
Optimization of Ethanolic Extraction of Enantia chloranta Bark, Phytochemical Composition, Green Synthesis of Silver Nanoparticles, and Antimicrobial Activity

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Material
2.1.1. Vegetal Materials
2.1.2. Microbial Strains and Their Profile of Susceptibility to Antibiotics
2.1.3. Galleria Mellonella
2.1.4. Chemicals
2.2. Method
2.2.1. Experimental Design for Extraction, Modelling, Validation of Model, and Optimization
2.2.2. HPLC-MS/MS Analysis
Sample Preparation
Analysis Conditions
2.3. Green Synthesis of Silver Nanoparticles
2.4. Characterization of Green-Synthetized AgNPs
2.5. Antimicrobial Activity
2.5.1. Inoculum Preparation
2.5.2. Preparation of Antimicrobial Solutions
2.5.3. Assessment of Antimicrobial Activity and Dose-Dependent Effect Using Well Diffusion Method
2.5.4. Determination of Minimum Inhibitory Concentration (MIC), Minimum Bactericidal Concentration (MBC), and Minimum Fungicidal Concentration (MFC)
2.5.5. Tolerance Level
2.5.6. Impact of Extract and AgNPs on Biofilm Formation
2.5.7. Acute Toxicity Test and In Vivo Antimicrobial Activity
2.5.8. Modulation of Common Antibiotics with Extract and AgNPs, and Antibiotic Resistance Reversal Activity
3. Results
3.1. Mathematical Modeling and Optimal Conditions for Ethanolic Extraction
3.1.1. Singular Influence of Factors on the Responses
3.1.2. Effect of Interactions between Factors on the Volume Yield, Mass Yield, and Inhibition Diameter of S. aureus ATCC 6538
3.1.3. Determination of Optimal Conditions
3.2. Phytochemical and Mineral Composition of the Optimized Extract (O-ECB)
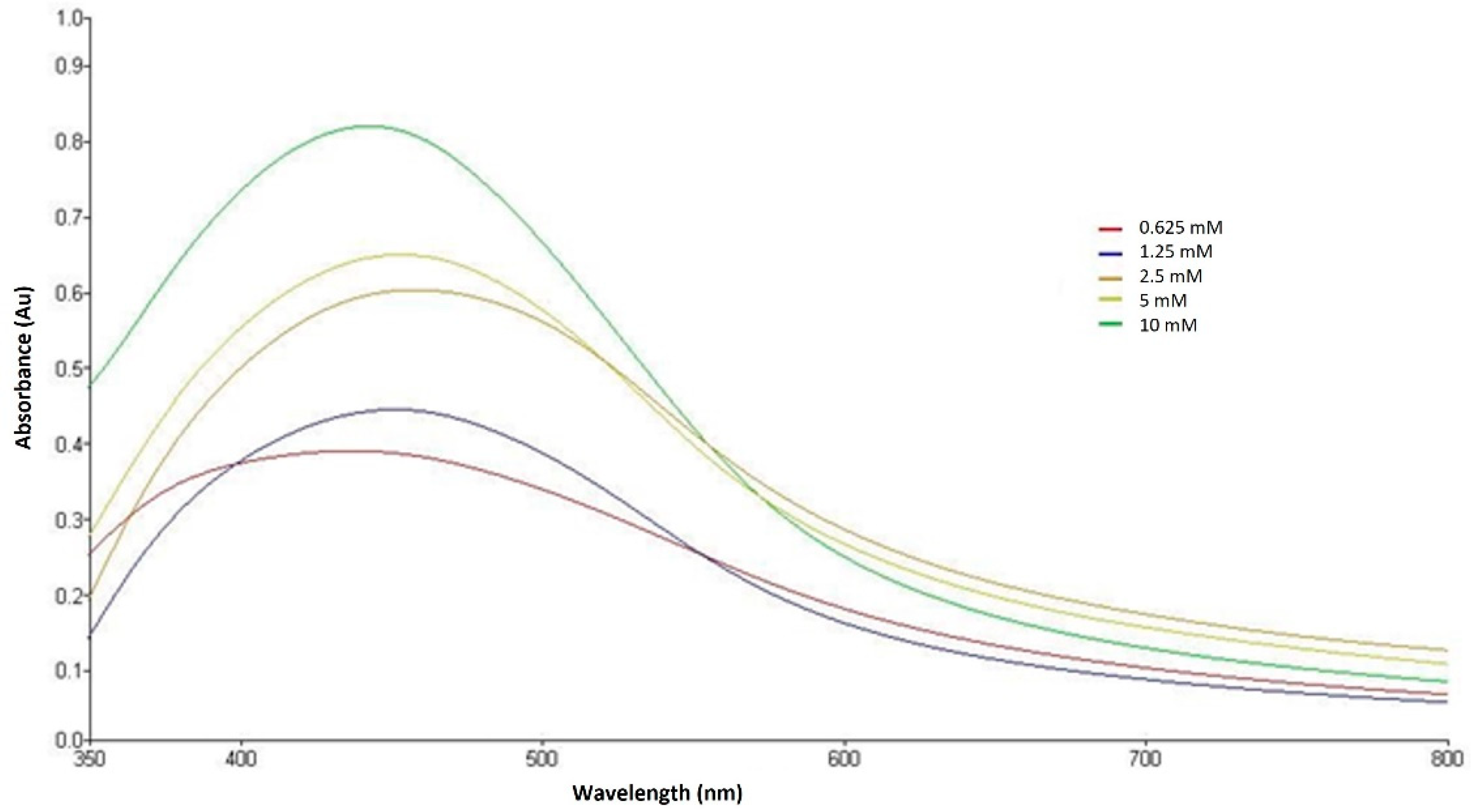
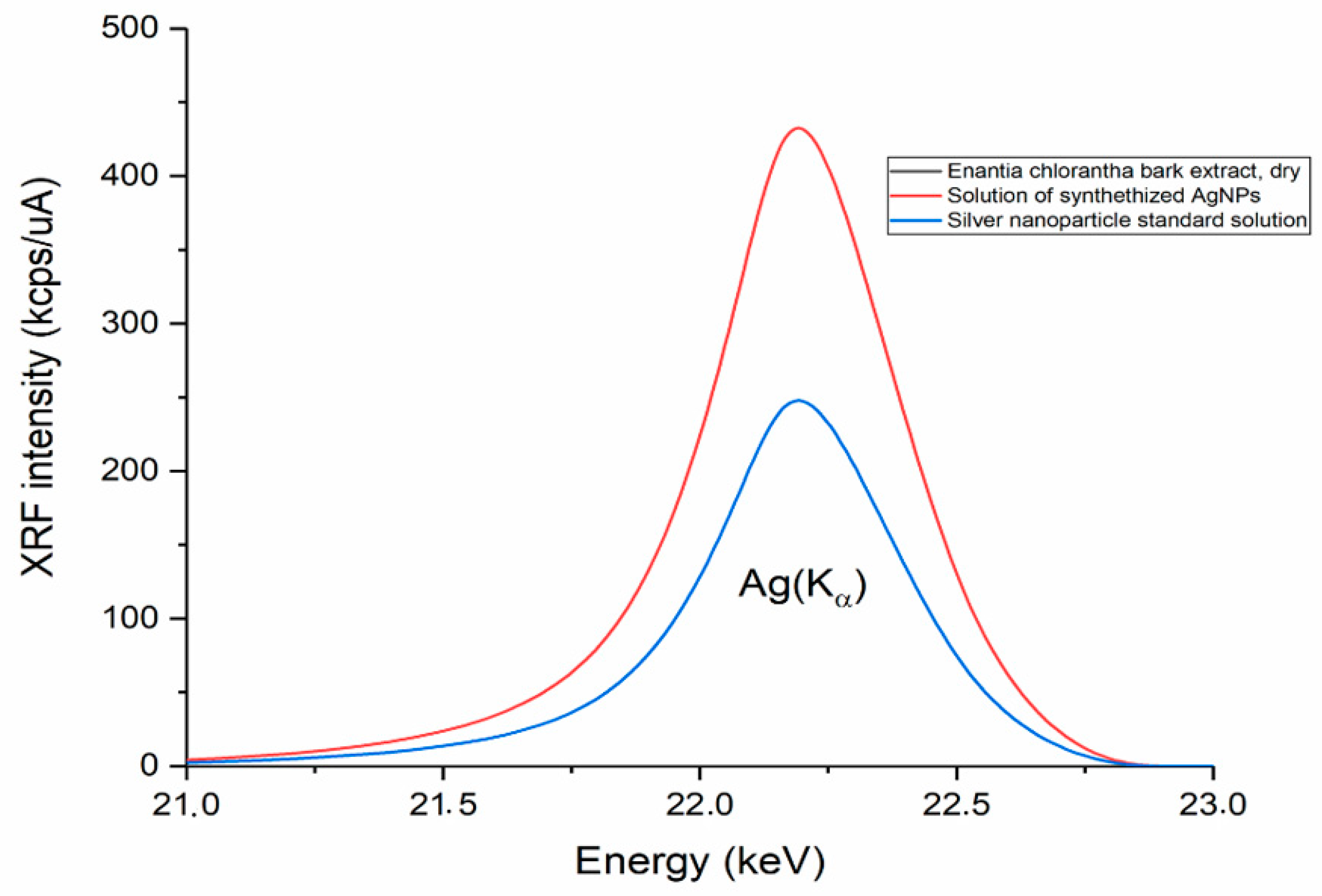
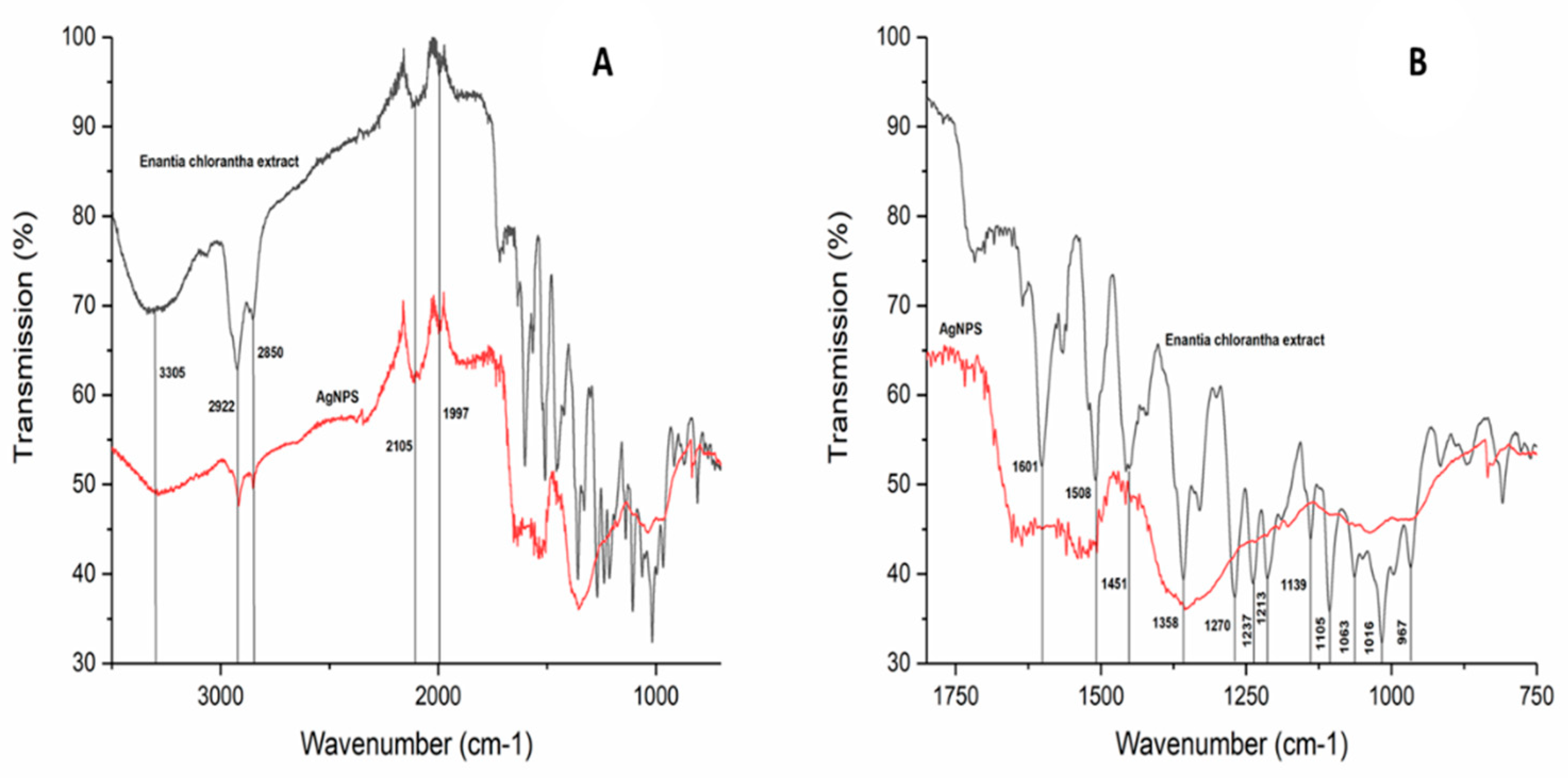
3.3. Silver Nanoparticles Green-Synthetized Using Enantia chlorantha Bark
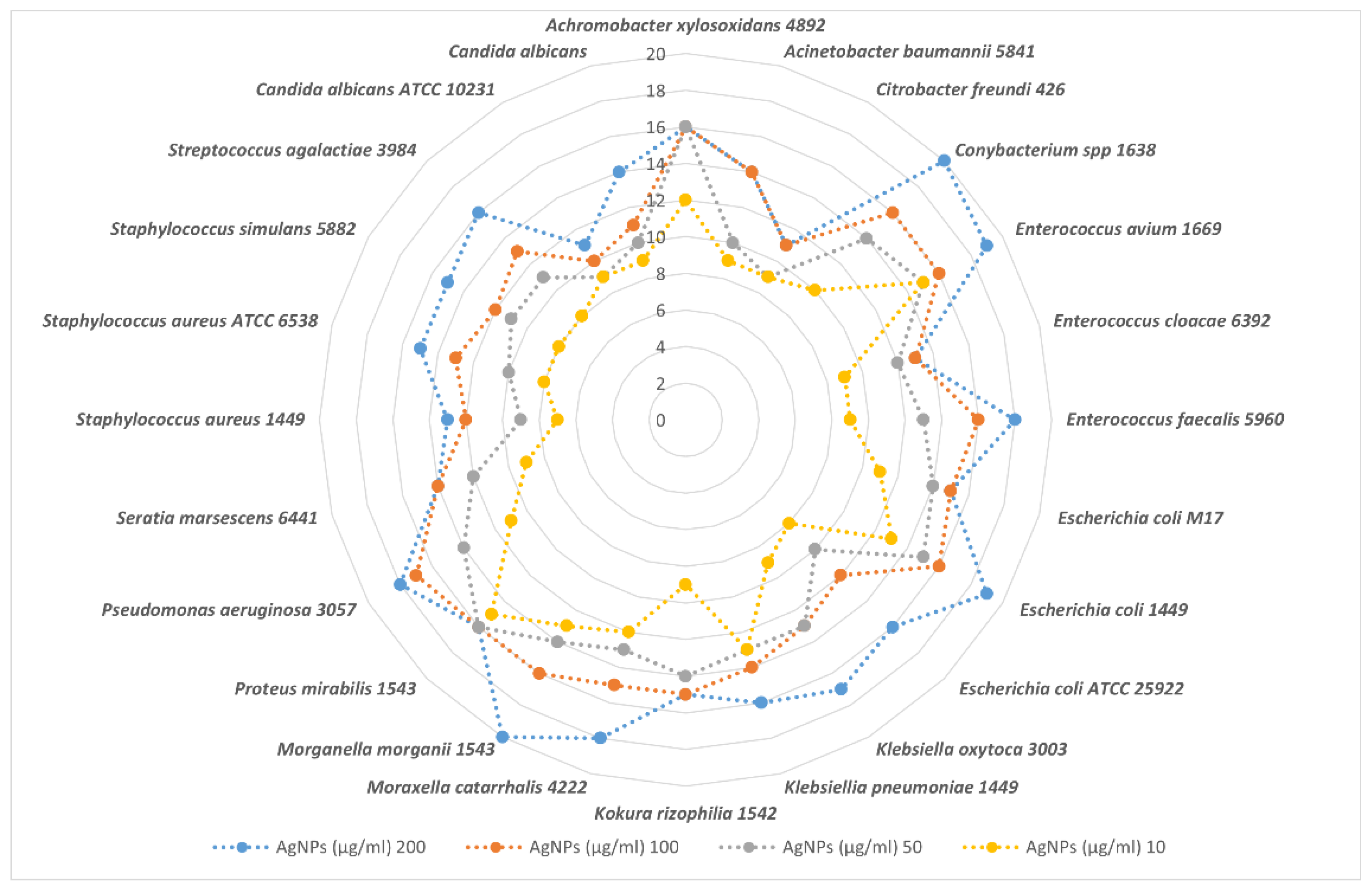
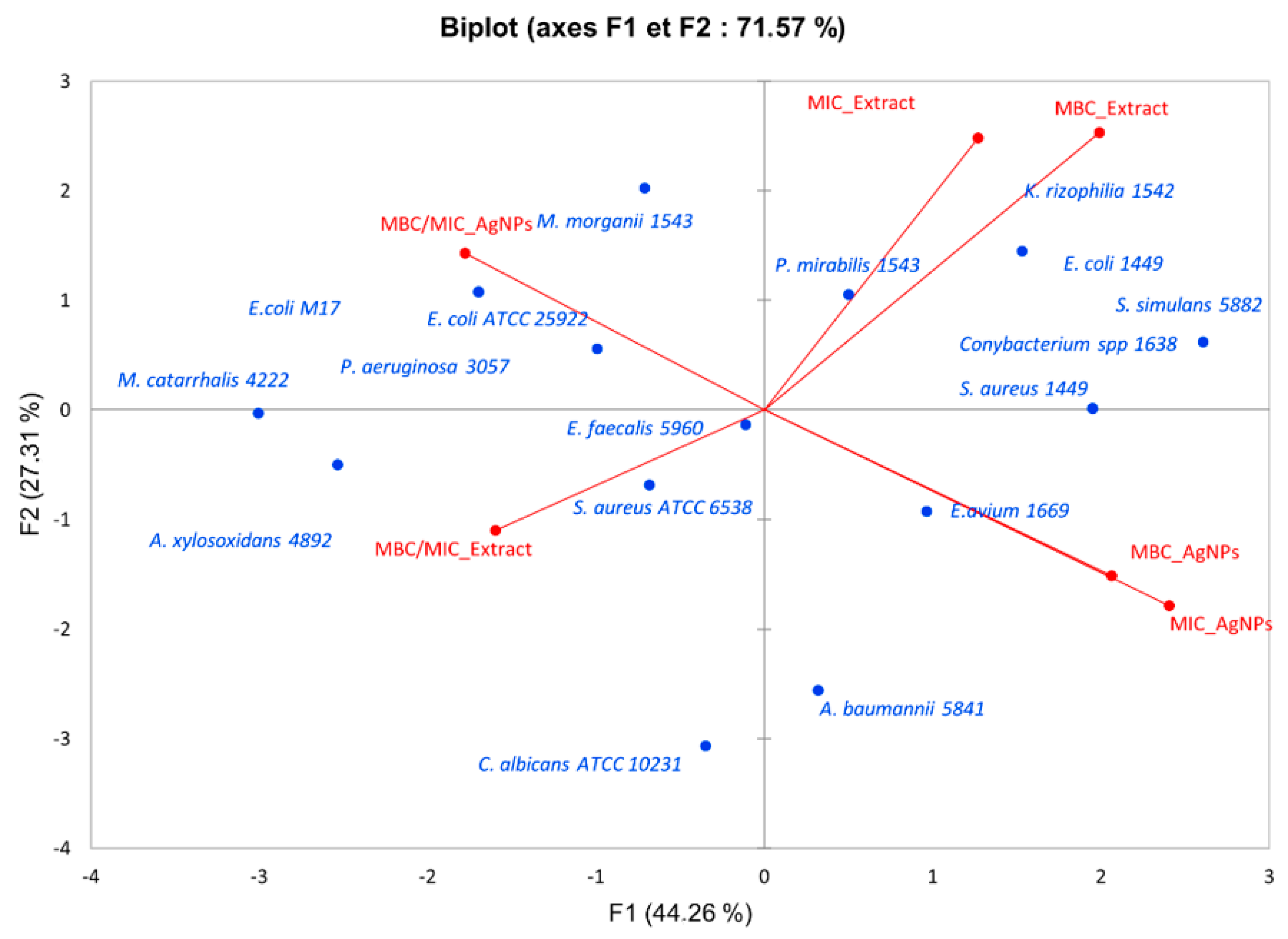
3.4. Antibacterial Activity of E. chlorantha Bark and Its AgNPs
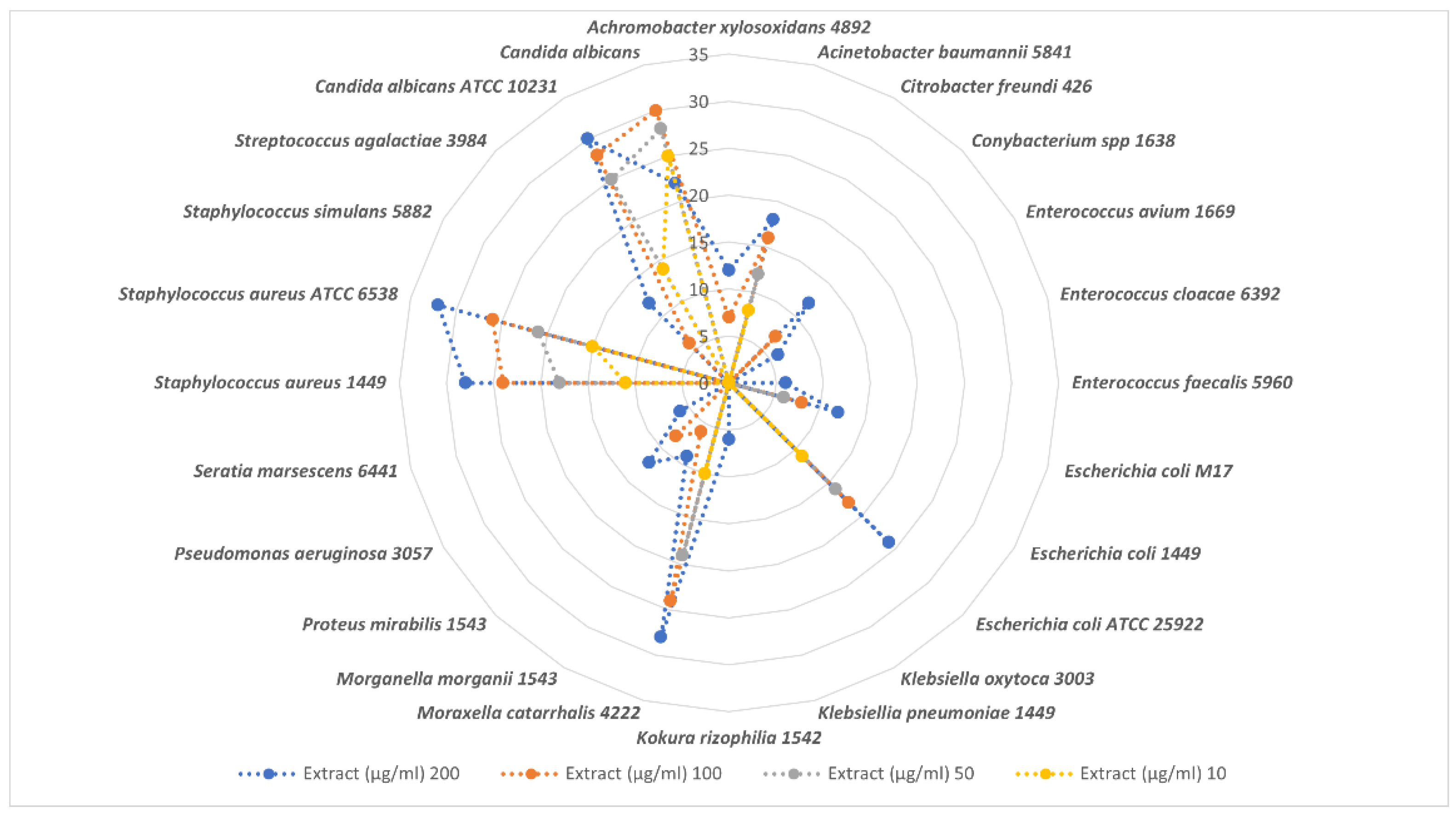
3.4.1. Dose-Dependent Effect, Minimum Inhibitory Concentration (MIC), Minimum Bactericidal Concentration (MBC), Minimum Fungicidal Concentration (MFC), and Ratio MBC/MIC
3.4.2. Antibiotic Resistance Reversal Activity
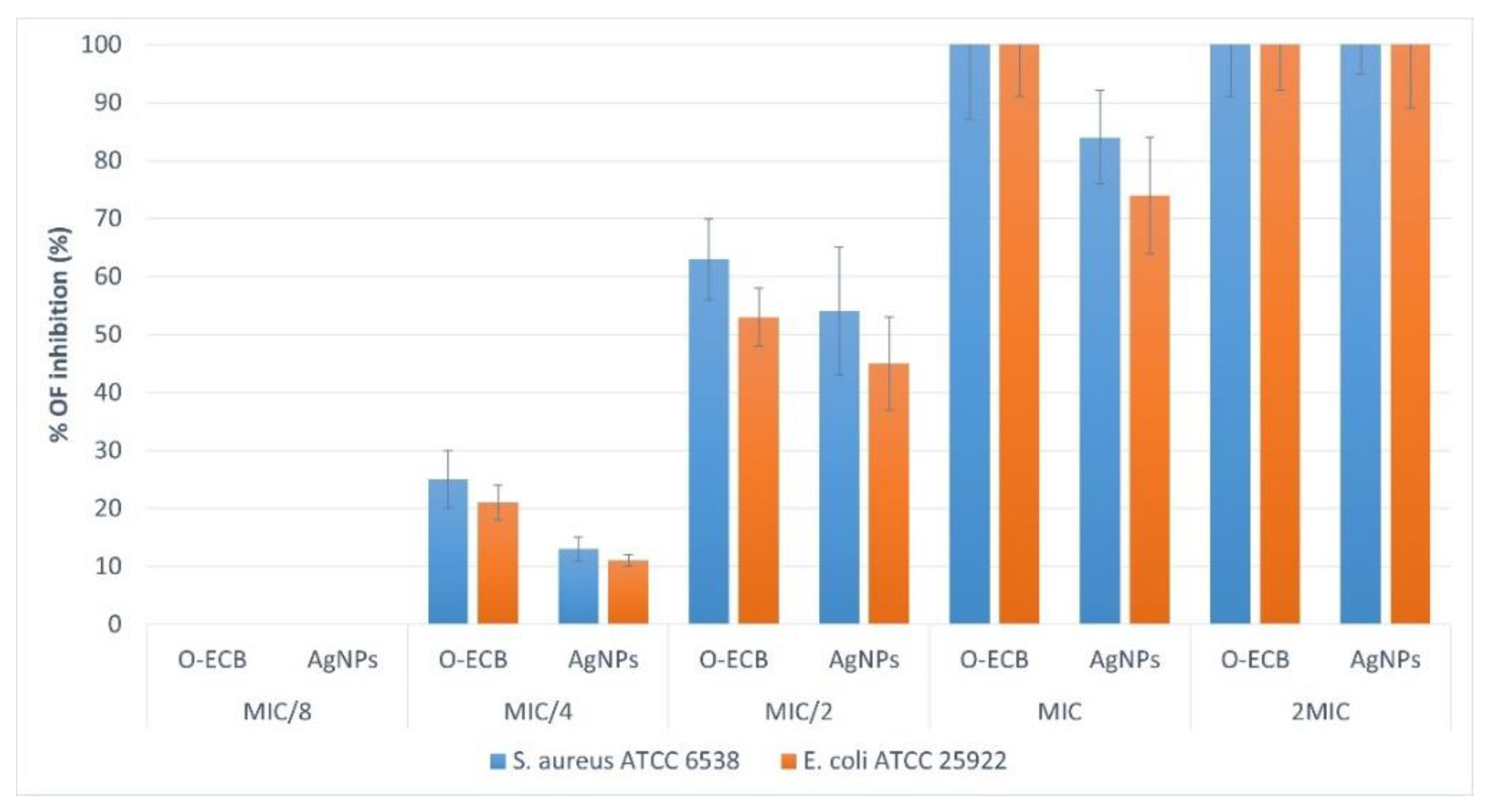
3.4.3. Impact of AgNPs and O-ECB on Biofilm Formation
