

Production of Xylooligosaccharide from Cassava Pulp’s Waste by Endo-β-1,4-D-Xylanase and Characterization of Its Prebiotic Effect by Fermentation of Lactobacillus acidophilus
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Production of XOS from Cassava Pulp
2.3. Analysis of XOS by Thin Layer Chromatography (TLC) and High-Performance Liquid Chromatography (HPLC)
2.4. MRS-Modified Media (MRSm) Production
2.5. Fermentation of L. acidophilus
2.6. Calculation of Lactic Acid Bacterial Populations
2.7. SCFA and Lactic Acid Analysis
3. Results
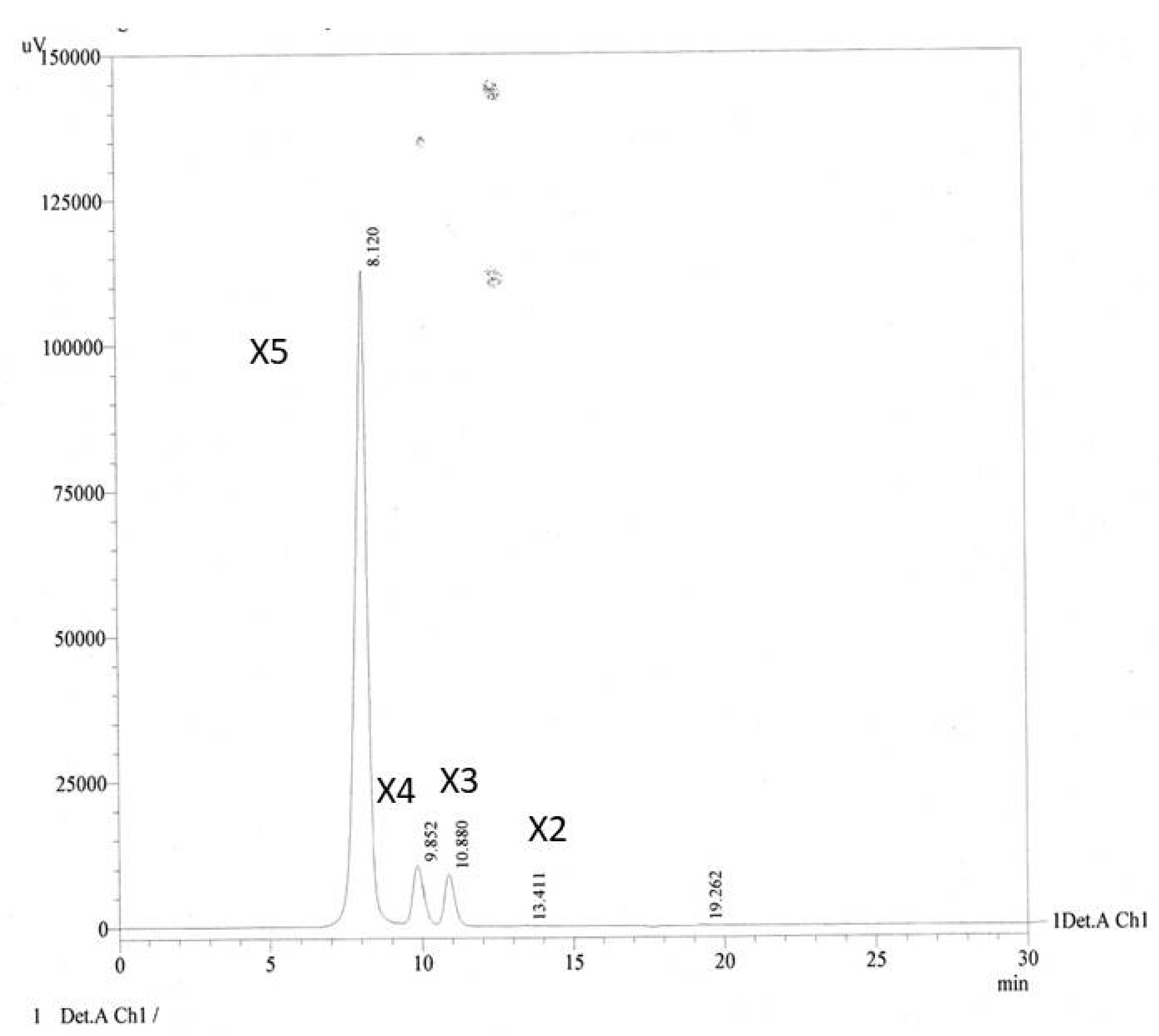
3.1. Production and Characterization of XOS from Cassava Pulp
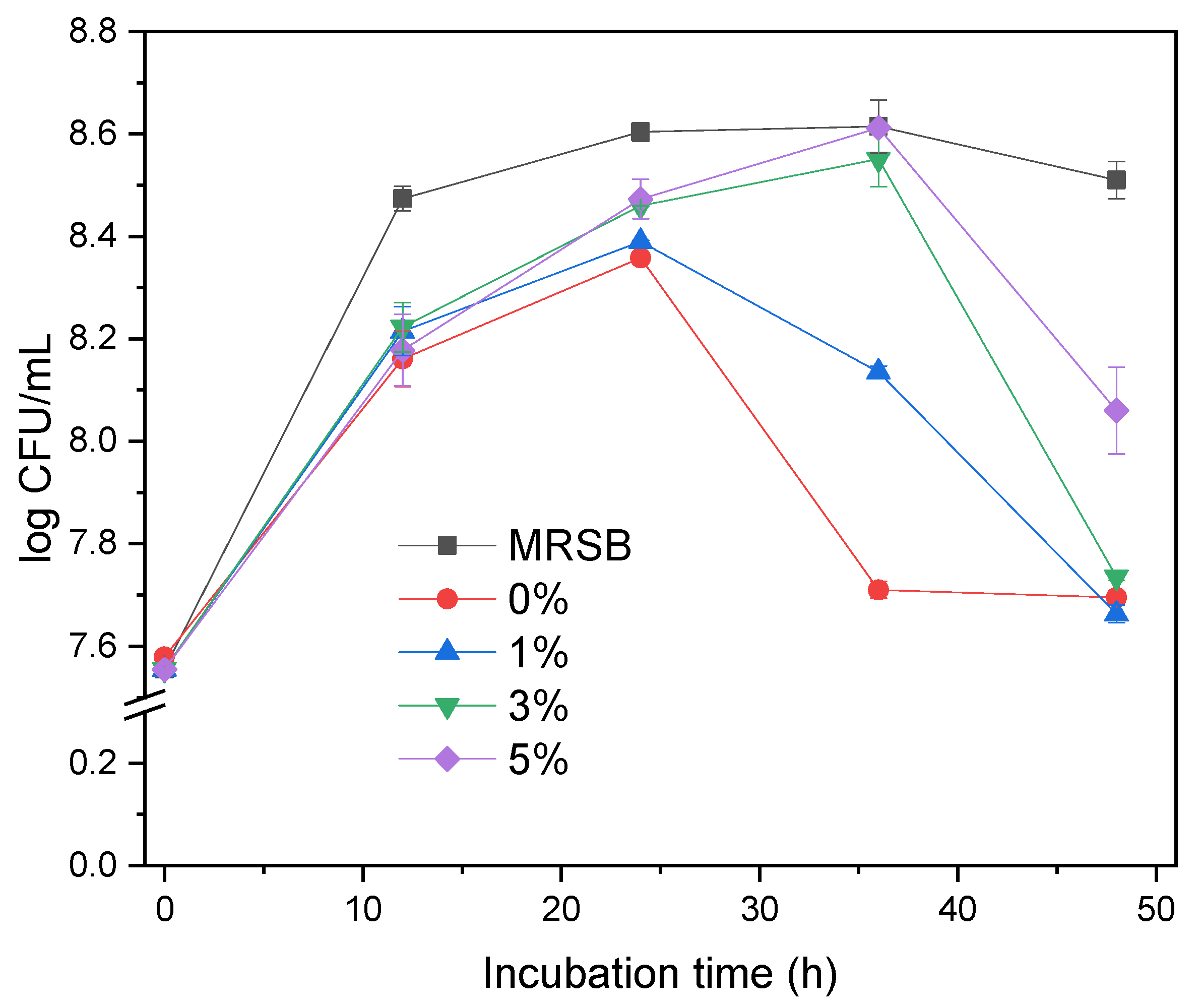
3.2. Growth of L. acidophilus
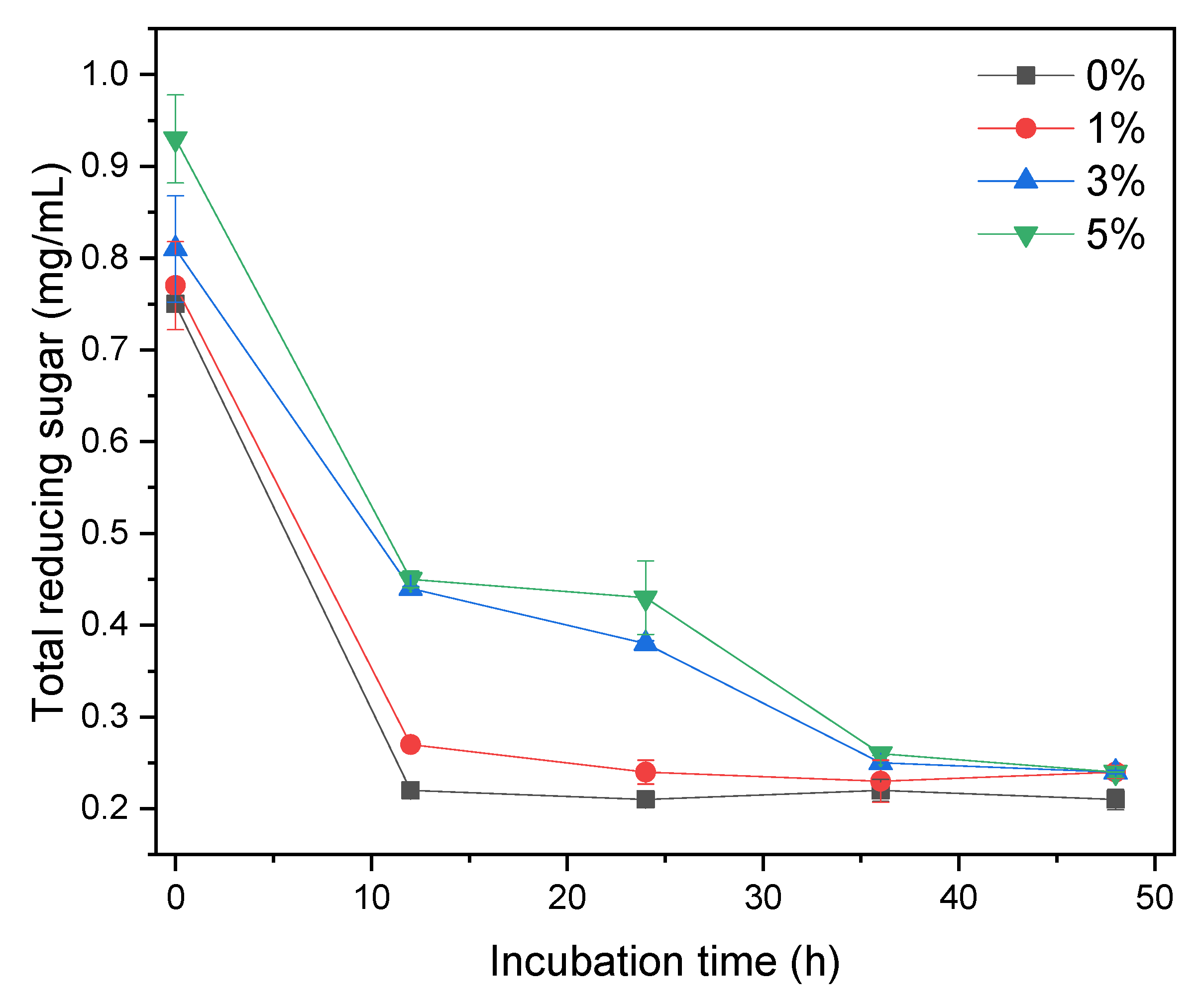
3.3. Total Reducing Sugar
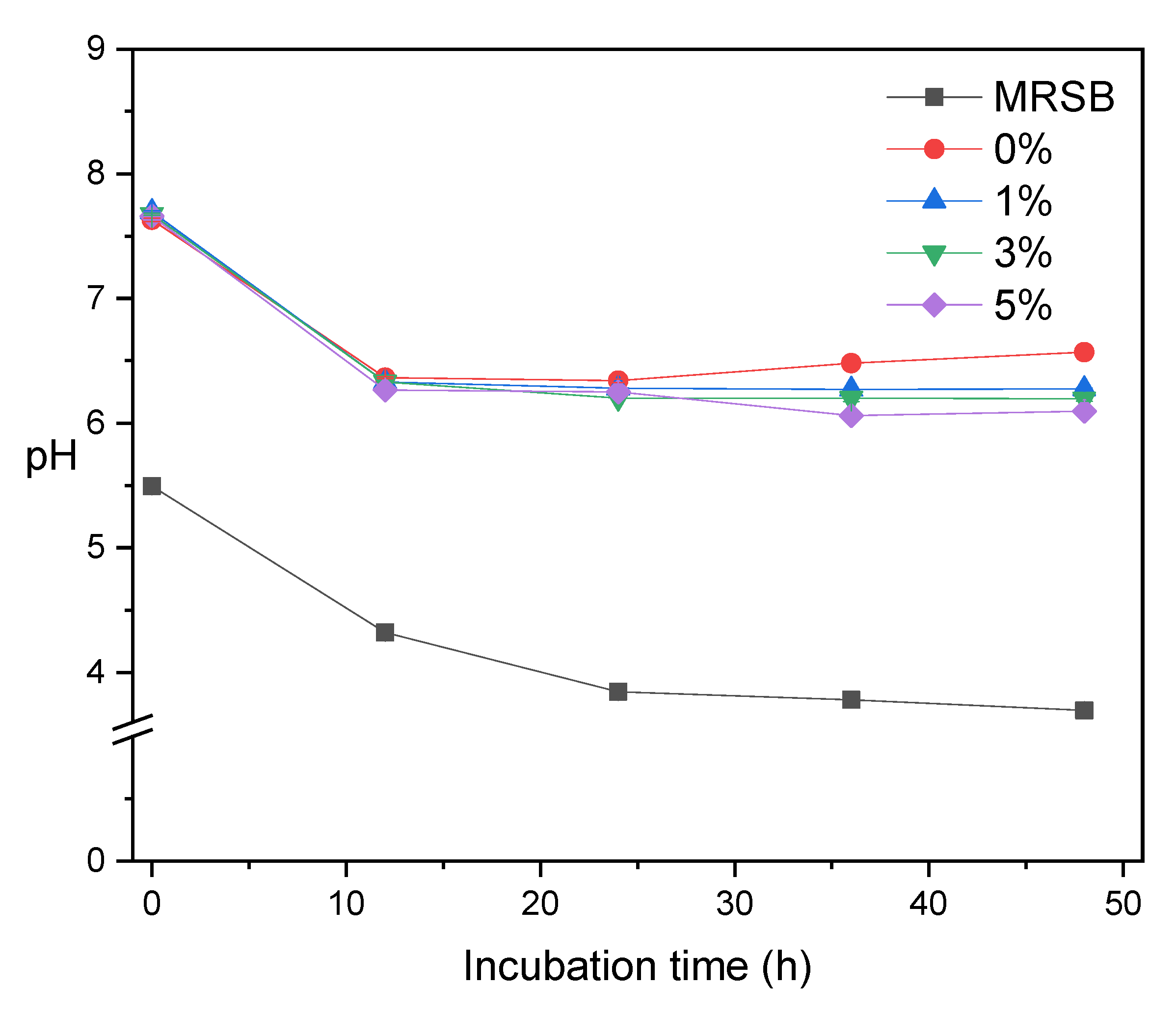
3.4. Short-Chain Fatty Acids (SCFA)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guarner, F.; Malagelada, J.-R. Gut flora in health and disease. Lancet 2003, 361, 512–519. [Google Scholar] [CrossRef]
- Fijan, S. Microorganisms with Claimed Probiotic Properties: An Overview of Recent Literature. Int. J. Environ. Res. Public Health 2014, 11, 4745–4767. [Google Scholar] [CrossRef] [PubMed]
- Kraehenbuhl, J.P.; Corbett, M. Keeping the Gut Microflora at Bay. Science 2004, 303, 1624–1625. [Google Scholar] [CrossRef]
- Guinane, C.M.; Cotter, P.D. Role of the gut microbiota in health and chronic gastrointestinal disease: Understanding a hidden metabolic organ. Ther. Adv. Gastroenterol. 2013, 6, 295–308. [Google Scholar] [CrossRef]
- Berg, R.D. The indigenous gastrointestinal microflora. Trends Microbiol. 1996, 4, 430–435. [Google Scholar] [CrossRef]
