
In Vitro Characterisation of Potential Probiotic Bacteria Isolated from a Naturally Fermented Carrot and Ginger Brine

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fermentation and Physicochemical Analysis of Carrot and Ginger Brine
pH and Titratable Acidity
2.2. Enumeration of Yeasts, Coliforms, and LAB
2.3. Isolation and Selection of LAB
2.4. Identification of LAB Using API 50 CHL Kits and 16S rDNA Sequencing
2.5. Assessment of Probiotic Potential of LAB
2.5.1. Tolerance to Low pH
2.5.2. Bile Salt Tolerance
2.5.3. Lysozyme Resistance
2.5.4. Cell Surface Hydrophobicity
2.5.5. Antibacterial Activity
2.5.6. Antioxidant Activity
Scavenging of Hydroxyl Radical with Intracellular Cell-Free Extract
Resistance to Hydrogen Peroxide
2.6. Statistical Analysis
3. Results and Discussion
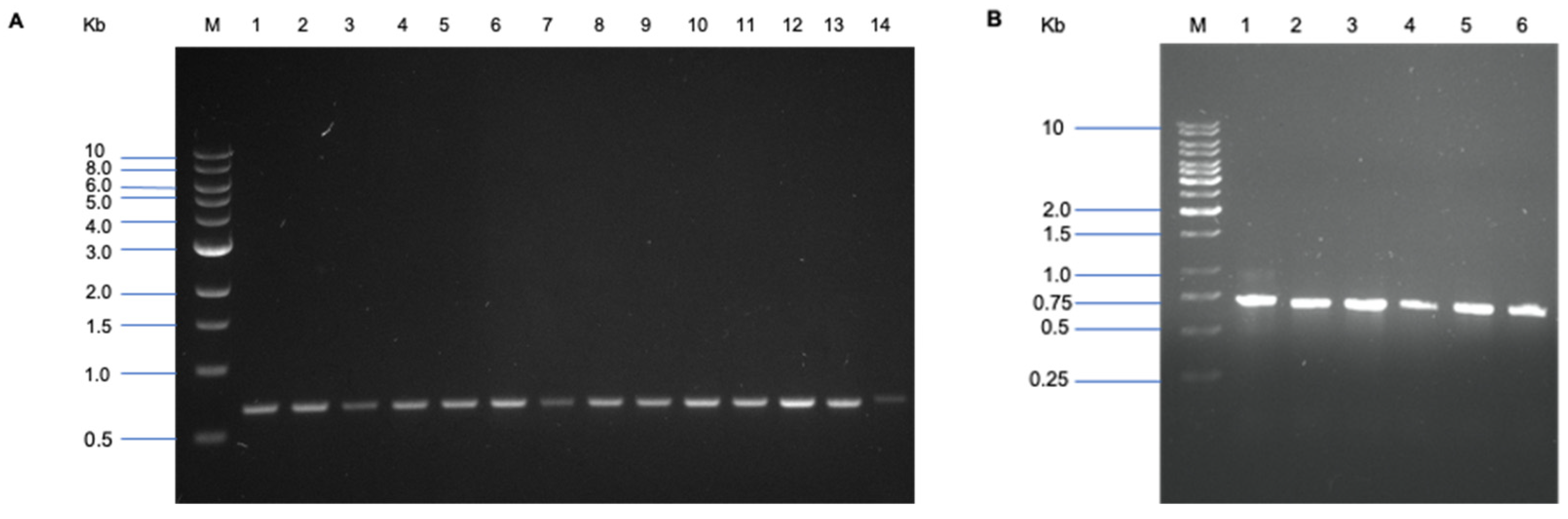
3.1. Genotypic Identification of the Isolates by 16S rDNA Sequencing
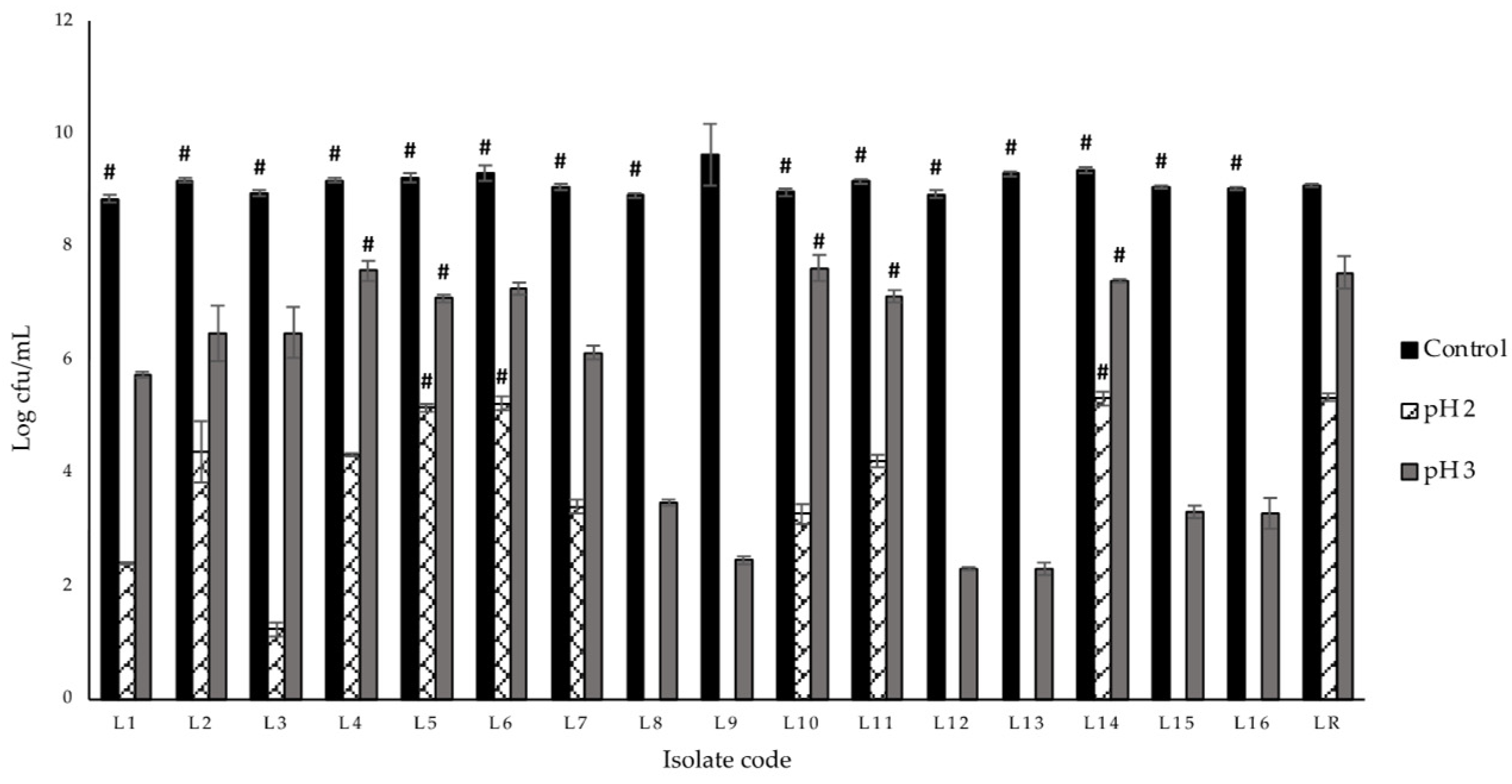
3.2. Acid Tolerance
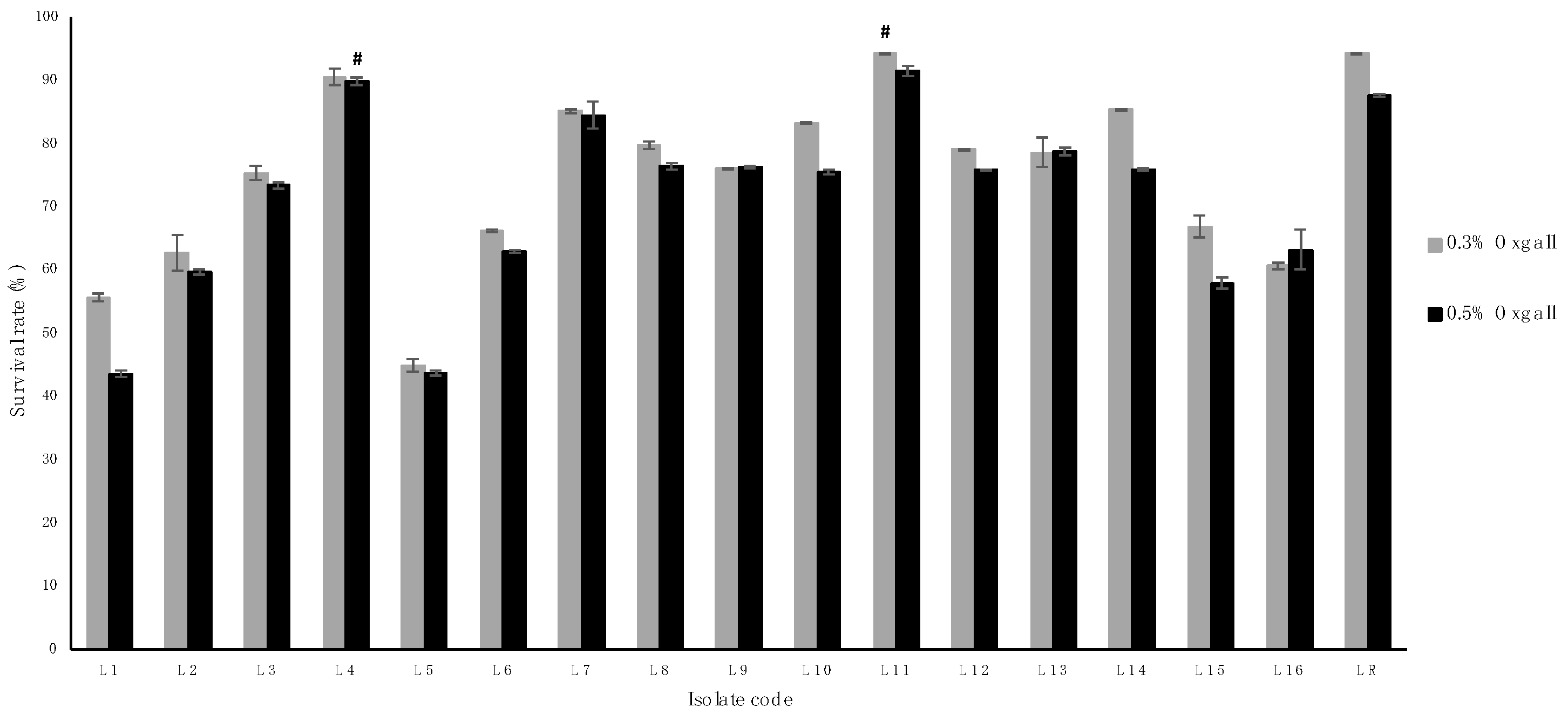
3.3. Bile Tolerance
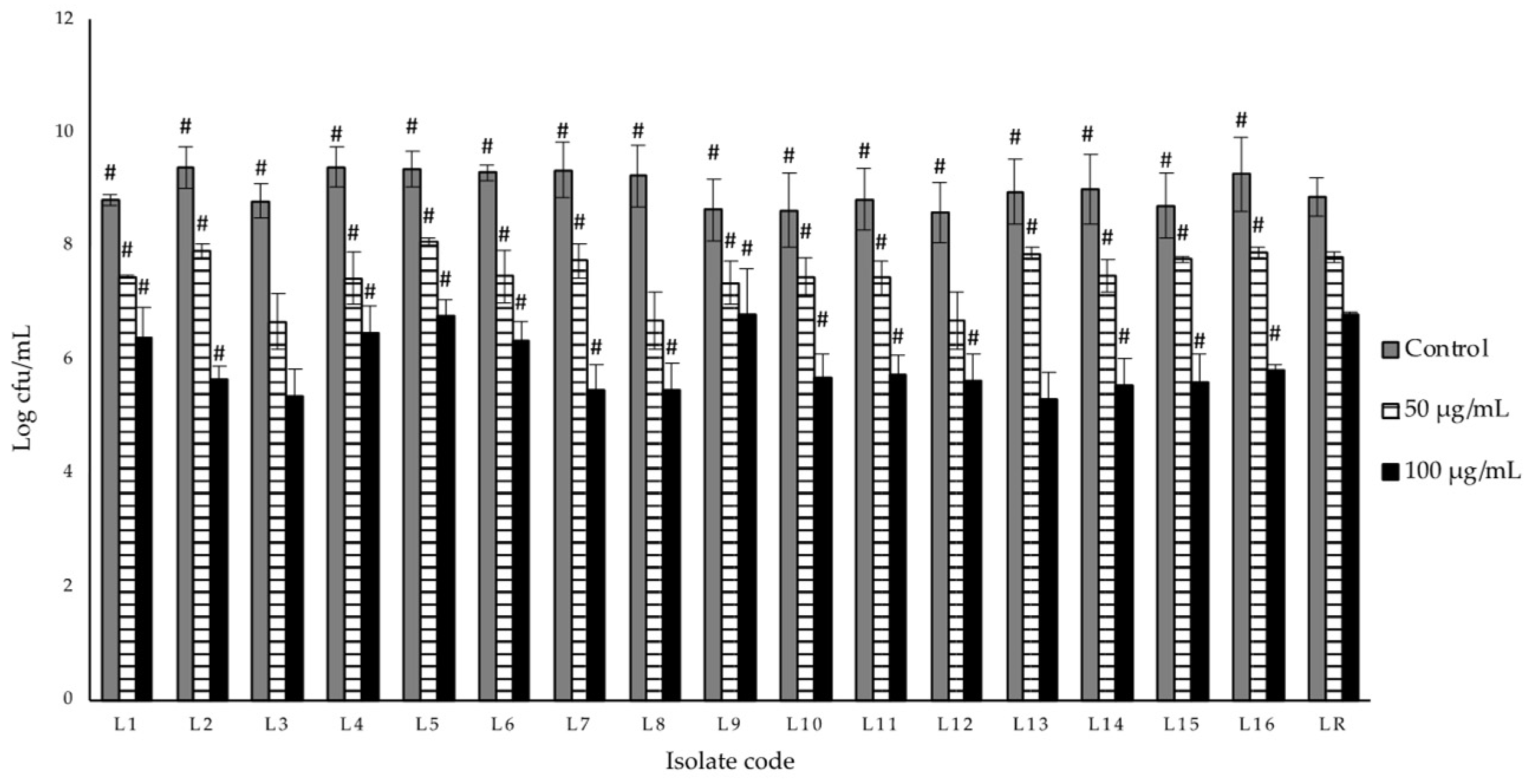
3.4. Lysozyme Resistance
3.5. Hydrophobicity
3.6. Antibacterial Activity Assay
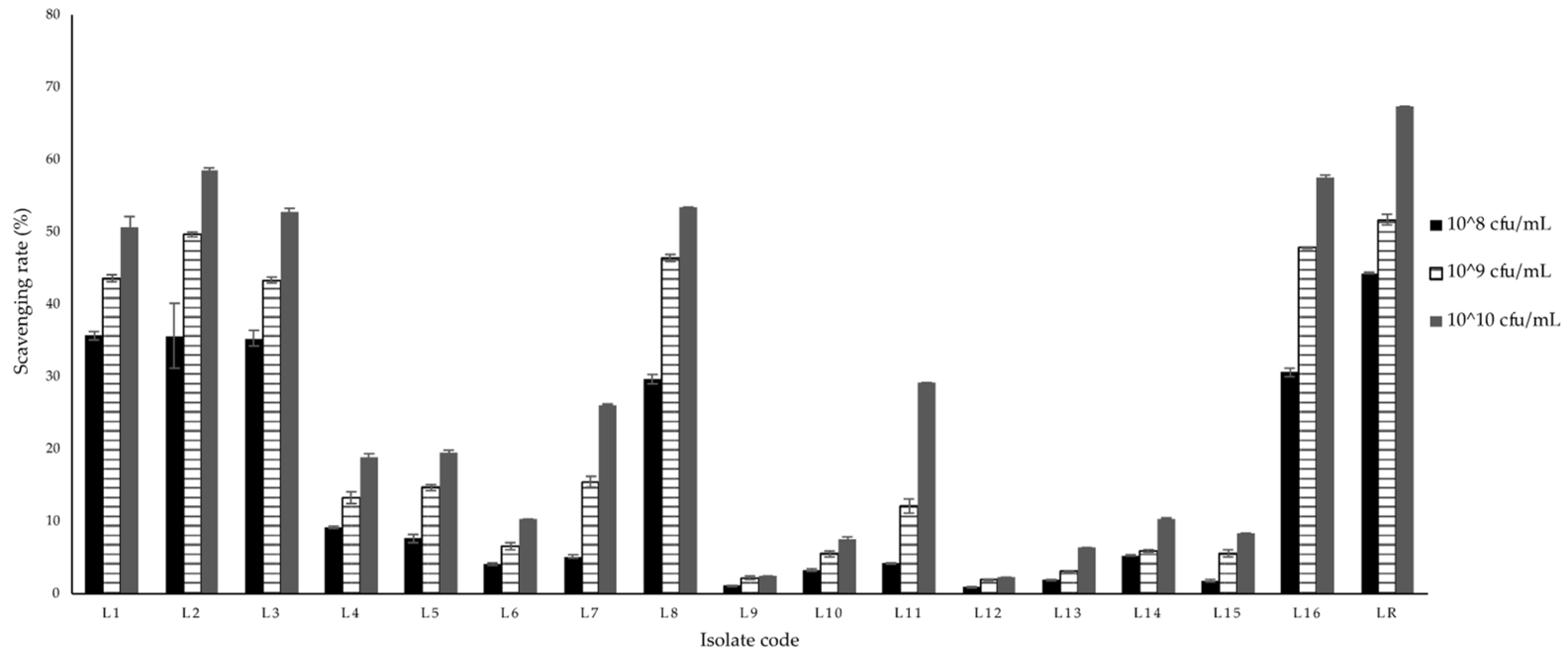
3.7. Antioxidant Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Caprara, G. Mediterranean-type dietary pattern and physical activity: The winning combination to counteract the rising burden of non-communicable diseases (NCDS). Nutrients 2021, 13, 429. [Google Scholar] [CrossRef]
- Swinburn, B.A.; Caterson, I.; Seidell, J.C.; James, W.P.T. Diet, nutrition and the prevention of excess weight gain and obesity. Public Health Nutr. 2004, 7, 123–146. [Google Scholar] [PubMed] [Green Version]
- Cencic, A.; Chingwaru, W. The role of functional foods, nutraceuticals, and food supplements in intestinal health. Nutrients 2010, 2, 611–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Sohaimy, S. Functional foods and nutraceuticals-modern approach to food science. World Appl. Sci. J. 2012, 20, 691–708. [Google Scholar]
- Mack, D.R. Probiotics: Mixed messages. Can. Fam. Physician 2005, 51, 1455. [Google Scholar] [PubMed]
- Holzapfel, W.H.; Haberer, P.; Geisen, R.; Björkroth, J.; Schillinger, U. Taxonomy and important features of probiotic microorganisms in food and nutrition. Am. J. Clin. Nutr. 2001, 73, 365s–373s. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.K.; Salminen, S. Handbook of Probiotics and Prebiotics; John Wiley & Sons: Hoboken, NJ, USA, 2009. [Google Scholar]