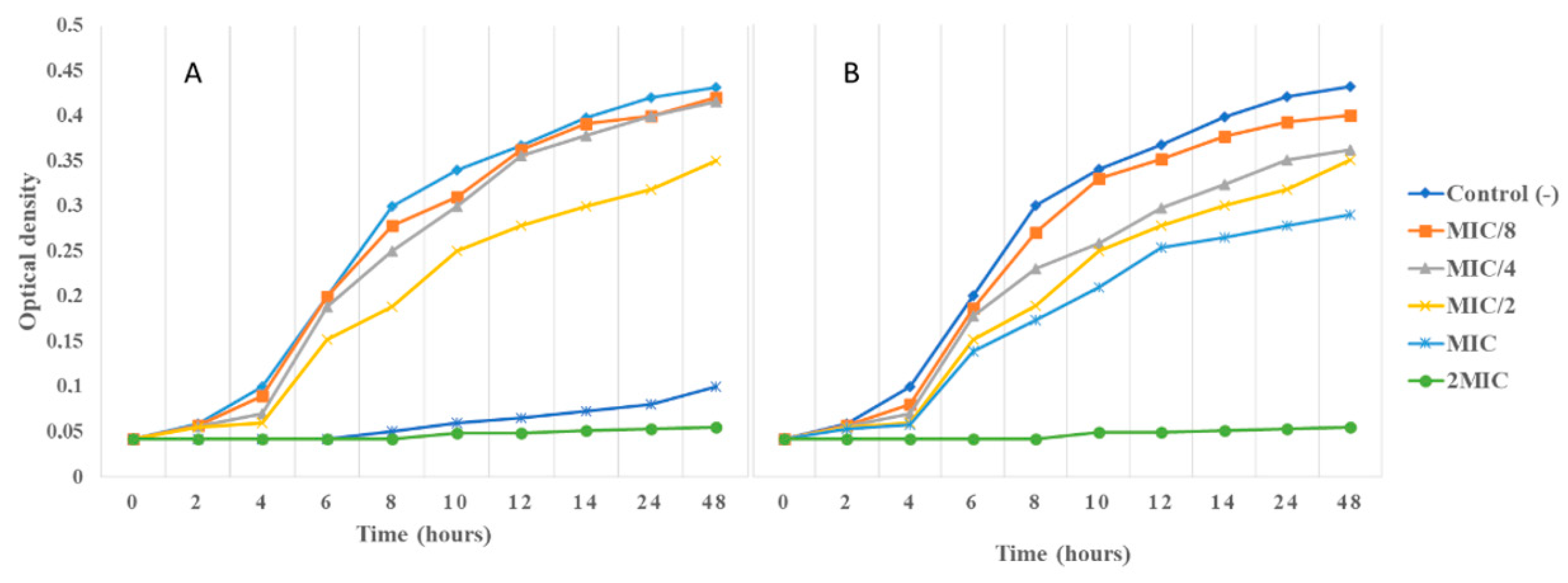
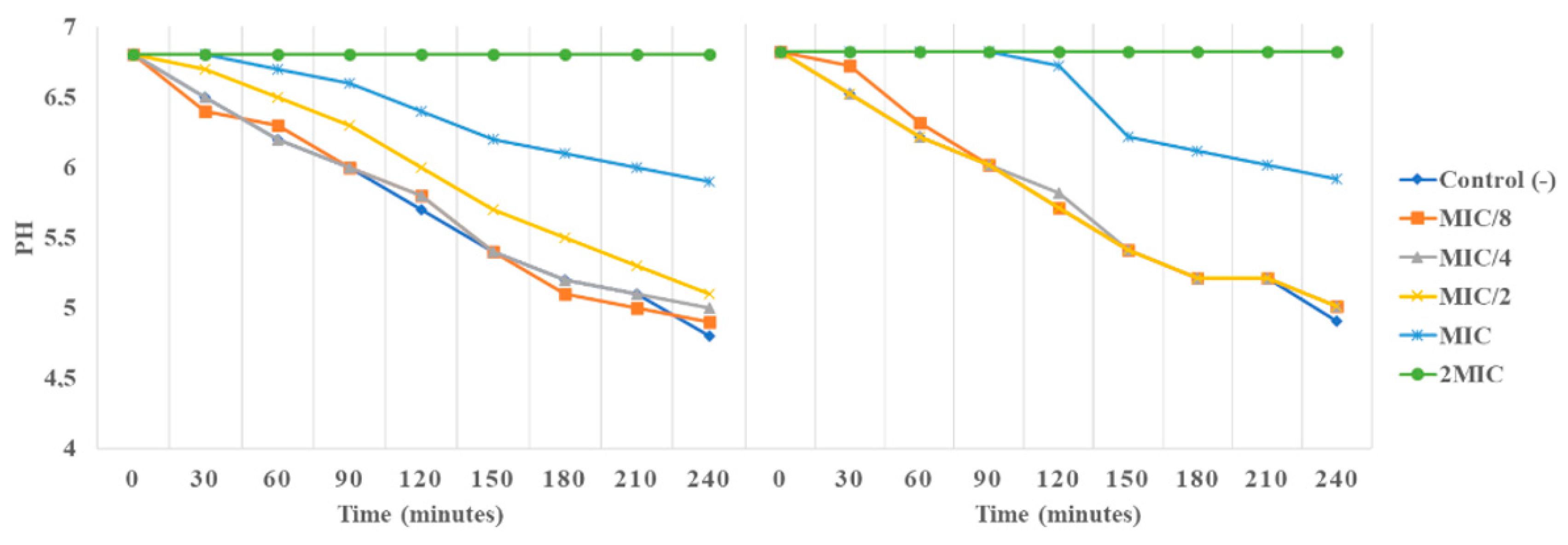
3.4.4. Impact of AgNPs and O-ECB on E. coli Growth Kinetic and H+-ATPase Proton Pumps
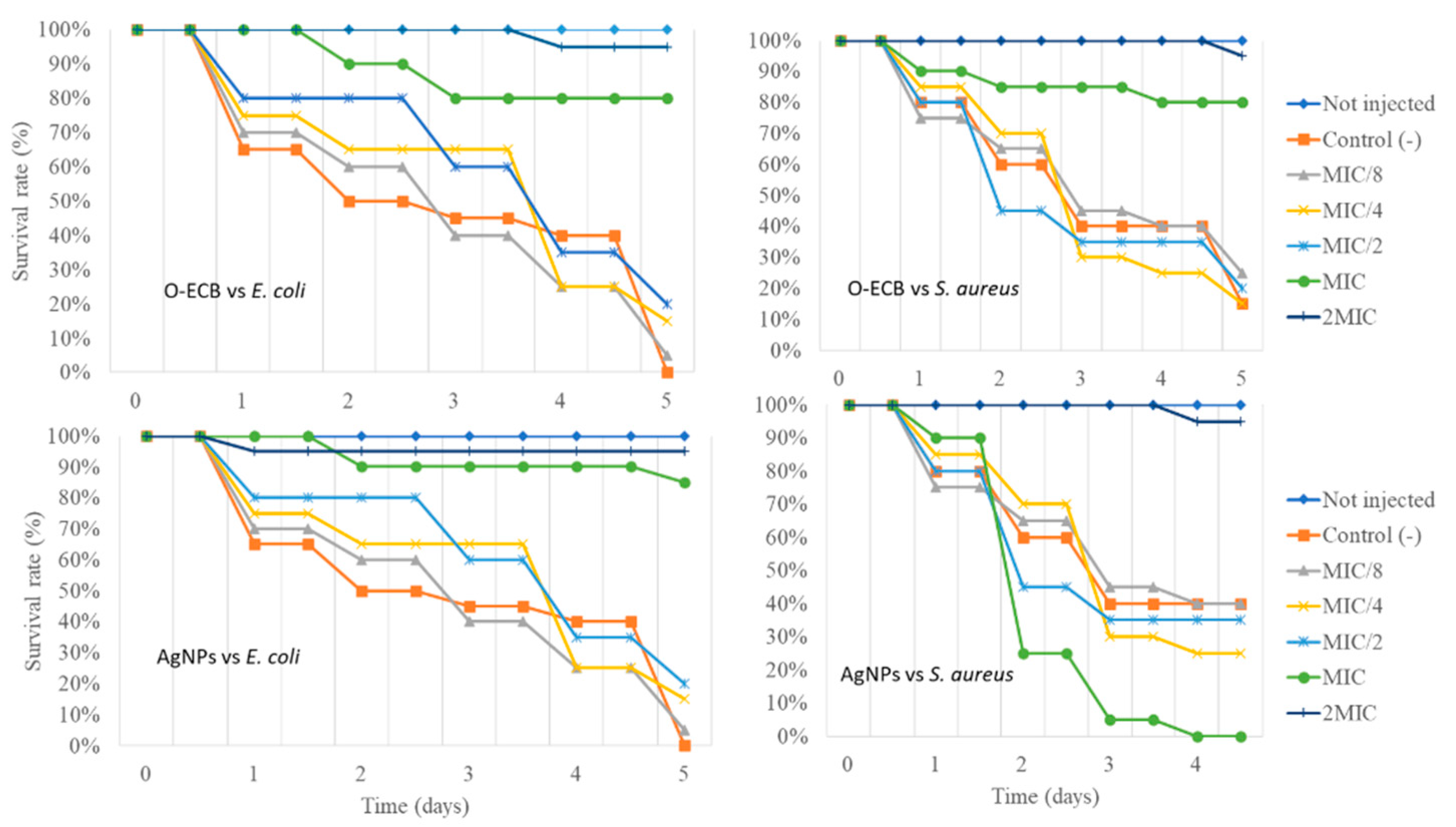
3.5. Toxicity Screen and In Vivo Antibacterial Activity of AgNPs and O-ECB
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tailor, G.; Yadav, B.L.; Chaudhary, J.; Joshi, M.; Suvalka, C. Green synthesis of silver nanoparticles using Ocimum canum and their anti-bacterial activity. Biochem. Biophys. Rep. 2020, 24, 100848. [Google Scholar] [CrossRef]
- Mostafa, M.; Kandile, N.G.; Mahmoud, M.K.; Ibrahim, H.M. Synthesis and characterization of polystyrene with embedded silver nanoparticle nanofibers to utilize as antibacterial and wound healing biomaterial. Heliyon 2022, 8, e08772. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.F.; Liu, Z.G.; Shen, W.; Gurunathan, S. Silver nanoparticles: Synthesis, characterization, properties, applications, and therapeutic approaches. Int. J. Mol. Sci. 2016, 17, 1534. [Google Scholar] [CrossRef] [PubMed]
- Shedrack, R.K.; Peter, K.; Judith, S.; Dominic, O.; Peter, M.; Naomi, M. Biogenic synthesis of silver nanoparticles using Azadirachta indica methanolic bark extract and their anti-proliferative activities against DU-145 human prostate cancer cells. Afr. J. Biotechnol. 2022, 21, 64–72. [Google Scholar]
- Chinnasamy, G.; Chandrasekharan, S.; Koh, T.W.; Bhatnagar, S. Synthesis, characterization, antibacterial and wound healing efficacy of silver nanoparticles from Azadirachta indica. Front. Microbiol. 2021, 12, 204. [Google Scholar] [CrossRef]
- Chandhirasekar, K.; Thendralmanikandan, A.; Thangavelu, P.; Nguyen, B.-S.; Nguyen, T.-A.; Sivashanmugan, K.; Nareshkumar, A.; Nguyen, V.-H. Plant-extract-assisted green synthesis and its larvicidal activities of silver nanoparticles using leaf extract of Citrus medica, Tagetes lemmonii, and Tarenna asiatica. Mater. Lett. 2021, 287, 129265. [Google Scholar] [CrossRef]
- Chakravarty, A.; Ahmad, I.; Singh, P.; Sheikh, M.U.D.; Aalam, G.; Sagadevan, S.; Ikram, S. Green synthesis of silver nanoparticles using fruits extracts of Syzygium cumini and their bioactivity. Chem. Phys. Lett. 2022, 795, 139493. [Google Scholar] [CrossRef]
