
Palm Oil Mill Effluent for Lipid Production by the Diatom Thalassiosira pseudonana

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microscopic Characterization
2.2. Maintenance and Inoculum Preparation
2.3. Characterization of Palm Oil Mill Effluent
2.4. Culture Conditions
2.5. Growth Measurements
2.6. Lipid Extraction and Transesterification
2.7. Lipid Composition Analysis
3. Results and Discussion
3.1. Diatom Identification
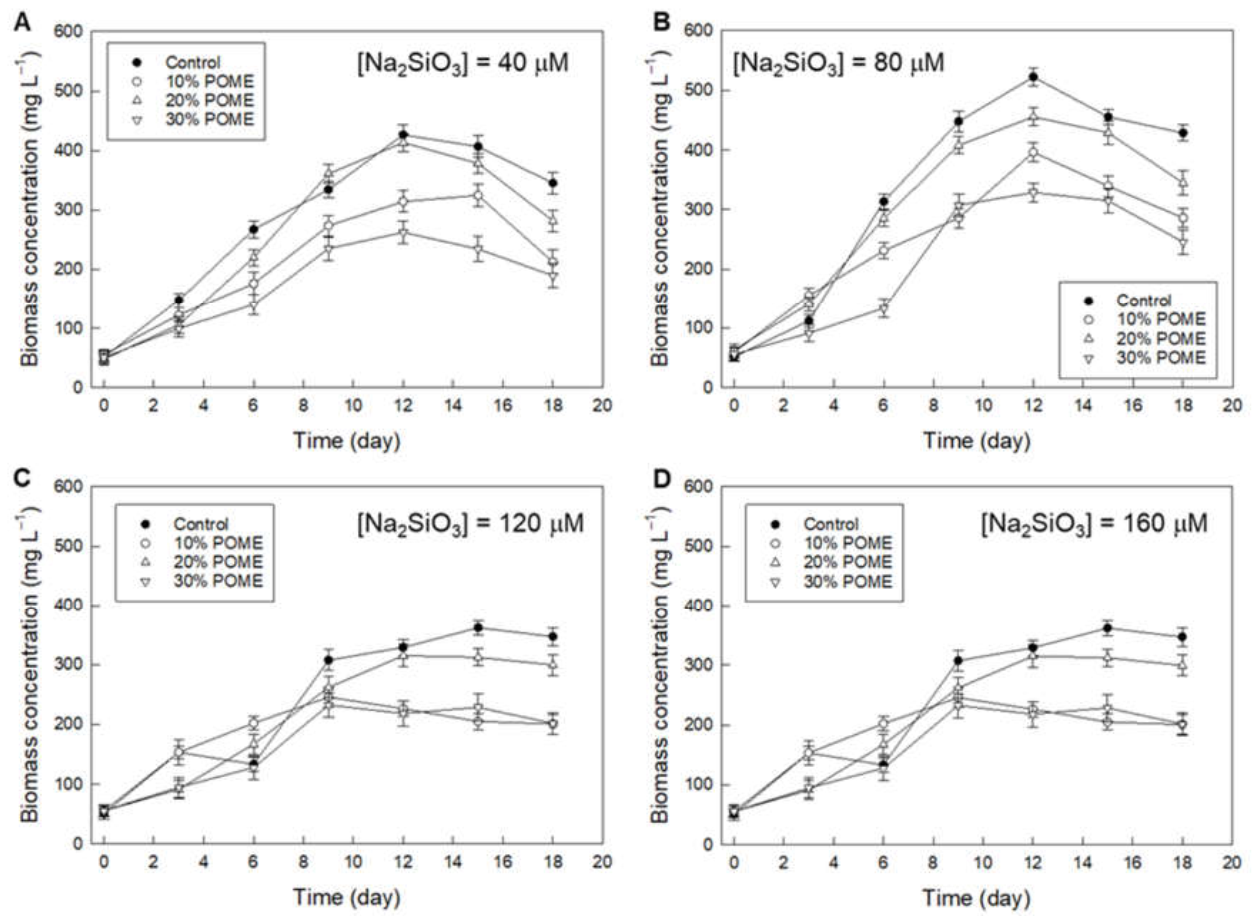
3.2. Effect of POME and Silicate Concentration on Biomass Production
3.3. Effect of Light–Dark Cycle on Biomass Production and Lipid Content
3.4. Biomass and Lipid Production under Optimal Conditions
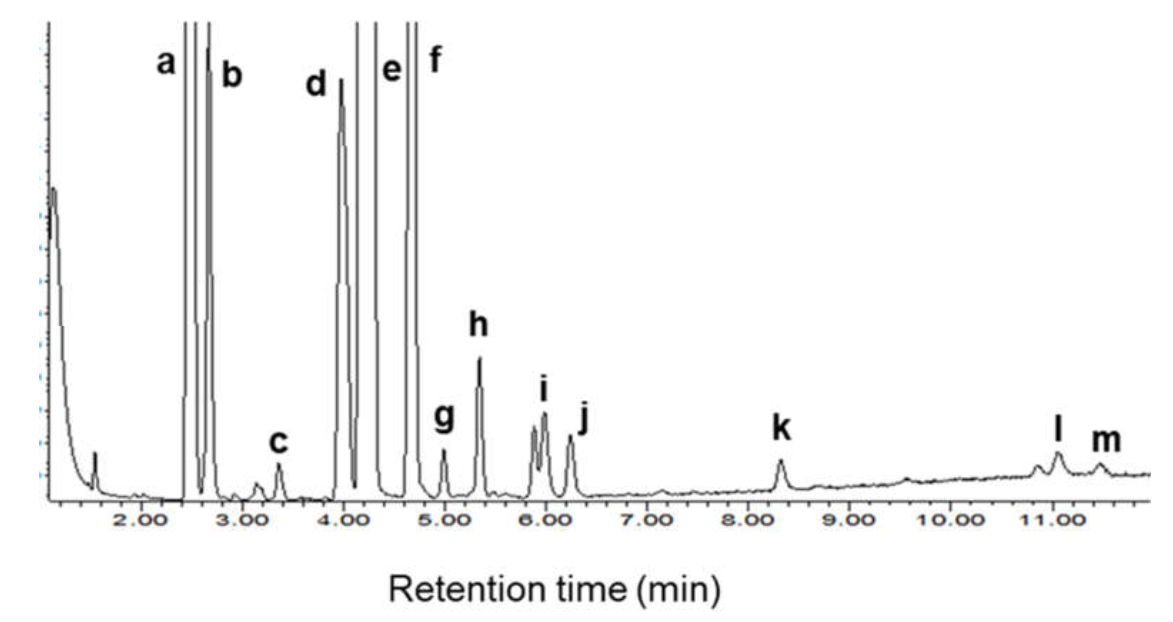
3.5. Fatty Acid Methyl Esters in Biodiesel
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kamyab, H.; Chelliapan, S.; Din, M.F.M.; Rezania, S.; Khademi, T.; Kumar, A. Palm Oil Mill Effluent as an Environmental Pollutant; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Poh, P.E.; Yong, W.-J.; Chong, M.F. Palm oil mill effluent (POME) characteristic in high crop season and the applicability of high-rate anaerobic bioreactors for the treatment of POME. Ind. Eng. Chem. Res. 2010, 49, 11732–11740. [Google Scholar] [CrossRef]
- Bux, F.; Chisti, Y. (Eds.) Algae Biotechnology: Products and Processes; Springer: New York, NY, USA, 2016. [Google Scholar] [CrossRef]
- Chisti, Y. Society and microalgae: Understanding the past and present. In Microalgae in Health and Disease Prevention; Levine, I.A., Fleurence, J., Eds.; Academic Press: London, UK, 2018; pp. 11–21. [Google Scholar] [CrossRef]
- Chisti, Y. Microalgae biotechnology: A brief introduction. In Handbook of Microalgae-Based Processes and Products: Fundamentals and Advances in Energy, Food, Feed, Fertilizer, and Bioactive Compounds; Jacob-Lopes, E., Maroneze, M.M., Queiroz, M.I., Zepka, L.Q., Eds.; Academic Press: London, UK, 2020; pp. 3–23. [Google Scholar] [CrossRef]
- Pandey, A.; Lee, D.J.; Chang, J.S.; Chisti, Y.; Soccol, C.R. (Eds.) Biomass, Biofuels, Biochemicals: Biofuels from Algae; Elsevier: New York, NY, USA, 2018. [Google Scholar] [CrossRef]
- Wang, J.K.; Seibert, M. Prospects for commercial production of diatoms. Biotechnol. Biofuels 2017, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Lim, C.I.; Biswas, W. Sustainability assessment for crude palm oil production in Malaysia using the palm oil sustainability assessment framework. Sust. Dev. 2019, 27, 253–269. [Google Scholar] [CrossRef]
