
Current Advances in Microbial Production of Acetoin and 2,3-Butanediol by Bacillus spp.



Abstract
:1. Introduction
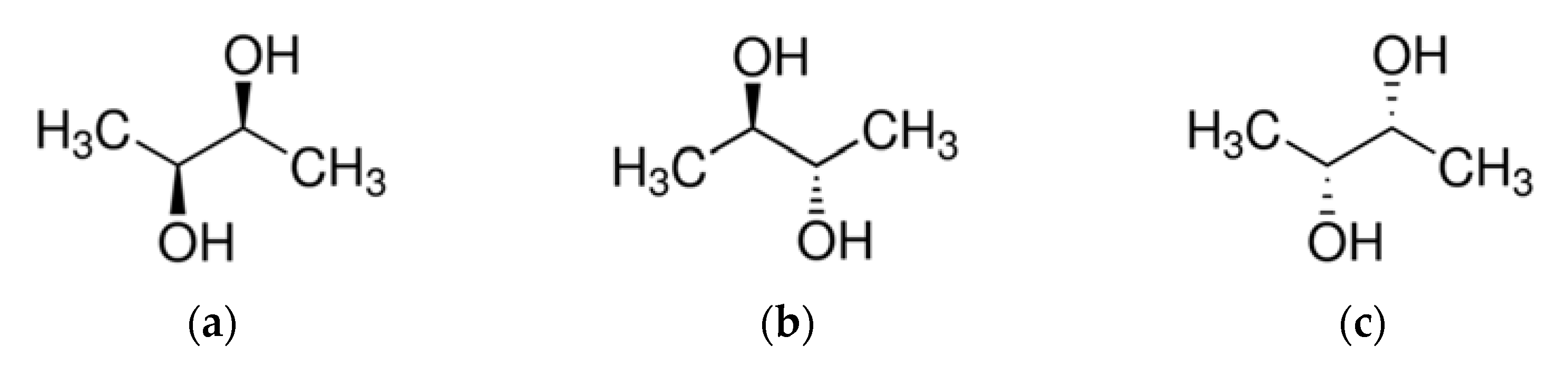
2. Chemical Structure of Acetoin and 2,3-Butanediol and Their Stereoisomers
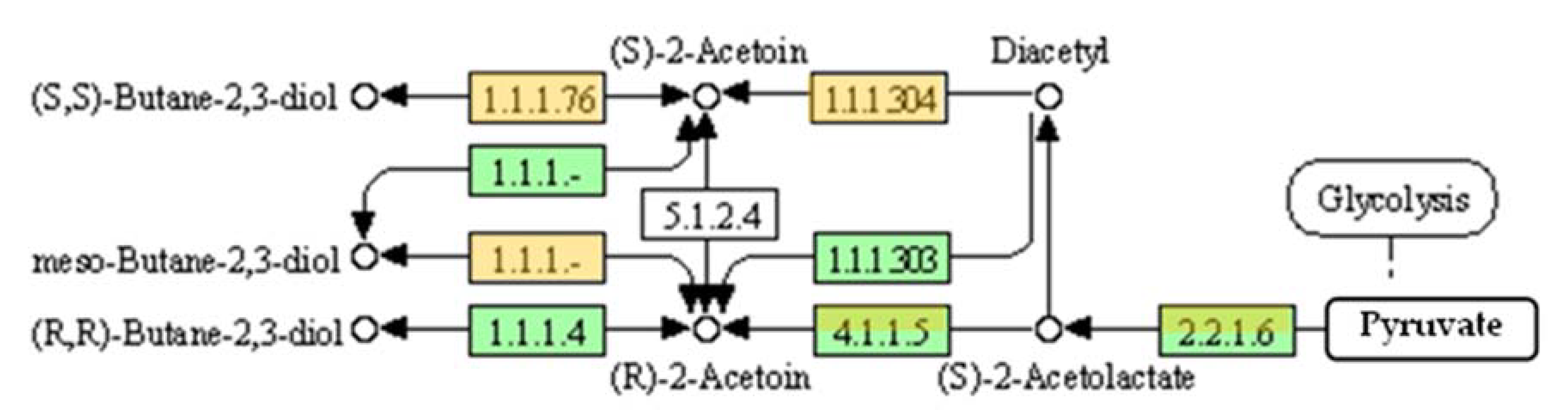
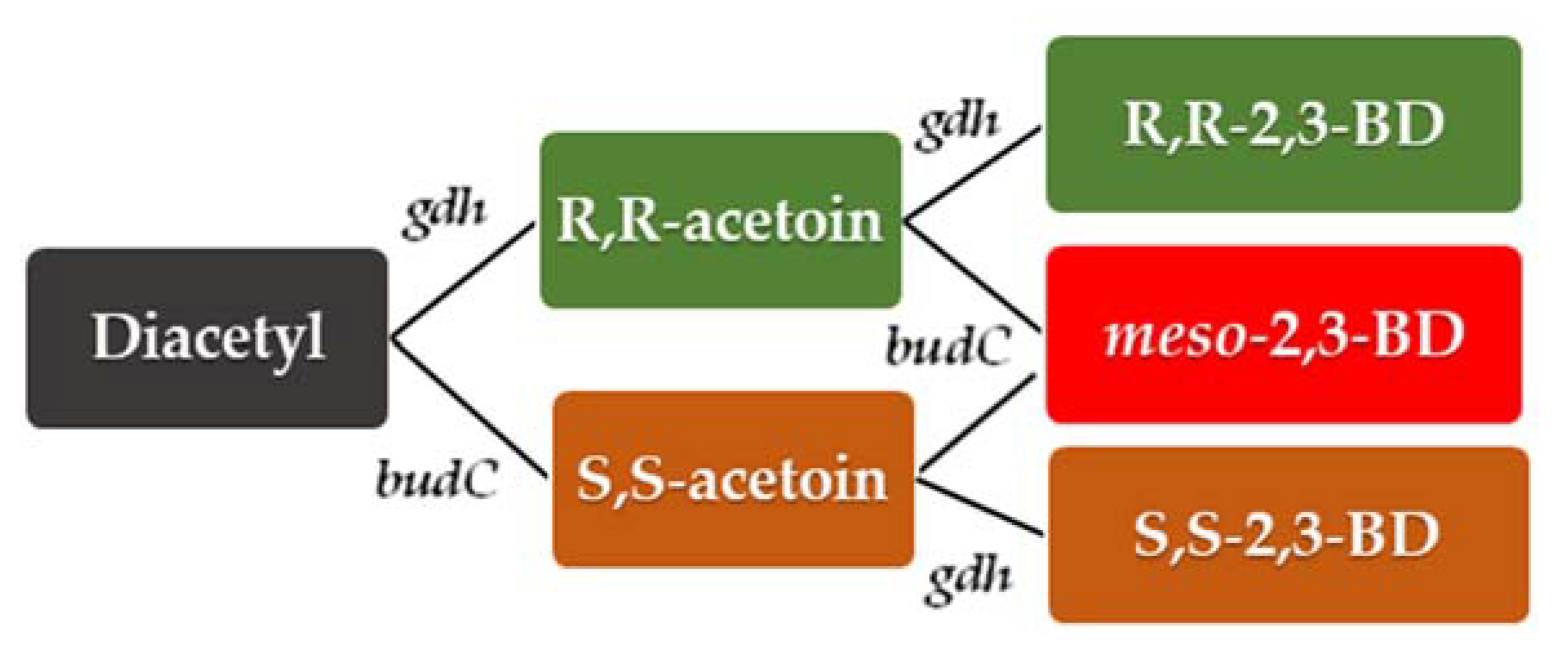
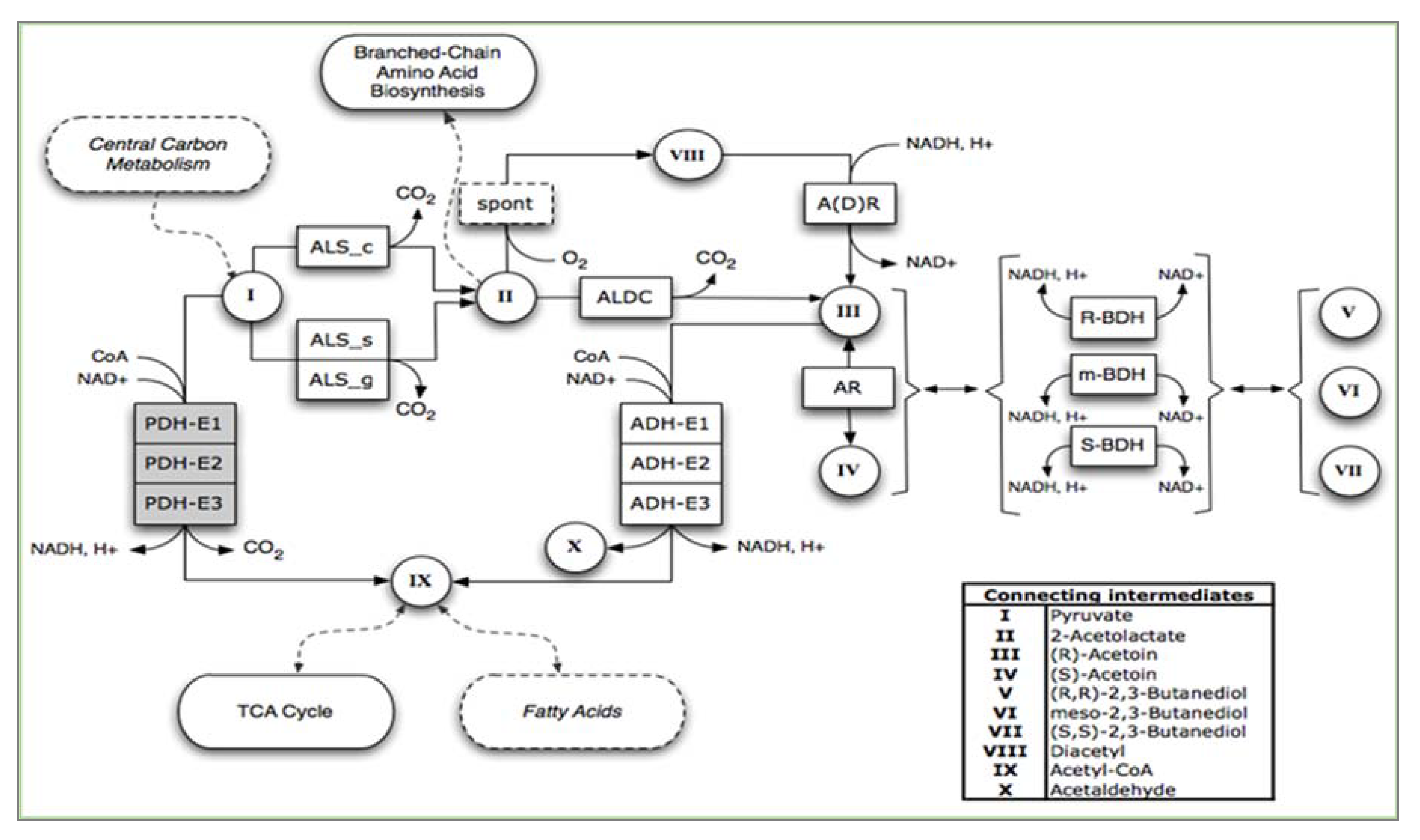
3. Metabolic Pathway for Acetoin and 2,3-Butanediol Synthesis by Bacillus spp.
4. Natural Producers of Acetoin and 2,3-BD
4.1. B. licheniformis
4.2. B. subtilis
4.3. B. amyloliquefaciens
4.4. Other 2,3-BD and Acetoin Producers
4.4.1. Bacillus spp.
4.4.2. Paenibacillus spp.
5. Engineered Producers of 2,3-BD and Acetoin
5.1. Engineered Producers of 2,3-BD
5.2. Engineered Producers of Acetoin
6. Substrates
7. Media and Process Parameters for 2,3-BD and Acetoin Production by Bacilli
7.1. Medium Content
7.2. Process Parameters
7.2.1. Temperature
7.2.2. pH
7.2.3. Aeration
7.3. By-Products Reduction Strategies
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, J.H.; Lee, D.Y.; Lee, S.K.; Kim, H.R.; Chun, Y.; Yoo, H.Y.; Kwak, H.S.; Park, C.; Lee, J.H.; Kim, S.W. Development of 2,3-Butanediol Production Process from Klebsiella aerogenes ATCC 29007 Using Extracted Sugars of Chlorella pyrenoidosa and Biodiesel-Derived Crude Glycerol. Processes 2021, 9, 517. [Google Scholar] [CrossRef]