- Cardoso-Avila, P.E.; Patakfalvi, R.; Rodríguez-Pedroza, C.; Aparicio-Fernández, X.; Loza-Cornejo, S.; Villa-Cruz, V.; Martínez-Cano, E. One-pot green synthesis of gold and silver nanoparticles using Rosa canina L. extract. RSC Adv. 2021, 11, 14624–14631. [Google Scholar] [CrossRef]
- Davares, A.K.L.; Arsene, M.M.J.; Viktorovna, P.I.; Shommya, D. Enantia chlorantha and its Multiple Therapeutic Virtues: A Mini Review. J. Pharm. Res. Int. 2021, 33, 254–259. [Google Scholar] [CrossRef]
- Etame, R.E.; Mouokeu, R.S.; Pouaha, C.L.C.; Kenfack, I.V.; Tchientcheu, R.; Assam, J.P.A.; Poundeu, F.S.M.; Tiabou, A.T.; Etoa, F.X.; Kuiate, J.R.; et al. Effect of fractioning on antibacterial activity of Enantia chlorantha Oliver (Annonaceae) methanol extract and mode of action. Evid. Based Complement. Altern. Med. 2018, 2018, 4831593. [Google Scholar] [CrossRef] [Green Version]
- Abike, T.O.; Osuntokun, O.T.; Modupe, A.O.; Adenike, A.F.; Atinuke, A.R. Antimicrobial Efficacy, Secondary Metabolite Constituents, Ligand Docking of Enantia chlorantha on Selected Multidrug Resistance Bacteria and Fungi. J. Adv. Biol. Biotechnol. 2020, 23, 17–32. [Google Scholar] [CrossRef]
- Olivier, D.K.; Van Vuuren, S.F.; Moteetee, A. Annickia affinis and A. chlorantha (Enantia chlorantha)—A review of two closely related medicinal plants from tropical Africa. J. Ethnopharmacol. 2015, 176, 438–462. [Google Scholar] [CrossRef] [PubMed]
- Arsene, M.M.J.; Podoprigora Irina, V.; Davares, A.K.L. Galleria mellonella (greater wax moth) as an eco-friendly in vivo approach for the assessment of the acute toxicity of medicinal plants: Application to some plants from Cameroon. Open Vet. J. 2021, 11, 651–666. [Google Scholar] [CrossRef] [PubMed]
- Faghri, J.; Dehbanipour, R.; Rastaghi, S.; Sedighi, M.; Maleki, N. High prevalence of multidrug-resistance uropathogenic Escherichia coli strains, Isfahan, Iran. J. Nat. Sci. Biol. Med. 2016, 7, 22–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arsene, M.M.J.; Zangue, D.S.C.; Ngoune, T.L.; Nyasha, K.; Louis, K. Antagonistic effects of raffia sap with probiotics against pathogenic microorganisms. Foods Raw Mater. 2021, 9, 24–31. [Google Scholar] [CrossRef]
- Motse, D.F.K.; Ngaba, G.P.; Koum, D.C.K.; Foko, L.P.K.; Ebongue, C.O.; Adiogo, D.D. Etiologic profile and sensitivity pattern of germs responsible for urinary tract infection among under-five children in Douala, Cameroon: A Hospital Based Study. Avicenna J. Clin. Microbiol. Infect. 2019, 6, 49–56. [Google Scholar] [CrossRef]