- Gibson, G.R.; Probert, H.M.; Loo JVan Rastall, R.A.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Updating the concept of prebiotics. Nutr. Res. Rev. 2004, 17, 259–275. [Google Scholar] [CrossRef]
- Gibson, G.R.; Roberfroid, M.B. Dietary Modulation of the Human Colonic Microbiota: Introducing the Concept of Prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef]
- Klessen, B.; Sykura, B.; Zunft, H.-J.; Blaut, M. Effects of inulin and lactose on fecal microflora, microbial activity, and bowel habit in elderly constipated. Am. J. Clin. Nutr. 1997, 65, 1397–1402. [Google Scholar] [CrossRef]
- Hsu, C.-K.; Liao, J.-W.; Chung, Y.-C.; Hsieh, C.-P.; Chan, Y.-C. Xylooligosaccharides and Fructooligosaccharides Affect the Intestinal Microbiota and Precancerous Colonic Lesion Development in Rats. J. Nutr. 2004, 134, 1523–1528. [Google Scholar] [CrossRef]
- Swennen, K.; Courtin, C.M.; Delcour, J.A. Non-digestible oligosaccharides with prebiotic properties. Crit. Rev. Food Sci. Nutr. 2006, 46, 459–471. [Google Scholar] [CrossRef]
- Laparra, J.M.; Sanz, Y. Interactions of gut microbiota with functional food components and nutraceuticals. Pharmacol. Res. 2010, 61, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R. Dietary Modulation of the Human Gut Microflora Using the Prebiotics Oligofructose and Inulin. J. Nutr. 1999, 129, 1438S–1441S. [Google Scholar] [CrossRef] [PubMed]
- Mai, V.; Morris, J.G. Colonic Bacterial Flora: Changing Understandings in the Molecular Age. J. Nutr. 2004, 134, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Ratnadewi, A.A.I.; Masruroh, H.; Suwardiyanto; Santoso, A.B. Application of coffee peel waste as raw material for Xylooligosaccharide Production. Coffe Sci. 2019, 14, 446–454. [Google Scholar] [CrossRef]