- Anadón, A.; Martínez-Larrañaga, M.R.; Caballero, V.; Castellano, V. Assessment of Prebiotics and Probiotics: An Overview, in Bioactive Foods in Promoting Health; Elsevier: Amsterdam, The Netherlands, 2010; pp. 19–41. [Google Scholar]
- Fuller, R. A review: Probiotics in man and animals. J. Appl. Bacteriol. 1989, 66, 365–378. [Google Scholar] [PubMed]
- Sarao, L.K.; Arora, M. Probiotics, prebiotics, and microencapsulation: A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 344–371. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.Z.; Ho, Y.W.; Abdullah, N.; Jalaludin, S. Acid and bile tolerance of Lactobacillus isolated from chicken intestine. Lett. Appl. Microbiol. 1998, 27, 183–185. [Google Scholar] [CrossRef] [PubMed]
- Masco, L.; Crockaert, C.; Hoorde, K.V.; Swings, J.; Huys, G. In vitro assessment of the gastrointestinal transit tolerance of taxonomic reference strains from human origin and probiotic product isolates of Bifidobacterium. J. Dairy Sci. 2007, 90, 3572–3578. [Google Scholar] [CrossRef] [Green Version]
- Tuomola, E.; Crittenden, R.; Playne, M.; Isolauri, E.; Salminen, S. Quality assurance criteria for probiotic bacteria. Am. J. Clin. Nutr. 2001, 73, 393s–398s. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turchi, B.; Mancini, S.; Fratini, F.; Pedonese, F.; Nuvoloni, R.; Bertelloni, F.; Ebani, V.V.; Cerri, D. Preliminary evaluation of probiotic potential of Lactobacillus plantarum strains isolated from Italian food products. World J. Microbiol. Biotechnol. 2013, 29, 1913–1922. [Google Scholar] [CrossRef] [PubMed]
- Del Carmen, S.; LeBlanc, A.d.M.d.; Levit, R.; Azevedo, V.; Langella, P.; Bermúdez-Humarán, L.G.; LeBlanc, J.G. Anti-cancer effect of lactic acid bacteria expressing antioxidant enzymes or IL-10 in a colorectal cancer mouse model. Int. Immunopharmacol. 2017, 42, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ringø, E.; Hoseinifar, S.H.; Lauzon, H.L.; Birkbeck, H.; Yang, D. The adherence and colonization of microorganisms in fish gastrointestinal tract. Rev. Aquac. 2019, 11, 603–618. [Google Scholar] [CrossRef]
- Gueimonde, M.; de los Reyes-Gavilán, C.; Sánchez, B. Stability of lactic acid bacteria in foods and supplements. Lactic Acid Bacteria. Microbiol. Funct. Asp. 2012, 361–384. [Google Scholar]
- Tripathi, M.K.; Giri, S.K. Probiotic functional foods: Survival of probiotics during processing and storage. J. Funct. Foods 2014, 9, 225–241. [Google Scholar] [CrossRef]
- Vasiljevic, T.; Shah, N.P. Probiotics—From Metchnikoff to bioactives. Int. Dairy J. 2008, 18, 714–728. [Google Scholar] [CrossRef]
