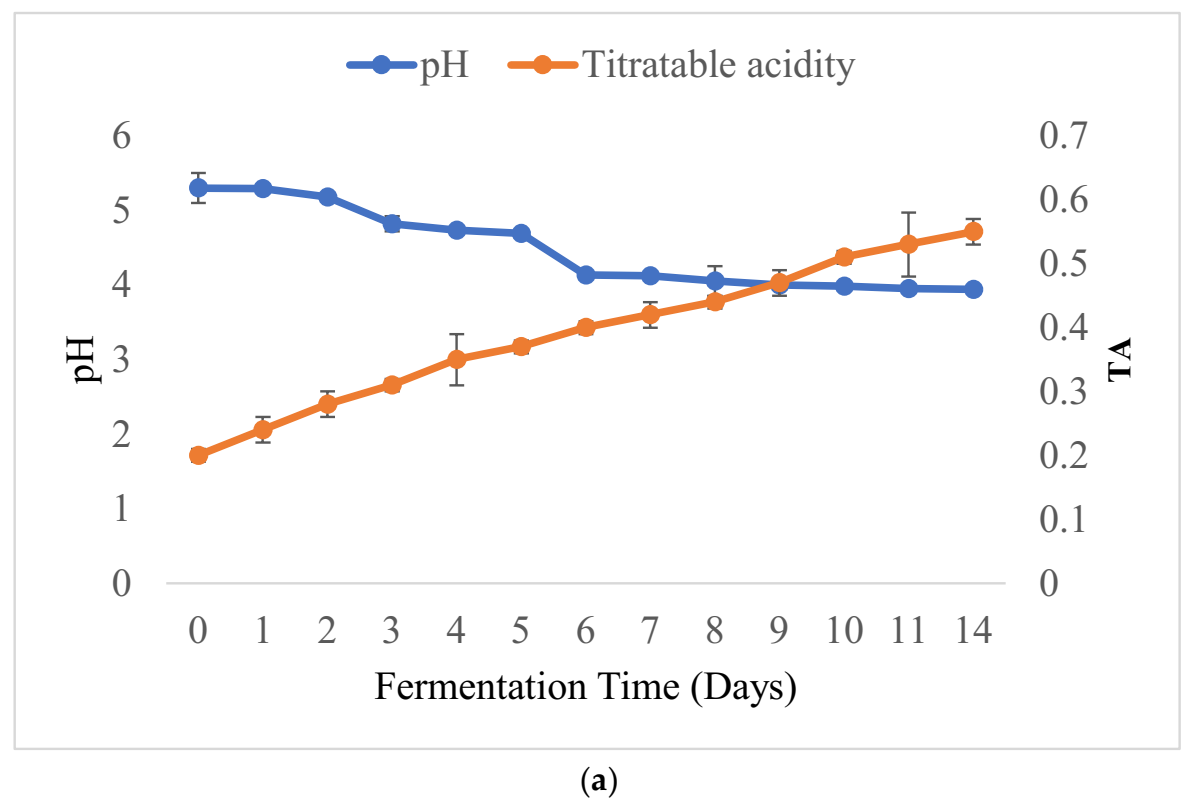
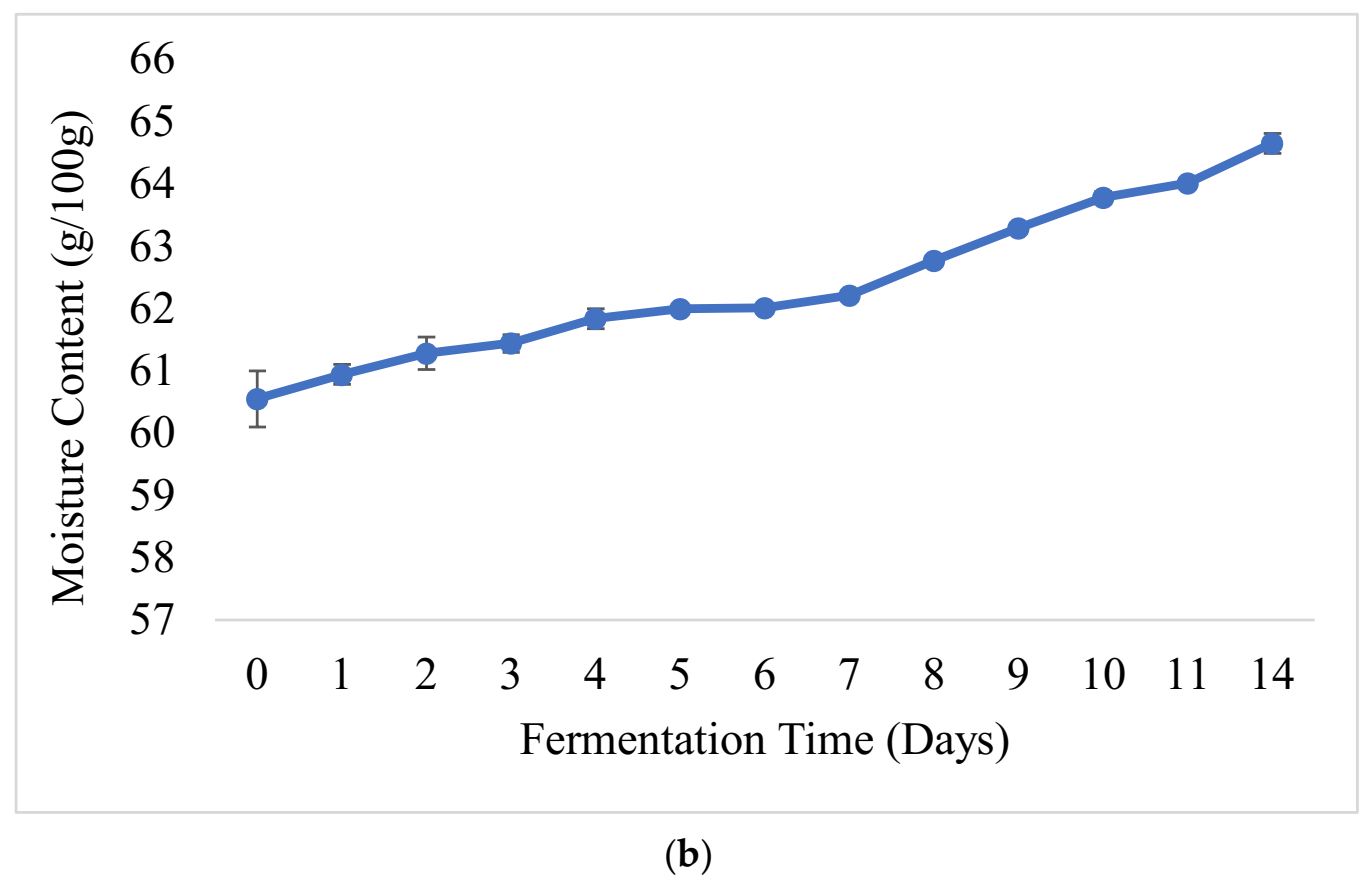
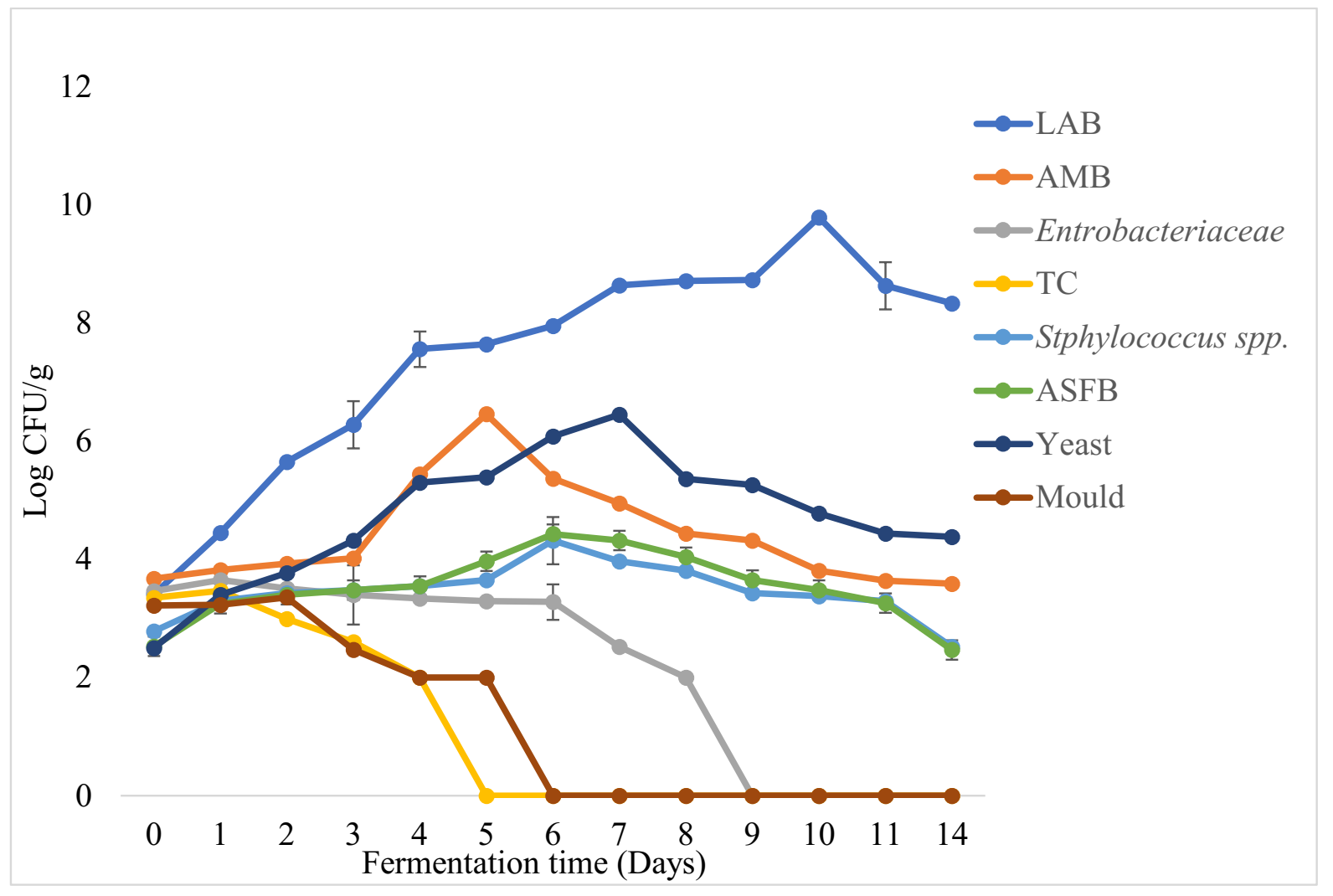
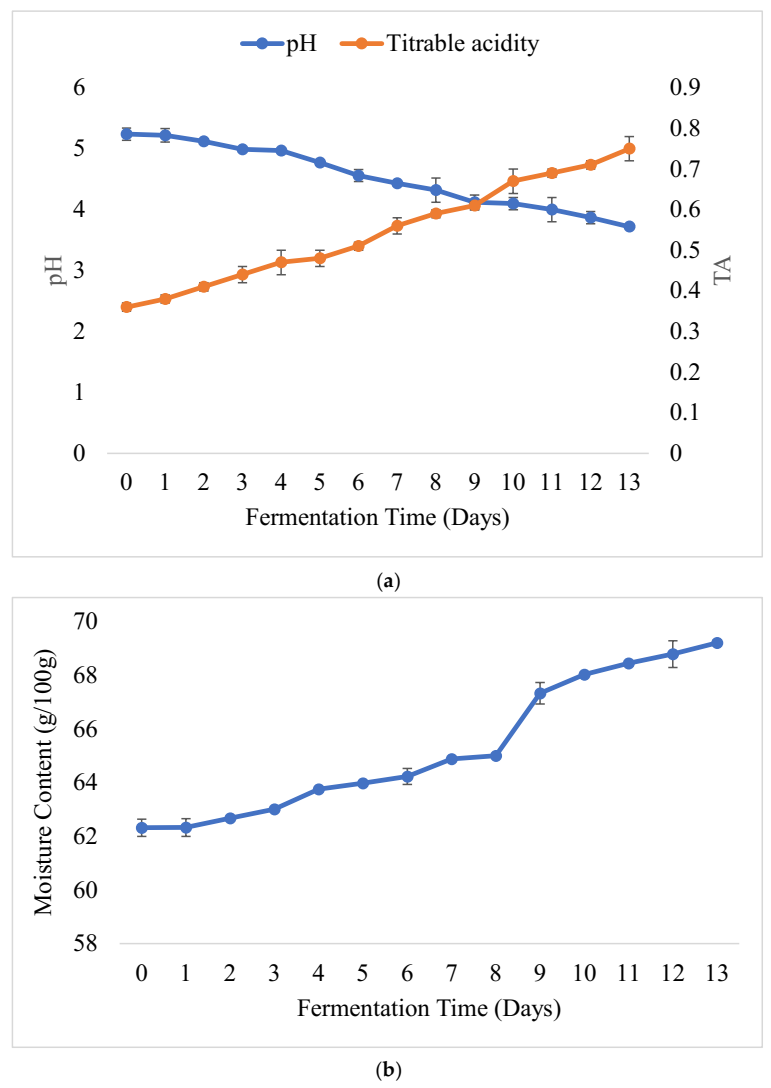
3.2. Physicochemical Changes and Microbial Dynamics during Barley-Based Shameta Fermentation
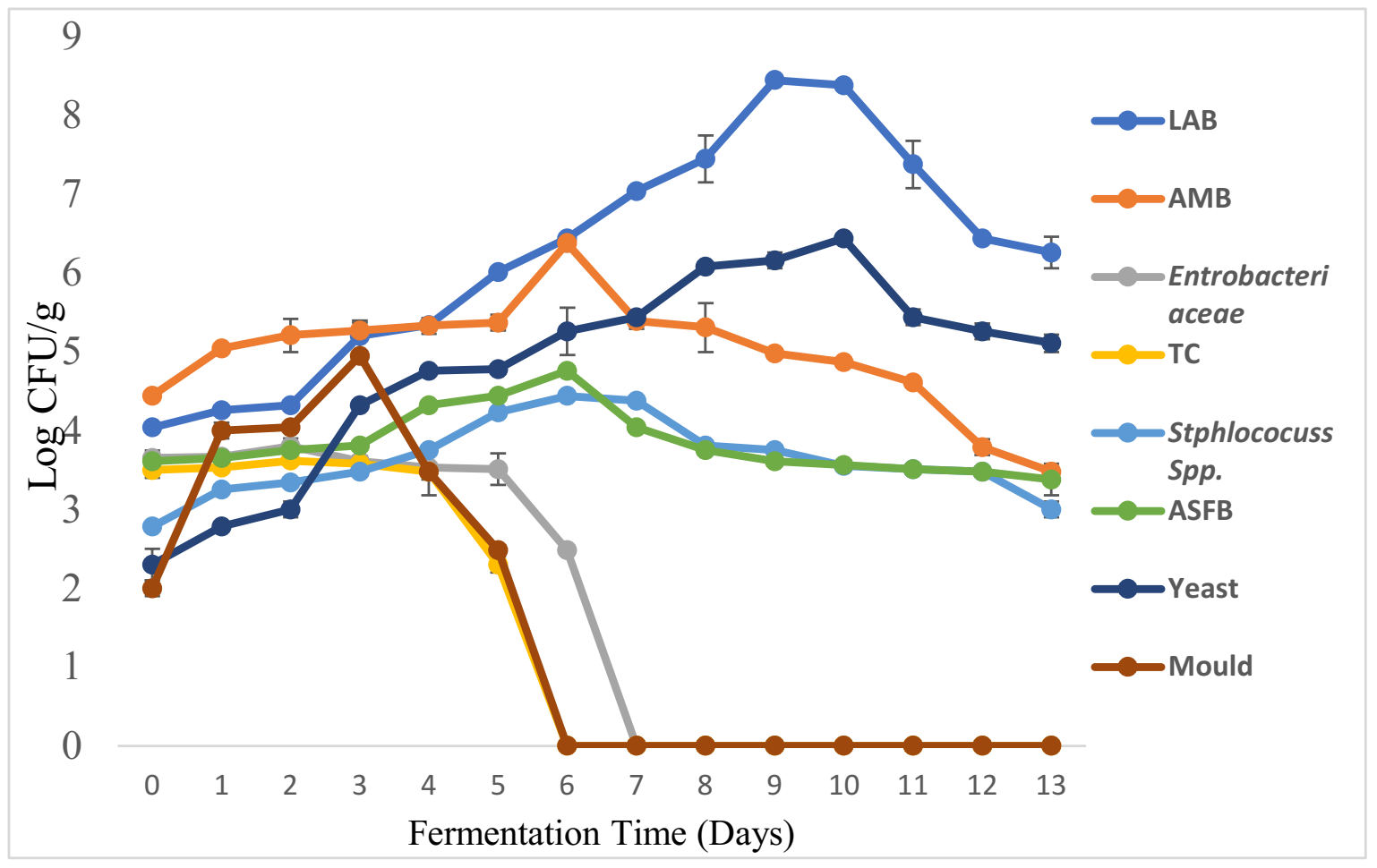
3.3. Physicochemical Changes and Microbial Dynamics during Maize-Based Shameta Fermentation

3.4. Identification of Dominant Lactic Acid Bacteria
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maria, L.M.; Dustin, H.; Sylvie, B.; Christopher, J.C.; Paul, D.C.; Benoit, F.; Michael, G.; Remco, K.; Gonca, P.; Anne, P.; et al. Health benefits of fermented foods: Microbiota and beyond. Curr. Opin. Biotechnol. 2017, 44, 94–102. [Google Scholar]
- Tou, E.H.; Mouquet-River, C.; Rochette, I.; Traoré, A.S.; Treche, S.; Guyot, J.P. Effect of different process combinations on the fermentation kinetics, microflora and energy density of ben-saalga, a fermented gruel from Burkina Faso. Food Chem. 2007, 100, 935–943. [Google Scholar] [CrossRef]
