
Characterization and Role of Sterols in Saccharomyces cerevisiae during White Wine Alcoholic Fermentation

 ,
, 
 , , and
, , and
Abstract
:1. Introduction
2. Importance of Sterols in Yeast Cell Survival
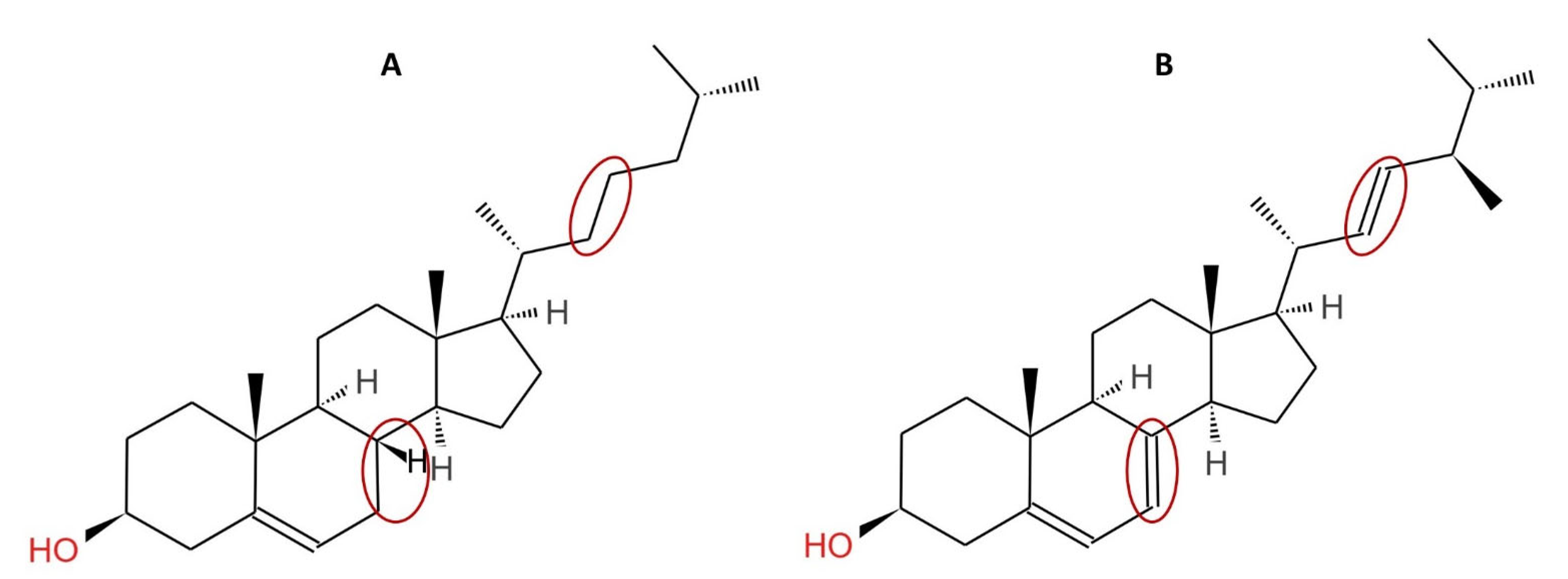
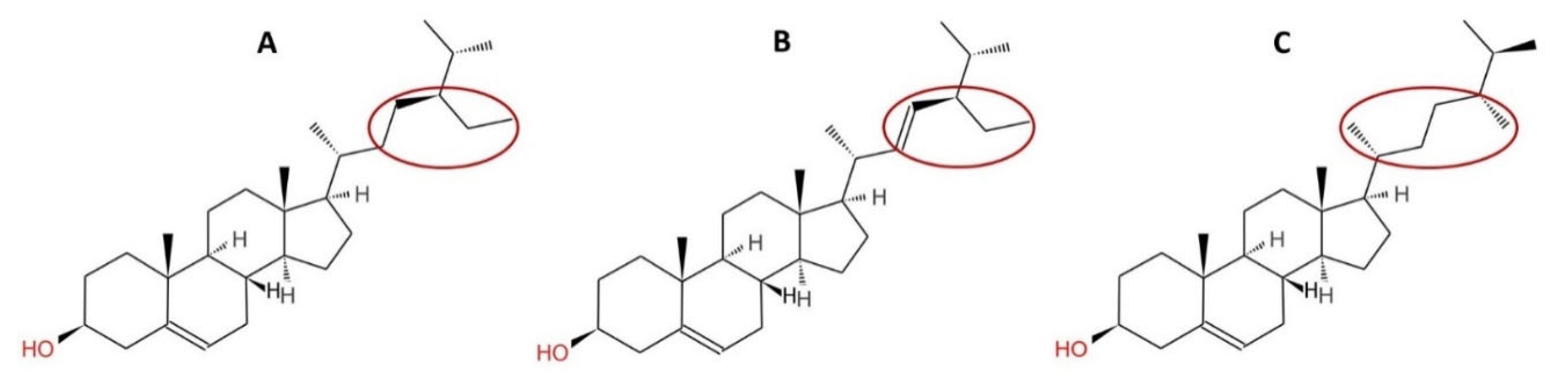
2.1. Composition, Structure and Source
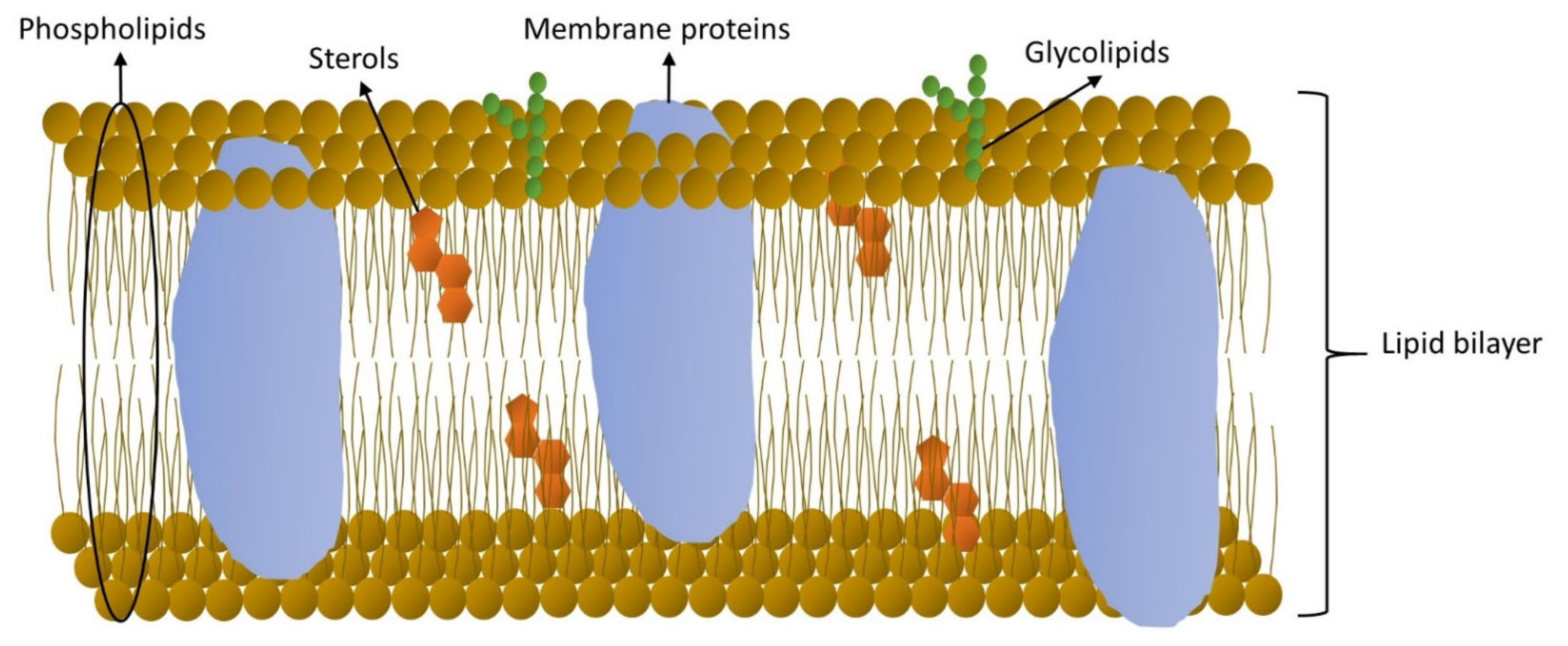
2.2. Location and Role
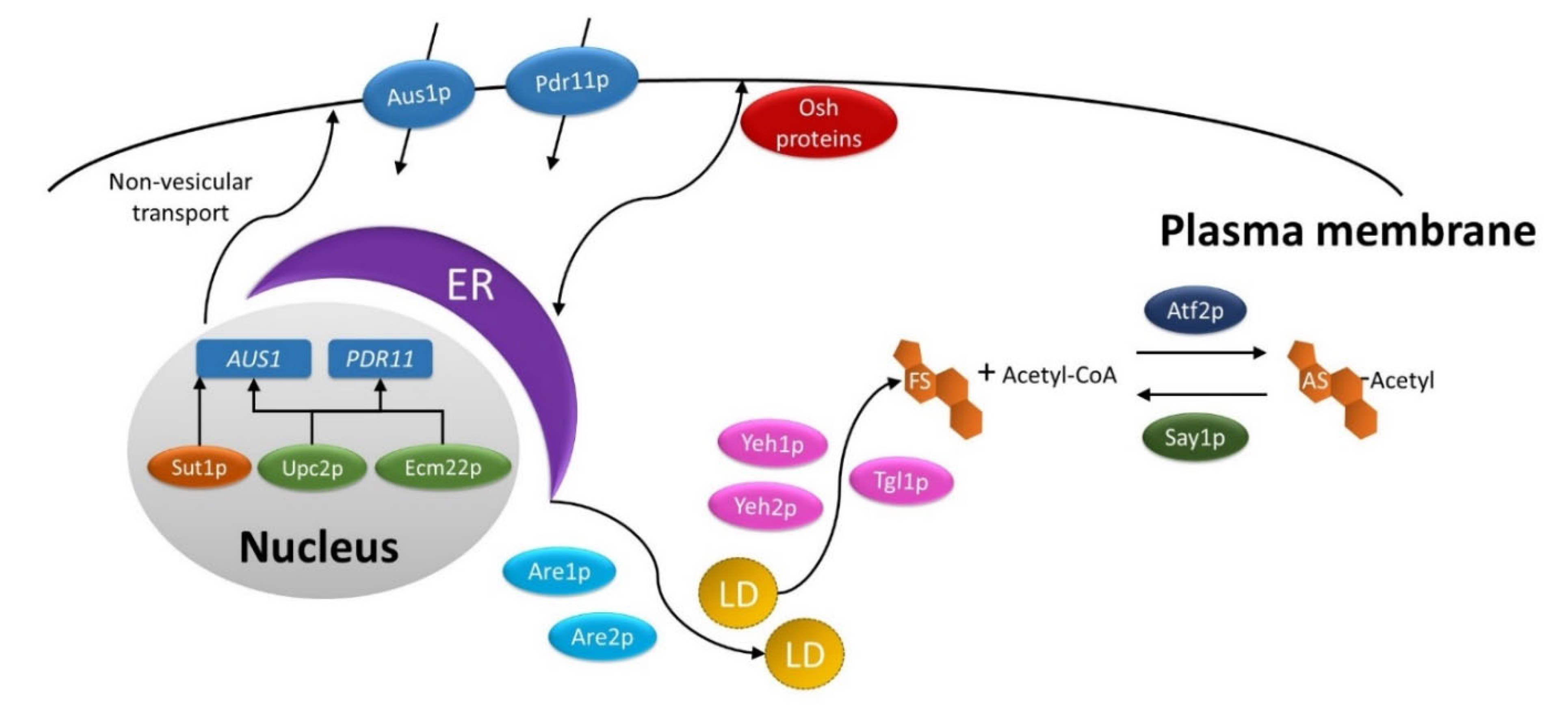
2.3. Sterol Synthesis, Metabolism Storage and Transport
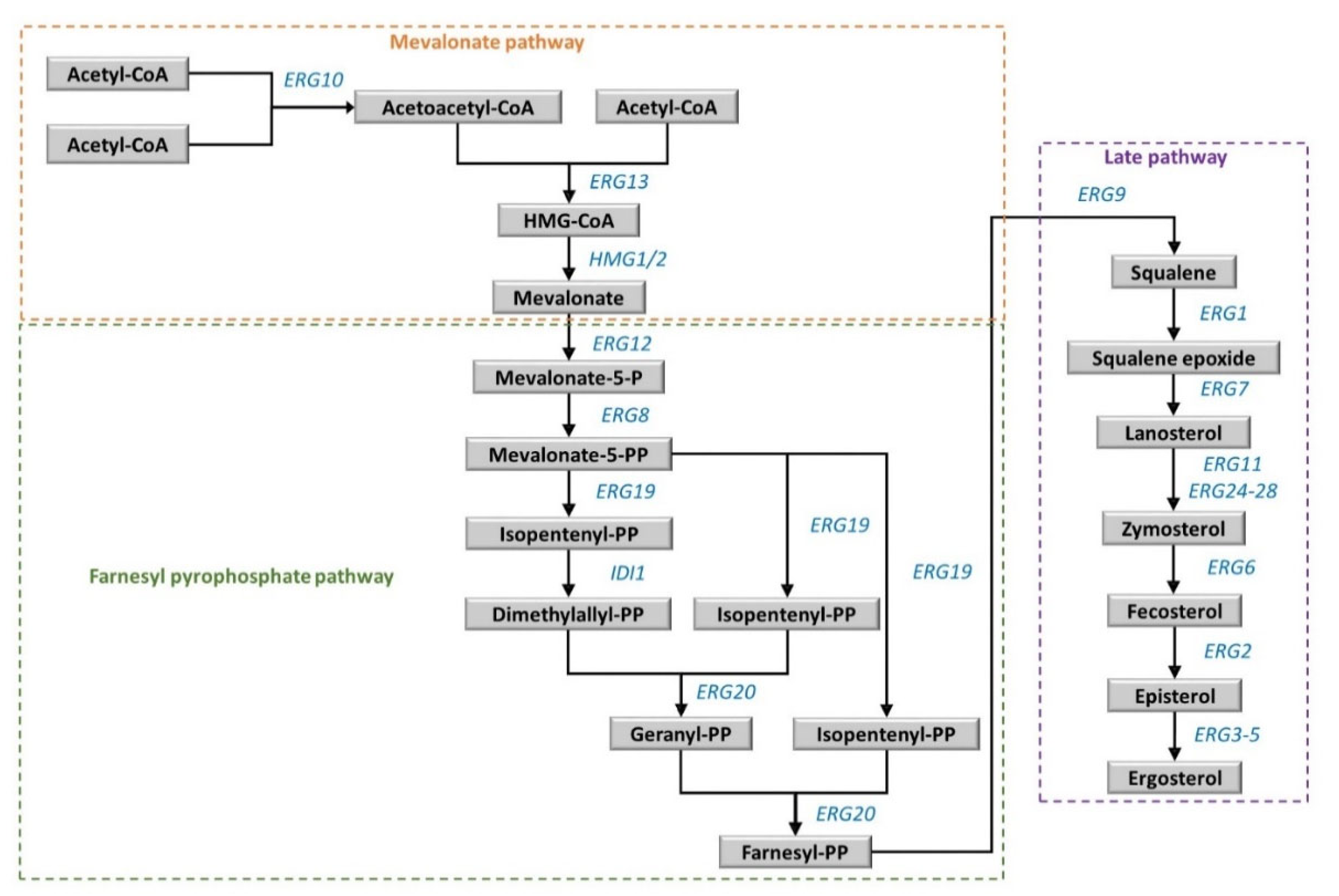
2.3.1. Sterol Synthesis and Metabolism
2.3.2. Sterol Storage and Transport
2.4. Phytosterol Assimilation
3. Impact of Sterols in White Wine Alcoholic Fermentation
3.1. Sterol Starvation
3.2. Ethanol Stress in S. cerevisiae
3.3. Effect of Temperature on Sterols
3.4. External Sources: The Addition of Grape Solid Particles, Exogenous Phytosterols, Inactive Dry Yeast Cells and Oxygen
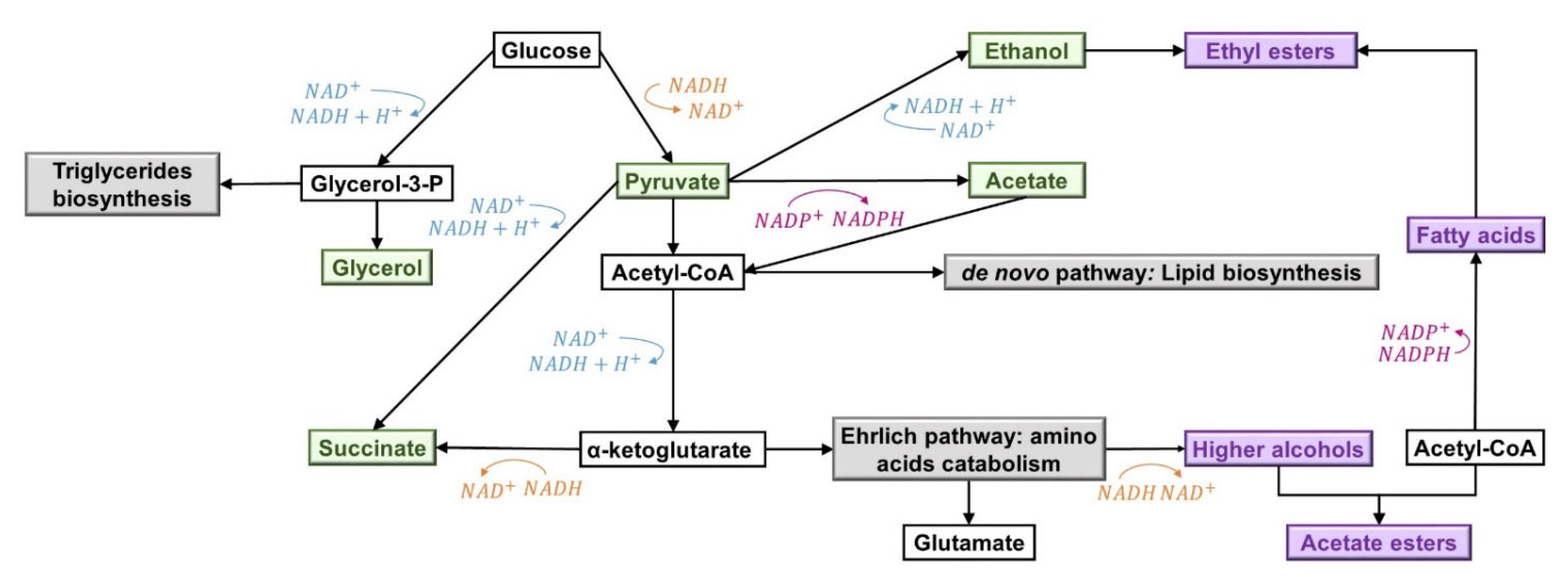
3.5. Effect of Sterols on Aroma Compounds
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wenk, M.R. The emerging field of lipidomics. Nat. Rev. Drug Discov. 2005, 4, 594–610. [Google Scholar] [CrossRef] [PubMed]
- Van Meer, G. Cellular lipidomics. EMBO J. 2005, 24, 3159–3165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mbuyane, L.L.; Bauer, F.F.; Divol, B. The metabolism of lipids in yeasts and applications in oenology. Food Res. Int. 2021, 141, 110142. [Google Scholar] [CrossRef] [PubMed]
- Fahy, E.; Subramaniam, S.; Murphy, R.C.; Nishijima, M.; Raetz, C.R.H.; Shimizu, T.; Spener, F.; Van Meer, G.; Wakelam, M.J.O.; Dennis, E.A. Update of the LIPID MAPS comprehensive classification system for lipids. J. Lipid Res. 2009, 50, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Rosenfeld, E.; Beauvoit, B.; Blondin, B.; Salmon, J.M. Oxygen consumption by anaerobic Saccharomyces cerevisiae under enological conditions: Effect on fermentation kinetics. Appl. Environ. Microbiol. 2003, 69, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Henneberry, A.L.; Sturley, S.L. Sterol homeostasis in the budding yeast, Saccharomyces cerevisiae. Semin. Cell Dev. Biol. 2005, 16, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Ruggiero, A.; Vitalini, S.; Burlini, N.; Bernasconi, S.; Iriti, M. Phytosterols in grapes and wine, and effects of agrochemicals on their levels. Food Chem. 2013, 141, 3473–3479. [Google Scholar] [CrossRef]
- Piironen, V.; Toivo, J.; Puupponen-Pimiä, R.; Lampi, A.M. Plant sterols in vegetables, fruits and berries. J. Sci. Food Agric. 2003, 83, 330–337. [Google Scholar] [CrossRef]
- Tumanov, S.; Zubenko, Y.; Greven, M.; Greenwood, D.R.; Shmanai, V.; Villas-Boas, S.G. Comprehensive lipidome profiling of Sauvignon blanc grape juice. Food Chem. 2015, 180, 249–256. [Google Scholar] [CrossRef]
- Daum, G.; Lees, N.D.; Bard, M.; Dickson, R. Biochemistry, cell biology and molecular biology of lipids of Saccharomyces cerevisiae. Yeast 1998, 14, 1471–1510. [Google Scholar] [CrossRef]
- Casalta, E.; Salmon, J.M.; Picou, C.; Sablayrolles, J.M. Grape solids: Lipid composition and role during alcoholic fermentation under enological conditions. Am. J. Enol. Vitic. 2019, 70, 147–154. [Google Scholar] [CrossRef]