- Tian, Y.; Fan, Y.; Liu, J.; Zhao, X.; Chen, W. Effect of nitrogen, carbon sources and agitation speed on acetoin production of Bacillus subtilis SF4-3. Electron. J. Biotechnol. 2016, 19, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.; Zhang, L.; Xie, Y.; Hu, C.; Zhang, Y.; Li, L.; Wang, Y.; Ma, C.; Xu, P. Production of (3S)-acetoin from diacetyl by using stereoselective NADPH-dependent carbonyl reductase and glucose dehydrogenase. Bioresour. Technol. 2013, 137, 111–115. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Chen, F.; Sun, M.; Gao, H.; Guo, Z.; Lin, H.; Chen, J.; Jin, W.; Yang, Y.; Zhang, L.; et al. Efficient (3S)-Acetoin and (2S,3S)-2,3-Butanediol Production from meso-2,3-Butanediol Using Whole-Cell Biocatalysis. Molecules 2018, 23, 691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maina, S.; Schneider, R.; Alexandri, M.; Papapostolou, H.; Nychas, G.-J.; Koutinas, A.; Venus, J. Volumetric oxygen transfer coefficient as fermentation control parameter to manipulate the production of either acetoin or D(−)2,3-butanediol using bakery waste. Bioresour. Technol. 2021, 335, 125155. [Google Scholar] [CrossRef] [PubMed]
- Global Acetoin Market 2018–2023. Available online: https://menafn.com/1099300202/Global-Acetoin-Market-2018–2023 (accessed on 3 November 2021).