- Abegaz, K.; Beyene, F.; Thor, L.; Judith, A.N. Indigenous processing methods and raw materials of borde: An Ethiopian traditional fermented beverage. Afr. J. Food Sci. 2002, 7, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Binitu, B.; Zewdu, A.; Fekadu, H. Indigenous processing methods of cheka: A traditional fermented beverage in Southwestern Ethiopia. J. Food Process Technol. 2015, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Holzapfel, W.H. Appropriate starter culture technologies for small-scale fermentation in developing countries. Int. J. Food Microbiol. 2002, 75, 197–212. [Google Scholar] [CrossRef]
- Mukuma, M.M. Microbiological quality and safety of the Zambian fermented cereal beverage: Chibwantu. Ph.D. Thesis, University of the Free State, Bloemfontein, South Africa, 2014. [Google Scholar]
- Katina, K.; Poutanen, K. Nutritional aspects of cereal fermentation with lactic acid bacteria and yeast. In Handbook on Sourdough Biotechnology; Gobbetti, M., Gänzle, M., Eds.; Springer: New York, NY, USA, 2013; pp. 229–244. [Google Scholar]
- Ukwuru, M.U.; Muritala, A.; Ukpomwan, S. Ecology of traditional cereal fermentation. UPI J. Chem. Life Sci. 2018, 1, 22–36. [Google Scholar]