- Mbarga, M.J.A.; Davares, A.K.L.; Podoprigora Irina, V.; Smoliakova Larisa, A.; Suadkia, S.; Khelifi, I.; Sergueïevna, D.M. The public health issue of antibiotic residues in food and feed: Causes, consequences, and potential solutions. Vet. World 2022, 15, 662–671. [Google Scholar] [CrossRef]
- Arsène, M.M.J.; Podoprigora, I.V.; Davares, A.K.L.; Razan, M.; Das, M.S.; Senyagin, A.N. Antibacterial activity of grapefruit peel extracts and green-synthesized silver nanoparticles. Vet. World 2021, 14, 1330–1341. [Google Scholar] [CrossRef]
- Arsene, M.M.J.; Viktorovna, P.I.; Grigorievna, V.E.; Davares, A.K.L.; Sergeevna, D.M.; Nikolaevna, S.I. Prolonged exposure to antimicrobials induces changes in susceptibility to antibiotics, biofilm formation and pathogenicity in Staphylococcus aureus. J. Pharm. Res. Int. 2021, 33, 140–151. [Google Scholar] [CrossRef]
- Yuri, K.K.; Arsene, M.M.J.; Aliya, M.V.; Podoprigora Irina, V.; Volina Elena, G. Assessment of antimicrobial activity of ethanolic and aqueous extracts of Aesculus hippocastanum L. (horse chestnut) bark against bacteria isolated from urine of patients diagnosed positive to urinary tract infections. bioRxiv, 2021; in print. [Google Scholar] [CrossRef]
- Joseph, M.M.A.; Podoprigora Irina, V.; Davares, A.K.L.; Mouafo, H.T.; Manga, I.A.M.; Pavlovna, S.I.; Sergueïevna, D.M. Screening of antimicrobial activity of aqueous and ethanolic extracts of some medicinal plants from Cameroon and assessment of their synergy with common antibiotics against multidrug-resistant uropathogenic bacteria. bioRxiv, 2021; in print. [Google Scholar] [CrossRef]
- Mondal, A.H.; Yadav, D.; Mitra, S.; Mukhopadhyay, K. Biosynthesis of silver nanoparticles using culture supernatant of shewanella sp. Ary1 and their antibacterial activity. Int. J. Nanomed. 2020, 15, 8295–8310. [Google Scholar] [CrossRef]