- Zhang, X.; Yang, T.-W.; Lin, Q.; Xu, M.-J.; Xia, H.-F.; Xu, Z.-H.; Li, H.-Z.; Rao, Z.-M. Isolation and identification of an acetoin high production bacterium that can reverse transform 2,3-butanediol to acetoin at the decline phase of fermentation. World J. Microbiol. Biotechnol. 2011, 27, 2785–2790. [Google Scholar] [CrossRef]
- Zhang, B.; Li, X.-L.; Fu, J.; Li, N.; Wang, Z.; Tang, Y.-J.; Chen, T. Production of Acetoin through Simultaneous Utilization of Glucose, Xylose, and Arabinose by Engineered Bacillus subtilis. PLoS ONE 2016, 11, e0159298. [Google Scholar] [CrossRef]
- Maina, S.; Prabhu, A.A.; Vivek, N.; Vlysidis, A.; Koutinas, A.; Kumar, V. Prospects on bio-based 2,3-butanediol and acetoin production: Recent progress and advances. Biotechnol. Adv. 2021, 107783. [Google Scholar] [CrossRef]
- Cui, X.; Zhao, X.; Liu, D. A novel route for the flexible preparation of hydrocarbon jet fuels from biomass-based platform chemicals: A case of using furfural and 2,3- butanediol as feedstocks. Green Chem. 2018, 20, 2018–2026. [Google Scholar] [CrossRef]
- Xiao, Z.; Xu, P. Acetoin Metabolism in Bacteria. Crit. Rev. Microbiol. 2007, 33, 127–140. [Google Scholar] [CrossRef]
- Hilmi, A.; Belgsir, E.M.; Leger, J.M.; Lamy, C. Electrocatalytic oxidation of aliphatic diols Part V. Electro-oxidation of butanediols on platinum-based electrodes. J. Electroanal. Chem. 1997, 435, 69–75. [Google Scholar] [CrossRef]
- Kochius, S.; Paetzold, M.; Scholz, A.; Merkens, H.; Vogel, A.; Ansorge-Schumacher, M.; Hollmann, F.; Schrader, J.; Holtmann, D. Enantioselective enzymatic synthesis of the α-hydroxy ketone (R)-acetoin from meso-2,3-butanediol. J. Mol. Catal. Enzym. 2014, 103, 61–66. [Google Scholar] [CrossRef]
- Garg, S.K.; Jain, A. Fermentative production of 2,3-butanediol: A review. Bioresour. Technol. 1995, 51, 103–109. [Google Scholar] [CrossRef]
- Celinska, E.; Grajek, W. Biotechnological production of 2,3-butanediol current state and prospects. Biotechnol. Adv. 2009, 27, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Cho, S.-Y.; Vu, T.-T.; Duong, H.-P.-Y.; Kim, E. Dehydration of 2,3-Butanediol to 1,3-Butadiene and Methyl Ethyl Ketone: Modeling, Numerical Analysis and Validation Using Pilot-Scale Reactor Data. Catalysts 2021, 11, 999. [Google Scholar] [CrossRef]
- Yu, E.K.; Saddler, J.N. Power of solvent production by Klebsiella pneumoniae grown on sugars present in wood hemicelluloses. Biotechnol. Lett. 1982, 4, 121–126. [Google Scholar] [CrossRef]
- Hazeena, S.H.; Sindhu, R.; Pandey, A.; Binod, P. Lignocellulosic bio-refinery approach for microbial 2,3-Butanediol production. Bioresour. Technol. 2020, 302, 122873. [Google Scholar] [CrossRef] [PubMed]
- Petrova, P.; Ivanov, I.; Tsigoriyna, L.; Valcheva, N.; Vasileva, E.; Parvanova-Mancheva, T.; Arsov, A.; Petrov, K. Traditional Bulgarian Dairy Products: Ethnic Foods with Health Benefits. Microorganisms 2021, 9, 480. [Google Scholar] [CrossRef] [PubMed]
- Petrini, P.; De Ponti, S.; Farea, S.; Tanzi, M.C. Polyurethane-maleamides for cardiovascular applications: Synthesis and properties. J. Mater. Sci.-Mater. Med. 1999, 10, 711–714. [Google Scholar] [CrossRef] [PubMed]
- Bartowsky, E.J.; Henschke, P.A. The ‘buttery’ attribute of wine-diacetyl-desirability, spoilage and beyond. Int. J. Food Microbiol. 2004, 96, 235–252. [Google Scholar] [CrossRef]
- Ji, X.J.; Huang, H.; Ouyang, P.K. Microbial 2,3-butanediol production: A state of the art review. Biotechnol. Adv. 2011, 29, 351–364. [Google Scholar] [CrossRef] [PubMed]
- Syu, M.J. Biological production of 2,3-butanediol. Appl. Microbiol. Biotechnol. 2001, 55, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Anchordoguy, T.J.; Cecchini, C.A.; Crowe, J.H.; Crowe, L.M. Insights into the cryoprotective mechanism of dimethyl sulfoxide for phospholipid bilayers. Cryobiology 1991, 28, 467–473. [Google Scholar] [CrossRef]
- Armitage., W.J.; Juss, B.K. Osmotic response of mammalian cells: Effects of permeating cryoprotectants on nonsolvent volume. J. Cell. Physiol. 1996, 168, 532–538. [Google Scholar] [CrossRef]
- Petrov, K.; Petrova, P. High production of 2,3-butanediol from glycerol by Klebsiella pneumoniae G31. Appl. Microbiol. Biotechnol. 2009, 84, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Levante, A.; Bancalari, E.; Tambassi, M.; Lazzi, C.; Neviani, E.; Gatti, M. Phenotypic Diversity of Lactobacillus casei Group Isolates as a Selection Criterion for Use as Secondary Adjunct Starters. Microorganisms 2020, 8, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Z.; Lu, J.R. Strategies for enhancing fermentative production of acetoin: A review. Biotechnol. Adv. 2014, 32, 492–503. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.Y.; Lee, S.H.; Lee, H.J.; Seo, H.Y.; Park, W.S.; Jeon, C.O. Effects of Leuconostoc mesenteroides starter cultures on microbial communities and metabolites during kimchi fermentation. Int. J. Food Microbiol. 2012, 53, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Laroute, V.; Tormo, H.; Couderc, C.; Mercier-Bonin, M.; Le Bourgeois, P.; Cocaign-Bousquet, M.; Daveran-Mingot, M.-L. From Genome to Phenotype: An Integrative Approach to Evaluate the Biodiversity of Lactococcus lactis. Microorganisms 2017, 5, 27. [Google Scholar] [CrossRef]
- Cañas, A.; Owens, J.D. Acetoin production in growing Leuconostoc mesenteroides. World J. Microbiol. Biotechnol. 1999, 62, 339–344. [Google Scholar] [CrossRef]