- Admassie, M. A Review on food fermentation and the biotechnology of lactic acid bacteria. World J. Food Sci. Technol. 2018, 2, 19–24. [Google Scholar] [CrossRef] [Green Version]
- Braide, W.; Azuwike, C.O.; Adeleye, S.A. The role of microorganisms in the production of some indigenous fermented foods in Nigeria. Int. J. Adv. Res. Biol. Sci. 2018, 5, 86–92. [Google Scholar]
- Giraffa, G. Studying the dynamics of microbial populations during food fermentation. FEMS Microbiol. Rev. 2004, 28, 251–260. [Google Scholar] [CrossRef] [Green Version]
- Nsofor, C.A.; Ume, S.C.; Uzor, B.C. Isolation and characterization of lactic acid bacteria from ogi sold in Elele. Niger. J. Biol. Food Sci. Res. 2014, 3, 19–22. [Google Scholar]
- Mobolaji, A.O.; Ifeoma, U.O.; Obadina, A.O.; Bankole, M.O.; Ayofemi, S.O.A. Microbiological assessment of maize ogi co-fermented with pigeon pea. Food Sci. Nutr. 2018, 6, 1238–1253. [Google Scholar]
- Bacha, K.; Tetemke, M.; Ashenafi, M. The microbial dynamics of “borde” fermentation: A traditional Ethiopian fermented beverage. SINET Ethiop. J. Sci. 1998, 21, 195–205. [Google Scholar] [CrossRef] [Green Version]