- Ratnadewi, A.A.I.; Santoso, A.B.; Sulistyaningsih, E.; Handayani, W. Application of Cassava Peel and Waste as Raw Materials for Xylooligosaccharide Production Using Endoxylanase from Bacillus subtilis of Soil Termite Abdomen. Procedia Chem. 2016, 18, 31–38. [Google Scholar] [CrossRef]
- Hafidah, A.H.; Sulistyaningsih, E.; Handayani, W.; Ratnadewi, A.A.I. Prebiotic Potential of Xylooligosaccharides Derived from Cassava Dregs in Balb/c Mice Colon. Pertanika J. Trop. Agric. Sci. 2018, 41, 1021–1031. [Google Scholar]
- Li, Z.; Summanen, P.H.; Komoriya, T.; Finegold, S.M. In Vitro study of the prebiotic xylooligosaccharide (XOS) on the growth of Bifidobacterium spp. and Lactobacillus spp. Int. J. Food Sci. Nutr. 2015, 66, 919–922. [Google Scholar] [CrossRef]
- Ratnadewi, A.A.I.; Zain, M.H.A.; Kusuma, A.A.N.N.; Handayani, W.; Nugraha, A.S.; Siswoyo, T.A. Lactobacillus casei fermentation towards xylooligosaccharide (XOS) obtained from coffee peel enzymatic hydrolysate. Biocatal. Agric. Biotechnol. 2020, 23, 101446. [Google Scholar] [CrossRef]
- Lasrado, L.D.; Gudipati, M. Antioxidant property of synbiotic combination of Lactobacillus sp. and wheat bran xylo-oligosaccharides. J. Food Sci. Technol. 2014, 52, 4551–4557. [Google Scholar] [CrossRef]
- Buruiană, C.T.; Vizireanu, C. Prebiotic xylooligosaccharides from lignocellulosic materials: Production, purification and applications-An overview. Ann. Univ. Dunarea De Jos Galati 2014, 38, 18–31. [Google Scholar]