- Humia, B.V.; Santos, K.S.; Barbosa, A.M.; Sawata, M.; Mendonça, M.d.C.; Padilha, F.F. Beer Molecules and Its Sensory and Biological Properties: A Review. Molecules 2019, 24, 1568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romano, P.; Suzzi, G.; Zironi, R.; Comi, G. Biometric study of acetoin production in Hanseniaspora guilliermondii and Kloeckera apiculata. Appl. Environ. Microbiol. 1993, 59, 1838–1841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, R.M.; Cavalheiro, D.; Ninow, J.L.; Furigo, A.J. Optimization of acetoin production by Hanseniaspora guilliermondii using experimental design. Braz. J. Chem. Eng. 2002, 19, 181–186. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhang, S.; Yong, Y.-C.; Ji, Z.; Ma, X.; Xu, Z.; Chen, S. Efficient production of acetoin by the newly isolated Bacillus licheniformis strain MEL09. Process Biochem. 2011, 46, 390–394. [Google Scholar] [CrossRef]
- Harden, A.; Walpole, G.S. 2,3-butylene glycol fermentation by Aerobacter aerogenes. Proc. R. Soc. 1906, 77, 399–405. [Google Scholar]
- Fulmer, E.I.; Christensen, L.M.; Kendall, A.R. Production of 2,3-butylene glycol by fermentation. Ind. Eng. Chem. 1933, 25, 798–800. [Google Scholar] [CrossRef]
- Voloch, M.; Jansen, N.B.; Ladish, M.R.; Tsao, G.T.; Narayan, R.; Rodwell, V.W. 2,3-butanediol. In Comprehensive Biotechnology: The Principles, Applications, and Regulations of Biotechnology in Industry, Agriculture, and Medicine, 1st ed.; Blanch, H.W., Drew, S., Wang, D.I.C., Eds.; Pergamon Press: Oxford, UK, 1985; pp. 933–944. [Google Scholar]
- Li, Z.J.; Ji, X.J.; Kan, S.L.; Qiao, H.Q.; Jiang, M.; Lu, D.; Wang, J.; Huang, H.; Jia, H.; Ouyuang, P.; et al. Past, present, and future industrial biotechnology in China. Adv. Biochem. Eng. Biotechnol. 2010, 122, 1–42. [Google Scholar] [CrossRef]
- Petrov, K.; Petrova, P. Isolation and molecular identification of Klebsiella pneumoniae strains, producing diols from glycerol. Biotechnol. Biotechnol. Equip. 2009, 23, 814–817. [Google Scholar] [CrossRef] [Green Version]
- Tsvetanova, F.; Petrova, P.; Petrov, K. 2,3-butanediol production from starch by engineered Klebsiella pneumoniae G31-A. Appl. Microbiol. Biotechnol. 2014, 98, 2441–2451. [Google Scholar] [CrossRef] [PubMed]
- Petrov, K.; Petrova, P. Enhanced production of 2,3-butanediol from glycerol by forced pH fluctuations. Appl. Microbiol. Biotechnol. 2010, 87, 943–949. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Guo, W.; Shi, L.; Yu, Y.; Zhang, C.; Yang, H. Characterization of acetoin production in a budC gene disrupted mutant of Serratia marcescens G12. J. Ind. Microbiol. Biotechnol. 2014, 41, 1267–1274. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Yang, H.H.; Feng, X.H.; Li, S.; Xu, H. A 2,3-butanediol dehydrogenase from Paenibacillus polymyxa ZJ-9 for mainly producing R,R-2,3-butanediol: Purification, characterization and cloning. J. Basic Microbiol. 2013, 53, 733–741. [Google Scholar] [CrossRef]
- Ge, Y.; Li, K.; Li, L.; Gao, C.; Zhang, L.; Ma, C.; Xu, P. Contracted but effective production of enantiopure 2,3-butanediol by thermophilic and GRAS Bacillus licheniformis. Green Chem. 2016, 18, 4693–4703. [Google Scholar] [CrossRef]
- Banoon, S.; Ali, Z.; Salih, T. Antibiotic resistance profile of local thermophilic Bacillus licheniformis isolated from Maysan province soil. Comun. Sci. 2020, 11, e3291. [Google Scholar] [CrossRef]
- Šovljanski, O.; Pezo, L.; Stanojev, J.; Bajac, B.; Kovač, S.; Tóth, E.; Ristić, I.; Tomić, A.; Ranitović, A.; Cvetković, D.; et al. Comprehensive Profiling of Microbiologically Induced CaCO3 Precipitation by Ureolytic Bacillus Isolates from Alkaline Soils. Microorganisms 2021, 9, 1691. [Google Scholar] [CrossRef]
- Mahdi, I.; Fahsi, N.; Hafidi, M.; Allaoui, A.; Biskri, L. Plant Growth Enhancement using Rhizospheric Halotolerant Phosphate Solubilizing Bacterium Bacillus licheniformis QA1 and Enterobacter asburiae QF11 Isolated from Chenopodium quinoa Willd. Microorganisms 2020, 8, 948. [Google Scholar] [CrossRef] [PubMed]
- Vinothkanna, A.; Sathiyanarayanan, G.; Balaji, P.; Mathivanan, K.; Pugazhendhi, A.; Ma, Y.; Sekar, S.; Thirumurugan, R. Structural characterization, functional and biological activities of an exopolysaccharide produced by probiotic Bacillus licheniformis AG-06 from Indian polyherbal fermented traditional medicine. Int. J. Biol. Macromol. 2021, 174, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Hirad, A.; Bahkali, A.; Khiyami, M.; Ahmed, M.; Santhapa, C.; Elgorban, A.; Al-Sum, B. Bioactivity of Marine Bacillus licheniformis Ksawd3 Isolated from Arabian Gulf, Saudi Arabia. J. Pure Appl. Microbiol. 2014, 8, 607–613. [Google Scholar]
- Biswas, J.K.; Banerjee, A.; Sarkar, B.; Sarkar, D.; Sarkar, S.K.; Rai, M.; Vithanage, M. Exploration of an Extracellular Polymeric Substance from Earthworm Gut Bacterium (Bacillus licheniformis) for Bioflocculation and Heavy Metal Removal Potential. Appl. Sci. 2020, 10, 349. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Hou, Q.; Guo, Q.; Zhang, J.; Sun, Y.; Wei, H.; Shen, L. Isolation and Characterization of a Deoxynivalenol-Degrading Bacterium Bacillus licheniformis YB9 with the Capability of Modulating Intestinal Microbial Flora of Mice. Toxins 2020, 12, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrova, P.; Arsov, A.; Ivanov, I.; Tsigoriyna, L.; Petrov, K. New Exopolysaccharides Produced by Bacillus licheniformis 24 Display Substrate-Dependent Content and Antioxidant Activity. Microorganisms 2021, 9, 2127. [Google Scholar] [CrossRef] [PubMed]
- Ilyas, N.; Mumtaz, K.; Akhtar, N.; Yasmin, H.; Sayyed, R.Z.; Khan, W.; Enshasy, H.A.E.; Dailin, D.J.; Elsayed, E.A.; Ali, Z. Exopolysaccharides Producing Bacteria for the Amelioration of Drought Stress in Wheat. Sustainability 2020, 12, 8876. [Google Scholar] [CrossRef]