- Gebrelibanos, L. Microbiological and Physicochemical Study of Azo: A Traditional Fermented Condiment Prepared from Sorghum and Leaves of Endod (Phytolacca dodecandra) in Kafta Humera, Tigray Regional State. Master’s Thesis, Addis Ababa University, Addis Ababa, Ethiopia, 2015. [Google Scholar]
- Safaye, H.A. Isolation and Characterization of the Dominant Yeast in the Traditional Beverages of Ethiopia; Tella and Tej. Master’s Thesis, Addis Ababa University, Addis Ababa, Ethiopia, 2011. [Google Scholar]
- Getnet, B.; Minyamer, T.; Masresha, G. The microbial profile and contaminants of bubugn: An Ethiopian house hold fermented beverage. Int. J. Biotechnol. 2018, 7, 17–24. [Google Scholar]
- Bacha, K.; Jonsson, H.; Ashenafi, M. Microbial Dynamics during the fermentation of wakalim: A traditional Ethiopian fermented sausage. J. Food Qual. 2010, 33, 370–390. [Google Scholar] [CrossRef]
- Central Statistical Agency (CSA). Population and Housing Census in Ethiopia. International Household Survey Network. Home/Central Data Catalog/ETH_2007_PHC_V01_M. 2007. Available online: https://microdata.worldbank.org/index.php/catalog/2747 (accessed on 11 February 2022).