- Carman, G.M.; Henry, S.A. Phosphatidic acid plays a central role in the transcriptional regulation of glycerophospholipid synthesis in Saccharomyces cerevisiae. J. Biol. Chem. 2007, 282, 37293–37297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickson, R.C. New insights into sphingolipid metabolism and function in budding yeast. J. Lipid Res. 2008, 49, 909–921. [Google Scholar] [CrossRef] [Green Version]
- Nandy, S.K.; Srivastava, R.K. A review on sustainable yeast biotechnological processes and applications. Microbiol. Res. 2018, 207, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Jordá, T.; Puig, S. Regulation of Ergosterol Biosynthesis in Saccharomyces cerevisiae. Genes 2020, 11, 795. [Google Scholar] [CrossRef] [PubMed]
- Van Der Rest, M.E.; Kamminga, A.H.; Nakano, A.; Anraku, Y.; Poolman, B.; Konings, W.N. The plasma membrane of Saccharomyces cerevisiae: Structure, function, and biogenesis. Microbiol. Rev. 1995, 59, 304–322. [Google Scholar] [CrossRef]
- Galea, A.M.; Brown, A.J. Special relationship between sterols and oxygen: Were sterols an adaptation to aerobic life? Free Radic. Biol. Med. 2009, 47, 880–889. [Google Scholar] [CrossRef]
- Jacquier, N.; Schneiter, R. Mechanisms of sterol uptake and transport in yeast. J. Steroid Biochem. Mol. Biol. 2012, 129, 70–78. [Google Scholar] [CrossRef]
- Karagiannis, S.; Lanaridis, P. Insoluble grape material present in must affects the overall fermentation aroma of dry white wines made from three grape cultivars cultivated in Greece. J. Food Sci. 2002, 67, 369–374. [Google Scholar] [CrossRef]
- Ma, T.-Z.; Gong, P.-F.; Lu, R.-R.; Zhang, B.; Morata, A.; Han, S.-Y. Effect of different clarification treatments on the volatile composition and aromatic attributes of “Italian riesling” icewine. Molecules 2020, 25, 2657. [Google Scholar] [CrossRef]
- Groat, M.; Ough, C.S. Effects of Insoluble Solids Added to Clarified Musts on Fermentation Rate, Wine Composition, and Wine Quality. Am. J. Enol. Vitic. 1978, 29, 112–119. [Google Scholar]
- Ochando, T.; Mouret, J.R.; Humbert-Goffard, A.; Sablayrolles, J.M.; Farines, V. Impact of initial lipid content and oxygen supply on alcoholic fermentation in champagne-like musts. Food Res. Int. 2017, 98, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Casalta, E.; Cervi, M.F.; Salmon, J.M.; Sablayrolles, J.M. White wine fermentation: Interaction of assimilable nitrogen and grape solids on alcoholic fermentation under oenological conditions. Aust. J. Grape Wine Res. 2013, 19, 47–52. [Google Scholar] [CrossRef]
- Bely, M.; Sablayrolles, J.M.; Barre, P. Description of Alcoholic Fermentation Kinetics: Its Variability and Significance. Am. J. Enol. Vitic. 1990, 41, 319–324. [Google Scholar]
- Bauer, F.F.; Pretorius, I.S. Yeast Stress Response and Fermentation Efficiency: How to Survive the Making of Wine—A Review. S. Afr. J. Enol. Vitic. 2000, 21, 27–51. [Google Scholar] [CrossRef]
- Ratledge, C.; Evans, C. Lipids and their Metabolism. In The Yeasts; Rose, A.H., Harrison, J.S., Eds.; Academic Press Limited: London, UK, 1989; Volume III, Metabolism and Physiology of Yeasts; pp. 367–455. [Google Scholar]
