
Valorization of a Pyrolytic Aqueous Condensate and Its Main Components for L-Malic Acid Production with Aspergillus oryzae DSM 1863


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism
2.2. Media and Cultivation Conditions
2.2.1. Preparation of the Spore Solution
2.2.2. Preculture Cultivations for Biomass Formation
2.2.3. Precultures for Main Culture Inoculation
2.2.4. Main Culture for Malic Acid Production in Shake Flasks
2.2.5. Bioreactor Cultivations for Malate Production
2.3. Formation and Detoxification of the PAC
2.4. Analytics
2.4.1. Quantification of the Fungal Cell Dry Weight
2.4.2. HPLC Analysis of Substrate Consumption and Malate Production
3. Results
3.1. Utilization of Acetol as Substrate for Biomass Formation of A. oryzae
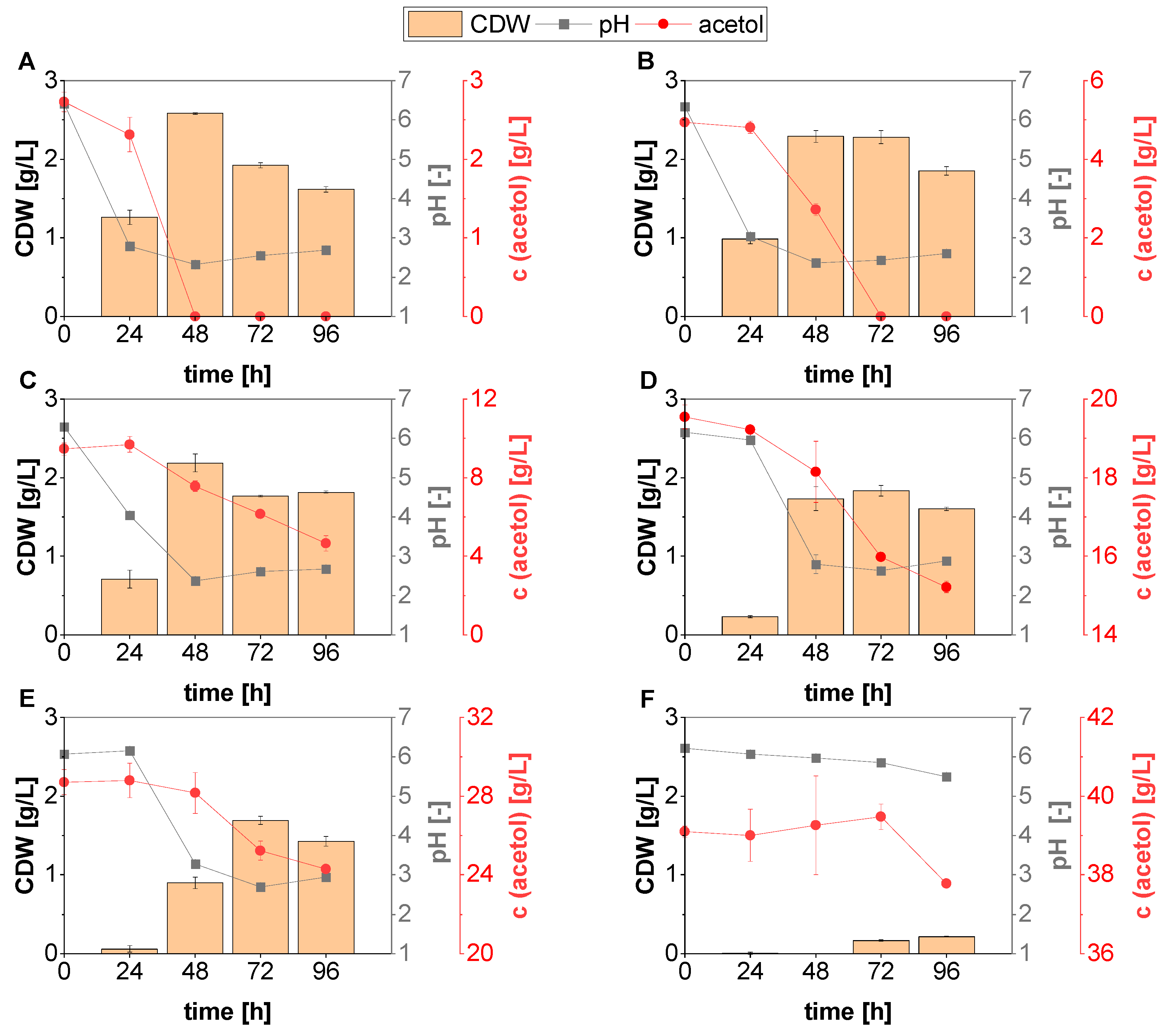
3.2. Acetate-Acetol Mixtures as Substrates for Fungal Growth
3.3. L-Malic Acid Production with Acetate-Acetol Mixtures
3.4. PAC as Sole Substrate for L-Malic Acid Formation
3.4.1. General Suitability of PAC for Fungal Acid Formation
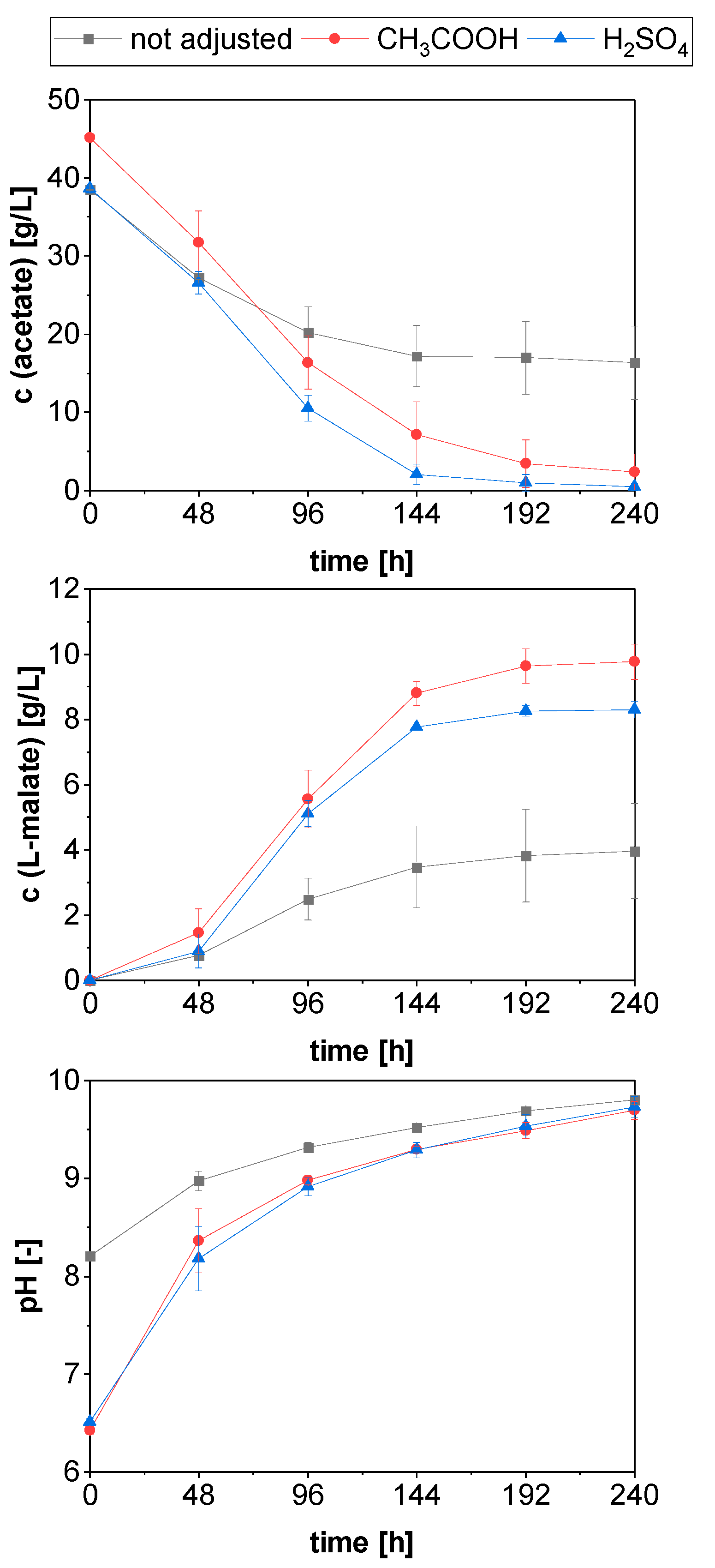
3.4.2. Role of the Initial Medium pH on L-Malic Acid Production from PAC
3.4.3. Malic Acid Production in Bioreactor Cultivations
4. Discussion
4.1. Acetol Utilization in A. oryzae Preculture
4.2. Malic Acid Production on Artificial and Real PAC
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grignard, R. New Quaternary Ammonium Salts of Malic Acid and Their Application in Cosmetics. U.S. Patent 3,954,846, 4 May 1976. [Google Scholar]
- Gore, V.; Gadkar, M.; Pokharkar, K. Novel Process to Prepare Almotriptan. U.S. Patent 2010/0292290 A1, 18 November 2010. [Google Scholar]
- Löffler, M.; Morschhäuser, R. Acid Cosmetic, Dermatological and Pharmaceutical Agents. U.S. Patent 7,025,973, 11 April 2006. [Google Scholar]
- Wu, X.; Chen, Z.; Li, Y.; Yu, Q.; Lu, Y.; Zhu, Q.; Li, Y.; An, D.; Qi, J.; Wu, W. Improving dermal delivery of hydrophilic macromolecules by biocompatible ionic liquid based on choline and malic acid. Int. J. Pharm. 2019, 558, 380–387. [Google Scholar] [CrossRef]
- El-Zaiat, H.M.; Kholif, A.E.; Mohamed, D.A.; Matloup, O.H.; Anele, U.Y.; Sallam, S.M.A. Enhancing lactational performance of Holstein dairy cows under commercial production: Malic acid as an option. J. Sci. Food Agric. 2019, 99, 885–892. [Google Scholar] [CrossRef] [PubMed]
- Ren, F.; Yang, W.; Hu, J.; Huang, P.; Jiao, X.-A.; Huang, J. Feeding Malic Acid to Chickens at Slaughter Age Improves Microbial Safety with Regard to Campylobacter. Animals 2021, 11, 1999. [Google Scholar] [CrossRef]
- Nguyen, D.H.; Seok, W.J.; Kim, I.H. Organic acids mixture as a dietary additive for pigs—A review. Animals 2020, 10, 952. [Google Scholar] [CrossRef] [PubMed]