- Palanisamy, K.M.; Paramasivam, P.; Jayakumar, S.; Maniam, G.P.; Rahim, M.H.A.; Govindan, N. Economical cultivation system of microalgae Spirulina platensis for lipid production. IOP Conf. Ser. Earth Environ. Sci. 2021, 641, 012022. [Google Scholar] [CrossRef]
- Palanisamy, K.M.; Paramasivam, P.; Maniam, G.P.; Rahim, M.H.A.; Govindan, N.; Chisti, Y. Production of lipids by Chaetoceros affinis in media based on palm oil mill effluent. J. Biotechnol. 2021, 327, 86–96. [Google Scholar] [CrossRef]
- Govindan, N.; Maniam, G.P.; Yusoff, M.M.; Rahim, M.H.A.; Chatsungnoen, T.; Ramaraj, R.; Chisti, Y. Statistical optimization of lipid production by the diatom Gyrosigma sp. grown in industrial wastewater. J. Appl. Phycol. 2020, 32, 375–387. [Google Scholar] [CrossRef]
- Govindan, N.; Maniam, G.P.; Sulaiman, A.Z.; Ajit, A.; Chatsungnoen, T.; Chisti, Y. Production of renewable lipids by the diatom Amphora copulata. Fermentation 2021, 7, 37. [Google Scholar] [CrossRef]
- Jayakumar, S.; Bhuyar, P.; Pugazhendhi, A.; Rahim, M.H.A.; Maniam, G.P.; Govindan, N. Effects of light intensity and nutrients on the lipid content of marine microalga (diatom) Amphiprora sp. for promising biodiesel production. Sci. Total Environ. 2021, 768, 145471. [Google Scholar] [CrossRef]
- Jasni, J.; Arisht, S.N.; Yasin, N.H.M.; Abdul, P.M.; Lin, S.-K.; Liu, C.-M.; Wu, S.-Y.; Jahim, J.M.; Takriff, M.S. Comparative toxicity effect of organic and inorganic substances in palm oil mill effluent (POME) using native microalgae species. J. Water Process Eng. 2020, 34, 101165. [Google Scholar] [CrossRef]
- Kamyab, H.; Din, M.F.M.; Keyvanfar, A.; Majid, M.Z.A.; Talaiekhozani, A.; Shafaghat, A.; Lee, C.T.; Shiun, L.J.; Ismail, H.H. Efficiency of microalgae Chlamydomonas on the removal of pollutants from palm oil mill effluent (POME). Energy Procedia 2015, 75, 2400–2408. [Google Scholar] [CrossRef] [Green Version]
- Nur, M.M.A.; Setyoningrum, T.M.; Budiaman, G.S. Potency of Botryococcus braunii cultivated on palm oil mill effluent wastewater as a source of biofuel. Environ. Eng. Res. 2017, 22, 417–425. [Google Scholar] [CrossRef] [Green Version]
- d’Ippolito, G.; Sardo, A.; Paris, D.; Vella, F.M.; Adelfi, M.G.; Botte, P.; Gallo, C.; Fontana, A. Potential of lipid metabolism in marine diatoms for biofuel production. Biotechnol. Biofuels 2015, 8, 28. [Google Scholar] [CrossRef] [Green Version]
- Savio, S.; Farrotti, S.; Paris, D.; Arnaìz, E.; Díaz, I.; Bolado, S.; Muñoz, R.; Rodolfo, C.; Congestri, R. Value-added co-products from biomass of the diatoms Staurosirella pinnata and Phaeodactylum tricornutum. Algal Res. 2020, 47, 101830. [Google Scholar] [CrossRef]