- Lilly, D.M.; Stillwell, R.H. Probiotics: Growth-promoting factors produced by microorganisms. Science 1965, 147, 747–748. [Google Scholar] [CrossRef]
- Collins, J.; Thornton, G.; Sullivan, G. Selection of probiotic strains for human applications. Int. Dairy J. 1998, 8, 487–490. [Google Scholar] [CrossRef]
- Piqué, N.; Berlanga, M.; Miñana-Galbis, D. Health benefits of heat-killed (Tyndallized) probiotics: An overview. Int. J. Mol. Sci. 2019, 20, 2534. [Google Scholar] [CrossRef] [Green Version]
- Bogere, P.; Choi, Y.; Heo, J. Probiotics as alternatives to antibiotics in treating post-weaning diarrhoea in pigs. S. Afr. J. Anim. Sci. 2019, 49, 403–416. [Google Scholar] [CrossRef] [Green Version]
- Ouwehand, A.C.; Salminen, S.; Isolauri, E. Probiotics: An overview of beneficial effects. In Lactic Acid Bacteria: Genetics, Metabolism and Applications; Springer: Berlin/Heidelberg, Germany, 2002; pp. 279–289. [Google Scholar]
- Giraffa, G. Overview of the Ecology and Biodiversity of the LAB. In Lactic Acid Bacteria; John Wiley & Sons: Hoboken, NJ, USA, 2014; pp. 45–54. [Google Scholar]
- Cassani, L.; Gomez-Zavaglia, A.; Simal-Gandara, J. Technological strategies ensuring the safe arrival of beneficial microorganisms to the gut: From food processing and storage to their passage through the gastrointestinal tract. Food Res. Int. 2019, 129, 108852. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.; Shukla, P. An overview of advanced technologies for selection of probiotics and their expediency: A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 3233–3242. [Google Scholar] [CrossRef] [PubMed]
- De Corato, U. Improving the shelf-life and quality of fresh and minimally-processed fruits and vegetables for a modern food industry: A comprehensive critical review from the traditional technologies into the most promising advancements. Crit. Rev. Food Sci. Nutr. 2020, 60, 940–975. [Google Scholar] [CrossRef] [PubMed]
- Yildiz, F.; Wiley, R.C. Minimally Processed Refrigerated Fruits and Vegetables; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Tyl, C.; Sadler, G.D. pH and Titratable Acidity. In Food Analysis; Springer: Berlin/Heidelberg, Germany, 2017; pp. 389–406. [Google Scholar]
- Kavitha, J.R.; Devasena, T. Isolation, characterization, determination of probiotic properties of lactic acid bacteria from human milk. OSR J. Pharm. Biol. Sci. 2013, 7, 1–7. [Google Scholar] [CrossRef]
- Müller, T. Comparison of methods for differentiation between homofermentative and heterofermentative lactic acid bacteria. Zent. Mikrobiol. 1990, 145, 363–366. [Google Scholar] [CrossRef]