- Kajiyama, T.; Kobayashi, H.; Taguchi, T.; Kataoka, K.; Tanaka, J. Improved synthesis with high yield and increased molecular weight of poly (α, β-malic acid) by direct polycondensation. Biomacromolecules 2004, 5, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Feng, G.; Shen, F.H.; Balian, G.; Laurencin, C.T.; Li, X. Novel biodegradable poly (1, 8-octanediol malate) for annulus fibrosus regeneration. Macromol. Biosci. 2007, 7, 1217–1224. [Google Scholar] [CrossRef]
- Werpy, T.; Petersen, G. Top Value Added Chemicals from Biomass: Volume I—Results of Screening for Potential Candidates from Sugars and Synthesis Gas; National Renewable Energy Lab: Golden, CO, USA, 2004.
- Kövilein, A.; Kubisch, C.; Cai, L.; Ochsenreither, K. Malic acid production from renewables: A review. J. Chem. Technol. Biotechnol. 2020, 95, 513–526. [Google Scholar] [CrossRef] [Green Version]
- Presečki, A.V.; Zelić, B.; Vasić-Rački, Đ. Comparison of the L-malic acid production by isolated fumarase and fumarase in permeabilized baker’s yeast cells. Enzyme Microb. Technol. 2007, 41, 605–612. [Google Scholar] [CrossRef]
- Naude, A.; Nicol, W. Malic acid production through the whole-cell hydration of fumaric acid with immobilised Rhizopus oryzae. Biochem. Eng. J. 2018, 137, 152–161. [Google Scholar] [CrossRef]
- Pandurić, N.; Šalić, A.; Zelić, B. Fully integrated biotransformation of fumaric acid by permeabilized baker’s yeast cells with in situ separation of L-malic acid using ultrafiltration, acidification and electrodialysis. Biochem. Eng. J. 2017, 125, 221–229. [Google Scholar] [CrossRef]
- Bressler, E.; Pines, O.; Goldberg, I.; Braun, S. Conversion of fumaric acid to l-malic by sol-gel immobilized Saccharomyces cerevisiae in a supported liquid membrane bioreactor. Biotechnol. Progr. 2002, 18, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Zhou, H.; Zhang, S.; Gu, H.; Yang, Q.; Zhang, W.; Dong, W.; Ma, J.; Fang, Y.; Jiang, M. Current advance in biological production of malic acid using wild type and metabolic engineered strains. Bioresour. Technol. 2018, 258, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, Y.; Yang, Y.; Zhang, H.; Wang, H.; Wu, Y.; Zhang, M.; Sun, T.; Cheng, J.; Wu, X. High levels of malic acid production by the bioconversion of corn straw hydrolyte using an isolated Rhizopus delemar strain. Biotechnol. Bioprocess Eng. 2014, 19, 478–492. [Google Scholar] [CrossRef]
- Zambanini, T.; Kleineberg, W.; Sarikaya, E.; Buescher, J.M.; Meurer, G.; Wierckx, N.; Blank, L.M. Enhanced malic acid production from glycerol with high-cell density Ustilago trichophora TZ1 cultivations. Biotechnol. Biofuels 2016, 9, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zambanini, T.; Sarikaya, E.; Kleineberg, W.; Buescher, J.M.; Meurer, G.; Wierckx, N.; Blank, L.M. Efficient malic acid production from glycerol with Ustilago trichophora TZ1. Biotechnol. Biofuels 2016, 9, 67. [Google Scholar] [CrossRef] [Green Version]