- Maeda, Y.; Nojima, D.; Yoshino, T.; Tanaka, T. Structure and properties of oil bodies in diatoms. Philos. Trans. R. Soc. B 2017, 372, 20160408. [Google Scholar] [CrossRef]
- Paasche, E. Silicon and the ecology of marine plankton diatoms. I. Thalassiosira pseudonana (Cyclotella nana) grown in a chemostat with silicate as limiting nutrient. Mar. Biol. 1973, 19, 117–126. [Google Scholar] [CrossRef]
- Armbrust, E.V.; Berges, J.A.; Bowler, C.; Green, B.R.; Martinez, D.; Putnam, N.H.; Zhou, S.; Allen, A.E.; Apt, K.E.; Bechner, M.; et al. The genome of the diatom Thalassiosira pseudonana: Ecology, evolution, and metabolism. Science 2004, 306, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, A.; Tiwari, A.; Srivastava, S. A genome-scale metabolic model of Thalassiosira pseudonana CCMP 1335 for a systems-level understanding of its metabolism and biotechnological potential. Microorganisms 2020, 8, 1396. [Google Scholar] [CrossRef]
- Hildebrand, M. Diatoms, biomineralization processes, and genomics. Chem. Rev. 2008, 108, 4855–4874. [Google Scholar] [CrossRef]
- Martin-Jézéquel, V.; Hildebrand, M.; Brzezinski, M.A. Silicon metabolism in diatoms: Implications for growth. J. Phycol. 2000, 36, 821–840. [Google Scholar] [CrossRef]
- Villanova, V.; Spetea, C. Mixotrophy in diatoms: Molecular mechanism and industrial potential. Physiol. Plant. 2021, 173, 603–611. [Google Scholar] [CrossRef]
- Baldisserotto, C.; Sabia, A.; Guerrini, A.; Demaria, S.; Maglie, M.; Ferroni, L.; Pancaldi, S. Mixotrophic cultivation of Thalassiosira pseudonana with pure and crude glycerol: Impact on lipid profile. Algal Res. 2021, 54, 102194. [Google Scholar] [CrossRef]
- Menegol, T.; Romero-Villegas, G.I.; López-Rodríguez, M.; Navarro-López, E.; López-Rosales, L.; Chisti, Y.; Molina-Grima, E. Mixotrophic production of polyunsaturated fatty acids and carotenoids by the microalga Nannochloropsis gaditana. J. Appl. Phycol. 2019, 31, 2823–2832. [Google Scholar] [CrossRef]
- Ashworth, J.; Coesel, S.; Lee, A.; Armbrust, E.V.; Orellana, M.V.; Baliga, N.S. Genome-wide diel growth state transitions in the diatom Thalassiosira pseudonana. Proc. Natl. Acad. Sci. USA 2013, 110, 7518–7523. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, R.L.; Collier, A.; Meeter, D.A. Growth response of Thalassiosira pseudonana Hasle and Heimdal clone 3H to illumination, temperature and nitrogen source. Chesapeake Sci. 1976, 17, 148–158. [Google Scholar] [CrossRef]