- Berlutti, F.; Frioni, A.; Natalizi, T.; Pantanella, F.; Valenti, P. Influence of sub-inhibitory antibiotics and flow condition on Staphylococcus aureus ATCC 6538 biofilm development and biofilm growth rate: BioTimer assay as a study model. J. Antibiot. 2014, 67, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Habibipour, R.; Moradi-Haghgou, L.; Farmany, A. Green synthesis of AgNPs@ PPE and its Pseudomonas aeruginosa biofilm formation activity compared to pomegranate peel extract. Int. J. Nanomed. 2019, 14, 6891–6899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demgne, O.M.F.; Mbougnia, J.F.T.; Seukep, A.J.; Mbaveng, A.T.; Tene, M.; Nayim, P.; Wamba, B.E.N.; Guefack, M.-G.F.; Beng, V.P.; Tane, P.; et al. Antibacterial phytocomplexes and compounds from Psychotria sycophylla (Rubiaceae) against drug-resistant bacteria. Adv. Tradit. Med. 2021, 1–12. [Google Scholar] [CrossRef]
- Thitilertdecha, N.; Teerawutgulrag, A.; Rakariyatham, N. Antioxidant and antibacterial activities of Nephelium lappaceum L. extracts. LWT Food Sci. Technol. 2008, 41, 2029–2035. [Google Scholar] [CrossRef]
- Muala, W.C.B.; Desobgo, Z.S.C.; Jong, N.E. Optimization of extraction conditions of phenolic compounds from Cymbopogon citratus and evaluation of phenolics and aroma profiles of extract. Heliyon 2021, 7, e06744. [Google Scholar] [CrossRef]
- Onivogui, G.; Letsididi, R.; Diaby, M.; Wang, L.; Song, Y. Influence of extraction solvents on antioxidant and antimicrobial activities of the pulp and seed of Anisophyllea laurina R. Br. ex Sabine fruits. Asian Pac. J. Trop. 2015, 6, 20–25. [Google Scholar] [CrossRef] [Green Version]
- Mouafo, H.T.; Tchuenchieu, A.D.K.; Nguedjo, M.W.; Edoun, F.L.E.; Tchuente, B.R.T.; Medoua, G.N. In vitro antimicrobial activity of Millettia laurentii De Wild and Lophira alata Banks ex CF Gaertn on selected foodborne pathogens associated to gastroenteritis. Heliyon 2021, 7, e06830. [Google Scholar] [CrossRef]
- Evbuomwan, L.; Chukwuka, E.P.; Obazenu, E.I.; Ilevbare, L. Antibacterial Activity of Vernonia amygdalina Leaf Extracts against Multidrug Resistant Bacterial Isolates. J. Appl. Sci. Environ. Manag. 2018, 22, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, L.B. Antidiabetic Activity and Pharmacodynamic Interaction of Combined Administration of Ethanolic Stem Bark Extract of Enantia chlorantha and Lisinopril in Type 2 Diabetic Rats. Ph.D. Thesis, Kwara State University, Malete, Nigeria, 2019. Available online: https://www.proquest.com/openview/ebed11153ea830f88d5d0ba9dcc21f80/1?pq-origsite=gscholar&cbl=18750&diss=y (accessed on 28 June 2021).