- Battat, E.; Peleg, Y.; Bercovitz, A.; Rokem, J.S.; Goldberg, I. Optimization of L-malic acid production by Aspergillus flavus in a stirred fermentor. Biotechnol. Bioeng. 1991, 37, 1108–1116. [Google Scholar] [CrossRef]
- Knuf, C.; Nookaew, I.; Brown, S.H.; McCulloch, M.; Berry, A.; Nielsen, J. Investigation of malic acid production in Aspergillus oryzae under nitrogen starvation conditions. Appl. Environ. Microbiol. 2013, 79, 6050–6058. [Google Scholar] [CrossRef] [Green Version]
- Ochsenreither, K.; Fischer, C.; Neumann, A.; Syldatk, C. Process characterization and influence of alternative carbon sources and carbon-to-nitrogen ratio on organic acid production by Aspergillus oryzae DSM1863. Appl. Microbiol. Biotechnol. 2014, 98, 5449–5460. [Google Scholar] [CrossRef] [PubMed]
- West, T.P. Malic acid production from thin stillage by Aspergillus species. Biotechnol. Lett 2011, 33, 2463–2467. [Google Scholar] [CrossRef]
- Iyyappan, J.; Bharathiraja, B.; Baskar, G.; Kamalanaban, E. Process optimization and kinetic analysis of malic acid production from crude glycerol using Aspergillus niger. Bioresour. Technol. 2019, 281, 18–25. [Google Scholar] [CrossRef]
- Dörsam, S.; Fesseler, J.; Gorte, O.; Hahn, T.; Zibek, S.; Syldatk, C.; Ochsenreither, K. Sustainable carbon sources for microbial organic acid production with filamentous fungi. Biotechnol. Biofuels 2017, 10, 242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oswald, F.; Dörsam, S.; Veith, N.; Zwick, M.; Neumann, A.; Ochsenreither, K.; Syldatk, C. Sequential mixed cultures: From syngas to malic acid. Front. Microbiol. 2016, 7, 891. [Google Scholar] [CrossRef] [PubMed]
- Pfitzer, C.; Dahmen, N.; Troger, N.; Weirich, F.; Sauer, J.; Gunther, A.; Muller-Hagedorn, M. Fast pyrolysis of wheat straw in the bioliq pilot plant. Energy Fuels 2016, 30, 8047–8054. [Google Scholar] [CrossRef]
- Hu, B.; Lu, Q.; Jiang, X.; Liu, J.; Cui, M.; Dong, C.; Yang, Y. Formation mechanism of hydroxyacetone in glucose pyrolysis: A combined experimental and theoretical study. Proc. Combust. Inst. 2019, 37, 2741–2748. [Google Scholar] [CrossRef]
- Shen, D.K.; Gu, S. The mechanism for thermal decomposition of cellulose and its main products. Bioresour. Technol. 2009, 100, 6496–6504. [Google Scholar] [CrossRef] [PubMed]
- Kiefer, D.; Merkel, M.; Lilge, L.; Hausmann, R.; Henkel, M. High cell density cultivation of Corynebacterium glutamicum on bio-based lignocellulosic acetate using pH-coupled online feeding control. Bioresour. Technol. 2021, 340, 125666. [Google Scholar] [CrossRef] [PubMed]