- Chi, Z.; Arneborg, N. Relationship between lipid composition, frequency of ethanol-induced respiratory deficient mutants, and ethanol tolerance in Saccharomyces cerevisiae. J. Appl. Microbiol. 1999, 86, 1047–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilera, F.; Peinado, R.A.; Millán, C.; Ortega, J.M.; Mauricio, J.C. Relationship between ethanol tolerance, H+-ATPase activity and the lipid composition of the plasma membrane in different wine yeast strains. Int. J. Food Microbiol. 2006, 110, 34–42. [Google Scholar] [CrossRef]
- Lucero, P.; Peñalver, E.; Moreno, E.; Lagunas, R. Internal trehalose protects endocytosis from inhibition by ethanol in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2000, 66, 4456–4461. [Google Scholar] [CrossRef] [Green Version]
- Dinh, T.N.; Nagahisa, K.; Hirasawa, T.; Furusawa, C.; Shimizu, H. Adaptation of Saccharomyces cerevisiae cells to high ethanol concentration and changes in fatty acid composition of membrane and cell size. PLoS ONE 2008, 3, e2623. [Google Scholar] [CrossRef]
- Bravim, F.; de Freitas, J.M.; Fernandes, A.A.R.; Fernandes, P.M.B. High hydrostatic pressure and the cell membrane: Stress response of Saccharomyces cerevisiae. Ann. N. Y. Acad. Sci. 2010, 1189, 127–132. [Google Scholar] [CrossRef]
- Turanlı-Yıldız, B.; Benbadis, L.; Alkım, C.; Sezgin, T.; Akşit, A.; Gökçe, A.; Öztürk, Y.; Baykal, A.T.; Çakar, Z.P.; François, J.M. In vivo evolutionary engineering for ethanol-tolerance of Saccharomyces cerevisiae haploid cells triggers diploidization. J. Biosci. Bioeng. 2017, 124, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Redón, M.; Guillamón, J.M.; Mas, A.; Rozès, N. Effect of lipid supplementation upon Saccharomyces cerevisiae lipid composition and fermentation performance at low temperature. Eur. Food Res. Technol. 2009, 228, 833–840. [Google Scholar] [CrossRef]
- Tesnière, C. Importance and role of lipids in wine yeast fermentation. Appl. Microbiol. Biotechnol. 2019, 103, 8293–8300. [Google Scholar] [CrossRef] [PubMed]
- Nes, W.D. Biosynthesis of cholesterol and other sterols. Chem. Rev. 2011, 111, 6423–6451. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J. Systems biology of lipid metabolism: From yeast to human. FEBS Lett. 2009, 583, 3905–3913. [Google Scholar] [CrossRef] [Green Version]
- Sturley, S. Conservation of eukaryotic sterol homeostasis: New insights from studies in budding yeast. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2001, 1529, 155–163. [Google Scholar] [CrossRef]
- Ejsing, C.S.; Sampaio, J.L.; Surendranath, V.; Duchoslav, E.; Ekroos, K.; Klemm, R.W.; Simons, K.; Shevchenko, A. Global analysis of the yeast lipidome by quantitative shotgun mass spectrometry. Proc. Natl. Acad. Sci. USA 2009, 106, 2136–2141. [Google Scholar] [CrossRef] [Green Version]
- Rattray, J.B.M.; Schibeci, A.; Kidby, D.K. Lipids of yeasts. Bacteriol. Rev. 1975, 39, 197–231. [Google Scholar] [CrossRef]
- Zinser, E.; Paltauf, F.; Daum, G. Sterol composition of yeast organelle membranes and subcellular distribution of enzymes involved in sterol metabolism. J. Bacteriol. 1993, 175, 2853–2858. [Google Scholar] [CrossRef] [Green Version]
- JSME Molecular Editor. Available online: https://vchem3d.univ-tlse3.fr/vM_2Djmol.html (accessed on 6 February 2022).