- Nelson, G.M.; George, S.E. Comparison of media for selection and enumeration of mouse fecal flora populations. J. Microbiol. Methods 1995, 22, 293–300. [Google Scholar] [CrossRef]
- Hedin, C.; Whelan, K.; Lindsay, J.O. Evidence for the use of probiotics and prebiotics in inflammatory bowel disease: A review of clinical trials. Proc. Nutr. Soc. 2007, 66, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.K.; Gilliland, S.E. Relationships among bile tolerance, bile salt deconjugation, and assimilation of cholesterol by Lactobacillus acidophilus. J. Dairy Sci. 1993, 76, 956–961. [Google Scholar] [CrossRef]
- Dias, R.; Vilas-Boas, E.; Campos, F.M.; Hogg, T.; Couto, J.A. Activity of lysozyme on Lactobacillus hilgardii strains isolated from Port wine. Food Microbiol. 2015, 49, 6–11. [Google Scholar] [CrossRef]
- Vinderola, C.G.; Reinheimer, J.A. Lactic acid starter and probiotic bacteria: A comparative “in vitro” study of probiotic characteristics and biological barrier resistance. Food Res. Int. 2003, 36, 895–904. [Google Scholar] [CrossRef]
- Mohankumar, A.; Murugalatha, N. Characterization and antibacterial activity of bacteriocin producing Lactobacillus isolated from raw cattle milk sample. Int. J. Biol. 2011, 3, 128. [Google Scholar] [CrossRef]
- Lin, M.Y.; Yen, C.L. Antioxidative ability of lactic acid bacteria. J. Agric. Food Chem. 1999, 47, 1460–1466. [Google Scholar] [CrossRef]
- He, Z.S.; Luo, H.; Cao, C.H.; Cui, Z.W. Photometric determination of hydroxyl free radical in Fenton system by brilliant green. Am. J. Chin. Med. 2004, 243, 236–237. [Google Scholar]
- Buchmeier, N.; Bossie, S.; Chen, C.Y.; Fang, F.C.; Guiney, D.G.; Libby, S.J. SlyA, a transcriptional regulator of Salmonella typhimurium, is required for resistance to oxidative stress and is expressed in the intracellular environment of macrophages. Infect. Immun. 1997, 65, 3725–3730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branquinho, R.; Meirinhos-Soares, L.; Carriço, J.A.; Pintado, M.; Peixe, L.V. Phylogenetic and clonality analysis of Bacillus pumilus isolates uncovered a highly heterogeneous population of different closely related species and clones. FEMS Microbiol. Ecol. 2014, 90, 689–698. [Google Scholar] [CrossRef] [Green Version]