- Arnold, S.; Tews, T.; Kiefer, M.; Henkel, M.; Hausmann, R. Evaluation of small organic acids present in fast pyrolysis bio-oil from lignocellulose as feedstocks for bacterial bioconversion. GCB Bioenergy 2019, 11, 1159–1172. [Google Scholar] [CrossRef] [Green Version]
- Kiefer, D.; Merkel, M.; Lilge, L.; Henkel, M.; Hausmann, R. From acetate to bio-based products: Underexploited potential for industrial biotechnology. Trends Biotechnol. 2021, 39, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Kövilein, A.; Umpfenbach, J.; Ochsenreither, K. Acetate as substrate for l-malic acid production with Aspergillus oryzae DSM 1863. Biotechnol. Biofuels 2021, 14, 48. [Google Scholar] [CrossRef]
- Dörsam, S.; Kirchhoff, J.; Bigalke, M.; Dahmen, N.; Syldatk, C.; Ochsenreither, K. Evaluation of Pyrolysis Oil as Carbon Source for Fungal Fermentation. Front. Microbiol. 2016, 7, 2059. [Google Scholar] [CrossRef]
- Lange, J.; Müller, F.; Bernecker, K.; Dahmen, N.; Takors, R.; Blombach, B. Valorization of pyrolysis water: A biorefinery side stream, for 1,2-propanediol production with engineered Corynebacterium glutamicum. Biotechnol. Biofuels 2017, 10, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lian, J.; Garcia-Perez, M.; Coates, R.; Wu, H.; Chen, S. Yeast fermentation of carboxylic acids obtained from pyrolytic aqueous phases for lipid production. Bioresour. Technol. 2012, 118, 177–186. [Google Scholar] [CrossRef]
- Kubisch, C.; Ochsenreither, K. Detoxification of a pyrolytic aqueous condensate from wheat straw for utilization as substrate in Aspergillus oryzae DSM 1863 cultivations. Biotechnol. Biofuels 2022, 15, 18. [Google Scholar] [CrossRef]
- Barratt, R.W.; Johnson, G.B.; Ogata, W.N. Wild-type and mutant stocks of Aspergillus nidulans. Genetics 1965, 52, 233. [Google Scholar] [CrossRef]
- Hill, T.W.; Kafer, E. Improved protocols for Aspergillus minimal medium: Trace element and minimal medium salt stock solutions. Fungal Genet. Rep. 2001, 48, 20–21. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Zhao, X.; Chi, Z.; Rover, M.; Johnston, P.; Brown, R.; Jarboe, L.; Wen, Z. Utilization of acetic acid-rich pyrolytic bio-oil by microalga Chlamydomonas reinhardtii: Reducing bio-oil toxicity and enhancing algal toxicity tolerance. Bioresour. Technol. 2013, 133, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Chi, Z.; Rover, M.; Brown, R.; Jarboe, L.; Wen, Z. Microalgae fermentation of acetic acid-rich pyrolytic bio-oil: Reducing bio-oil toxicity by alkali treatment. Environ. Prog. Sustain. Energy 2013, 32, 955–961. [Google Scholar] [CrossRef]