- Kwon, J.-H.; Won, S.-J.; Moon, J.-H.; Lee, U.; Park, Y.-S.; Maung, C.E.H.; Ajuna, H.B.; Ahn, Y.S. Bacillus licheniformis PR2 Controls Fungal Diseases and Increases Production of Jujube Fruit under Field Conditions. Horticulturae 2021, 7, 49. [Google Scholar] [CrossRef]
- Matei, M.-C.; Andrei, S.M.; Buza, V.; Cernea, M.S.; Dumitras, D.A.; Neagu, D.; Rafa, H.; Popovici, C.P.; Szakacs, A.R.; Catinean, A.; et al. Natural Endotoxemia in Dogs—A Hidden Condition That Can Be Treated with a Potential Probiotic Containing Bacillus subtilis, Bacillus licheniformis, and Pediococcus acidilactici: A Study Model. Animals 2021, 11, 1367. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Qin, J.; Gao, C.; Hua, D.; Ma, C.; Li, L.; Wang, Y.; Xu, P. Production of (2S,3S)-2,3-butanediol and (3S)-acetoin from glucose using resting cells of Klebsiella pneumonia and Bacillus subtilis. Bioresour. Technol. 2011, 102, 10741–10744. [Google Scholar] [CrossRef]
- Wang, D.; Oh, B.R.; Lee, S.; Kim, D.-H.; Joe, M.-H. Process optimization for mass production of 2,3-butanediol by Bacillus subtilis CS13. Biotechnol. Biofuels 2021, 14, 15. [Google Scholar] [CrossRef] [PubMed]
- Marwoto, B.; Nakashimada, Y.; Kakizono, T.; Nishio, N. Metabolic analysis of acetate accumulation during xylose consumption by Paenibacillus polymyxa. Appl. Microbiol. Biotechnol. 2004, 64, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Okonkwo, C.C.; Ujor, V.; Ezeji, T.C. Investigation of relationship between 2,3-butanediol toxicity and production during growth of Paenibacillus polymyxa. New Biotechnol. 2017, 34, 23–31. [Google Scholar] [CrossRef]
- Okonkwo, C.C.; Ujor, V.C.; Mishra, P.K.; Ezeji, T.C. Process Development for Enhanced 2,3-Butanediol Production by Paenibacillus polymyxa DSM 365. Fermentation 2017, 3, 18. [Google Scholar] [CrossRef] [Green Version]
- Häßler, T.; Schieder, D.; Pfaller, R.; Faulstich, M.; Sieber, V. Enhanced fed-batch fermentation of 2,3-butanediol by Paenibacillus polymyxa DSM 365, Bioresour. Technol. 2012, 124, 237–244. [Google Scholar] [CrossRef]
- Yang, T.; Rao, Z.; Zhang, X.; Lin, Q.; Xia, H.; Xu, Z.; Yang, S. Production of 2,3-butanediol from glucose by GRAS microorganism Bacillus amyloliquefaciens. J. Basic Microbiol. 2011, 51, 650–658. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, S.; Liu, L.; Wu, J. Acetoin production enhanced by manipulating carbon flux in a newly isolated Bacillus amyloliquefaciens. Bioresour. Technol. 2013, 130, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Nilegaonkar, S.; Bhosale, S.; Kshirsagar, D.; Kapadi, A. Production of 2,3-butanediol from glucose by Bacillus licheniformis. World J. Microbiol. Biotechnol. 1992, 8, 378–381. [Google Scholar] [CrossRef] [PubMed]
- Nilegaonkar, S.S.; Bhosale, S.B.; Dandage, C.N.; Kapadi, A.H. Potential of Bacillus licheniformis for the production of 2,3-butanediol. J. Ferment. Bioeng. 1996, 82, 408–410. [Google Scholar] [CrossRef]
- Tsigoriyna, L.; Ganchev, D.; Petrova, P.; Petrov, K. Highly Efficient 2,3-Butanediol Production by Bacillus licheniformis via Complex Optimization of Nutritional and Technological Parameters. Fermentation 2021, 7, 118. [Google Scholar] [CrossRef]
- Perego, P.; Converti, A.; del Borghi, M. Effects of temperature, inoculum size and starch hydrolyzate concentration on butanediol production by Bacillus licheniformis. Bioresour. Technol. 2003, 89, 125–131. [Google Scholar] [CrossRef]
- Petrova, P.; Petlichka, S.; Petrov, K. New Bacillus spp. with potential for 2,3-butanediol production from biomass. J. Biosci. Bioeng. 2020, 130, 20–28. [Google Scholar] [CrossRef]
- Li, L.; Zhang, L.; Li, K.; Wang, Y.; Gao, C.; Han, B.; Ma, C.; Xu, P. A newly isolated Bacillus licheniformis strain thermophilically produces 2,3-butanediol, a platform and fuel bio-chemical. Biotechnol. Biofuels 2013, 6, 123. [Google Scholar] [CrossRef] [Green Version]
- Jurchescu, I.M.; Hamann, J.; Zhou, X.; Ortmann, T.; Kuenz, A.; Prusse, U.; Lang, S. Enhanced 2,3-butanediol production in fed batch cultures of free and immobilized Bacillus licheniformis DSM 8785. Appl. Microbiol. Biotechnol. 2013, 97, 6715–6723. [Google Scholar] [CrossRef] [PubMed]
- Lü, C.; Ge, Y.; Cao, M.; Guo, X.; Liu, P.; Gao, C.; Xu, P.; Ma, C. Metabolic Engineering of Bacillus licheniformis for Production of Acetoin. Front. Bioeng. Biotechnol. 2020, 8, 125. [Google Scholar] [CrossRef]
- Li, L.X.; Li, K.; Wang, K.; Chen, C.; Gao, C.; Ma, C.Q.; Xu, P. Efficient production of 2,3-butanediol from corn stover hydrolysate by using a thermophilic Bacillus licheniformis strain. Bioresour. Technol. 2014, 170, 256–261. [Google Scholar] [CrossRef]
- Rebecchi, S.; Pinelli, D.; Zanaroli, G.; Fava, F.; Frascari, D. Effect of oxygen mass transfer rate on the production of 2,3-butanediol from glucose and agro-industrial byproducts by Bacillus licheniformis ATCC 9789. Biotechnol. Biofuels 2018, 11, 145. [Google Scholar] [CrossRef] [PubMed]
- Song, C.W.; Rathnasingh, C.; Park, J.M.; Lee, J.; Song, H. Isolation and evaluation of Bacillus strains for industrial production of 2,3-butanediol. J. Microbiol. Biotechnol. 2018, 28, 409–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kallbach, M.; Horn, S.; Kuenz, A.; Prusse, U. Screening of novel bacteria for the 2,3-butanediol production. Appl. Microbiol. Biotechnol. 2017, 101, 1025–1033. [Google Scholar] [CrossRef]
- PubChem Database at the National Institutes of Health (NIH). Available online: https://pubchem.ncbi.nlm.nih.gov (accessed on 3 November 2021).