- Ibrahim, L.B.; Idowu, P.F.; Moses, O.A.; Alabi, M.A.; Ajani, E.O. Antidiabetic Potential of Stem Bark Extract of Enantia chlorantha and Lack of Modulation of Its Therapeutic Efficacy in Diabetic Rats Co-Administered with Lisinopril. Acta Chim. Slov. 2021, 68, 118–127. [Google Scholar] [CrossRef]
- Priya, D.S.; Sankaravadivu, S.; Sudha, S.; Christy, H.K.S. Green synthesis and Characterisation of Silver Nanoparticles Using Phallusia nigra. Ann. Rom. Soc. Cell Biol. 2021, 25, 12948–12957. [Google Scholar]
- Gudimalla, A.; Jose, J.; Varghese, R.J.; Thomas, S. Green synthesis of silver nanoparticles using Nymphae odorata extract incorporated films and antimicrobial activity. J. Polym. Environ. 2021, 29, 1412–1423. [Google Scholar] [CrossRef]
- Vadlapudi, V.; Amanchy, R. Phytofabrication of silver nanoparticles using Myriostachya wightiana as a novel bioresource, and evaluation of their biological activities. Braz. Arch. Biol. Technol. 2017, 60, e17160329. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.W.; Shin, D.; Yang, M. Optical Property Change of Silver Nanowire Thin Films in Laser Patterning Process. Int. J. Precis. Eng. Manuf. 2020, 21, 301–308. [Google Scholar] [CrossRef]
- Scolaro, C.; Visco, A.; Torrisi, L. Laser welding of polymeric nanocomposites filled with silver nanoparticles produced by laser ablation. J. Instrum. 2020, 15, C02037. [Google Scholar] [CrossRef]
- Długosz, O.; Banach, M. Continuous synthesis of metal and metal oxide nanoparticles in microwave reactor. Colloids Surf. A Physicochem. Eng. 2020, 606, 125453. [Google Scholar] [CrossRef]
- Kumar, R.; Kumar, S.; Jewaria, P.K.; Devi, P. A study on green synthesis of silver nanoparticles using Murraya koenigii aqueous leaf extract. Int. J. Chem. Stud. 2020, 8, 2757–2759. [Google Scholar] [CrossRef]
- Obodovskiy, I. Radiation methods of matter composition analysis. In Radiation; Elsevier: Amsterdam, The Netherlands, 2019; pp. 293–301. [Google Scholar]
- Mittelman, A.M.; Fortner, J.D.; Pennell, K.D. Effects of ultraviolet light on silver nanoparticle mobility and dissolution. Environ. Sci. Nano. 2015, 2, 683–691. [Google Scholar] [CrossRef]
- Shankar, P.D.; Shobana, S.; Karuppusamy, I.; Pugazhendhi, A.; Ramkumar, V.S.; Arvindnarayan, S.; Kumar, G. A review on the biosynthesis of metallic nanoparticles (gold and silver) using bio-components of microalgae: Formation mechanism and applications. Enzym. Microb. Technol. 2016, 95, 28–44. [Google Scholar] [CrossRef]
- Pandey, S.; De Klerk, C.; Kim, J.; Kang, M.; Fosso-Kankeu, E. Eco friendly approach for synthesis, characterization and biological activities of milk protein stabilized silver nanoparticles. Polymers. 2020, 12, 1418. [Google Scholar] [CrossRef]
- Wang, L.; Hu, C.; Shao, L. The antimicrobial activity of nanoparticles: Present situation and prospects for the future. Int. J. Nanomed. 2017, 12, 1227. [Google Scholar] [CrossRef]
- Volleková, A.; Košt’álová, D.; Kettmann, V.; Tóth, J. Antifungal activity of Mahonia aquifolium extract and its major protoberberine alkaloids. Phytother. Res. 2003, 17, 834–837. [Google Scholar] [CrossRef] [PubMed]
- Khameneh, B.; Iranshahy, M.; Soheili, V.; Bazzaz, B.S.F. Review on plant antimicrobials: A mechanistic viewpoint. Antimicrob. Resist. Infect. Control. 2019, 8, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahamoud, A.; Chevalier, J.; Alibert-Franco, S.; Kern, W.V.; Pagès, J.M. Antibiotic efflux pumps in Gram-negative bacteria: The inhibitor response strategy. J. Antimicrob. Chemother. 2007, 59, 1223–1229. [Google Scholar] [CrossRef] [PubMed]
- Padilla, E.; Llobet, E.; Doménech-Sánchez, A.; Martínez-Martínez, L.; Bengoechea, J.A.; Albertí, S. Klebsiella pneumoniae AcrAB efflux pump contributes to antimicrobial resistance and virulence. Antimicrob. Agents Chemother. 2010, 54, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Ni, R.T.; Onishi, M.; Mizusawa, M.; Kitagawa, R.; Kishino, T.; Matsubara, F.; Tsuchiya, T.; Kuroda, T.; Ogawa, W. The role of RND-type efflux pumps in multidrug-resistant mutants of Klebsiella pneumoniae. Sci. Rep. 2020, 10, 10876. [Google Scholar] [CrossRef]
- Rai, M.; Kon, K.; Ingle, A.; Duran, N.; Galdiero, S.; Galdiero, M. Broad-spectrum bioactivities of silver nanoparticles: The emerging trends and future prospects. Appl. Microbiol. Biotechnol. 2014, 98, 1951–1961. [Google Scholar] [CrossRef]
- Sondi, I.; Salopek-Sondi, B. Silver nanoparticles as antimicrobial agent: A case study on E. coli as a model for Gram-negative bacteria. J. Colloid Interface Sci. 2004, 275, 177–182. [Google Scholar] [CrossRef]
- Banerjee, M.; Mallick, S.; Paul, A.; Chattopadhyay, A.; Ghosh, S.S. Heightened reactive oxygen species generation in the antimicrobial activity of a three components iodinated chitosan—Silver nanoparticle composite. Langmuir 2010, 26, 5901–5908. [Google Scholar] [CrossRef]
- Pal, S.; Tak, Y.K.; Song, J.M. Does the antibacterial activity of silver nanoparticles depend on the shape of the nanoparticle? A study of the gram-negative bacterium Escherichia coli. Appl. Environ. Microbiol. 2007, 73, 1712–1720. [Google Scholar] [CrossRef] [Green Version]
- Kuete, V. Potential of Cameroonian plants and derived products against microbial infections: A review. Planta Med. 2010, 76, 1479–1491. [Google Scholar] [CrossRef]
- Kuete, V.; Efferth, T. Cameroonian medicinal plants: Pharmacology and derived natural products. Front. Pharmacol. 2010, 1, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oussou, K.R.; Coffi, K.; Nathalie, G.S.; Gerard, K.; Mireille, D.; Yao, T.N.; Gille, F.; Jean-Claude, C.H. Activités antibactériennes des huiles essentielles de trois plantes aromatiques de Côte d’Ivoire. Comptes Rendus. Chim. 2008, 7, 1081–1086. [Google Scholar] [CrossRef]
- Teke, G.N.; Kuiate, J.R.; Kuete, V.; Teponno, R.B.; Tapondjou, L.A.; Tane, P.; Giacinti, G.; Vilarem, G. Bio guided isolation of potential antimicrobial and antioxidant agents from the stem bark of Trilepisium madagascariense. S. Afr. J. Bot. 2011, 77, 319–327. [Google Scholar] [CrossRef] [Green Version]
- Ngongang, F.C.M.; Fankam, A.G.; Mbaveng, A.T.; Wamba, B.E.N.; Nayim, P.; Beng, V.P.; Kuete, V. Methanol extracts from Manilkara zapota with moderate antibacterial activity displayed strong antibiotic-modulating effects against multidrug-resistant phenotypes. Pharmacology 2020, 3, 37. [Google Scholar] [CrossRef]