- Zhao, X.; Davis, K.; Brown, R.; Jarboe, L.; Wen, Z. Alkaline treatment for detoxification of acetic acid-rich pyrolytic bio-oil for microalgae fermentation: Effects of alkaline species and the detoxification mechanisms. Biomass Bioenergy 2015, 80, 203–212. [Google Scholar] [CrossRef]
- Lian, J.; Chen, S.; Zhou, S.; Wang, Z.; O’Fallon, J.; Li, C.-Z.; Garcia-Perez, M. Separation, hydrolysis and fermentation of pyrolytic sugars to produce ethanol and lipids. Bioresour. Technol. 2010, 101, 9688–9699. [Google Scholar] [CrossRef] [PubMed]
- Cameron, D.C.; Cooney, C.L. A novel fermentation: The production of R (–)–1,2–propanediol and acetol by Clostridium thermosaccharolyticum. Biotechnology 1986, 4, 651–654. [Google Scholar] [CrossRef]
- Altaras, N.E.; Etzel, M.R.; Cameron, D.C. Conversion of Sugars to 1,2-Propanediol by Thermoanaerobacterium thermosaccharolyticum HG-8. Biotechnol. Progr. 2001, 17, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Tran-Din, K.; Gottschalk, G. Formation of D (-)-1,2-propanediol and D (-)-lactate from glucose by Clostridium sphenoides under phosphate limitation. Arch. Microbiol. 1985, 142, 87–92. [Google Scholar] [CrossRef]
- Bennett, G.N.; San, K.-Y. Microbial formation, biotechnological production and applications of 1,2-propanediol. Appl. Microbiol. Biotechnol. 2001, 55, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.; Rhee, H.; Watanabe, K.; Murata, K.; Kimura, A. Metabolism of 2-oxoaldehyde in mold: Purification and characterization of two methylglyoxal reductases from Aspergillus niger. Eur. J. Biochem. 1988, 171, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Schuurink, R.; Busink, R.; Hondmann, D.H.; Wltteveen, C.F.; Vlsser, J. Purification and properties of NADP+-dependent glycerol dehydrogenases from Aspergillus nidulans and A. niger. Microbiology 1990, 136, 1043–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walti, A. Action of Aspergillus niger on normal 1,2-diols. J. Am. Chem. Soc. 1934, 56, 2723–2726. [Google Scholar] [CrossRef]
- Murata, K.; Fukuda, Y.; Watanabe, K.; Saikusa, T.; Shimosaka, M.; Kimura, A. Characterization of methylglyoxal synthase in Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 1985, 131, 190–198. [Google Scholar] [CrossRef]
- Nakamura, K.; Kondo, S.; Kawai, Y.; Nakajima, N.; Ohno, A. Amino acid sequence and characterization of aldo-keto reductase from bakers’ yeast. Biosci. Biotechnol. Biochem. 1997, 61, 375–377. [Google Scholar] [CrossRef]
- Jung, J.-Y.; Choi, E.-S.; Oh, M.-K. Enhanced production of 1,2-propanediol by tpil deletion in Saccharomyces cerevisiae. J. Microbiol. Biotechnol. 2008, 18, 1797–1802. [Google Scholar]