- Trabelsi, I.; Bejar, W.; Ayadi, D.; Chouayekh, H.; Kammoun, R.; Bejar, S.; Salah, R.B. Encapsulation in alginate and alginate coated-chitosan improved the survival of newly probiotic in oxgall and gastric juice. Int. J. Biol. Macromol. 2013, 61, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Benmechernene, Z.; Chentouf, H.F.; Yahia, B.; Fatima, G.; Quintela-Baluja, M.; Calo-Mata, P.; Barros-Velázquez, J. Technological aptitude and applications of Leuconostoc mesenteroides bioactive strains isolated from Algerian raw camel milk. BioMed Res. Int. 2013, 2013, 418132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, P.K.; Mital, B.K.; Garg, S.K. Characterization of Lactobacillus acidophilus strains for use as dietary adjunct. Int. J. Food Microbiol. 1996, 29, 105–109. [Google Scholar] [CrossRef]
- Argyri, A.A.; Zoumpopoulou, G.; Karatzas, K.-A.G.; Tsakalidou, E.; Nychas, G.-J.E.; Panagou, E.Z.; Tassou, C.C. Selection of potential probiotic lactic acid bacteria from fermented olives by in vitro tests. Food Microbiol. 2013, 33, 282–291. [Google Scholar] [CrossRef]
- Yu, Z.; Zhang, X.; Li, S.; Li, C.; Li, D.; Yang, Z. Evaluation of probiotic properties of Lactobacillus plantarum strains isolated from Chinese sauerkraut. World J. Microbiol. Biotechnol. 2013, 29, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Dunne, C.; Murphy, L.; Flynn, S.; O’Mahony, L.; O’Halloran, S.; Feeney, M.; Morrissey, D.; Thornton, G.; Fitzgerald, G.; Daly, C.; et al. Probiotics: From myth to reality. Demonstration of functionality in animal models of disease and in human clinical trials. Lact. Acid Bact. Genet. Metab. Appl. 1999, 279–292. [Google Scholar]
- Hawaz, E. Isolation and identification of probiotic lactic acid bacteria from curd and in vitro evaluation of its growth inhibition activities against pathogenic bacteria. Afr. J. Microbiol. Res. 2014, 8, 1419–1425. [Google Scholar]
- Mishra, V.; Prasad, D.N. Application of in vitro methods for selection of Lactobacillus casei strains as potential probiotics. Int. J. Food Microbiol. 2005, 103, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Hamon, E.; Horvatovich, P.; Izquierdo, E.; Bringel, F.; Marchioni, E.; Aoudé-Werner, D.; Ennahar, S. Comparative proteomic analysis of Lactobacillus plantarumfor the identification of key proteins in bile tolerance. BMC Microbiol. 2011, 11, 63. [Google Scholar] [CrossRef] [PubMed]
- De Paula, A.T.; Jeronymo-Ceneviva, A.B.; Silva, L.F.; Todorov, S.D.; Franco, B.D.G.M.; Penna, A.L.B. Leuconostoc mesenteroides SJRP55: A potential probiotic strain isolated from Brazilian water buffalo mozzarella cheese. Ann. Microbiol. 2015, 65, 899–910. [Google Scholar] [CrossRef]
- Joel, T.J.; Suguna, S.; Steffi, S.R. Antimicrobial activity of lysozyme against oral pathogens. Asian J. Pharm. Res. Health Care 2016, 8, 42–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surono, I.S. In vitro probiotic properties of indigenous dadih lactic acid bacteria. Asian-Australas. J. Anim. Sci. 2003, 16, 726–731. [Google Scholar] [CrossRef]