- León-Saiki, G.M.; Remmers, I.M.; Martens, D.E.; Lamers, P.P.; Wijffels, R.H.; van der Veen, D. The role of starch as transient energy buffer in synchronized microalgal growth in Acutodesmus obliquus. Algal Res. 2017, 25, 160–167. [Google Scholar] [CrossRef]
- Dong, H.-P.; Dong, Y.-L.; Cui, L.; Balamurugan, S.; Gao, J.; Lu, S.-H.; Jiang, T. High light stress triggers distinct proteomic responses in the marine diatom Thalassiosira pseudonana. BMC Genom. 2016, 17, 994. [Google Scholar] [CrossRef] [Green Version]
- Vásquez-Suárez, A.; Guevara, M.; González, M.; Cortez, R.; Arredondo-Vega, B. Crecimiento y composición bioquímica de Thalassiosira pseudonana (Thalassiosirales: Thalassiosiraceae) bajo cultivo semi-continuo en diferentes medios y niveles de irradiancies. (Growth and biochemical composition of Thalassiosira pseudonana (Thalassiosirales: Thalassiosiraceae) cultivated in semicontinuous system at different culture media and irradiances). Rev. Biol. Trop. 2013, 61, 1003–1013. [Google Scholar]
- Tonon, T.; Harvey, D.; Larson, T.R.; Graham, I.A. Long chain polyunsaturated fatty acid production and partitioning to triacylglycerols in four microalgae. Phytochemistry 2002, 61, 15–24. [Google Scholar] [CrossRef]
- Zhukova, N.V. Changes in the lipid composition of Thalassiosira pseudonana during its life cycle. Russ. J. Plant Physiol. 2004, 51, 702–707. [Google Scholar] [CrossRef]
- Alverson, A.J.; Beszteri, B.; Julius, M.L.; Theriot, E.C. The model marine diatom Thalassiosira pseudonana likely descended from a freshwater ancestor in the genus Cyclotella. BMC Evol. Biol. 2011, 11, 125. [Google Scholar] [CrossRef] [Green Version]
- Aké-Castillo, J.A.; Hernández-Becerril, D.U.; Meave del Castillo, M.E. Species of the genus Thalassiosira (Bacillariophyceae) from the Gulf of Tehuantepec, Mexico. Bot. Mar. 1999, 42, 487–503. [Google Scholar] [CrossRef]
- Gastineau, R.; Hamedi, C.; Hamed, M.B.B.; Abi-Ayad, S.-M.-A.; Bąk, M.; Lemieux, C.; Turmel, M.; Dobosz, S.; Wróbel, R.J.; Kierzek, A.; et al. Morphological and molecular identification reveals that waters from an isolated oasis in Tamanrasset (extreme South of Algerian Sahara) are colonized by opportunistic and pollution-tolerant diatom species. Ecol. Indic. 2021, 121, 107104. [Google Scholar] [CrossRef]
- Kipp, R.M.; Mccarthy, M.; Fusaro, A. Thalassiosira pseudonana (Hustedt) Hasle and Heimdal, (1957) 1970: U.S Geological Survey, Nonindigenous Aquatic Species Database, Gainesville, FL, and NOAA Great Lakes Aquatic Nonindigenous Species Information System, Ann Arbor, MI. Revision Date: 9/12/2019. Available online: https://nas.er.usgs.gov/queries/GreatLakes/FactSheet.aspx?SpeciesID=1692 (accessed on 2 November 2021).