- Jung, J.-Y.; Yun, H.-S.; Lee, J.-W.; Oh, M.-K. Production of 1,2-propanediol from glycerol in Saccharomyces cerevisiae. J. Microbiol. Biotechnol. 2011, 21, 846–853. [Google Scholar] [CrossRef] [Green Version]
- Jeon, E.; Lee, S.; Kim, D.; Yoon, H.; Oh, M.; Park, C.; Lee, J. Development of a Saccharomyces cerevisiae strain for the production of 1, 2-propanediol by gene manipulation. Enzyme Microb. Technol. 2009, 45, 42–47. [Google Scholar] [CrossRef]
- Lee, W.; DaSilva, N.A. Application of sequential integration for metabolic engineering of 1,2-propanediol production in yeast. Metab. Eng. 2006, 8, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Kometani, T.; Kitasuji, E.; Matsuno, R. Baker’s yeast mediated bioreduction. A new procedure using ethanol as an energy source. Chem. Lett. 1989, 1465–1466. [Google Scholar] [CrossRef]
- Kometani, T.; Kitatsuji, E.; Matsuno, R. Baker’s yeast mediated bioreduction: Practical procedure using EtOH as energy source. J. Ferment. Bioeng. 1991, 71, 197–199. [Google Scholar] [CrossRef]
- Kometani, T.; Yoshii, H.; Takeuchi, Y.; Matsuno, R. Large-scale preparation of (R)-1,2-propanediol through baker’s yeast-mediated bioreduction. J. Ferment. Bioeng. 1993, 76, 414–415. [Google Scholar] [CrossRef]
- Kometani, T.; Kitatsuji, E.; Matsuno, R. Bioreduction of ketones mediated by baker’s yeast with acetate as ultimate reducing agent. Agric. Biol. Chem. 1991, 55, 867–868. [Google Scholar]
- Yu, A.-N.; Tan, Z.-W.; Wang, F.-S. Mechanistic studies on the formation of pyrazines by Maillard reaction between L-ascorbic acid and L-glutamic acid. LWT Food Sci. Technol. 2013, 50, 64–71. [Google Scholar] [CrossRef]
- Alcano, M.D.J.; Jahn, R.C.; Scherer, C.D.; Wigmann, É.F.; Moraes, V.M.; Garcia, M.V.; Mallmann, C.A.; Copetti, M.V. Susceptibility of Aspergillus spp. to acetic and sorbic acids based on pH and effect of sub-inhibitory doses of sorbic acid on ochratoxin A production. Food Res. Int. 2016, 81, 25–30. [Google Scholar] [CrossRef]
- O’Mahony, R.J.; Burns, A.T.H.; Millam, S.; Hooley, P.; Fincham, D.A. Isotropic growth of spores and salt tolerance in Aspergillus nidulans. Mycol. Res. 2002, 106, 1480–1486. [Google Scholar] [CrossRef]
- Wang, Y.; Yan, H.; Neng, J.; Gao, J.; Yang, B.; Liu, Y. The Influence of NaCl and Glucose content on Growth and Ochratoxin A production by Aspergillus ochraceus, Aspergillus carbonarius and Penicillium nordicum. Toxins 2020, 12, 515. [Google Scholar] [CrossRef]
- Dörsam, S. Evaluation of Renewable Resources as Carbon Sources for Organic Acid Production with Filamentous Fungi. Ph.D. Thesis, Karlsruhe Institute of Technology, Karlsruhe, Germany, 2018. Available online: https://d-nb.info/115547435x/34 (accessed on 24 January 2022).