- Fiorini, D.; Boarelli, M.C.; Gabbianelli, R.; Ballini, R.; Pacetti, D. A quantitative headspace–solid-phase microextraction–gas chromatography–flame ionization detector method to analyze short chain free fatty acids in rat feces. Anal. Biochem. 2016, 508, 12–14. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.M.; Barnes, M.B.; Gras, S.L.; McSweeney, C.; Lockett, T.; Augustin, M.A.; Gooley, P.R. Esterification of high amylose starch with short chain fatty acids modulates degradation by Bifidobacterium spp. J. Funct. Foods 2014, 6, 137–146. [Google Scholar] [CrossRef]
- Moura, P.; Barata, R.; Carvalheiro, F.; Gírio, F.; Loureiro-Dias, M.C.; Esteves, M.P. In Vitro fermentation of xylo-oligosaccharides from corn cobs autohydrolysis by Bifidobacterium and Lactobacillus strains. LWT Food Sci. Technol. 2007, 40, 963–972. [Google Scholar] [CrossRef]
- Samanta, A.K.; Jayapal, N.; Jayaram, C.; Roy, S.; Kolte, A.P.; Senani, S.; Sridhar, M. Xylooligosaccharides as prebiotics from agricultural by-products: Production and applications. Bioact. Carbohydr. Diet. Fibre 2015, 5, 62–71. [Google Scholar] [CrossRef]
- Aachary, A.A.; Prapulla, S.G. Xylooligosaccharides (XOS) as an Emerging Prebiotic: Microbial Synthesis, Utilization, Structural Characterization, Bioactive Properties, and Applications. Compr. Rev. Food Sci. Food Saf. 2011, 10, 2–16. [Google Scholar] [CrossRef]
- Sims, I.M.; Ryan, J.L.J.; Kim, S.H. In Vitro fermentation of prebiotic oligosaccharides by Bifidobacterium lactis HN019 and Lactobacillus spp. Anaerobe 2014, 25, 11–17. [Google Scholar] [CrossRef]
- Wang, J.; Sun, B.; Cao, Y.; Wang, C. In Vitro fermentation of xylooligosaccharides from wheat bran insoluble dietary fiber by Bifidobacteria. Carbohydr. Polym. 2010, 82, 419–423. [Google Scholar] [CrossRef]