- Kiss, K.T. Occurrence of Thalassiosira pseudonana new record Bacillariophyceae in some rivers of Hungary. Acta Bot. Hung. 1984, 30, 277–288. [Google Scholar]
- Lowe, R.L.; Busch, D.E. Morphological observations on two species of the diatom genus Thalassiosira from fresh-water habitats in Ohio. Trans. Am. Microsc. Soc. 1975, 94, 118–123. [Google Scholar] [CrossRef]
- Muylaert, K.; Sabbe, K. The diatom genus Thalassiosira (Bacillariophyta) in the estuaries of the Schelde (Belgium/The Netherlands) and the Elbe (Germany). Bot. Mar. 1996, 39, 103–115. [Google Scholar] [CrossRef]
- Gleich, S.J.; Plough, L.V.; Glibert, P.M. Photosynthetic efficiency and nutrient physiology of the diatom Thalassiosira pseudonana at three growth temperatures. Mar. Biol. 2020, 167, 124. [Google Scholar] [CrossRef]
- Sabia, A.; Clavero, E.; Pancaldi, S.; Rovira, J.S. Effect of different CO2 concentrations on biomass, pigment content, and lipid production of the marine diatom Thalassiosira pseudonana. Appl. Microbiol. Biotechnol. 2018, 102, 1945–1954. [Google Scholar] [CrossRef]
- Hewes, C.D. Timing is everything: Optimizing crop yield for Thalassiosira pseudonana (Bacillariophyceae) with semi-continuous culture. J. Appl. Phycol. 2016, 28, 3213–3223. [Google Scholar] [CrossRef]
- Price, N.M.; Thompson, P.A.; Harrison, P.J. Selenium: An essential element for growth of the coastal marine diatom Thalassiosira pseudonana (Bacillariophyceae). J. Phycol. 1987, 23, 1–9. [Google Scholar] [CrossRef]
- Doucette, G.J.; Price, N.M.; Harrison, P.J. Effects of selenium deficiency on the morphology and ultrastructure of the coastal marine diatom Thalassiosira pseudonana (Bacillariophyceae). J. Phycol. 1987, 23, 9–17. [Google Scholar] [CrossRef]
- Cong, N.V.; Vien, D.T.H.; Hong, D.D. Fatty acid profile and nutrition values of the microalga (Thalassiosira pseudonana) used in white shrimp culture. Vietnam J. Sci. Technol. 2018, 56, 138–145. [Google Scholar]
- Mai, T.D.; Lee-Chang, K.J.; Jameson, I.D.; Hoang, T.; Cai, N.B.A.; Pham, H.Q. Fatty acid profiles of selected microalgae used as live feeds for shrimp postlarvae in Vietnam. Aquacult. J. 2021, 1, 26–38. [Google Scholar] [CrossRef]
- Volkman, J.K.; Jeffrey, S.W.; Nichols, P.D.; Rogers, G.I.; Garlan, C.D. Fatty acid and lipid composition of 10 species of microalgae used in mariculture. J. Exp. Mar. Biol. Ecol. 1989, 128, 219–240. [Google Scholar] [CrossRef]
- Blanco, S. Diatom taxonomy and identification keys. In Modern Trends in Diatom Identification; Cristobal, G., Blanco, S., Bueno, G., Eds.; Springer: Cham, Switzerland, 2020; pp. 25–38. [Google Scholar] [CrossRef]
- Hustedt, F. Die Süsswasser-Flora Mitteleuropas: Heft 10: Bacillariophyta (Diatomeae), 2nd ed.; Verlag Von Gustav Fischer: Jena, Germany, 1930; p. 466. [Google Scholar]
- Hendey, N.I. An Introductory Account of the Smaller Algae of British Coastal Waters, Part V, Bacillariophyceae (Diatoms); HMSO: London, UK, 1964. [Google Scholar]
- Wehr, J.D.; Sheath, R.G. Freshwater Algae of North America: Ecology and Classification; Academic Press: London, UK, 2003. [Google Scholar] [CrossRef]
- Taylor, J.C.; Harding, W.R.; Archibald, C.G.M. A Methods Manual for the Collection, Preparation and Analysis of Diatom Samples; WRC Report No TT 281/07; Water Research Commission: Pretoria, South Africa, 2007. [Google Scholar]
- Hasle, G.R. Examination of diatom type material: Nitzschia delicatissima Cleve, Thalassiosira minuscula Krasske, and Cyclotella nana Hustedt. Br. Phycol. J. 1976, 11, 101–110. [Google Scholar] [CrossRef]
- Rea, I.; Terracciano, M.; Chandrasekaran, S.; Voelcker, N.H.; Dardano, P.; Martucci, N.M.; De Stefano, L. Bioengineered silicon diatoms: Adding photonic features to a nanostructured semiconductive material for biomolecular sensing. Nanoscale Res. Lett. 2016, 11, 405. [Google Scholar] [CrossRef] [Green Version]
- Guillard, R.R.L.; Lorenzen, C.J. Yellow-green algae with chlorophyllide C12. J. Phycol. 1972, 8, 10–14. [Google Scholar] [CrossRef]