- Pérez, P.F.; Minnaard, Y.; Disalvo, E.A.; Antoni, G.L.D. Surface properties of bifidobacterial strains of human origin. Appl. Environ. Microbiol. 1998, 64, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Raghavendra, P.; Halami, P.M. Screening, selection and characterization of phytic acid degrading lactic acid bacteria from chicken intestine. Int. J. Food Microbiol. 2009, 133, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Ouwehand, A.C.; Vesterlund, S. Antimicrobial components from lactic acid bacteria. Food Sci. Technol.-N. Y.-Marcel Dekker 2004, 139, 375–396. [Google Scholar]
- Aswathy, R.G.; Ismail, B.; John, R.P.; Nampoothiri, K.M. Evaluation of the probiotic characteristics of newly isolated lactic acid bacteria. Appl. Biochem. Biotechnol. 2008, 151, 244–255. [Google Scholar] [CrossRef] [PubMed]
- Woodmansey, E.J. Intestinal bacteria and ageing. J. Appl. Microbiol. 2007, 102, 1178–1186. [Google Scholar] [CrossRef] [PubMed]
- Abushelaibi, A.; Almahadin, S.; El-Tarabily, K.A.; Shah, N.P.; Ayyash, M. Characterization of potential probiotic lactic acid bacteria isolated from camel milk. LWT-Food Sci. Technol. 2017, 79, 316–325. [Google Scholar] [CrossRef]
- Ren, D.; Zhu, J.; Gong, S.; Liu, H.; Yu, H. Antimicrobial characteristics of lactic acid bacteria isolated from homemade fermented foods. BioMed Res. Int. 2018, 2018, 5416725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paula, A.T.d.; Jeronymo-Ceneviva, A.B.; Silva, L.F.; Todorov, S.D.; Franco, B.D.G.d.M.; Choiset, Y.; Haertlé, T.; Chobert, J.M.; Dousset, X.; Penna, A.L.B. Leuconostoc mesenteroides SJRP55: A bacteriocinogenic strain isolated from Brazilian water buffalo mozzarella cheese. Probiotics Antimicrob. Proteins 2014, 6, 186–197. [Google Scholar] [CrossRef]
- Li, L.; Shin, S.Y.; Lee, K.W.; Han, N.S. Production of natural antimicrobial compound d-phenyllactic acid using L euconostoc mesenteroides ATCC 8293 whole cells involving highly active d-lactate dehydrogenase. Lett. Appl. Microbiol. 2014, 59, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Ouwehand, A.C.; Salminen, S.J. The health effects of cultured milk products with viable and non-viable bacteria. Int. Dairy J. 1998, 8, 749–758. [Google Scholar] [CrossRef]