- Piironen, V.; Lindsay, D.G.; Miettinen, T.A.; Toivo, J.; Lampi, A.M. Plant sterols: Biosynthesis, biological function and their importance to human nutrition. J. Sci. Food Agric. 2000, 80, 939–966. [Google Scholar] [CrossRef]
- Burlini, N.; Iriti, M.; Daghetti, A.; Faoro, F.; Ruggiero, A.; Bernasconi, S. Benzothiadiazole (BTH) activates sterol pathway and affects vitamin D3 metabolism in Solanum malacoxylon cell cultures. Plant Cell Rep. 2011, 30, 2131–2141. [Google Scholar] [CrossRef] [PubMed]
- Caspeta, L.; Chen, Y.; Ghiaci, P.; Feizi, A.; Baskov, S.; Hallström, B.M.; Petranovic, D.; Nielsen, J. Altered sterol composition renders yeast thermotolerant. Science 2014, 346, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, P.S.; Heiman, M.G.; Walther, T.C.; Engel, A.; Schwudke, D.; Gushwa, N.; Kurzchalia, T.; Walter, P. Structure of sterol aliphatic chains affects yeast cell shape and cell fusion during mating. Proc. Natl. Acad. Sci. USA 2010, 107, 4170–4175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escribá, P.V.; González-Ros, J.M.; Goñi, F.M.; Kinnunen, P.K.J.; Vigh, L.; Sánchez-Magraner, L.; Fernández, A.M.; Busquets, X.; Horváth, I.; Barceló-Coblijn, G. Membranes: A meeting point for lipids, proteins and therapies: Translational Medicine. J. Cell. Mol. Med. 2008, 12, 829–875. [Google Scholar] [CrossRef] [Green Version]
- Rego, A.; Trindade, D.; Chaves, S.R.; Manon, S.; Costa, V.; Sousa, M.J.; Côrte-Real, M. The yeast model system as a tool towards the understanding of apoptosis regulation by sphingolipids. FEMS Yeast Res. 2014, 14, 160–178. [Google Scholar] [CrossRef] [Green Version]
- Sokolov, S.S.; Trushina, N.I.; Severin, F.F.; Knorre, D.A. Ergosterol Turnover in Yeast: An Interplay between Biosynthesis and Transport. Biochemistry 2019, 84, 346–357. [Google Scholar] [CrossRef]
- Klug, L.; Daum, G. Yeast lipid metabolism at a glance. FEMS Yeast Res. 2014, 14, 369–388. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.J.; Crowley, J.H.; Parks, L.W. Transcriptional regulation by ergosterol in the yeast Saccharomyces cerevisiae. Mol. Cell. Biol. 1996, 16, 5427–5432. [Google Scholar] [CrossRef] [Green Version]
- Leber, R.; Zenz, R.; Schröttner, K.; Fuchsbichler, S.; Pühringer, B.; Turnowsky, F. A novel sequence element is involved in the transcriptional regulation of expression of the ERG1 (squalene epoxidase) gene in Saccharomyces cerevisiae. Eur. J. Biochem. 2001, 268, 914–924. [Google Scholar] [CrossRef]
- Mollinedo, F. Lipid raft involvement in yeast cell growth and death. Front. Oncol. 2012, 2, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, F.J.; Douglas, L.M.; Konopka, J.B. Sterol-rich plasma membrane domains in fungi. Eukaryot. Cell 2007, 6, 755–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merzendorfer, H.; Heinisch, J.J. Microcompartments within the yeast plasma membrane. Biol. Chem. 2013, 394, 189–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ermakova, E.; Zuev, Y. Effect of ergosterol on the fungal membrane properties. All-atom and coarse-grained molecular dynamics study. Chem. Phys. Lipids 2017, 209, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Kodedová, M.; Sychrová, H. Changes in the sterol composition of the plasma membrane affect membrane potential, salt tolerance and the activity of multidrug resistance pumps in Saccharomyces cerevisiae. PLoS ONE 2015, 10, e0139306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.C. Implications of sterol structure for membrane lipid composition, fluidity and phospholipid asymmetry in Saccharomyces cerevisiae. FEMS Yeast Res. 2006, 6, 1047–1051. [Google Scholar] [CrossRef] [Green Version]
- Guan, X.L.; Souza, C.M.; Pichler, H.; Dewhurst, G.; Schaad, O.; Kentaro Kajiwara, H.W.; Ivanova, T.; Castillon, G.A.; Piccolis, M.; Abe, F.; et al. Functional Interactions between Sphingolipids and Sterols in Biological Membranes Regulating Cell Physiology. Mol. Biol. Cell 2009, 20, 2673–2683. [Google Scholar] [CrossRef] [Green Version]
- Abe, F.; Hiraki, T. Mechanistic role of ergosterol in membrane rigidity and cycloheximide resistance in Saccharomyces cerevisiae. Biochim. Biophys. Acta-Biomembr. 2009, 1788, 743–752. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, R.A.; Godinho, C.P.; Vitorino, M.V.; Robalo, T.T.; Fernandes, F.; Rodrigues, M.S.; Sá-Correia, I. Crosstalk between Yeast Cell Plasma Membrane Ergosterol Content and Cell Wall Stiffness under Acetic Acid Stress Involving Pdr18. J. Fungi 2022, 8, 103. [Google Scholar] [CrossRef]
- Silva, C.; Aranda, F.J.; Ortiz, A.; Martinez, V.; Carvajal, M.; Teruel, J.A. Molecular aspects of the interaction between plants sterols and DPPC bilayers—An experimental and theoretical approach. J. Colloid Interface Sci. 2011, 358, 192–201. [Google Scholar] [CrossRef]
- Wojciechowski, Z. Biochemistry of Phytosterol Conjugates. In Physiology and Biochemistry of Sterols; Nes, W., Patterson, G.W., Eds.; American Oil Chemists’ Society: Champaign, IL, USA, 1991; pp. 361–395. [Google Scholar]
- Dyas, L.; Goad, L.J. Steryl fatty acyl esters in plants. Phytochemistry 1993, 34, 17–29. [Google Scholar] [CrossRef]
- Hartmann, M.A. Plant sterols and the membrane environment. Trends Plant Sci. 1998, 3, 170–175. [Google Scholar] [CrossRef]
- Clouse, S.D. Plant development: A role for sterols in embryogenesis. Curr. Biol. 2000, 10, R601–R604. [Google Scholar] [CrossRef] [Green Version]
- Altmann, K.; Westermann, B. Role of essential genes in mitochondrial morphogenesis in Saccharomyces cerevisiae. Mol. Biol. Cell 2005, 16, 5410–5417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, S.; Esquivel, B.D.; White, T.C. Overexpression or deletion of ergosterol biosynthesis genes alters doubling time, response to stress agents, and drug susceptibility in Saccharomyces cerevisiae. MBio 2018, 9, e01291-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Z.; He, B.; Ma, L.; Sun, Y.; Niu, Y.; Zeng, B. Recent Advances in Ergosterol Biosynthesis and Regulation Mechanisms in Saccharomyces cerevisiae. Indian J. Microbiol. 2017, 57, 270–277. [Google Scholar] [CrossRef]
- Hiser, L.; Basson, M.E.; Rine, J. ERG10 from Saccharomyces cerevisiae encodes acetoacetyl-CoA thiolase. J. Biol. Chem. 1994, 269, 31383–31389. [Google Scholar] [CrossRef]
- Miziorko, H.M. Enzymes of the mevalonate pathway of isoprenoid biosynthesis. Arch. Biochem. Biophys. 2011, 505, 131–143. [Google Scholar] [CrossRef] [Green Version]
- Basson, M.E.; Thorsness, M.; Rine, J. Saccharomyces cerevisiae contains two functional genes encoding 3-hydroxy-3-methylglutaryl-coenzyme A reductase. Proc. Natl. Acad. Sci. USA 1986, 83, 5563–5567. [Google Scholar] [CrossRef] [Green Version]
- Kristan, K.; Rižner, T.L. Steroid-transforming enzymes in fungi. J. Steroid Biochem. Mol. Biol. 2012, 129, 79–91. [Google Scholar] [CrossRef]
- Espenshade, P.J.; Hughes, A.L. Regulation of sterol synthesis in eukaryotes. Annu. Rev. Genet. 2007, 41, 401–427. [Google Scholar] [CrossRef]
- Davies, B.S.J.; Rine, J. A role for sterol levels in oxygen sensing in Saccharomyces cerevisiae. Genetics 2006, 174, 191–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foresti, O.; Ruggiano, A.; Hannibal-Bach, H.K.; Ejsing, C.S.; Carvalho, P. Sterol homeostasis requires regulated degradation of squalene monooxygenase by the ubiquitin ligase Doa10/Teb4. Elife 2013, 2013, e00953. [Google Scholar] [CrossRef] [PubMed]