- Maina, S.; Mallouchos, A.; Nychas, G.-J.E.; Freire, D.M.; de Castro, A.M.; Papanikolaou, S.; Kookos, I.K.; Koutinas, A. Bioprocess development for (2R,3R)-butanediol and acetoin production using very high polarity cane sugar and sugarcane molasses by a Bacillus amyloliquefaciens strain. J. Chem. Technol. Biotechnol. 2019, 94, 2167–2177. [Google Scholar] [CrossRef]
- KEGG BRITE Database. Available online: https://www.genome.jp/kegg/brite.html (accessed on 6 November 2021).
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 2014, 42, D206–D214. [Google Scholar] [CrossRef] [PubMed]
- Petrova, P.; Velikova, P.; Petrov, K. Genome sequence of Bacillus velezensis 5RB, an overproducer of 2,3-butanediol. Microbiol. Resour. Announc. 2019, 8, e01475-18. [Google Scholar] [CrossRef] [Green Version]
- Voloch, M.; Ladish, M.R.; Rodwell, V.W.; Tsao, G.T. Reduction of acetoin to 2,3-butanediol in Klebsiella pneumoniae: A new model. Biotechnol. Bioeng. 1983, 25, 173–183. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, Y.; Yang, D.C.; Zhang, J.L. Expression and functional analysis of three nicosulfuron-degrading enzymes from Bacillus subtilis YB1. J. Environ. Sci. Health B 2018, 53, 476–485. [Google Scholar] [CrossRef]
- Gao, H.; Jiang, X.; Pogliano, K.; Aronson, A.I. The E1-beta and E2 subunits of the Bacillus subtilis pyruvate dehydrogenase complex are involved in regulation of sporulation. J. Bacteriol. 2002, 184, 2780–2788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thanh, T.N.; Jurgen, B.; Bauch, M.; Liebeke, M.; Lalk, M.; Ehrenreich, A.; Evers, S.; Maurer, K.H.; Antelmann, H.; Ernst, F.; et al. Regulation of acetoin and 2,3-butanediol utilization in Bacillus licheniformis. Appl. Microbiol. Biotechnol. 2010, 87, 2227–2235. [Google Scholar] [CrossRef]
- Raspoet, D.; Pot, B.; De Deyn, D.; De Vos, P.; Kersters, K.; De Ley, J. Differentiation between 2,3-Butanediol Producing Bacillus licheniformis and B. polymyxa Strains by Fermentation Product Profiles and Whole-Cell Protein Electrophoretic Patterns. Syst. Appl. Microbiol. 1991, 14, 1–7. [Google Scholar] [CrossRef]
- Shariati, P.; Mitchell, W.J.; Boyd, A.; Priest, F.G. Anaerobic metabolism in Bacillus licheniformis NClB 6346. Microbiology 1995, 141, 1117–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Chen, C.; Li, K.; Wang, Y.; Gao, C.; Ma, C.; Xua, P. Efficient Simultaneous Saccharification and Fermentation of Inulin to 2,3-Butanediol by Thermophilic Bacillus licheniformis ATCC 14580. Appl. Environ. Microbiol. 2014, 80, 6458–6464. [Google Scholar] [CrossRef] [Green Version]
- Białkowska, A.M.; Gromek, E.; Krysiak, J.; Sikora, B.; Kalinowska, H.; Jędrzejczak-Krzepkowska, M.; Celina Kubik, C.; Lang, S.; Schütt, F.; Turkiewicz, M. Application of enzymatic apple pomace hydrolysate to production of 2,3-butanediol by alkaliphilic Bacillus licheniformis NCIMB 8059. J. Ind. Microbiol. Biotechnol. 2015, 42, 1609–1621. [Google Scholar] [CrossRef] [PubMed]
- Moes, J.; Griot, M.; Keller, J.; Heinzle, E.; Dunn, I.J.; Bourne, J.R. A microbial culture with oxygen-sensitive product distribution as a potential tool for characterizing bioreactor oxygen-transport. Biotechnol. Bioeng. 1985, 27, 482–489. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.W.; Rao, Z.M.; Zhang, X.; Xu, M.J.; Xu, Z.H.; Yang, S.T. Fermentation of biodiesel-derived glycerol by Bacillus amyloliquefaciens: Effects of co-substrates on 2,3-butanediol production. Appl. Microbiol. Biotechnol. 2013, 97, 7651–7658. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Xu, H.; Li, Q.-J.; Feng, X.-H.; Li, S. Optimization of medium for one-step fermentation of inulin extract from Jerusalem artichoke tubers using Paenibacillus polymyxa ZJ-9 to produce R,R-2,3-butanediol. Bioresour. Technol. 2010, 101, 7076–7082. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.-J.; Cheng, J.-S.; Liang, Y.-Q.; Jiang, T.; Yuan, Y.-J. Regulation of extracellular oxidoreduction potential enhanced (R,R)-2,3-butanediol production by Paenibacillus polymyxa CJX518. Bioresour. Technol. 2014, 167, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Adlakha, N.; Yazdani, S.S. Efficient production of (R,R)-2,3-butanediol from cellulosic hydrolysate using Paenibacillus polymyxa ICGEB2008. J. Ind. Microbiol. Biotechnol. 2015, 42, 21–28. [Google Scholar] [CrossRef]
- Ma, K.; He, M.; You, H.; Pan, L.; Wang, Z.; Wang, Y.; Hu, G.; Cui, Y.; Maeda, T. Improvement of (R,R)-2,3-butanediol production from corn stover hydrolysate by cell recycling continuous fermentation. Chem. Eng. J. 2018, 332, 361–369. [Google Scholar] [CrossRef]
- Dias, B.D.C.; Lima, M.E.D.N.V.; Vollú, R.E.; da Mota, F.F.; da Silva, A.J.R.; de Castro, A.M.; Freire, D.M.G.; Seldin, L. 2,3-Butanediol production by the non-pathogenic bacterium Paenibacillus brasilensis. Appl. Microbiol. Biotechnol. 2018, 102, 8773–8782. [Google Scholar] [CrossRef]
- Xiao, Z.J.; Liu, P.H.; Qin, J.Y.; Xu, P. Statistical optimization of medium components for enhanced acetoin production from molasses and soybean meal hydrolysate. Appl. Microbiol. Biotechnol. 2007, 74, 61–68. [Google Scholar] [CrossRef]
- Dai, J.Y.; Cheng, L.; He, Q.F.; Xiu, Z.L. High acetoin production by a newly isolated marine Bacillus subtilis strain with low requirement of oxygen supply. Process Biochem. 2015, 50, 1730–1734. [Google Scholar] [CrossRef]