- Degefa, K.; Biru, G.; Abebe, G. Farming system characterization and analysis of East Wollega Zone, Oromia, Ethiopia. Int. J. Manag. Fuzzy Syst. 2020, 6, 14–28. [Google Scholar] [CrossRef]
- Abegaz, K. Isolation, characterization and identification of lactic acid bacteria involved in traditional fermentation of borde: An Ethiopian cereal-based beverage. Afr. J. Biotechnol. 2007, 6, 1469–1478. [Google Scholar]
- Antony, U.; Chandra, T. Microbial Population and biochemical changes in fermenting finger millet (Eleusine coracana). World J. Microbiol. Biotechnol. 1997, 13, 533–537. [Google Scholar] [CrossRef]
- Association of Official Agricultural Chemists (AOAC). Official Methods of Analysis of the AOAC, 18th ed.; AOAC International: Washington, DC, USA, 2000. [Google Scholar]
- Mugula, J.K.; Nnko, S.A.M.; Sorhaug, T. Changes in quality attributes during storage of togwa: A lactic acid fermented gruel. J. Food Saf. 2001, 21, 181–194. [Google Scholar] [CrossRef]
- Todd, M.S.; Elliot, T.R.; Catherine, W.D. Comparison of methods for determining coliform and Escherichia coli levels in apple cider. J. Food Prot. 1997, 4, 1302–1471. [Google Scholar]
- Soda, M.E.; Ahmed, N.; Omran, N.; Osman, G.; Morsi, A. Isolation, identification and selection of lactic acid bacteria cultures for cheese making. Emir. J. Agric. Sci. 2003, 15, 51–71. [Google Scholar] [CrossRef]
- Nemo, R.; Bacha, K. Microbial, physico-chemical and proximate analysis of selected Ethiopian traditional fermented beverage. LWT Food Sci. Technol. 2020, 131, 109713. [Google Scholar] [CrossRef]
- Idris, A.; Mehari, T.; Ashenafi, M. Some microbiological and biochemical studies on the fermentation of “awaze” and “datta”: A traditional Ethiopian condiment. Int. J. Food Sci. Nutr. 2001, 52, 5–14. [Google Scholar] [PubMed]
- Nikita, C.; Hemangi, D. Isolation, identification and characterization of lactic acid bacteria from dairy sludge sample. J. Environ. Res. Dev. 2012, 7, 234–244. [Google Scholar]
- Okoro, I.A.; Ojimelukwe, P.C.; Ekwenye, U.N.; Akaerue, B.; Atuonwu, A.C. Quality characteristics of indigenous fermented beverage; pito using Lactobacillus sake as a starter culture. Cont. J. Appl. Sci. 2011, 6, 15–20. [Google Scholar]
- Onyeagba, A.C. Laboratory Guide for Microbiology, 1st ed.; Crystal Publishers: Okigwe, Nigeria, 2004; pp. 75–94. [Google Scholar]
- Kimaryo, V.; Massawe, G.; Olasupo, N.; Holzapfel, W. The use of starter culture in the fermentation of cassava for the production of “kivunde”: A traditional Tanzanian food product. Int. J. Food Microbiol. 2000, 56, 179–190. [Google Scholar] [CrossRef]
- Seeley, H.W.; Vandemark, P.J. Microbes in Action—A Laboratory Manual of Microbiology; D.B. Taraporevala Sons and Company Pvt. Ltd.: Mumbai, India, 1970; Volume 39, pp. 85–86. [Google Scholar]
- Chowdhury, A.; Raju, K.K.; Kalurupalle, S.; Tharun, S. Both Sm-domain and C-terminal extension of Lsm1 are important for the RNA-binding Activity of the Lsm1-7-Pat1 Complex. RNA 2014, 20, 1465–1475. [Google Scholar] [CrossRef] [Green Version]
- Hasan, M.N.; Sultan, M.Z.; Mareum, M. Significance of fermented food in nutrition and food science. J. Sci. 2014, 6, 373–386. [Google Scholar] [CrossRef]
- Garbutt, J. Essentials of Food Microbiology; Hodder Headline Group: London, UK, 1997; pp. 116–170. [Google Scholar]
- Battcock, M.; Azam-Ali, S. Basic Principles of Fermentation. In Fermented Fruits and Vegetables: A Global Perspective; FAO Agricultural Services Bulletin No. 134; FAO: Rome, Italy, 1998; Chapter 2. [Google Scholar]
- Binitu, B.W.; Fekadu, H.G.; Zewdu, A.W. Nutritional and alcoholic contents of cheka: A traditional fermented beverage in Southwestern Ethiopia. Food Sci. Nutr. 2018, 6, 2466–2472. [Google Scholar] [CrossRef]
- Ashenafi, M.; Mehari, T. Some microbiological and nutritional properties of “borde” and “shamita, traditional Ethiopian fermented beverages. Ethiop. J. Health Dev. 1995, 9, 105–110. [Google Scholar]
- Lechiancole, T.; Ricciardi, A.; Parente, E. Optimization of media and fermentation conditions for the growth of Lactobacillus sakei. Ann. Microbiol. 2002, 52, 257–274. [Google Scholar]
- Liptáková, D.; Matejčeková, Z.; Valík, Ľ. Lactic acid bacteria and fermentation of cereals and Pseudocereals. In Fermentation Processes; InTech: London, UK, 2017; pp. 224–242. [Google Scholar]
- Asfaw, D. Review on use of bioactive compounds in some spices in food preservation. Food Sci. Qual. Manag. 2019, 87, 6–13. [Google Scholar]
- Chibuike, A.O.; Ify, D.A.; Ejike, M.O.; Awache, I.; James, L.A. Study on the bacteriological quality of fura sold in Wukari, North-East Nigeria. Food Microbiol. 2018, 2, 24–29. [Google Scholar]
- Ogodo, A.C.; Ugbogu, O.C.; Agwaranze, D.I.; Ihiabe, F.U. Some studies on the bacteriological quality of sorghum-based commercially prepared fermented Ogi (Akamu) in Wukari, Nigeria. Am. J. Food Sci. Nutr. 2017, 4, 48–51. [Google Scholar]
- Centre for Food Safety (CFS). Microbiological Guidelines for Food: Ready-to-Eat food in General and Specific Food Items; Centre for Food Safety, Food and Environmental Hygiene Department: Hong Kong, China, 2014. Available online: microbio-guide2014-cvr_E(cfs.gov.hk) (accessed on 20 January 2022).