- Burg, J.S.; Espenshade, P.J. Regulation of HMG-CoA reductase in mammals and yeast. Prog. Lipid Res. 2011, 50, 403–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köffel, R.; Tiwari, R.; Falquet, L.; Schneiter, R. TGL1 Genes Encode a Novel Family of Membrane-Anchored Lipases That Are Required for Steryl Ester Hydrolysis. Mol. Cell Biol. 2005, 25, 1655–1668. [Google Scholar] [CrossRef] [Green Version]
- Maxfield, F.R.; Menon, A.K. Intracellular sterol transport and distribution. Curr. Opin. Cell Biol. 2006, 18, 379–385. [Google Scholar] [CrossRef]
- Welte, M.A.; Gould, A.P. Lipid droplet functions beyond energy storage. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2017, 1862, 1260–1272. [Google Scholar] [CrossRef]
- Tiwari, R.; Köffel, R.; Schneiter, R. An acetylation/deacetylation cycle controls the export of sterols and steroids from S. cerevisiae. EMBO J. 2007, 26, 5109–5119. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, V.; Schneiter, R. Pathogen-related yeast (PRY) proteins and members of the CAP superfamily are secreted sterol-binding proteins. Proc. Natl. Acad. Sci. USA 2012, 109, 16882–16887. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, D.P.; Ohvo-Rekilä, H.; Baumann, N.A.; Beh, C.T.; Menon, A.K. Sterol trafficking between the endoplasmic reticulum and plasma membrane in yeast. Biochem. Soc. Trans. 2006, 34, 356–358. [Google Scholar] [CrossRef]
- Schulz, T.; Prinz, W. Sterol Transport in Yeast and the Oxysterol Binding Protein Homologue (OSH) Family. Biochim. Biophys. Acta BBA-Mol. Cell Biol. Lipid 2007, 1771, 769–780. [Google Scholar] [CrossRef] [Green Version]
- Beh, C.T.; Cool, L.; Phillips, J.; Rine, J. Overlapping functions of the yeast oxysterol-binding protein homologues. Genetics 2001, 157, 1117–1140. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhang, Y.; Li, H.; Hai, K.C.; Munn, A.L.; Yang, H. AAA ATPases regulate membrane association of yeast oxysterol binding proteins and sterol metabolism. EMBO J. 2005, 24, 2989–2999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tinkelenberg, A.H.; Liu, Y.; Alcantara, F.; Khan, S.; Guo, Z.; Bard, M.; Sturley, S.L. Mutations in Yeast ARV1 Alter Intracellular Sterol Distribution and Are Complemented by Human ARV1. J. Biol. Chem. 2000, 275, 40667–40670. [Google Scholar] [CrossRef] [Green Version]
- Malathi, K.; Higaki, K.; Tinkelenberg, A.H.; Balderes, D.A.; Almanzar-Paramio, D.; Wilcox, L.J.; Erdeniz, N.; Redican, F.; Padamsee, M.; Liu, Y.; et al. Mutagenesis of the putative sterol-sensing domain of yeast Niemann Pick C-related protein reveals a primordial role in subcellular sphingolipid distribution. J. Cell Biol. 2004, 164, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Ren, J.; Li, H.; Zhang, Q.; Armstrong, J.S.; Munn, A.L.; Yang, H. Ncr1p, the yeast ortholog of mammalian Niemann Pick C1 protein, is dispensable for endocytic transport. Traffic 2004, 5, 1017–1030. [Google Scholar] [CrossRef] [PubMed]
- Berger, A.C.; Vanderford, T.H.; Gernert, K.M.; Nichols, J.W.; Faundez, V.; Corbett, A.H. Saccharomyces cerevisiae Npc2p is a functionally conserved homologue of the human Niemann-Pick Disease Type C 2 protein, hNPC2. Eukaryot. Cell 2005, 4, 1851–1862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Prinz, W.A. ATP-binding cassette (ABC) transporters mediate nonvesicular, raft-modulated sterol movement from the plasma membrane to the endoplasmic reticulum. J. Biol. Chem. 2004, 279, 45226–45234. [Google Scholar] [CrossRef] [Green Version]
- Valachovič, M.; Hronská, L.; Hapala, I. Anaerobiosis induces complex changes in sterol esterification pattern in the yeast Saccharomyces cerevisiae. FEMS Microbiol. Lett. 2001, 197, 41–45. [Google Scholar] [CrossRef] [Green Version]
- Köffel, R.; Schneiter, R. Yeh1 constitutes the major steryl ester hydrolase under heme-deficient conditions in Saccharomyces cerevisiae. Eukaryot. Cell 2006, 5, 1018–1025. [Google Scholar] [CrossRef] [Green Version]
- Scarcelli, J.J.; Hodge, C.A.; Cole, C.N. The yeast integral membrane protein Apq12 potentially links membrane dynamics to assembly of nuclear pore complexes. J. Cell Biol. 2007, 178, 799–812. [Google Scholar] [CrossRef] [Green Version]
- Hodge, C.A.; Choudhary, V.; Wolyniak, M.J.; Scarcelli, J.J.; Schneiter, R.; Cole, C.N. Integral membrane proteins Brr6 and Apq12 link assembly of the nuclear pore complex to lipid homeostasis in the endoplasmic reticulum. J. Cell Sci. 2010, 123, 141–151. [Google Scholar] [CrossRef] [Green Version]
- Schneiter, R.; Cole, C.N. Integrating complex functions: Coordination of nuclear pore complex assembly and membrane expansion of the nuclear envelope requires a family of integral membrane proteins. Nucleus 2010, 1, 387–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raychaudhuri, S.; Prinz, W.A. Uptake and trafficking of exogenous sterols in Saccharomyces cerevisiae. Biochem. Soc. Trans. 2006, 34, 359–362. [Google Scholar] [CrossRef]
- Lorenz, R.T.; Rodriguez, R.J.; Lewis, T.A.; Parks, L.W. Characteristics of sterol uptake in Saccharomyces cerevisiae. J. Bacteriol. 1986, 167, 981–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casalta, E.; Cervi, M.; Sablayrolles, J.; Salmon, J. Effet combiné des niveaux d’azote assimilable et de bourbes: Nouveau paramètre à prendre en compte pour la maîtrise de la fermentation alcoolique. Rev. Fr. D’oenol. 2012, 255, 9–15. [Google Scholar]
- Lorenz, R.T.; Parks, L.W. Involvement of heme components in sterol metabolism of Saccharomyces cerevisiae. Lipids 1991, 26, 598–603. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, L.J.; Balderes, D.A.; Wharton, B.; Tinkelenberg, A.H.; Rao, G.; Sturley, S.L. Transcriptional profiling identifies two members of the ATP-binding cassette transporter superfamily required for sterol uptake in yeast. J. Biol. Chem. 2002, 277, 32466–32472. [Google Scholar] [CrossRef] [Green Version]
- Alimardani, P.; Régnacq, M.; Moreau-Vauzelle, C.; Ferreira, T.; Rossignol, T.; Blondin, B.; Bergès, T. SUT1-promoted sterol uptake involves the ABC transporter Aus1 and the mannoprotein Dan1 whose synergistic action is sufficient for this process. Biochem. J. 2004, 381, 195–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tesnière, C.; Pradal, M.; Legras, J.L. Sterol uptake analysis in Saccharomyces and non-Saccharomyces wine yeast species. FEMS Yeast Res. 2021, 21, foab020. [Google Scholar] [CrossRef] [PubMed]
- Henry, S. Membrane lipids of yeast: Biochemical and genetic studies. In Molecular Biology of the Yeast Saccharomyces Cerevisiae: Metabolism and Gene Expression; Strathern, J.N., Jones, E.W., Broach, J.R., Eds.; Cold Spring Harbor Laboratory: New York, NY, USA, 1982; pp. 101–158. [Google Scholar]
- Deroite, A.; Legras, J.L.; Rigou, P.; Ortiz-Julien, A.; Dequin, S. Lipids modulate acetic acid and thiol final concentrations in wine during fermentation by Saccharomyces cerevisiae × Saccharomyces kudriavzevii hybrids. AMB Exp. 2018, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Bardi, L.; Crivelli, C.; Marzona, M. Esterase activity and release of ethyl esters of medium-chain fatty acids by Saccharomyces cerevisiae during anaerobic growth. Can. J. Microbiol. 1998, 44, 1171–1176. [Google Scholar] [CrossRef] [PubMed]