- Benelli, G.; Maggi, F.; Petrelli, R.; Canale, A.; Nicoletti, M.; Rakotosaona, R.; Rasoanaivo, P. Not ordinary antimalarial drugs: Madagascar plant decoctions potentiating the chloroquine action against Plasmodium parasites. Ind. Crops Prod. 2017, 103, 19–38. [Google Scholar] [CrossRef]
- Casciaro, B.; Mangiardi, L.; Cappiello, F.; Romeo, I.; Loffredo, M.R.; Iazzetti, A.; Calcaterra, A.; Goggiamani, A.; Ghirga, F.; Mangoni, M.L.; et al. Naturally-occurring alkaloids of plant origin as potential antimicrobials against antibiotic-resistant infections. Molecules 2020, 25, 3619. [Google Scholar] [CrossRef]
- Kumar, P.; Srivastava, V.; Chaturvedi, R.; Sundar, D.; Bisaria, V.S. Elicitor enhanced production of protoberberine alkaloids from in vitro cell suspension cultures of Tinospora cordifolia (Willd.) Miers ex Hook. F. Thoms. Plant Cell Tissue Organ Cult. 2017, 130, 417–426. [Google Scholar] [CrossRef]
- Alam, K.; Al Farraj, D.A.; Mah-E-Fatima, S.; Yameen, M.A.; Elshikh, M.S.; Alkufeidy, R.M.; Mustafa, A.E.-Z.M.; Bhasme, P.; Alshammari, M.K.; Alkubaisi, N.A.; et al. Anti-biofilm activity of plant derived extracts against infectious pathogen-Pseudomonas aeruginosa PAO1. J. Infect. Public Health 2020, 13, 1734–1741. [Google Scholar] [CrossRef]
- Martinez-Gutierrez, F.; Boegli, L.; Agostinho, A.; Sánchez, E.M.; Bach, H.; Ruiz, F.; James, G. Anti-biofilm activity of silver nanoparticles against different microorganisms. Biofouling 2013, 29, 651–660. [Google Scholar] [CrossRef]
- Swidwinska-Gajewska, A.M.; Czerczak, S. Nanosrebro-szkodliwe skutki dzialania biologicznego/nanosilver-harmful effects of biological activity. Med. Pr. 2014, 65, 831. [Google Scholar]
- Goller, C.C.; Romeo, T. Environmental influences on biofilm development. Bact. Biofilms 2008, 322, 37–66. [Google Scholar] [CrossRef]
- Mah, T.F.C.; O’Toole, G.A. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 2001, 9, 34–39. [Google Scholar] [CrossRef]
- Stewart, P.S.; Costerton, J.W. Antibiotic resistance of bacteria in biofilms. Lancet 2001, 358, 135–138. [Google Scholar] [CrossRef]
- Bakaletz, L.O. Bacterial biofilms in otitis media: Evidence and relevance. Pediatr. Infect. Dis. J. 2007, 26, S17–S19. [Google Scholar] [CrossRef] [PubMed]
- Brady, R.A.; Leid, J.G.; Calhoun, J.H.; Costerton, J.W.; Shirtliff, M.E. Osteomyelitis and the role of biofilms in chronic infection. FEMS Immunol. Med. Microbiol. 2008, 52, 13–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Høiby, N. Understanding bacterial biofilms in patients with cystic fibrosis: Current and innovative approaches to potential therapies. J. Cyst. Fibros. 2002, 1, 249–254. [Google Scholar] [CrossRef] [Green Version]
- Post, J.C.; Stoodley, P.; Hall–Stoodley, L.; Ehrlich, G.D. The role of biofilms in otolaryngologic infections. Curr. Opin. Otolaryngol. Head Neck Surg. 2004, 12, 185–190. [Google Scholar] [CrossRef]
- Ames, G.A.; Swogger, E.; Wolcott, R.; deLancey Pulcini, E.; Secor, P.; Sestrich, J.; Costerton, J.W.; Stewart, P.S. Biofilms in chronic wounds. Wound Repair Regen. 2008, 16, 37–44. [Google Scholar] [CrossRef]
- Canadian Centre for Occupational Health and Safety Act (CCOHS). What Is a LD50 and LC50? Available online: https://www.ccohs.ca/oshanswers/chemicals/ld50.html (accessed on 2 June 2021).
- Mann, R.; Holmes, A.; McNeilly, O.; Cavaliere, R.; Sotiriou, G.A.; Rice, S.A.; Gunawan, C. Evolution of biofilm-forming pathogenic bacteria in the presence of nanoparticles and antibiotic: Adaptation phenomena and cross-resistance. J. Nanobiotechnol. 2021, 19, 291. [Google Scholar] [CrossRef]
- Latimer, J.; Forbes, S.; McBain, A.J. Attenuated virulence and biofilm formation in Staphylococcus aureus following sublethal exposure to triclosan. Antimicrob Agents Chemother. 2012, 56, 3092–3100. [Google Scholar] [CrossRef] [Green Version]
- Sachivkina, N.P.; Karamyan, A.S.; Kuznetsova, O.M.; Byakhova, V.M. Development of therapeutic transdermal systems for microbial biofilm destruction. FEBS Open Bio. 2019, 9, 386. [Google Scholar]
- Kravtsov, E.G.; Anokhina, I.V.; Rybas, Y.A.; Sachivkina, N.P.; Ermolaev, A.V.; Brodskaya, S.B. Effects of female sex hormones on adhesion of candida albicans yeast-like fungi to the buccal epithelium. Bull. Exp. Biol. Med. 2014, 157, 246–248. [Google Scholar] [CrossRef] [PubMed]
- Sachivkina, N.P.; Kravtsov, E.G.; Vasilyeva, E.A.; Anokhina, I.V.; Dalin, M.V. Study of antimycotic activity of Lyticase. Bull. Exp. Biol. Med. 2009, 148, 214–216. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coded Variables | Experimental Variables | Volume Yield (%) | Mass Yield (%) | Inhibition Diameter of S. aureus (mm) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Trial | % EtOH (X1) | Ratio m/v (X2) | Temps (X3) | % EtOH (X1) | Ratio m/v (X2) | Time in Hours (X3) | Exp | Theo | Exp | Theo | Exp | Theo |
| 1 | 1 | 0 | 1 | 100 | 15/100 | 24 | 93.00 | 92.37 | 13.10 | 12.20 | 27.00 | 23.88 |
| 2 | 1 | −1 | 0 | 100 | 10/100 | 15 | 97.00 | 97.12 | 10.50 | 9.93 | 24.00 | 24.38 |
| 3 | 0 | 1 | −1 | 50 | 20/100 | 6 | 87.00 | 86.50 | 9.50 | 8.83 | 21.00 | 18.25 |
| 4 | −1 | −1 | 0 | 0 | 10/100 | 15 | 94.00 | 93.12 | 7.30 | 6.28 | 8.00 | 7.88 |
| 5 | −1 | 1 | 0 | 0 | 20/100 | 15 | 87.00 | 86.87 | 7.50 | 6.68 | 14.00 | 13.63 |
| 6 | 1 | 1 | 0 | 100 | 20/100 | 15 | 84.00 | 84.87 | 12.80 | 10.23 | 26.00 | 26.13 |
| 7 | 0 | 0 | 0 | 50 | 15/100 | 15 | 93.00 | 93.33 | 8.60 | 8.73 | 19.00 | 17.00 |
| 8 | 0 | 0 | 0 | 50 | 15/100 | 15 | 94.00 | 93.33 | 8.90 | 8.73 | 15.00 | 17.00 |
| 9 | −1 | 0 | −1 | 0 | 15/100 | 6 | 90.00 | 90.62 | 7.60 | 8.50 | 11.00 | 9.63 |
| 10 | 1 | 0 | −1 | 100 | 15/100 | 6 | 92.00 | 91.62 | 13.00 | 12.05 | 27.00 | 25.13 |
| 11 | 0 | −1 | −1 | 50 | 10/100 | 6 | 96.00 | 96.25 | 8.40 | 8.53 | 16.00 | 13.00 |
| 12 | 0 | 0 | 0 | 50 | 15/100 | 15 | 93.00 | 93.33 | 8.70 | 8.73 | 17.00 | 17.00 |
| 13 | −1 | 0 | 1 | 0 | 15/100 | 24 | 91.00 | 91.37 | 7.60 | 8.55 | 13.00 | 10.38 |
| 14 | 0 | −1 | 1 | 50 | 10/100 | 24 | 96.00 | 96.50 | 8.50 | 8.58 | 16.00 | 14.25 |
| 15 | 0 | 1 | 1 | 50 | 20/100 | 24 | 88.00 | 87.75 | 9.10 | 8.98 | 18.00 | 16.50 |
| Volume Yield (%) | Mass Yield (%) | Inhibition Diameter (mm) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Source | Sum of Squares | F-Value | p-Value | Sum of Squares | F-Value | p-Value | Sum of Squares | F-Value | p-Value |
| Model | 197.417 | 28.00 | 0.001 | 56.3427 | 36.12 | 0.001 | 467.983 | 22.13 | 0.002 |
| Linear | 174.250 | 74.15 | 0.000 | 49.2550 | 94.72 | 0.000 | 448.750 | 63.65 | 0.000 |
| X1 | 2.000 | 2.55 | 0.171 | 47.0450 | 271.41 | 0.000 | 420.500 | 178.94 | 0.000 |