- Cox, L.J.; Keller, N.; Vanschothorst, M. The use and misuse of quantitative determinations of Enterobacteriaceae in Food Microbiology. J. Appl. Bacteriol. Symp. Suppl. 1988, 17, 237S–249S. [Google Scholar] [CrossRef]
- Samuel, C.T. Street food safety, types and microbiological quality in Ethiopia: A Critical review. Am. J. Appl. Sci. Res. 2020, 6, 67–71. [Google Scholar]
- Martin, N.H.; Trmcic, A.; Hsieh, T.; Boor, K.J.; Wiedmann, M. The evolving role of coliforms as indicators of unhygienic processing conditions in dairy foods. Front. Microbial. 2016, 7, 1549. [Google Scholar] [CrossRef]
- Food Standards Australia New Zealand (FSANZ). Compendium of Microbiological Criteria for Food; Food Standards Australia New Zealand, the Terrace: Wellington, New Zealand, 2016; pp. 1–51.
- Reed, G.H. Foodborne illness (part 8): Escherichia coli. Dairy Food Environ. San. 1994, 14, 329–330. [Google Scholar]
- Ray, B. Fundamental Food Microbiology, 3rd ed.; CRC Press: New York, NY, USA, 2004; pp. 359–390. [Google Scholar]
- Ogodo, A.C.; Agwaranze, D.I.; Nwaneri, C.B.; Okoronkwo, U.C.; Ekeleme, U.G. Activity of leave and stem bark cuttings of Ocimum gratissimum extracts on foodborne pathogens. AASCIT J. Biosci. 2017, 3, 5–11. [Google Scholar]
- Tewari, A.; Abdullah, S. Bacillus cereus food poisoning: International and Indian perspective. J. Food Sci. Technol. 2015, 52, 2500–2511. [Google Scholar] [CrossRef] [Green Version]
- Lücking, G.; Stoeckel, M.; Atamer, Z.; Hinrichs, J. Characterization of aerobic spore-forming bacteria associated with industrial dairy processing environments and product spoilage. Int. J. Food Microbiol. 2013, 166, 270–279. [Google Scholar] [CrossRef]
- Health Protection Agency (HPA). Guidelines for Assessing the Microbiological Safety of Ready-to-Eat Foods; Health Protection Agency: London, UK, 2009; pp. 8–10.
- Food Safety Authority of Ireland (FSAI). Guidelines for the Interpretation of Results of Microbiological Testing of Ready-to-Eat Foods Placed on the Market; Revision 4; Guidance Note; European Comission: Brussels, Belgium, 2020. [Google Scholar]
- Rawat, S. Food Spoilage: Microorganisms and their prevention. Asian J. Plant Sci. Res. 2015, 5, 47–56. [Google Scholar]
- Ogbulie, J.N. Production of Tempeh from African Yam Bean (Sphenostylis stenocarpa Harms). Master’s Thesis, University of Port Harcourt, Port Harcourt, Nigeria, 1991; p. 33. [Google Scholar]
- Olumide, A.; Odeyemi, O.; Oluwaseun, A.; Mariyana, S.; Deyan, S. Understanding spoilage microbial community and spoilage mechanisms in foods of animal origin. Compr. Rev. Food Sci. Food Saf. 2020, 19, 311–331. [Google Scholar]
- Mohammed, S.I.; Steenson, L.R.; Kirleis, A.W. Isolation and characterization of microorganisms associated with the traditional sorghum fermentation for production of Sudanese kisra. Appl. Environ. Microbiol. 1991, 57, 2529–2533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogbonnaya, N.; Bernice, C.C. Studies on Akamu: A Traditional fermented maize food. Rev. Chil. Nutr. 2012, 39, 180–184. [Google Scholar]
- Minton, P.E. Handbook of Evaporation Technology; Noyes Publications: West Wood, NJ, USA, 1986. [Google Scholar]
- Berggren, S.; Hedren, E.; Edman, K. Water Holding Capacity and Viscosity of Ingredients from Oats. The Effect of Beta-Glucan and Starch Content, Particle Size, pH and Temperature; Linnaeus University: Växjö, Sweden, 2017. [Google Scholar]
- Halm, M.; Lillie, A.; Sorensen, A.K.; Jakobsen, M. Microbiological and aromatic characteristics of fermented maize doughs for kenkey production in Ghana. Int. J. Food Microbiol. 1993, 19, 135–143. [Google Scholar] [CrossRef]
- Alonso-Calleja, C.; Martínez-Fernández, B.; Prieto, M.; Capita-González, R. Microbiological quality of vacuum-packed retail ostrich meat in Spain. Food Microbiol. 2004, 21, 241–246. [Google Scholar] [CrossRef]
- Almeida, E.G.; Rachid, C.C.; Schwan, R.F. Microbial population present in fermented beverage ‘cauim’ produced by Brazilian Amerindians. Int. J. Food Microbiol. 2007, 120, 146–151. [Google Scholar] [CrossRef]
- Steinkraus, K.H. Hand Book of Indigenous Fermented Foods; Revised and Expanded; Marcel Dekker: New York, NY, USA, 1996. [Google Scholar]
- Fredlund, E.; Druvefors, U.Ä.; Olstorpe, M.N.; Passoth, V.; Schnürer, J. Influence of ethyl acetate production and ploidy on the anti-mould activity of Pichia anomala. FEMS Microbiol. Lett. 2004, 238, 133–137. [Google Scholar]