- Parks, L.W. Metabolism of sterols in yeast. CRC Crit. Rev. Microbiol. 1978, 6, 301–341. [Google Scholar] [CrossRef] [PubMed]
- Beltran, G.; Novo, M.; Leberre, V.; Sokol, S.; Labourdette, D.; Guillamon, J.-M.; Mas, A.; François, J.; Rozes, N. Integration of transcriptomic and metabolic analyses for understanding the global responses of low-temperature winemaking fermentations. FEMS Yeast Res. 2006, 6, 1167–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deytieux, C.; Mussard, L.; Biron, M.J.; Salmon, J.M. Fine measurement of ergosterol requirements for growth of Saccharomyces cerevisiae during alcoholic fermentation. Appl. Microbiol. Biotechnol. 2005, 68, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Jollow, D.; Kellerman, G.M.; Linnane, A.W. The biogenesis of mitochondria. 3. The lipid composition of aerobically and anaerobically grown Saccharomyces cerevisiae as related to the membrane systems of the cells. J. Cell Biol. 1968, 37, 221–230. [Google Scholar] [CrossRef]
- Jahnke, L.; Klein, H.P. Oxygen requirements for formation and activity of the squalene epoxidase in Saccharomyces cerevisiae. J. Bacteriol. 1983, 155, 488–492. [Google Scholar] [CrossRef] [Green Version]
- Fornairon-Bonnefond, C.; Demaretz, V.; Rosenfeld, E.; Salmon, J.M. Oxygen addition and sterol synthesis in Saccharomyces cerevisiae during enological fermentation. J. Biosci. Bioeng. 2002, 93, 176–182. [Google Scholar] [CrossRef]
- Da Costa, B.L.V.; Raghavendran, V.; Franco, M.; De Britto, A.; Filho, C.; Yoshinaga, M.Y.; Miyamoto, S.; Basso, T.O.; Gombert, A.K. Forever panting and forever growing: Physiology of Saccharomyces cerevisiae at extremely low oxygen availability in the absence of ergosterol and unsaturated fatty acids. FEMS Yeast Res. 2019, 19, foz054. [Google Scholar] [CrossRef]
- Julien, A.; Roustan, J.L.; Dulau, L.; Sablayrolles, J.M. Comparison of nitrogen and oxygen demands of enological yeasts: Technological consequences. Am. J. Enol. Vitic. 2000, 51, 215–222. [Google Scholar]
- Tesnière, C.; Delobel, P.; Pradal, M.; Blondin, B. Impact of Nutrient Imbalance on Wine Alcoholic Fermentations: Nitrogen Excess Enhances Yeast Cell Death in Lipid-Limited Must. PLoS ONE 2013, 8, e61645. [Google Scholar] [CrossRef] [Green Version]
- Duc, C.; Pradal, M.; Sanchez, I.; Noble, J.; Tesnière, C.; Blondin, B. A set of nutrient limitations trigger yeast cell death in a nitrogen-dependent manner during wine alcoholic fermentation. PLoS ONE 2017, 12, e0184838. [Google Scholar] [CrossRef] [PubMed]
- Abe, F. Induction of DAN/TIR yeast cell wall mannoprotein genes in response to high hydrostatic pressure and low temperature. FEBS Lett. 2007, 581, 4993–4998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumann, K.; Dato, L.; Graf, A.B.; Frascotti, G.; Dragosits, M.; Porro, D.; Mattanovich, D.; Ferrer, P.; Branduardi, P. The impact of oxygen on the transcriptome of recombinant S. cerevisiae and P. pastoris—A comparative analysis. BMC Genom. 2011, 12, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rachidi, N.; Barre, P.; Blondin, B. Examination of the transcriptional specificity of an enological yeast. A pilot experiment on the chromosome-III right arm. Curr. Genet. 2000, 37, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zitomer, R.S.; Carrico, P.; Deckert, J. Regulation of hypoxic gene expression in yeast. Kidney Int. 1997, 51, 507–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duc, C.; Pradal, M.; Sanchez, I.; Noble, J.; Blondin, B.; Tesnière, C. Specific gene regulations of unusual micronutrient starvations leading to cell death during wine fermentation. OENO One 2020, 54, 359–371. [Google Scholar] [CrossRef]
- Luparia, V.; Soubeyrand, V.; Berges, T.; Julien, A.; Salmon, J.M. Assimilation of grape phytosterols by Saccharomyces cerevisiae and their impact on enological fermentations. Appl. Microbiol. Biotechnol. 2004, 65, 25–32. [Google Scholar] [CrossRef]
- Belviso, S.; Bardi, L.; Bartolini, A.B.; Marzona, M. Lipid nutrition of Saccharomyces cerevisiae in winemaking. Can. J. Microbiol. 2004, 50, 669–674. [Google Scholar] [CrossRef]
- Deroite, A.; Legras, J.L.; Ortiz-Julien, A.; Dequin, S. Reduction of acetic acid production during wine fermentation by Saccharomyces cerevisiae × Saccharomyces kudriavzevii hybrids using adaptive evolution under lipids limitation. In Proceedings of the ISSY 34, Bariloche, Argentina, 1–4 October 2018. [Google Scholar]
- Rollero, S.; Bloem, A.; Camarasa, C.; Sanchez, I.; Ortiz-Julien, A.; Sablayrolles, J.-M.; Dequin, S.; Mouret, J.-R. Combined effects of nutrients and temperature on the production of fermentative aromas by Saccharomyces cerevisiae during wine fermentation. Appl. Microbiol. Biotechnol. 2015, 99, 2291–2304. [Google Scholar] [CrossRef]
- You, K.M.; Rosenfield, C.L.; Knipple, D.C. Ethanol tolerance in the yeast Saccharomyces cerevisiae is dependent on cellular oleic acid content. Appl. Environ. Microbiol. 2003, 69, 1499–1503. [Google Scholar] [CrossRef] [Green Version]
- Mannazzu, I.; Angelozzi, D.; Belviso, S.; Budroni, M.; Farris, G.A.; Goffrini, P.; Lodi, T.; Marzona, M.; Bardi, L. Behaviour of Saccharomyces cerevisiae wine strains during adaptation to unfavourable conditions of fermentation on synthetic medium: Cell lipid composition, membrane integrity, viability and fermentative activity. Int. J. Food Microbiol. 2008, 121, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Liu, Z.L. Mechanisms of ethanol tolerance in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2010, 87, 829–845. [Google Scholar] [CrossRef] [PubMed]
- Madeira, A.; Leitão, L.; Soveral, G.; Dias, P.; Prista, C.; Moura, T.; Loureiro-Dias, M.C. Effect of ethanol on fluxes of water and protons across the plasma membrane of Saccharomyces cerevisiae. FEMS Yeast Res. 2010, 10, 252–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucero, P.; Peñalver, E.; Moreno, E.; Lagunas, R. Moderate concentrations of ethanol inhibit endocytosis of the yeast maltose transporter. Appl. Environ. Microbiol. 1997, 63, 3831–3836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, P. Tolerance of fungi to ethanol. In Stress Tolerance of Fungi; Jennings, D.H., Ed.; CRC Press: Boca Raton, FL, USA, 1993; pp. 189–208. [Google Scholar]
- Piper, P.W.; Talreja, K.; Panaretou, B.; Moradas-Ferreira, P.; Byrne, K.; Praekelt, U.M.; Meacock, P.; Récnacq, M.; Boucherie, H. Induction of major heat-shock proteins of Saccharomyces cerevisiae, including plasma membrane Hsp30, by ethanol levels above a critical threshold. Microbiology 1994, 140, 3031–3038. [Google Scholar] [CrossRef] [Green Version]
- Piper, P.; Ortiz-Calderon, C.; Holyoak, C.; Coote, P.; Cole, M. Hsp30, the integral plasma membrane heat shock protein of Saccharomyces cerevisiae, is a stress-inducible regulator of plasma membrane H(+)-ATPase. Cell Stress Chaperones 1997, 2, 12–24. [Google Scholar] [CrossRef] [Green Version]
- Dickey, A.N.; Yim, W.S.; Faller, R. Using ergosterol to mitigate the deleterious effects of ethanol on bilayer structure. J. Phys. Chem. B 2009, 113, 2388–2397. [Google Scholar] [CrossRef]
- Rose, A.H. Composition of the envelope layers of Saccharomyces cerevisiae in relation to flocculation and ethanol tolerance. J. Appl. Bacteriol. 1993, 74, 110S–118S. [Google Scholar] [CrossRef]
- Jones, R.P. Biological principles for the effects of ethanol. Enzym. Microb. Technol. 1989, 11, 130–153. [Google Scholar] [CrossRef]