- Rycroft, C.E.; Jones, M.R.; Gibson, G.R.; Rastall, R.A. A comparative in vitro evaluation of the fermentation properties of prebiotic oligosaccharides. J. Appl. Microbiol. 2001, 91, 878–887. [Google Scholar] [CrossRef]
- Ooi, L.G.; Liong, M.T. Cholesterol-lowering effects of probiotics and prebiotics: A review of in Vivo and in Vitro Findings. Int. J. Mol. Sci. 2010, 11, 2499–2522. [Google Scholar] [CrossRef]
- Pertami, S.D.; Pancasiyanuar, M.; Irasari, S.A.; Rahardjo, M.B.; Wasilah, W. Lactobacillus acidophilus Probiotic Inhibits the Growth of Candida albicans. J. Dent. Indones. 2013, 20, 64–67. [Google Scholar] [CrossRef]
- Delzenne, N.M.; Kok, N. Effects of fructans-type prebiotics on lipid metabolism. Am. J. Clin. Nutr. 2001, 73, 456–458. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Soccol, C.R.; Nigam, P.; Soccol, V.T.; Vandenberghe, L.P.S.; Mohan, R. Biotechnological potential of agro-industrial residues. II: Cassava bagasse. Bioresour. Technol. 2000, 74, 81–87. [Google Scholar] [CrossRef]
- Amenaghawon, N.A.; Ogbeide, S.E.; Okieimen, C.O. Application of statistical experimental design for the optimisation of dilute sulphuric acid hydrolysis of Cassava Bagasse. Acta Polytech. Hung. 2014, 11, 239–250. [Google Scholar]
- Safitri, E.; Hanifah; Previta; Sudarko; Puspaningsih, N.N.T.; Ratnadewi, A.A.I. Cloning, purification, and characterization of recombinant endo- β-1,4-D-xylanase of Bacillus sp. From soil termite abdomen. Biocatal. Agric. Biotechnol. 2020, 31, 101877. [Google Scholar] [CrossRef]
- Carvalho, A.F.A.; de Oliva Neto, P.; da Silva, D.F.; Pastore, G.M. Xylo-oligosaccharides from lignocellulosic materials: Chemical structure, health benefits and production by chemical and enzymatic hydrolysis. Food Res. Int. 2013, 51, 75–85. [Google Scholar] [CrossRef]