- Pinchai, N.; Juvvadi, P.R.; Fortwendel, J.R.; Perfect, B.Z.; Rogg, L.E.; Asfaw, Y.G.; Steinbach, W.J. The Aspergillus fumigatus P-type Golgi apparatus Ca2+/Mn2+ ATPase PmrA is involved in cation homeostasis and cell wall integrity but is not essential for pathogenesis. Eukaryot. Cell 2010, 9, 472–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markina-Iñarrairaegui, A.; Spielvogel, A.; Etxebeste, O.; Ugalde, U.; Espeso, E.A. Tolerance to alkaline ambient pH in Aspergillus nidulans depends on the activity of ENA proteins. Sci. Rep. 2020, 10, 14325. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, G. The Effect of Aeration on the Metabolism of Acetate by Aspergillus niger. J. Exp. Bot. 1953, 4, 369–376. [Google Scholar] [CrossRef]
- Brown, S.H.; Bashkirova, L.; Berka, R.; Chandler, T.; Doty, T.; McCall, K.; McCulloch, M.; McFarland, S.; Thompson, S.; Yaver, D. Metabolic engineering of Aspergillus oryzae NRRL 3488 for increased production of L-malic acid. Appl. Microbiol. Biotechnol. 2013, 97, 8903–8912. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhou, Y.; Cao, W.; Liu, H. Improved production of malic acid in Aspergillus niger by abolishing citric acid accumulation and enhancing glycolytic flux. ACS Synth. Biol. 2020, 9, 1418–1425. [Google Scholar] [CrossRef]
- Liu, J.; Li, J.; Liu, Y.; Shin, H.; Ledesma-Amaro, R.; Du, G.; Chen, J.; Liu, L. Synergistic rewiring of carbon metabolism and redox metabolism in cytoplasm and mitochondria of Aspergillus oryzae for increased L-malate production. ACS Synth. Biol. 2018, 7, 2139–2147. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Xie, Z.; Shin, H.; Li, J.; Du, G.; Chen, J.; Liu, L. Rewiring the reductive tricarboxylic acid pathway and L-malate transport pathway of Aspergillus oryzae for overproduction of L-malate. J. Biotechnol. 2017, 253, 1–9. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| c Acetol [g/L] | Consumed Acetate [g/L] | Consumed Acetol [g/L] | Consumed Substrate [g/L] | Final CDW [g/L] | Yx/s 1 [g/g] |
|---|---|---|---|---|---|
| 0 | 11.74 ± 1.18 | 0.00 ± 0.00 | 11.74 ± 1.18 | 3.29 ± 0.21 | 0.28 ± 0.03 |
| 3 | 11.21 ± 0.49 | 2.77 ± 0.00 | 13.97 ± 0.49 | 3.57 ± 0.35 | 0.26 ± 0.03 |
| 5 | 12.42 ± 1.57 | 4.38 ± 0.01 | 16.80 ± 1.57 | 4.37 ± 0.36 | 0.26 ± 0.03 |
| 10 | 15.09 ± 0.51 | 8.35 ± 0.13 | 23.44 ± 0.53 | 4.46 ± 0.64 | 0.19 ± 0.03 |
| 20 | 9.71 ± 1.17 | 7.96 ± 0.92 | 17.66 ± 1.49 | 3.47± 0.40 | 0.21 ± 0.02 |
| c Acetol [g/L] | Consumed Acetate [g/L] | Consumed Acetol [g/L] | Consumed Carbon [g/L] | Final Malate Titer [g/L] | Maximum Yield Yp/s, Carbon 1 [g/g] | Maximum Productivity P Max 1 [g/(L∙h)] | P Overall 2 [g/(L∙h)] |