- Henderson, C.M.; Block, D.E. Examining the role of membrane lipid composition in determining the ethanol tolerance of Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2014, 80, 2966–2972. [Google Scholar] [CrossRef] [Green Version]
- Johnston, E.J.; Moses, T.; Rosser, S.J. The wide-ranging phenotypes of ergosterol biosynthesis mutants, and implications for microbial cell factories. Yeast 2020, 37, 27–44. [Google Scholar] [CrossRef] [Green Version]
- Alexandre, H.; Rousseaux, I.; Charpentier, C. Relationship between ethanol tolerance, lipid composition and plasma membrane fluidity in Saccharomyces cerevisiae and Kloeckera apiculata. FEMS Microbiol. Lett. 1994, 124, 17–22. [Google Scholar] [CrossRef]
- Novotný, C.; Flieger, M.; Panos, J.; Karst, F. Effect of 5,7-unsaturated sterols on ethanol tolerance in Saccharomyces cerevisiae. Biotechnol. Appl. Biochem. 1992, 15, 314–320. [Google Scholar]
- Tierney, K.J.; Block, D.E.; Longo, M.L. Elasticity and phase behavior of DPPC membrane modulated by cholesterol, ergosterol, and ethanol. Biophys. J. 2005, 89, 2481–2493. [Google Scholar] [CrossRef] [Green Version]
- Vanegas, J.M.; Contreras, M.F.; Faller, R.; Longo, M.L. Role of unsaturated lipid and ergosterol in ethanol tolerance of model yeast biomembranes. Biophys. J. 2012, 102, 507–516. [Google Scholar] [CrossRef] [Green Version]
- Vanegas, J.M.; Faller, R.; Longo, M.L. Influence of Ethanol on Lipid/Sterol Membranes: Phase Diagram Construction from AFM Imaging. Langmuir 2010, 26, 10415–10418. [Google Scholar] [CrossRef]
- Liu, G.; Chen, Y.; Færgeman, N.J.; Nielsen, J. Elimination of the last reactions in ergosterol biosynthesis alters the resistance of Saccharomyces cerevisiae to multiple stresses. FEMS Yeast Res. 2017, 17, fox063. [Google Scholar] [CrossRef]
- Inoue, T.; Iefuji, H.; Fujii, T.; Soga, H.; Satoh, K. Cloning and Characterization of a Gene Complementing the Mutation of an Ethanol-sensitive Mutant of Sake Yeast. Biosci. Biotechnol. Biochem. 2000, 64, 229–236. [Google Scholar] [CrossRef]
- López-Malo, M.; García-Ríos, E.; Chiva, R.; Guillamon, J.M. Functional analysis of lipid metabolism genes in wine yeasts during alcoholic fermentation at low temperature. Microb. Cell 2014, 1, 365–375. [Google Scholar] [CrossRef]
- Bisson, L.F. Stuck and Sluggish Fermentations. Am. J. Enol. Vitic. 1999, 50, 107–119. [Google Scholar]
- Torija, M.J.; Beltran, G.; Novo, M.; Poblet, M.; Guillamón, J.M.; Mas, A.; Rozès, N. Effects of fermentation temperature and Saccharomyces species on the cell fatty acid composition and presence of volatile compounds in wine. Int. J. Food Microbiol. 2003, 85, 127–136. [Google Scholar] [CrossRef]
- López-Malo, M. Metabolic and Molecular Adaptation of Wine Yeasts at Low Temperature Fermentation: Strategies for Their Genetic Improvement. Ph.D. Thesis, Universitat Rovira i Virgili, Tarragona, Spain, 2013. [Google Scholar]
- Tronchoni, J.; Rozès, N.; Querol, A.; Guillamón, J.M. Lipid composition of wine strains of Saccharomyces kudriavzevii and Saccharomyces cerevisiae grown at low temperature. Int. J. Food Microbiol. 2012, 155, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Casalta, E.; Vernhet, A.; Sablayrolles, J.M.; Tesnière, C.; Salmon, J.M. Review: Characterization and role of grape solids during alcoholic fermentation under enological conditions. Am. J. Enol. Vitic. 2016, 67, 133–138. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Vargas, S.; Sánchez-García, A.; Martínez-Rivas, J.M.; Prieto, J.A.; Randez-Gil, F. Fluidization of membrane lipids enhances the tolerance of Saccharomyces cerevisiae to freezing and salt stress. Appl. Environ. Microbiol. 2007, 73, 110–116. [Google Scholar] [CrossRef] [Green Version]
- Sablayrolles, J.M.; Barre, P. Evaluation of oxygen requirement of alcoholic fermentations under simulated oenological conditions. Sci. Aliment. 1986, 6, 373–383. [Google Scholar]
- Waldbauer, J.R.; Newman, D.K.; Summons, R.E. Microaerobic steroid biosynthesis and the molecular fossil record of Archean life. Proc. Natl. Acad. Sci. USA 2011, 108, 13409–13414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreasen, A.A.; Stier, T.J.B. Anaerobic nutrition of Saccharomyces cerevisiae. I. Ergosterol requirement for growth in a defined medium. J. Cell. Comp. Physiol. 1953, 41, 23–36. [Google Scholar] [CrossRef]
- Zavrel, M.; Hoot, S.J.; White, T.C. Comparison of sterol import under aerobic and anaerobic conditions in three fungal species, Candida albicans, Candida glabrata, and Saccharomyces cerevisiae. Eukaryot. Cell 2013, 12, 725–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zara, G.; Bardi, L.; Belviso, S.; Farris, G.A.; Zara, S.; Budroni, M. Correlation between cell lipid content, gene expression and fermentative behaviour of two Saccharomyces cerevisiae wine strains. J. Appl. Microbiol. 2008, 104, 906–914. [Google Scholar] [CrossRef]
- Cocito, C.; Delfini, C. Experiments for developing selective clarification techniques: Sterol and fatty acid loss from grape must related to clarification technique. J. Wine Res. 1997, 8, 187–197. [Google Scholar] [CrossRef]
- Soubeyrand, V.; Luparia, V.; Williams, P.; Doco, T.; Vernhet, A.; Ortiz-Julien, A.; Salmon, J.M. Formation of micella containing solubilized sterols during rehydration of active dry yeasts improves their fermenting capacity. J. Agric. Food Chem. 2005, 53, 8025–8032. [Google Scholar] [CrossRef] [PubMed]
- Sablayrolles, J.M.; Dubois, C.; Manginot, C.; Roustan, J.L.; Barre, P. Effectiveness of combined ammoniacal nitrogen and oxygen additions for completion of sluggish and stuck wine fermentations. J. Ferment. Bioeng. 1996, 82, 377–381. [Google Scholar] [CrossRef]
- Lambrechts, M.G.; Pretorius, I.S. Yeast and its Importance to Wine Aroma. Underst. Wine Chem. 2000, 21, 97–129. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, C.; Mendes-Faia, A.; Mendes-Ferreira, A. The nitrogen source impacts major volatile compounds released by Saccharomyces cerevisiae during alcoholic fermentation. Int. J. Food Microbiol. 2012, 160, 87–93. [Google Scholar] [CrossRef]
- Vilanova, M.; Ugliano, M.; Varela, C.; Siebert, T.; Pretorius, I.S.; Henschke, P.A. Assimilable nitrogen utilisation and production of volatile and non-volatile compounds in chemically defined medium by Saccharomyces cerevisiae wine yeasts. Appl. Microbiol. Biotechnol. 2007, 77, 145–157. [Google Scholar] [CrossRef] [Green Version]
- Molina, A.M.; Swiegers, J.H.; Varela, C.; Pretorius, I.S.; Agosin, E. Influence of wine fermentation temperature on the synthesis of yeast-derived volatile aroma compounds. Appl. Microbiol. Biotechnol. 2007, 77, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Rollero, S.; Mouret, J.R.; Sanchez, I.; Camarasa, C.; Ortiz-Julien, A.; Sablayrolles, J.M.; Dequin, S. Key role of lipid management in nitrogen and aroma metabolism in an evolved wine yeast strain. Microb. Cell Fact. 2016, 15, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Saerens, S.M.G.; Delvaux, F.; Verstrepen, K.J.; Van Dijck, P.; Thevelein, J.M.; Delvaux, F.R. Parameters affecting ethyl ester production by Saccharomyces cerevisiae during fermentation. Appl. Environ. Microbiol. 2008, 74, 454–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swiegers, J.; Bartowsky, E.; Henschke, P.A.; Pretorius, I. Yeast and bacterial modulation of wine aroma and flavour. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