- McLaughlin, H.P.; Motherway, M.O.C.; Lakshminarayanan, B.; Stanton, C.; Paul Ross, R.; Brulc, J.; Menon, R.; O’Toole, P.W.; van Sinderen, D. Carbohydrate catabolic diversity of bifidobacteria and lactobacilli of human origin. Int. J. Food Microbiol. 2015, 203, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Zhou, H.; Wang, Z. Effects of xylooligosaccharide on growth performance, activities of digestive enzymes, and intestinal microflora of juvenile Pelodiscus sinensis. Front. Agric. China 2011, 5, 612–617. [Google Scholar] [CrossRef]
- Kedia, G.; Vázquez, J.A.; Pandiella, S.S. Enzymatic digestion and in vitro fermentation of oat fractions by human lactobacillus strains. Enzyme Microb. Technol. 2008, 43, 355–361. [Google Scholar] [CrossRef]
- Reddy, S.S.; Krishnan, C. Production of high-pure xylooligosaccharides from sugarcane bagasse using crude β-xylosidase-free xylanase of Bacillus subtilis KCX006 and their bifidogenic function. LWT Food Sci. Technol. 2016, 65, 237–245. [Google Scholar] [CrossRef]
- Chapla, D.; Pandit, P.; Shah, A. Production of xylooligosaccharides from corncob xylan by fungal xylanase and their utilization by probiotics. Bioresour. Technol. 2012, 115, 215–221. [Google Scholar] [CrossRef]
- Madhukumar, M.S.; Muralikrishna, G. Fermentation of xylo-oligosaccharides obtained from wheat bran and Bengal gram husk by lactic acid bacteria and bifidobacteria. J. Food Sci. Technol. 