| X2 | 171.125 | 218.46 | 0.000 | 2.2050 | 12.72 | 0.016 | 28.125 | 11.97 | 0.018 |
| X3 | 1.125 | 1.44 | 0.284 | 0.0050 | 0.03 | 0.872 | 0.125 | 0.05 | 0.827 |
| Square | 13.917 | 5.92 | 0.042 | 5.9202 | 11.38 | 0.011 | 11.983 | 1.70 | 0.282 |
| X1 * X1 | 7.619 | 11.20 | 0.020 | 4.6058 | 26.76 | 0.004 | 6.519 | 2.97 | 0.145 |
| X2 * X2 | 5.984 | 7.86 | 0.038 | 0.4959 | 2.31 | 0.189 | 0.791 | 0.22 | 0.658 |
| X3 * X3 | 0.314 | 0.40 | 0.554 | 0.8185 | 4.72 | 0.082 | 4.673 | 1.99 | 0.218 |
| Two-Way Interaction | 9.250 | 3.94 | 0.087 | 1.1675 | 2.25 | 0.201 | 7.250 | 1.03 | 0.455 |
| X1 * X2 | 9.000 | 11.49 | 0.019 | 1.1025 | 6.36 | 0.053 | 4.000 | 1.70 | 0.249 |
| X1 * X3 | 0.000 | 0.00 | 1.000 | 0.0025 | 0.01 | 0.909 | 1.000 | 0.43 | 0.543 |
| X2 * X3 | 0.250 | 0.32 | 0.597 | 0.0625 | 0.36 | 0.574 | 2.250 | 0.96 | 0.373 |
| Error | 3.917 | 0.8667 | 11.750 | ||||||
| Lack of Fit | 3.250 | 3.25 | 0.244 | 0.8200 | 11.71 | 0.080 | 3.750 | 0.31 | 0.820 |
| Pure Error | 0.667 | 0.0467 | 8.000 | ||||||
| Total | 201.333 | 57.2093 | 479.733 | ||||||
| R2 | R2 Adj | AAD | Bf | Af | |
|---|---|---|---|---|---|
| Volume yield | 98.05 | 94.55 | 0.008 | 1.000 | 1.008 |
| Mass yield | 98.49 | 95.76 | 0.069 | 0.970 | 1.069 |
| Inhibition diameter | 97.55 | 93.14 | 0.088 | 0.936 | 1.089 |
| % EtOH | Ratio m/v | Time (h) | D (mm) | Mass Yield (%) | Volume Yield (%) | Composite Desirability | |
|---|---|---|---|---|---|---|---|
| 100 (1) | 20/100 (1) | 6 (−1) | Solution | 28.625 | 13.595 | - | 0.96 |
| Experimental value | 31 | 14.3 | 93 |
| No. | Name | RT, min | Structure | m/z, MS Spectra | m/z, MS/MS Spectra (CE 30 eV) | % |
|---|---|---|---|---|---|---|
| 1 | Compound 1 | 15, 24 |  | 314 | 269, 253, 237, 211, 209, 192, 175, 160, 145, 143, 137, 121, 107 | 1.86 |
| 2 | Compound 2 | 17, 68 |  | 328 | 283, 269, 253, 237, 189, 174, 151, 121, 107 | 0.64 |
| 3 | Compound 3 | 19, 66 |  | 314 | 269, 253, 237, 211, 209, 192, 175, 145, 143, 137, 121, 107 | 0.69 |
| 4 | Pseudorotundine | 20, 33 |  | 356 | 192, 190, 177 | 1.68 |
| 5 | Tetrahydropalmatine (Rotundine) | 23, 54 |  | 356 | 192, 190, 177 | 3.12 |
| 6 | Compound 6 + Unknown compound | 24, 19 |  | 328 (78%), 368 (22%) | 283, 268, 252, 237, 189, 174, 145, 121, 107 + 353, 352, 338, 336, 324, 310, 307 | 1.83 |
| 7 | Jatrorrhizine | 24, 73 |  | 338 | 323, 322, 308, 294, 279 | 11.02 |
| 8 | Pseudocolumbamine | 25, 88 |  | 338 | 323, 322, 308, 294, 279, 265 | 6.33 |
| 9 | Columbamine +7,8-dihydro-8-hydroxypalmatine | 26, 29 |  | 338 (95%) | 323, 322, 308, 306, 294, 279, 277, 265 | 19.21 |
| 370 (5%) | 355, 354, 340, 326, 312, 311 | |||||
| 10 | Pseudopalmatine | 28, 19 |  | 352 | 336, 320, 308, 294, 292, 279 | 1.99 |
| 11 | Palmatine | 28, 98 |  | 352 | 336, 322, 320, 308, 294, 292, 278 | 51.63 |
| Chemical Element | Mean Fluorescence Intensity, cps/uA | Standard Deviation |
|---|---|---|
| Si | 0.0581 | 0.0024 |
| S | 0.2052 | 0.0028 |
| Cl | 2.5942 | 0.0097 |
| K | 0.6511 | 0.0010 |
| Ca | 0.6185 | 0.0253 |
| Mn | 0.1482 | 0.0033 |
| Fe | 3.4854 | 0.0127 |
| Zn | 0.1624 | 0.0014 |
| Br | 4.6529 | 0.0114 |
| Extract | AgNPs | |||||
|---|---|---|---|---|---|---|
| MIC (µg/mL) | MBC (or MFC) (µg/mL) | MBC (or MFC)/MIC | MIC (µg/mL) | MBC (or MFC) (µg/mL) | MBC (or MFC)/MIC | |
| A. xylosoxidans 4892 | 32 | 128 | 4 | 4 | 8 | 2 |
| A. baumannii 5841 | 16 | 32 | 2 | 64 | 64 | 1 |
| C. freundi 426 | 512 | >1024 | - | 64 | 128 | 2 |
| Conybacterium spp 1638 | 256 | 512 | 2 | 64 | 128 | 2 |
| E. avium 1669 | 128 | 512 | 4 | 64 | 128 | 2 |
| E. cloacae 6392 | 512 | >1024 | - | 32 | 32 | 1 |
| E. faecalis 5960 | 128 | 512 | 4 | 32 | 64 | 2 |
| E. coli M17 | 128 | 512 | 4 | 8 | 32 | 4 |
| E. coli 1449 | 256 | 512 | 2 | 64 | 128 | 2 |
| E. coli ATCC 25922 | 128 | 512 | 4 | 16 | 64 | 4 |
| K. oxytoca 3003 | >1024 | >1024 | - | 32 | 64 | 2 |
| K. pneumoniae 1449 | >1024 | >1024 | - | 64 | 128 | 2 |
| K. rizophilia 1542 | 512 | 512 | 1 | 32 | 64 | 2 |
| M. catarrhalis 4222 | 16 | 32 | 2 | 2 | 8 | 4 |
| M. morganii 1543 | 256 | 512 | 2 | 8 | 32 | 4 |
| Ps mirabilis 1543 | 256 | 512 | 2 | 16 | 16 | 1 |
| P. aeruginosa 3057 | 128 | 512 | 4 | 8 | 32 | 4 |
| S. marsescens 6441 | >1024 | >1024 | - | 64 | 128 | 2 |
| S. aureus 1449 | 256 | 512 | 2 | 64 | 128 | 2 |
| S. aureus ATCC 6538 | 64 | 512 | 8 | 32 | 64 | 2 |
| S. simulans 5882 | 512 | 512 | 1 | 64 | 64 | 1 |
| S. agalactiae 3984 | 512 | >1024 | - | 64 | 128 | 2 |
| C. albicans ATCC 10231 | 8 | 32 | 4 | 64 | 128 | 2 |
| C. albicans | 128 | 256 | 2 | 512 | 512 | 1 |
| E. coli 1449 | S. aureus 1449 | E. faecalis 5960 | ||
|---|---|---|---|---|
| Ampicillin | MIC/MBC | 1024/ND | 64/512 | 32/64 |
| FIC (AMP + O-ECB) | 0.015 (1/128) | 0.125 (1/16) | 0.125 (1/16) | |
| FIC(AMP + AgNPs) | 0.250 (1/8) | 0.125 (1/16) | 0.125 (1/16) | |
| Kanamycin | MIC/MBC | 32/128 | 8/16 | 128/128 |
| FIC (KA + O-ECB) | 0.250 (1/8) | 0.250 (1/8) | 0.031 (1/64) | |
| FIC(KA + AgNPs) | 0.125 (1/16) | 0.125 (1/16) | 0.125 (1/16) | |
| Cefazolin | MIC/MBC | 64/128 | 8/16 | 64/256 |
| FIC (CZ + O-ECB) | 0.250 (1/8) | 0.125 (1/8) | 0.031 (1/64) | |
| FIC(CZ + AgNPs) | 0.500 (1/16) | 0.500 (1/16) | 0.500(1/16) |
| LD50 | ||
|---|---|---|
| mg/mL | g/kg of Body Weight | |
| O-ECB | 40.00 | 29.75 |
| AgNPs | 0.6 | 0.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arsene, M.M.J.; Viktorovna, P.I.; Alla, M.V.; Mariya, M.A.; Sergei, G.V.; Cesar, E.; Davares, A.K.L.; Parfait, K.; Wilfrid, K.N.; Nikolay, T.S.; et al. Optimization of Ethanolic Extraction of Enantia chloranta Bark, Phytochemical Composition, Green Synthesis of Silver Nanoparticles, and Antimicrobial Activity. Fermentation 2022, 8, 530. https://doi.org/10.3390/fermentation8100530
Arsene MMJ, Viktorovna PI, Alla MV, Mariya MA, Sergei GV, Cesar E, Davares AKL, Parfait K, Wilfrid KN, Nikolay TS, et al. Optimization of Ethanolic Extraction of Enantia chloranta Bark, Phytochemical Composition, Green Synthesis of Silver Nanoparticles, and Antimicrobial Activity. Fermentation. 2022; 8(10):530. https://doi.org/10.3390/fermentation8100530
Chicago/Turabian StyleArsene, Mbarga M. J., Podoprigora I. Viktorovna, Marukhlenko V. Alla, Morozova A. Mariya, Goriainov V. Sergei, Esparza Cesar, Anyutoulou K. L. Davares, Kezimana Parfait, Kamgang N. Wilfrid, Tuturov S. Nikolay, and et al. 2022. "Optimization of Ethanolic Extraction of Enantia chloranta Bark, Phytochemical Composition, Green Synthesis of Silver Nanoparticles, and Antimicrobial Activity" Fermentation 8, no. 10: 530. https://doi.org/10.3390/fermentation8100530