- Chelule, P.; Mokoena, M.; Gqaleni, N. Advantages of Traditional Lactic Acid Bacteria Fermentation of Food in Africa; Current Research, Technology and Education Topics in Applied Microbiology and Biotechnology; Mendez–Vilas, A., Ed.; FORMATEX Microbiology Series; Formatex Research Center: Badajoz, Spain, 2010; Volume 2. [Google Scholar]
- Anteneh, T.; Tetemke, M.; Mogessie, A. Antagonism of lactic acid bacteria against food borne pathogens during fermentation and storage of borde and shameta: Traditional Ethiopian fermented beverages. Int. Food Res. J. 2011, 18, 1189–1194. [Google Scholar]
- Achi, O.K.; Ukwuru, M. Cereal-Based fermented foods of Africa as functional foods. Int. J Microbiol. Appl. 2015, 2, 71–83. [Google Scholar]
- Ogodo, A.C.; Ugbogu, O.C.; Ugbogu, A.E.; Ezeonu, C.S. Production of mixed fruit (pawpaw, banana and watermelon) wine using Saccharomyces cerevisiae isolated from palm wine. Springer Plus 2015, 4, 683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zambrano, M.; Dutta, B.; Mercer, D.G.; MacLean, H.L.; Touchie, M.F. Assessment of moisture content measurement methods of dried food products in small-scale operations in developing countries: A review. Trends Food Sci. Technol. 2019, 88, 484–496. [Google Scholar] [CrossRef]
- Nout, M.J.R.; Rombouts, F.M.; Hautfast, G.J. Accelerated natural lactic fermentation of infant food formulations. Food Nutr. Bull. 1989, 11, 65–73. [Google Scholar] [CrossRef]
- Odunfa, S.A. African fermented foods. In Microbiology of Fermented Foods; Wood, B.J., Ed.; Elsevier Applied Science Publishers: London, UK; New York, NY, USA, 1985; Volume 2, pp. 155–199. [Google Scholar]
- Jay, J.M.; Loessner, M.J.; Golden, D.A. Modern Food Microbiology, 7th ed.; Springer Science and Business Media Inc.: Los Angeles, CA, USA, 2005; pp. 125–143. [Google Scholar]
- Jespersen, I.; Halm, M.; Kpodo, K.; Jakobsen, M. Significance of yeasts and moulds occurring in maize dough fermentation for kenkey production. Int. J Food Microbiol. 1994, 24, 239–248. [Google Scholar] [CrossRef]
- Omemu, A.M.; Oyewole, O.B.; Bankole, M.O. Significance of yeasts in the fermentation of maize for ogi production. Food Microbiol. 2007, 24, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Omemu, A.M. Fermentation dynamics during production of ogi: A Nigerian fermented cereal porridge. Rep. Opin. 2011, 3, 8–17. [Google Scholar]
- Nout, M.J.R.; Aidoo, K.E. Asian Fungal Fermented Food; Osiewacz, H.D., Ed.; The Mycota X Industrial Applications; Springer: Berlin/Heidelberg, Germany, 2002; pp. 23–47. [Google Scholar]
- Thapa, S.; Tamang, J.P. Product characterization of kodo ko jaanr: A fermented finger millet beverage of the Himalayas. Food Microbiol. 2004, 21, 617–622. [Google Scholar] [CrossRef]
- Petrova, P.; Petrov, K.; Stoyancheva, G. Starch-modifying enzymes of lactic acid bacteria—Structures, Properties and Applications. Starch-Stärke 2013, 65, 34–47. [Google Scholar] [CrossRef]
- Owusu-Kwarteng, J.; Tano-Debrah, K.; Glover, R.L.K.; Akabanda, F. Process characteristics and microbiology of fura produced in Ghana. J. Nat. Sci. 2010, 8, 41–51. [Google Scholar]
- Adegoke, G.O.; Babaola, A.K. Characteristics of microorganisms of importance in the fermentation of fufu and ogi: The two Nigerian fermented foods. J. Appl. Bacteriol. 1988, 65, 449–453. [Google Scholar] [CrossRef]
- Nche, P.F.; Nout, M.J.R.; Rombouts, F.M. Effect of cowpea supplementation on the quality of kenkey: A traditional Ghanaian fermented food. J. Cereal Sci. 1994, 19, 191–197. [Google Scholar] [CrossRef]
- Lukjancenko, O.; Ussery, D.W.; Wassenaar, T.M. Comparative genomics of Bifidobacterium, Lactobacillus and related probiotic genera. Microbiol. Ecol. 2012, 63, 651–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franz, C.M.A.P.; Stiles, M.E.; Schleifer, K.H.; Holzapfel, W.H. Enterococci in foods: A Conundrum for food safety. Int. J. Food Microbiol. 2003, 88, 105–122. [Google Scholar] [CrossRef]
- Olatunde, O.O.; Obadina, A.O.; Omemu, A.M.; Oyewole, O.B.; Olugbile, A.; Olukomaiya, O.O. Screening and molecular identification of potential probiotic lactic acid bacteria in effluents generated during ogi production. Ann. Microbiol. 2018, 68, 433–443. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Sources (Districts) | Samples Code | Major Ingredients | Approximate Premix Added | |