- Verstrepen, K.J.; Van Laere, S.D.M.; Vanderhaegen, B.M.P.; Derdelinckx, G.; Dufour, J.-P.; Pretorius, I.S.; Winderickx, J.; Thevelein, J.M.; Delvaux, F.R. Expression levels of the yeast alcohol acetyltransferase genes ATF1, Lg-ATF1, and ATF2 control the formation of a broad range of volatile esters. Appl. Environ. Microbiol. 2003, 69, 5228–5237. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Petteys, B.J.; McClure, J.M.; Valsakumar, V.; Bekiranov, S.; Frank, E.L.; Smith, J.S. Thiamine biosynthesis in Saccharomyces cerevisiae is regulated by the NAD+-dependent histone deacetylase Hst1. Mol. Cell. Biol. 2010, 30, 3329–3341. [Google Scholar] [CrossRef] [Green Version]
- Mauricio, J.C.; Moreno, J.; Zea, L.; Ortega, J.M.; Medina, M. The effects of grape must fermentation conditions on volatile alcohols and esters formed by Saccharomyces cerevisiae. J. Sci. Food Agric. 1997, 75, 155–160. [Google Scholar] [CrossRef]
- Varela, C.; Torrea, D.; Schmidt, S.A.; Ancin-Azpilicueta, C.; Henschke, P.A. Effect of oxygen and lipid supplementation on the volatile composition of chemically defined medium and Chardonnay wine fermented with Saccharomyces cerevisiae. Food Chem. 2012, 135, 2863–2871. [Google Scholar] [CrossRef]
- Fairbairn, S.; Ferreira, A.C.S.; Bauer, F.F. Modulation of Yeast-Derived Volatile Aromas by Oleic Acid and Sterols. S. Afr. J. Enol. Vitic. 2019, 40, 1–11. [Google Scholar] [CrossRef]
- Guittin, C.; Maçna, F.; Sanchez, I.; Poitou, X.; Sablayrolles, J.M.; Mouret, J.R.; Farines, V. Impact of high lipid contents on the production of fermentative aromas during white wine fermentation. Appl. Microbiol. Biotechnol. 2021, 105, 6435–6449. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Kobayashi, O.; Yoshimoto, H.; Furukawa, S.; Tamai, Y. Effect of aeration and unsaturated fatty acids on expression of the Saccharomyces cerevisiae alcohol acetyltransferase gene. Appl. Environ. Microbiol. 1997, 63, 910–915. [Google Scholar] [CrossRef] [Green Version]
- Minetoki, T.; Bogaki, T.; Iwamatsu, A.; Fujii, T.; Hamachi, M. The purification, properties and internal peptide sequences of alcohol acetyltransferase isolated from Saccharomyces cerevisiae Kyokai No. 7. Biosci. Biotechnol. Biochem. 1993, 57, 2094–2098. [Google Scholar] [CrossRef]
- Dufour, J.; Malcorps, P.H.; Silcock, P. Control of Ester Synthesis During Brewery Fermentation. In Brewing Yeast Fermentation Performance; Wiley Online Library: Hoboken, NJ, USA, 2003; pp. 213–233. ISBN 9780470696040. [Google Scholar]
- Saerens, S.M.G.; Delvaux, F.R.; Verstrepen, K.J.; Thevelein, J.M. Production and biological function of volatile esters in Saccharomyces cerevisiae. Microb. Biotechnol. 2010, 3, 165–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, L.L.; Shi, Y.; Jiang, R.; Yang, Q.; Wang, Y.Q.; Liu, P.T.; Duan, C.Q.; Yan, G.L. Effects of adding unsaturated fatty acids composition of Saccharomyces cerevisiae and compounds in wine on fatty acid major volatile. S. Afr. J. Enol. Vitic. 2015, 36, 285–295. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.T.; Zhang, B.Q.; Duan, C.Q.; Yan, G.L. Pre-fermentative supplementation of unsaturated fatty acids alters the effect of overexpressing ATF1 and EEB1 on esters biosynthesis in red wine. LWT 2020, 120, 108925. [Google Scholar] [CrossRef]
- Plata, C.; Mauricio, J.C.; Millán, C.; Ortega, J.M. Influence of glucose and oxygen on the production of ethyl acetate and isoamyl acetate by a Saccharomyces cerevisiae strain during alcoholic fermentation. World J. Microbiol. Biotechnol. 2005, 21, 115–121. [Google Scholar] [CrossRef]
- Saharan, R.K.; Kanwal, S.; Sharma, S.C. Role of glutathione in ethanol stress tolerance in yeast Pachysolen tannophilus. Biochem. Biophys. Res. Commun. 2010, 397, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Roland, A.; Schneider, R.; Razungles, A.; Cavelier, F. Varietal Thiols in Wine: Discovery, Analysis and Applications. Chem. Rev. 2011, 111, 7355–7376. [Google Scholar] [CrossRef] [PubMed]
- Benkwitz, F.; Tominaga, T.; Kilmartin, P.A.; Lund, C.; Wohlers, M.; Nicolau, L. Identifying the chemical composition related to the distinct aroma characteristics of New Zealand Sauvignon blanc wines. Am. J. Enol. Vitic. 2012, 63, 62–72. [Google Scholar] [CrossRef]
- Landolfo, S.; Zara, G.; Zara, S.; Budroni, M.; Ciani, M.; Mannazzu, I. Oleic acid and ergosterol supplementation mitigates oxidative stress in wine strains of Saccharomyces cerevisiae. Int. J. Food Microbiol. 2010, 141, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Swiegers, J.; Pretorius, I.; Swiegers, J.H.; Pretorius, I.S. Modulation of volatile sulfur compounds by wine yeast. Appl. Microbiol. Biotechnol. 2007, 74, 954–960. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Eukaryotes | Mammals | Fungi | Plants |
|---|---|---|---|
| Sterol | Cholesterol | Ergosterol | |
| Zymosterol | β-sitosterol | ||
| Fecosterol | Stigmasterol | ||
| Episterol | Campesterol | ||
| Lanosterol |
| Higher Alcohols | Sterol with Positive Impact on Higher Alcohol Biosynthesis | References |
|---|---|---|
| Propanol | Ergosterol | [169,171] |
| 3-ethoxy-1-propanol | Ergosterol and phytosterols | [171] |
| Isoamyl alcohol | Ergosterol | [169,171] |
| Isoamyl alcohol | Phytosterols | [124,171] |
| 2-phenylethanol | Ergosterol | [169,170,171] |
| 2-phenylethanol | Phytosterols | [172] |
| Propanol and 2-phenylethanol | Phytosterols | [171] |
| 2-methylbutanol and 3-methylbutanol | Ergosterol | [170] |
| Isobutanol | Ergosterol | [169,170,171] |
| Isobutanol | Phytosterols | [124,171,172] |
| Esters | Sterol and Its Impact on Ester Biosynthesis | References | |
|---|---|---|---|
| Acetate esters | Isoamyl acetate | (+) Ergosterol and phytosterols | [171] |
| Ethyl acetate, isobutyl acetate, 2-methylbutyl acetate, isoamyl acetate and phenylethyl acetate | (+) Ergosterol | [170] | |
| Ethyl acetate, isobutyl acetate and isoamyl acetate | (−) Phytosterols | [164,172] | |
| Ethyl esters | Ethyl hexanoate and ethyl octanoate | (−) Phytosterols | [124,164] |
| Ethyl acetate | (+) Ergosterol and phytosterols | [171] | |
| Ethyl propanoate, ethyl butanoate, ethyl hexanoate, ethyl octanoate and ethyl decanoate | (+) Ergosterol | [170] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Girardi Piva, G.; Casalta, E.; Legras, J.-L.; Tesnière, C.; Sablayrolles, J.-M.; Ferreira, D.; Ortiz-Julien, A.; Galeote, V.; Mouret, J.-R. Characterization and Role of Sterols in Saccharomyces cerevisiae during White Wine Alcoholic Fermentation. Fermentation 2022, 8, 90. https://doi.org/10.3390/fermentation8020090
Girardi Piva G, Casalta E, Legras J-L, Tesnière C, Sablayrolles J-M, Ferreira D, Ortiz-Julien A, Galeote V, Mouret J-R. Characterization and Role of Sterols in Saccharomyces cerevisiae during White Wine Alcoholic Fermentation. Fermentation. 2022; 8(2):90. https://doi.org/10.3390/fermentation8020090
Chicago/Turabian StyleGirardi Piva, Giovana, Erick Casalta, Jean-Luc Legras, Catherine Tesnière, Jean-Marie Sablayrolles, David Ferreira, Anne Ortiz-Julien, Virginie Galeote, and Jean-Roch Mouret. 2022. "Characterization and Role of Sterols in Saccharomyces cerevisiae during White Wine Alcoholic Fermentation" Fermentation 8, no. 2: 90. https://doi.org/10.3390/fermentation8020090