2012, 49, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Hinnebusch, B.F.; Meng, S.; Wu, J.T.; Archer, S.Y.; Hodin, R.A. The Effects of Short-Chain Fatty Acids on Human Colon Cancer Cell Phenotype Are Associated with Histone Hyperacetylation. J. Nutr. 2002, 132, 1012–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Types of Short Chain Fatty Acids | Content (mM) | |
|---|---|---|
| 0% | 5% | |
| Acetic acid | 43.1 | 14.42 |
| Propionic acid | 0.63 | 0.25 |
| Isobutyric acid | 0.22 | 0.13 |
| n-butyric acid | 0.18 | 0.41 |
| Isovaleric acid | 0.17 | 0.25 |
| n-valeric acid | 0.03 | 0.02 |
| Lactic acid | 25.08 | 57.42 |
| Total | 69.41 | 72.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ratnadewi, A.A.I.; Rahma, M.T.; Nurhayati, N.; Santoso, A.B.; Senjarini, K.; Labes, A.; Reza, M. Production of Xylooligosaccharide from Cassava Pulp’s Waste by Endo-β-1,4-D-Xylanase and Characterization of Its Prebiotic Effect by Fermentation of Lactobacillus acidophilus. Fermentation 2022, 8, 488. https://doi.org/10.3390/fermentation8100488
Ratnadewi AAI, Rahma MT, Nurhayati N, Santoso AB, Senjarini K, Labes A, Reza M. Production of Xylooligosaccharide from Cassava Pulp’s Waste by Endo-β-1,4-D-Xylanase and Characterization of Its Prebiotic Effect by Fermentation of Lactobacillus acidophilus. Fermentation. 2022; 8(10):488. https://doi.org/10.3390/fermentation8100488
Chicago/Turabian StyleRatnadewi, Anak Agung Istri, Marena Thalita Rahma, Nurhayati Nurhayati, Agung Budi Santoso, Kartika Senjarini, Antje Labes, and Muhammad Reza. 2022. "Production of Xylooligosaccharide from Cassava Pulp’s Waste by Endo-β-1,4-D-Xylanase and Characterization of Its Prebiotic Effect by Fermentation of Lactobacillus acidophilus" Fermentation 8, no. 10: 488. https://doi.org/10.3390/fermentation8100488