|---|---|---|---|---|---|---|---|
| 0 | 37.16 ± 1.58 | 0.00 ± 0.00 | 15.12 ± 0.64 | 6.29 ± 0.46 | 0.18 ± 0.05 (96 h) | 0.046 ± 0.008 (96 h) | 0.026 ± 0.002 |
| 3 | 37.23 ± 1.89 | 2.73 ± 0.12 | 16.47 ± 0.77 | 6.02 ± 0.17 | 0.15 ± 0.00 (144 h) | 0.039 ± 0.000 (144 h) | 0.025 ± 0.001 |
| 5 | 36.81 ± 0.68 | 4.51 ± 0.20 | 17.17 ± 0.29 | 5.71 ± 0.38 | 0.14 ± 0.01 (144 h) | 0.037 ± 0.003 (144 h) | 0.024 ± 0.002 |
| 10 | 34.57 ± 5.53 | 9.54 ± 0.21 | 18.71 ± 2.25 | 5.64 ± 0.59 | 0.11 ± 0.02 (240 h) | 0.032 ± 0.008 (144 h) | 0.024 ± 0.002 |
| 20 | 20.86 ± 2.85 | 14.23 ± 1.41 | 15.41 ± 1.35 | 3.72 ± 0.37 | 0.09 ± 0.01 (240 h) | 0.016 ± 0.002 (240 h) | 0.016 ± 0.002 |
| Cultivation | Condition | Consumed Acetate [g/L] | Maximum Malate Titer [g/L] | Maximum Yield Yp/s 1 [g/g] | Maximum Yield Yp/s, Carbon [g/g] | Maximum Productivity P Max [g/(L∙h)] | Overall Productivity P 2 [g/(L∙h)] |
|---|---|---|---|---|---|---|---|
| first shake flask cultivation | pH not adjusted | 17.45 ± 3.01 | 3.37± 0.61 (192 h) | 0.20 ± 0.05 (192 h) | 0.18 ± 0.05 (192 h) | 0.026 ± 0.006 (96 h) | 0.013 ± 0.003 |
| pH optimization in shake flasks | pH not adjusted | 22.07 ± 4.69 | 3.96 ± 1.46 (240 h) | 0.18 ± 0.08 (240 h) | 0.16 ± 0.07 (240 h) | 0.026 ± 0.007 (96 h) | 0.016 ± 0.006 |
| pH adjusted with H2SO4 | 38.19 ± 0.75 | 8.29 ± 0.26 (240 h) | 0.22 ± 0.01 (192 h) | 0.19 ± 0.01 (192 h) | 0.054 ± 0.001 (144 h) | 0.035 ± 0.001 | |
| pH adjusted with CH3COOH | 42.82 ± 2.36 | 9.77 ± 0.55 (240 h) | 0.23 ± 0.03 (144 h) | 0.21 ± 0.02 (144 h) | 0.061 ± 0.002 (144 h) | 0.041 ± 0.002 | |
| bioreactor cultivations | acetate control | 29.71 ± 2.30 | 2.73 ± 0.47 (144 h) | 0.09 ± 0.02 (192) | 0.08 ± 0.02 (192) | 0.032 ± 0.008 (72 h) | 0.019 ± 0.003 |
| PAC | 40.82 ± 0.37 | 7.30 ± 0.29 (144 h) | 0.18 ± 0.01 (144 h) | 0.16 ± 0.01 (240) | 0.054 ± 0.010 (120 h) | 0.051 ± 0.002 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kubisch, C.; Ochsenreither, K. Valorization of a Pyrolytic Aqueous Condensate and Its Main Components for L-Malic Acid Production with Aspergillus oryzae DSM 1863. Fermentation 2022, 8, 107. https://doi.org/10.3390/fermentation8030107
Kubisch C, Ochsenreither K. Valorization of a Pyrolytic Aqueous Condensate and Its Main Components for L-Malic Acid Production with Aspergillus oryzae DSM 1863. Fermentation. 2022; 8(3):107. https://doi.org/10.3390/fermentation8030107
Chicago/Turabian StyleKubisch, Christin, and Katrin Ochsenreither. 2022. "Valorization of a Pyrolytic Aqueous Condensate and Its Main Components for L-Malic Acid Production with Aspergillus oryzae DSM 1863" Fermentation 8, no. 3: 107. https://doi.org/10.3390/fermentation8030107