- Xu, P.; Xiao, Z.J.; Du, Y.; Wei, Z. Acetoin High-Yield Bacillus pumilus Strain. U.S. Patent US 8,158,402 B2, 17 April 2012. [Google Scholar]
- Zhang, L.; Chen, S.; Xie, H.; Tian, Y.; Hu, K. Efficient acetoin production by optimization of medium components and oxygen supply control using a newly isolated Paenibacillus polymyxa CS107. J. Chem. Technol. Biotechnol. 2012, 87, 1551–1557. [Google Scholar] [CrossRef]
- Qi, G.; Kang, Y.; Li, L.; Xiao, A.; Zhang, S.; Wen, Z.; Xu, D.; Chen, S. Deletion of meso-2,3-butanediol dehydrogenase gene budC for enhanced D(−)2,3-butanediol production in Bacillus licheniformis. Biotechnol. Biofuels 2014, 7, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Y.; Zhang, J.; Li, L.; Wen, Z.; Nomura, C.T.; Wu, S.; Chen, S. Engineering Bacillus licheniformis for the production of meso-2,3-butanediol. Biotechnol. Biofuels 2016, 9, 117. [Google Scholar] [CrossRef] [Green Version]
- Białkowska, A.M.; Jędrzejczak-Krzepkowska, M.; Gromek, E.; Krysiak, J.; Sikora, B.; Kalinowska, H.; Kubik, C.; Schütt, F.; Turkiewicz, M. Effects of genetic modifications and fermentation conditions on 2,3-butanediol production by alkaliphilic Bacillus subtilis. Appl. Microbiol. Biotechnol. 2016, 100, 2663–2676. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Rao, Z.; Zhang, X.; Xu, M.; Xu, Z.; Yang, S.-T. Enhanced 2,3-butanediol production from biodiesel-derived glycerol by engineering of cofactor regeneration and manipulating carbon flux in Bacillus amyloliquefaciens. Microb. Cell Fact. 2015, 14, 122. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Rao, Z.; Zhang, X.; Xu, M.; Xu, Z.; Yang, S.T. Improved production of 2,3-butanediol in Bacillus amyloliquefaciens by over-expression of glyceraldehyde-3-phosphate dehydrogenase and 2,3-butanediol dehydrogenase. PLoS ONE 2013, 8, e76149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Cao, C.; Jiang, R.; Xu, H.; Xue, F.; Huang, W.; Ni, H.; Gao, J. Production of R,R-2,3-butanediol of ultra-high optical purity from Paenibacillus polymyxa ZJ-9 using homologous recombination. Bioresour. Technol. 2018, 261, 272–278. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, Y.; Gao, J.; Xu, H.; Cao, C.; Xue, F.; Ding, G.; Peng, Y. Introduction of the exogenous NADH coenzyme regeneration system and its influence on intracellular metabolic flux of Paenibacillus polymyxa. Bioresour. Technol. 2016, 201, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Bao, T.; Zhang, X.; Rao, Z.; Zhao, X.; Zhang, R.; Yang, T.; Xu, Z.; Yang, S. Efficient Whole-Cell Biocatalyst for Acetoin Production with NAD+ Regeneration System through Homologous Co-Expression of 2,3-Butanediol Dehydrogenase and NADH Oxidase in Engineered Bacillus subtilis. PLoS ONE 2019, 9, e102951. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, R.; Bao, T.; Rao, Z.; Yang, T.; Xu, M.; Xu, Z.; Li, H.; Yang, S. The rebalanced pathway significantly enhances acetoin production by disruption of acetoin reductase gene and moderate-expression of a new water-forming NADH oxidase in Bacillus subtilis. Metab. Eng. 2014, 23, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhao, X.; Zhang, J.; Zhao, C.; Liu, J.; Tian, Y.; Yang, L. Effect of deletion of 2, 3-butanediol dehydrogenase gene (bdhA) on acetoin production of Bacillus subtilis. Prep. Biochem. Biotechnol. 2017, 47, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wei, X.; Yu, W.; Wen, Z.; Chen, S. Enhancement of acetoin production from Bacillus licheniformis by 2,3-butanediol conversion strategy: Metabolic engineering and fermentation control. Process Biochem. 2017, 57, 35–42. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, R.; Bao, T.; Yang, T.; Xu, M.; Li, H.; Xu, Z.; Rao, Z. Moderate expression of the transcriptional regulator ALsR enhances acetoin production by Bacillus subtilis. J. Ind. Microbiol. Biotechnol. 2013, 40, 1067–1076. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Fu, J.; Zhang, X.; Chen, T. Metabolic engineering of Bacillus subtilis for enhanced production of acetoin. Biotechnol. Lett. 2012, 34, 1877–1885. [Google Scholar] [CrossRef]
- Xu, H.; Jia, S.; Liu, J. Production of acetoin by Bacillus subtilis TH-49. In Proceedings of the 2011 International Conference on Consumer Electronics, Communications and Networks (CECNet), Xianning, China, 11–13 March 2011; pp. 1524–1527. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, R.; Yang, T.; Zhang, J.; Xu, M.; Li, H.; Xu, Z.; Rao, Z. Mutation breeding of acetoin high producing Bacillus subtilis blocked in 2,3-butanediol dehydrogenase. World J. Microbiol. Biotechnol. 2013, 29, 1783–1789. [Google Scholar] [CrossRef]
- Jia, X.; Peng, X.; Liu, Y.; Han, Y. Conversion of cellulose and hemicellulose of biomass simultaneously to acetoin by thermophilic simultaneous saccharification and fermentation. Biotechnol. Biofuels 2017, 10, 232. [Google Scholar] [CrossRef] [Green Version]
- Luo, Q.; Wu, J.; Wu, M. Enhanced acetoin production by Bacillus amyloliquefaciens through improved acetoin tolerance. Process Biochem. 2014, 49, 1223–1230. [Google Scholar] [CrossRef]
- Feng, J.; Gu, Y.; Yan, P.-F.; Song, C.; Wang, Y. Recruiting Energy-Conserving Sucrose Utilization Pathways for Enhanced 2,3-Butanediol Production in Bacillus subtilis. ACS Sustain. Chem. Eng. 2017, 5, 11221–11225. [Google Scholar] [CrossRef]