- American Public Health Association. Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 1999. [Google Scholar]
- Neoh, C.H.; Lam, C.Y.; Ghani, S.M.; Ware, I.; Sarip, S.H.M.; Ibrahim, Z. Bioremediation of high-strength agricultural wastewater using Ochrobactrum sp. strain SZ1. 3 Biotech 2016, 6, 143. [Google Scholar] [CrossRef] [Green Version]
- Manaf, I.S.A.; Rahim, M.H.A.; Govindan, N.; Maniam, G.P. A first report on biodiesel production from Aglaia korthalsii seed oil using waste marine barnacle as a solid catalyst. Ind. Crops Prod. 2018, 125, 395–400. [Google Scholar] [CrossRef]
- Malek, M.N.F.A.; Veerappan, V.; Rahim, M.H.A.; Maniam, G.P. Various adsorbents to improve the filterability of biodiesel. Phys. Chem. Earth 2020, 120, 102910. [Google Scholar] [CrossRef]
- Manaf, I.S.A.; Yi, C.J.; Rahim, M.H.A.; Maniam, G.P. Utilization of waste fish bone as catalyst in transesterification of RBD palm oil. Mater. Today Proc. 2019, 19, 1294–1302. [Google Scholar] [CrossRef]
- Hogan, P.; Otero, P.; Murray, P.; Saha, S.K. Effect of biomass pre-treatment on supercritical CO2 extraction of lipids from marine diatom Amphora sp. and its biomass evaluation as bioethanol feedstock. Heliyon 2021, 7, e05995. [Google Scholar] [CrossRef]
- Hildebrand, M.; York, E.; Kelz, J.I.; Davis, A.K.; Frigeri, L.G.; Allison, D.P.; Doktycz, M.J. Nanoscale control of silica morphology and three-dimensional structure during diatom cell wall formation. J. Mater. Res. 2006, 21, 2689–2698. [Google Scholar] [CrossRef]
- Thamatrakoln, K.; Hildebrand, M. Analysis of Thalassiosira pseudonana silicon transporters indicates distinct regulatory levels and transport activity through the cell cycle. Eukaryot. Cell 2007, 6, 271–279. [Google Scholar] [CrossRef] [Green Version]
- Moore, E.R.; Bullington, B.S.; Weisberg, A.J.; Jiang, Y.; Chang, J.; Halsey, K.H. Morphological and transcriptomic evidence for ammonium induction of sexual reproduction in Thalassiosira pseudonana and other centric diatoms. PLoS ONE 2017, 12, e0181098. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.H.; Li, Y.Y.; Zhang, H.; Liu, J.L.; Xie, Z.X.; Lin, L.; Wang, D.Z. Quantitative proteomics reveals common and specific responses of a marine diatom Thalassiosira pseudonana to different macronutrient deficiencies. Front. Microbiol. 2018, 9, 2761. [Google Scholar] [CrossRef]
- Cheah, W.Y.; Show, P.L.; Juan, J.C.; Chang, J.S.; Ling, T.C. Microalgae cultivation in palm oil mill effluent (POME) for lipid production and pollutants removal. Energy Convers. Manag. 2018, 174, 430–438. [Google Scholar] [CrossRef]
- Mock, T.; Samanta, M.P.; Iverson, V.; Berthiaume, C.; Robison, M.; Holtermann, K.; Durkin, C.; BonDurant, S.S.; Richmond, K.; Rodesch, M.; et al. Whole-genome expression profiling of the marine diatom Thalassiosira pseudonana identifies genes involved in silicon bioprocesses. Proc. Natl. Acad. Sci. USA 2008, 105, 1579–1584. [Google Scholar] [CrossRef] [Green Version]
- Brzezinski, M.A.; Olson, R.J.; Chisholm, S.W. Silicon availability and cell-cycle progression in marine diatoms. Mar. Ecol. Prog. Ser. 1990, 67, 83–96. [Google Scholar] [CrossRef]
- Hildebrand, M.; Frigeri, L.G.; Davis, A.K. Synchronized growth of Thalassiosira pseudonana (Bacillariophyceae) provides novel insights into cell-wall synthesis processes in relation to the cell cycle. J. Phycol. 2007, 43, 730–740. [Google Scholar] [CrossRef]
- McNair, H.M.; Brzezinski, M.A.; Krause, J.W. Diatom populations in an upwelling environment decrease silica content to avoid growth limitation. Environ. Microbiol. 2018, 20, 4184–4193. [Google Scholar] [CrossRef]
- Amo, Y.D.; Brzezinski, M.A. The chemical form of dissolved Si taken up by marine diatoms. J. Phycol. 1999, 35, 1162–1170. [Google Scholar] [CrossRef]