|---|---|---|---|---|
| Types of Spices and Herbs Added | Rapeseed Oil (%) | |||
| Sibu Sire | BMWS | Barley (86%) + Maize (5%) + wheat (5%) | F + BCU + Gr + R (0.5%) | 3.5 |
| BMS | Barley (89%) + Maize (5%) | BC + BC2 + R (1.5%) | 4.5 | |
| MS | Maize (95%) | F + WC + Gr + R (0.5%) | 4.5 | |
| Guto Gida | MFG | Maize (86.5%) + Faba bean (5%) | F + BC + BCU + Gr + B (1%) | 7.5 |
| MG | Maize (91%) | G + BC + Gr + B (1%) | 8 | |
| MBFG | Maize (81%) + Barley (5%) + Faba bean (5%) | BC + WC + Gr + R (1.5%) | 7.5 | |
| Jimma Arjo | BMJ | Barley (87%) + Maize (5%) | F + Gr (0.5%) | 7.5 |
| BJ | Barley (96%) | F + Gr (0.5%) | 3.5 | |
| Wayu Tuka | MW | Maize (91%) | BCU + R (1.5%) | 7.5 |
| Coded Samples | pH | Titratable Acidity (%) |
|---|---|---|
| BMWS | 4.1 ± 0.1 d | 0.78 + 0.01 c |
| BMS | 4.4 + 0.01 a | 0.57 + 0.01 f |
| MS | 4.1 + 0.01 d | 0.74 + 0.02 d |
| MFG | 3.9 + 0.00 e | 0.82 + 0.01 b |
| MG | 3.6 + 0.02 f | 1.1 + 0.03 a |
| MBFG | 3.9 + 0.02 e | 0.82 + 0.00 b |
| BMJ | 4.2 + 0.01 b | 0.61 + 0.00 e |
| BJ | 3.9 + 0.00 e | 0.81 + 0.01 b |
| MW | 4.2 + 0.01 b | 0.76 + 0.01 c,d |
| CV | 0.56 | 1.14 |
| MSD | 0.1 | 0.03 |
| CODE * | LAB | AMB | Entrobacteriaceae | TC | Staphylococcus spp. | ASFB | Yeast | Mould |
|---|---|---|---|---|---|---|---|---|
| BMWS | 7.54 ± 0.02 c | 4.40 ± 0.1 b | 3.69 ± 0.00 a | 3.36 ± 0.02 a | 3.22 ± 0.00 c | 2.47 ± 0.01 b | 4.32 ± 0.02 c,d | 3.26 ± 0.02 c |
| BMS | 5.88 ± 0.01 h | 4.18 ± 0.01 c | 3.38 ± 0.01 b | 3.27 ± 0.01 a,b | <2 | <2 | 5.26 ± 0.01 a | 3.32 ± 0.02 b |
| MS | 6.74 + 0.02 f | 3.50 + 0.1 d | 2.74 + 0.02 d | <2 | 3.22 + 0.01 b | <2 | 4.18 + 0.01 d | 2.26 + 0.02 f |
| MFG | 7.74 ± 0.02 b | 4.58 ± 0.01 a | 3.38 ± 0.01 b | 3.28 ± 0.01 a,b | 3.28 ± 0.01 a | <2 | 3.5 ± 0.10 e | 2.11 ± 0.01 g |
| MG | 6.74 ± 0.02 f | 3.32 ± 0.01 f | 3.32 ± 0.01 b,c | 3.22 ± 0.00 b | <2 | <2 | 4.78 ± 0.02 b | <2 |
| MBFG | 6.81 ± 0.01 e,f | 3.36 ± 0.02 e,f | 2.65 ± 0.01 d | <2 | 3.25 ± 0.01 a,b | <2 | 4.44 ± 0.02 d | <2 |
| BMJ | 6.88 ± 0.01 e | 3.37 ± 0.00 e,f | 3.33 ± 0.01 b,c | 2.48 ± 0.01 c | 2.47 ± 0.02 e | 2.60 ± 0.1 a | 4.26 ± 0.03 c,d | 2.47 ± 0.01 e |
| BJ | 6.28 ± 0.01 g | 3.18 ± 0.01 g | 3.28 ± 0.01 c | 2.50 ± 0.1 c | 2.52 ± 0.02 d | 2.5 ± 0.1 a,b | 3.65 ± 0.01 e | 3.67 ± 0.01 a |
| MW | 7.40 ± 0.01 d | 3.32 ± 0.01 f | 2.70 ± 0.1 d | <2 | 2.47 ± 0.02 e | 2.51 ± 0.00 a,b | 3.51 ± 0.01 e | 2.62 ± 0.01 d |
| Lab1 | 8.33 ± 0.01 a | 3.59 ± 0.01 d | <2 | <2 | <2 | <2 | 4.38 ± 0.02 c | <2 |
| Lab2 | 6.26 ± 0.1 g | 3.48 ± 0.02 d,e | <2 | <2 | <2 | <2 | 5.11 ± 0.11 a | <2 |
| CV | 1.20 | 1.01 | 1.16 | 1.36 | 0.50 | 2.79 | 1.48 | 0.58 |
| MSD | 0.13 | 0.10 | 0.10 | 0.10 | 0.04 | 0.13 | 0.19 | 0.04 |
| Group of LAB | |||
|---|---|---|---|
| I (n = 128) | II (n = 27) | III (n = 16) | |
| Characteristics | |||
| Microscopic morphology | Rod | Cocci | Cocci in tetrads |
| Gram reaction | + | + | + |
| Catalase Test | - | - | - |
| Oxidation fermentation | - | - | - |
| Motility test | - | - | - |
| KOH Test | - | - | - |
| CO2 from Glucose | - | - | - |
| Growth at temperature | |||
| 10 °C | - | - | - |
| 15 °C | −/+ | - | - |
| 32 °C | + | + | + |
| 37 °C | + | + | + |
| 45 °C | +/− | + | + |
| Tolerance to NaCl (%) | |||
| 2.5 | +/− | +/− | +/− |
| 4 | + | + | + |
| 6.5 | +/− | + | + |
| pH tolerance | |||
| pH 4.4 | +/− | + | + |
| pH 9.6 | - | + | - |
| Possible Identity of the Isolates | Lactobacillus | Entrococcus | Pediococcus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kitessa, D.A.; Bacha, K.; Tola, Y.B.; Murimi, M.; Gershe, S.; Guta, M. Microbial Quality and Growth Dynamics in Shameta: A Traditional Ethiopian Cereal-Based Fermented Porridge. Fermentation 2022, 8, 124. https://doi.org/10.3390/fermentation8030124
Kitessa DA, Bacha K, Tola YB, Murimi M, Gershe S, Guta M. Microbial Quality and Growth Dynamics in Shameta: A Traditional Ethiopian Cereal-Based Fermented Porridge. Fermentation. 2022; 8(3):124. https://doi.org/10.3390/fermentation8030124
Chicago/Turabian StyleKitessa, Daniel A., Ketema Bacha, Yetenayet B. Tola, Mary Murimi, Soressa Gershe, and Meseret Guta. 2022. "Microbial Quality and Growth Dynamics in Shameta: A Traditional Ethiopian Cereal-Based Fermented Porridge" Fermentation 8, no. 3: 124. https://doi.org/10.3390/fermentation8030124