- Hazra, B.; Sarkar, R.; Biswas, S. Nripendranath Mandal Comparative study of the antioxidant and reactive oxygen species scavenging properties in the extracts of the fruits of Terminalia chebula, Terminalia belerica and Emblica officinalis. BMC Complement. Altern. Med. 2010, 10, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kao, T.H.; Chen, B.H. Functional components in soybean cake and their effects on antioxidant activity. J. Agric. Food Chem. 2006, 54, 7544–7555. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Hwang, T.K.; Chung, M.Y.; Cho, D.H.; Park, C.S. Resistance of Lactobacillus casei KCTC 3260 to reactive oxygen species (ROS): Role for a metal ion chelating effect. J. Food Sci. 2005, 70, m388–m391. [Google Scholar] [CrossRef]
- Kullisaar, T.; Zilmer, M.; Mikelsaar, M.; Vihalemm, T.; Annuk, H.; Kairane, C.; Kilk, A. Two antioxidative lactobacilli strains as promising probiotics. Int. J. Food Microbiol. 2002, 72, 215–224. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate Code | Phylogenetic Affiliation | Accession No. | Similarity (%) |
|---|---|---|---|
| L1 | Leuconostoc mesenteroides subsp. mesenteroides | NC008531.1 | 81.25 |
| L2 | Leuconostoc mesenteroides subsp. mesenteroides | NC008531.1 | 89.34 |
| L3 | Leuconostoc mesenteroides subsp. mesenteroides | NC008531.1 | 94.71 |
| L4 | Leuconostoc mesenteroides subsp. mesenteroides | NC008531.1 | 94.79 |
| L5 | Leuconostoc mesenteroides subsp. mesenteroides | NC008531.1 | 98.75 |
| L6 | Leuconostoc mesenteroides subsp. mesenteroides | NC008531.1 | 98.91 |
| L7 | Leuconostoc mesenteroides subsp. mesenteroides | NC008531.1 | 99.06 |
| L8 | Leuconostoc mesenteroides subsp. mesenteroides | NC008531.1 | 98.76 |
| L9 | Leuconostoc mesenteroides subsp. mesenteroides | NC008531.1 | 99.21 |
| L10 | Leuconostoc mesenteroides subsp. mesenteroides | NC008531.1 | 99.21 |
| L11 | Leuconostoc mesenteroides subsp. mesenteroides | NC008531.1 | 99.21 |
| L12 | Leuconostoc mesenteroides subsp. mesenteroides | NC008531.1 | 99.04 |
| L13 | Leuconostoc mesenteroides subsp. mesenteroides | NC008531.1 | 99.20 |
| L14 | Leuconostoc mesenteroides subsp. mesenteroides | NC008531.1 | 97.45 |
| L15 | Leuconostoc mesenteroides subsp. mesenteroides | NC008531.1 | 97.81 |
| L16 | Leuconostoc mesenteroides subsp. mesenteroides | NC008531.1 | 97.81 |
| L17 | Bacillus safensis | NZCP018191.1 | 97.50 |
| L18 | Bacillus licheniformis | NC006270.3 | 96.89 |
| Isolate Code | Hydrophobicity (%) |
|---|---|
| L1 | 45.64 ± 0.58 |
| L2 | 33.98 ± 2.65 |
| L3 | 83.68 ± 0.51 |
| L4 | 56.59 ± 4.59 |
| L5 | 54.58 ± 0.51 |
| L6 | 36.28 ± 0.16 |
| L7 | 36.11 ± 1.58 |
| L8 | 33.89 ± 5.32 |
| L9 | 36.74 ± 1.06 |
| L10 | 84.35 ± 1.85 |