- Cermeño, P.; Falkowski, P.G.; Romero, O.E.; Schaller, M.F.; Vallina, S.M. Continental erosion and the Cenozoic rise of marine diatoms. Proc. Natl. Acad. Sci. USA 2015, 112, 4239–4244. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, R.P.; Hildebrand, M. Evidence for a regulatory role of diatom silicon transporters in cellular silicon responses. Eukaryot. Cell 2015, 14, 29–40. [Google Scholar] [CrossRef] [Green Version]
- Lewin, J.C. Silicon metabolism in diatoms. II. Sources of silicon for growth of Navicula pelliculosa. Plant Physiol. 1955, 30, 129–134. [Google Scholar] [CrossRef]
- Sicko-Goad, L.; Andresen, N.A. Effect of growth and light/dark cycles on diatom lipid content and composition. J. Phycol. 1991, 27, 710–718. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Talmy, D.; Campbell, D.A. Diatom growth responses to photoperiod and light are predictable from diel reductant generation. J. Phycol. 2017, 53, 95–107. [Google Scholar] [CrossRef]
- Wu, Y.; Zeng, Y.; Qu, J.Y.; Wang, W.-X. Mercury effects on Thalassiosira weissflogii: Applications of two-photon excitation chlorophyll fluorescence lifetime imaging and flow cytometry. Aquatic Toxicol. 2012, 110, 133–140. [Google Scholar] [CrossRef]
- Chisholm, S.W.; Costello, J.C. Influence of environmental factors and population composition on the timing of cell division in Thalassiosira fluviatilis (Bacillariophyceae) grown on light/dark cycles. J. Phycol. 1980, 16, 375–383. [Google Scholar] [CrossRef]
- Nelson, D.M.; Brand, L.E. Cell division periodicity in 13 species of marine phytoplankton on a light: Dark cycle. J. Phycol. 1979, 15, 67–75. [Google Scholar] [CrossRef]
- Brand, L.E.; Guillard, R.R.L. The effects of continuous light and light intensity on the reproduction rates of twenty-two species of marine phytoplankton. J. Exp. Mar. Biol. Ecol. 1981, 50, 119–132. [Google Scholar] [CrossRef]
- Li, G.; Woroch, A.D.; Donaher, N.A.; Cockshutt, A.M.; Campbell, D.A. A hard day’s night: Diatoms continue recycling photosystem II in the dark. Front. Mar. Sci. 2016, 3, 218. [Google Scholar] [CrossRef] [Green Version]
- Camacho Rubio, F.; García Camacho, F.; Fernández Sevilla, J.M.; Chisti, Y.; Molina Grima, E. A mechanistic model of photosynthesis in microalgae. Biotechnol. Bioeng. 2003, 81, 459–473. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Wu, S.; Gu, W.; Wang, L.; Wang, J.; Gao, S.; Wang, G. Photosynthesis acclimation under severely fluctuating light conditions allows faster growth of diatoms compared with dinoflagellates. BMC Plant Biol. 2021, 21, 164. [Google Scholar] [CrossRef]
- Zachleder, V.; Bišová, K.; Vítová, M. The cell cycle of microalgae. In The Physiology of Microalgae; Borowitzka, M., Beardall, J., Raven, J., Eds.; Springer: Cham, Switzerland, 2016; pp. 3–46. [Google Scholar] [CrossRef]
- Jacquet, S.; Partensky, F.; Lennon, J.F.; Vaulot, D. Diel patterns of growth and division in marine picoplankton in culture. J. Phycol. 2001, 37, 357–369. [Google Scholar] [CrossRef] [Green Version]
- Joseph, M.M.; Renjith, K.R.; John, G.; Nair, S.M.; Chandramohanakumar, N. Biodiesel prospective of five diatom strains using growth parameters and fatty acid profiles. Biofuels 2017, 8, 81–89. [Google Scholar] [CrossRef]
- Ying, L.; Mai, K.-S.; Sun, S.-C. Total lipid and fatty acid composition of eight strains of marine diatoms. Chin. J. Oceanol. Limnol. 2000, 18, 345–349. [Google Scholar] [CrossRef]
- Chen, Y.-C. The biomass and total lipid content and composition of twelve species of marine diatoms cultured under various environments. Food Chem. 2012, 131, 211–219. [Google Scholar] [CrossRef]
- Fisher, N.S.; Schwarzenbach, R.P. Fatty acid dynamics in Thalassiosira pseudonana (Bacillariophyceae): Implications for physiological ecology. J. Phycol. 1978, 14, 143–150. [Google Scholar] [CrossRef]
- Bromke, M.A.; Giavalisco, P.; Willmitzer, L.; Hesse, H. Metabolic analysis of adaptation to short-term changes in culture conditions of the marine diatom Thalassiosira pseudonana. PLoS ONE 2013, 8, e67340. [Google Scholar] [CrossRef]
- Hunter, J.E.; Brandsma, J.; Dymond, M.K.; Koster, G.; Moore, C.M.; Postle, A.D.; Mills, R.A.; Attard, G.S. Lipidomics of Thalassiosira pseudonana under phosphorus stress reveal underlying phospholipid substitution dynamics and novel diglycosylceramide substitutes. Appl. Environ. Microbiol. 2018, 84, e02034-17. [Google Scholar] [CrossRef] [Green Version]
- Boelen, P.; van Mastrigt, A.; van de Bovenkamp, H.H.; Heeres, H.J.; Buma, A.G.J. Growth phase significantly decreases the DHA-to-EPA ratio in marine microalgae. Aquacult. Int. 2017, 25, 577–587. [Google Scholar] [CrossRef] [Green Version]
- Cook, O.; Hildebrand, M. Enhancing LC-PUFA production in Thalassiosira pseudonana by overexpressing the endogenous fatty acid elongase genes. J. Appl. Phycol. 2016, 28, 897–905. [Google Scholar] [CrossRef]
- Tonon, T.; Qing, R.; Harvey, D.; Li, Y.; Larson, T.R.; Graham, I.A. Identification of a long-chain polyunsaturated fatty acid acyl-coenzyme A synthetase from the diatom Thalassiosira pseudonana. Plant Physiol. 2005, 138, 402–408. [Google Scholar] [CrossRef] [Green Version]
- Mortensen, S.H.; Børsheim, K.Y.; Rainuzzo, J.R.; Knutsen, G. Fatty acid and elemental composition of the marine diatom Chaetoceros gracilis Schütt. Effects of silicate deprivation, temperature and light intensity. J. Exp. Mar. Biol. Ecol. 1988, 122, 173–185. [Google Scholar] [CrossRef]
- Shene, C.; Paredes, P.; Flores, L.; Leyton, A.; Asenjo, J.A.; Chisti, Y. Dynamic flux balance analysis of biomass and lipid production by Antarctic thraustochytrid Oblongichytrium sp. RT2316-13. Biotechnol. Bioeng. 2020, 117, 3006–3017. [Google Scholar] [CrossRef]
- Opute, F.I. Lipid and fatty-acid composition of diatoms. J. Exp. Bot. 1974, 25, 823–835. [Google Scholar] [CrossRef]
- Sicko-Goad, L.; Simmons, M.S.; Lazinsky, D.; Hall, J. Effect of light cycle on diatom fatty acid composition and quantitative morphology. J. Phycol. 1988, 24, 1–7. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Parameter | Value |
|---|---|
| pH | 5.2 ± 0.9 |
| Ammoniacal nitrogen, NH3-N (mg L−1) | 16.3 ± 2.1 |
| Biochemical oxygen demand, BOD (mg L−1) | 2600 ± 325 |
| Chemical oxygen demand, COD (mg L−1) | 2664 ± 21 |
| Total nitrogen (mg L−1) | 141.4 ± 15.3 |
| Total phosphorus (mg L−1) | 21.5 ± 1.4 |
| Total suspended solids (mg L−1) | 94.6 ± 5.3 |
| Experiment Set a | POME (% v/v) |
|---|---|
| A (40 μM Na2SiO3) | 0 (control) b |
| 10 | |
| 20 | |
| 30 | |
| B (80 μM Na2SiO3) | 0 (control) b |
| 10 | |
| 20 | |
| 30 | |
| C (120 μM Na2SiO3) | 0 (control) b |
| 10 | |
| 20 | |
| 30 | |
| D (160 μM Na2SiO3) | 0 (control) |
| 10 | |
| 20 | |
| 30 |
| Parameter | Light–Dark Cycle (h:h) | |||
|---|---|---|---|---|
| 24:0 | 16:8 | 12:12 | 8:16 | |
| Biomass concentration (mg L−1) | 812 ± 11 a | 869 ± 14 b | 791 ± 15 c | 451 ± 12 d |
| Biomass productivity (mg L−1 d−1) | 54.1 ± 0.7 a | 57.9 ± 0.9 b | 52.7 ± 1.0 a | 30.1 ± 2.7 c |
| Lipids in biomass (%, w/w) | 38.1 ± 0.8 a | 46.2 ± 1.1 b | 35.7 ± 0.9 c | 12.6 ± 1.1 d |
| Fatty Acid Class | This Work b | Mai et al. [48] c | Volkman et al. [49] d |
|---|---|---|---|
| Saturated | 40.0 | 22.8 | 27.4 |
| Monounsaturated | 48.9 | 30.0 | 19.6 |
| Polyunsaturated | 11.1 | 47.2 | 53.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palanisamy, K.M.; Maniam, G.P.; Sulaiman, A.Z.; Ab. Rahim, M.H.; Govindan, N.; Chisti, Y. Palm Oil Mill Effluent for Lipid Production by the Diatom Thalassiosira pseudonana. Fermentation 2022, 8, 23. https://doi.org/10.3390/fermentation8010023
Palanisamy KM, Maniam GP, Sulaiman AZ, Ab. Rahim MH, Govindan N, Chisti Y. Palm Oil Mill Effluent for Lipid Production by the Diatom Thalassiosira pseudonana. Fermentation. 2022; 8(1):23. https://doi.org/10.3390/fermentation8010023
Chicago/Turabian StylePalanisamy, Karthick Murugan, Gaanty Pragas Maniam, Ahmad Ziad Sulaiman, Mohd Hasbi Ab. Rahim, Natanamurugaraj Govindan, and Yusuf Chisti. 2022. "Palm Oil Mill Effluent for Lipid Production by the Diatom Thalassiosira pseudonana" Fermentation 8, no. 1: 23. https://doi.org/10.3390/fermentation8010023