| L11 | 33.18 ± 1.86 |
| L12 | 39.07 ± 0.07 |
| L13 | 38.30 ± 2.04 |
| L14 | 35.99 ± 1.18 |
| L15 | 54.93 ± 4.30 |
| L16 | 51.44 ± 0.87 |
| LR | 73.64 ± 1.12 |
| Isolate Code | Antimicrobial Activity | |
|---|---|---|
| Escherichia coli | Staphylococcus aureus | |
| L1 | ++ | +++ |
| L2 | +++ | +++ |
| L3 | + | +++ |
| L4 | ++ | ++ |
| L5 | +++ | +++ |
| L6 | +++ | +++ |
| L7 | +++ | +++ |
| L8 | +++ | +++ |
| L9 | +++ | +++ |
| L10 | +++ | +++ |
| L11 | +++ | +++ |
| L12 | ++ | ++ |
| L13 | +++ | +++ |
| L14 | + | +++ |
| L15 | ++ | +++ |
| L16 | +++ | +++ |
| LR | +++ | +++ |
| Isolate Code | Hydrogen Peroxide (mM) | |||
|---|---|---|---|---|
| 0 | 0.4 | 0.7 | 1.0 | |
| L1 | 2.58 ± 0.27 # | 2.16 ± 0.02 # | 1.41 ± 0.02 # | 0.72 ± 0.02 |
| L2 | 2.67 ± 0.24 # | 2.06 ± 0.06 # | 1.51 ± 0.05 # | 0.83 ± 0.05 # |
| L3 | 2.59 ± 0.25 # | 2.17 ± 0.11 # | 1.46 ± 0.01 # | 0.73 ± 0.02 |
| L4 | 2.54 ± 0.22 # | 1.86 ± 0.06 | 1.08 ± 0.03 | 0.53 ± 0.17 |
| L5 | 2.28 ± 0.06 # | 1.56 ± 0.08 | 0.99 ± 0.03 | 0.41 ± 0.02 |
| L6 | 2.28 ± 0.13 # | 1.56 ± 0.07 | 0.96 ± 0.01 | 0.34 ± 0.06 |
| L7 | 2.07 ± 0.05 | 1.93 ± 0.02 # | 1.34 ± 0.06 # | 0.64 ± 0.03 |
| L8 | 2.60 ± 0.26 | 2.05 ± 0.05 # | 1.42 ± 0.14 # | 0.74 ± 0.04 |
| L9 | 1.96 ± 0.06 | 1.48 ± 0.10 | 0.71 ± 0.23 | 0.37 ± 0.13 |
| L10 | 1.95 ± 0.07 | 1.50 ± 0.03 | 0.75 ± 0.22 | 0.33 ± 0.12 |
| L11 | 2.15 ± 0.04 | 1.92 ± 0.01 # | 1.39 ± 0.02 # | 0.60 ± 0.10 |
| L12 | 1.92 ± 0.07 | 1.52 ± 0.04 | 0.79 ± 0.16 | 0.25 ± 0.03 |
| L13 | 2.28 ± 0.03 | 1.72 ± 0.04 | 0.90 ± 0.09 | 0.32 ± 0.10 |
| L14 | 2.35 ± 0.04 | 1.75 ± 0.02 | 0.94 ± 0.13 | 0.37 ± 0.02 |
| L15 | 2.04 ± 0.02 | 1.76 ± 0.03 | 1.20 ± 0.11 | 0.39 ± 0.08 |
| L16 | 2.01 ± 0.03 | 1.90 ± 0.05 | 1.41 ± 0.03 # | 0.82 ± 0.14 # |
| LR | 2.68 ± 0.22 | 2.11 ± 0.13 | 1.71 ± 0.31 | 1.01 ± 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cele, N.; Nyide, B.; Khoza, T. In Vitro Characterisation of Potential Probiotic Bacteria Isolated from a Naturally Fermented Carrot and Ginger Brine. Fermentation 2022, 8, 534. https://doi.org/10.3390/fermentation8100534
Cele N, Nyide B, Khoza T. In Vitro Characterisation of Potential Probiotic Bacteria Isolated from a Naturally Fermented Carrot and Ginger Brine. Fermentation. 2022; 8(10):534. https://doi.org/10.3390/fermentation8100534
Chicago/Turabian StyleCele, Nontobeko, Babalwa Nyide, and Thandeka Khoza. 2022. "In Vitro Characterisation of Potential Probiotic Bacteria Isolated from a Naturally Fermented Carrot and Ginger Brine" Fermentation 8, no. 10: 534. https://doi.org/10.3390/fermentation8100534