- Deshmukh, A.N.; Nipanikar-Gokhale, P.; Jain, R. Engineering of Bacillus subtilis for the Production of 2,3-Butanediol from Sugarcane Molasses. Appl. Biochem. Biotechnol. 2016, 179, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Sikora, B.; Kubik, C.; Kalinowska, H.; Gromek, E.; Białkowska, A.; Jędrzejczak-Krzepkowska, M.; Schüett, F.; Turkiewicz, M. Application of byproducts from food processing for production of 2,3-butanediol using Bacillus amyloliquefaciens TUL 308. Prep. Biochem. Biotechnol. 2016, 46, 610–619. [Google Scholar] [CrossRef]
- Białkowska, A. Strategies for efficient and economical 2,3-butanediol production: New trends in this field. World J. Microbiol. Biotechnol. 2016, 32, 200. [Google Scholar] [CrossRef]
- Koutinas, A.A.; Yepez, B.; Kopsahelis, N.; Freire, D.M.G.; de Castro, A.M.; Papanikolaou, S.; Kookos, I.K. Techno-economic evaluation of a complete bioprocess for 2,3-butanediol production from renewable resources. Bioresour. Technol. 2016, 204, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Huang, H.; Chen, S.; Qi, G. Production of optically pure 2,3-butanediol from Miscanthus floridulus hydrolysate using engineered Bacillus licheniformis strains. World J. Microbiol. Biotechnol. 2018, 34, 66. [Google Scholar] [CrossRef]
- Khelil, O.; Choubane, S.; Cheba, B.A. Polyphenols content of spent coffee grounds subjected to physico-chemical pretreatments influences lignocellulolytic enzymes production by Bacillus sp. R2. Bioresour. Technol. 2016, 211, 769e773. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Tanabe, S.H.; Xie, C.; Honda, D.; Sun, J.; Ai, L. Bacillus ligniniphilus sp. nov.; an alkaliphilic and halotolerant bacterium isolated from sediments of the South China Sea. Int. J. Syst. Evol. Microbiol. 2014, 64, 1712e1717. [Google Scholar] [CrossRef]
- Borriss, R.; Chen, X.H.; Rueckert, C.; Blom, J.; Becker, A.; Baumgarth, B.; Fan, B.; Pukall, R.; Schumann, P.; Spröer, C.; et al. Relationship of Bacillus amyloliquefaciens clades associated with strains DSM 7 T and FZB42 T: A proposal for Bacillus amyloliquefaciens subsp. nov. and Bacillus amyloliquefaciens subsp. plantarum subsp. nov. based on complete genome sequence comparisons. Int. J. Syst. Evol. Microbiol. 2011, 61, 1786e1801. [Google Scholar] [CrossRef]
- Chen, L.; Gu, W.; Xu, H.-Y.; Yang, G.-L.; Shan, X.-F.; Chen, G.; Kang, Y.-H.; Wang, C.-F.; Qian, A.-D. Comparative genome analysis of Bacillus velezensis reveals a potential for degrading lignocellulosic biomass. 3 Biotech 2018, 8, 253. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Yang, W.; Meng, F.; Ji, S.; Xin, H.; Cao, B. Characterization of an acidic cellulase produced by Bacillus subtilis BY-4 isolated from gastrointestinal tract of Tibetan pig. J. Taiwan Inst. Chem. Eng. 2015, 56, 67–72. [Google Scholar] [CrossRef]
- Wei, K.S.C.; Teoh, T.C.; Koshy, P.; Salmah, I.; Zainudin, A. Cloning, expression and characterization of the endoglucanase gene from Bacillus subtilis UMC7 isolated from the gut of the indigenous termite Macrotermes malaccensis in Escherichia coli. Electron. J. Biotechnol. 2015, 18, 103–109. [Google Scholar] [CrossRef] [Green Version]
- Fatani, S.; Saito, Y.; Alarawi, M.; Gojobori, T.; Mineta, K. Genome sequencing and identification of cellulase genes in Bacillus paralicheniformis strains from the Red Sea. BMC Microbiol. 2021, 21, 254. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.M.; Xin, X.J.; Li, C.X.; Xu, J.H.; Bao, J. Cloning of thermostable cellulase genes of Clostridium thermocellum and their secretive expression in Bacillus subtilis. Appl. Biochem. Biotechnol. 2012, 166, 652–662. [Google Scholar] [CrossRef] [PubMed]
- Arsov, A.; Petrov, K.; Petrova, P. Enhanced Activity by Genetic Complementarity: Heterologous Secretion of Clostridial Cellulases by Bacillus licheniformis and Bacillus velezensis. Molecules 2021, 26, 5625. [Google Scholar] [CrossRef]
- Nakashimada, Y.; Marwoto, B.; Kashiwamura, T.; Kakizono, T.; Nishio, N. Enhanced 2,3-butanediol production by addition of acetic acid in Paenibacillus polymyxa. J. Biosci. Bioeng. 2000, 90, 661–664. [Google Scholar] [CrossRef]
- Deshmukh, A.; Mistry, S.; Yewale, T.; Mahajan, D.; Jain, R. Production of 2,3-butanediol from Sugarcane Molasses Using Bacillus subtilis. Int. J. Adv. Biotechnol. Res. 2015, 6, 66–79. Available online: https://www.researchgate.net/publication/273453911_Production_of_2_3-Butanediol_from_Sugarcane_Molasses_Using_Bacillus_subtilis (accessed on 9 December 2021).
- Wang, Q.; Chen, T.; Zhao, X.; Chamu, J. Metabolic engineering of thermophilic Bacillus licheniformis for chiral pure D(−)2,3-butanediol production. Biotechnol. Bioeng. 2012, 109, 1610–1621. [Google Scholar] [CrossRef]
- Jansen, N.B.; Flickinger, M.C.; Tsao, G.T. Production of 2,3-butanediol from D(−)xylose by Klebsiella oxytoca ATCC 8724. Biotechnol. Bioeng. 1984, 26, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Hakizimana, O.; Emmanuel Matabaro, E.; Lee, B.H. The current strategies and parameters for the enhanced microbial production of 2,3-butanediol. Biotechnol. Rep. 2020, 25, e00397. [Google Scholar] [CrossRef]
- Harvianto, G.R.; Haider, J.; Hong, J.; Long, N.V.D.; Shim, J.-J.; Cho, M.H.; Kim, W.K.; Lee, M. Purification of 2,3-butanediol from fermentation broth: Process development and techno-economic analysis. Biotechnol. Biofuels 2018, 11, 18. [Google Scholar] [CrossRef]
- Fu, J.; Huo, G.; Feng, L.; Mao, Y.; Wang, Z.; Ma, H.; Chen, T.; Zhao, X. Metabolic engineering of Bacillus subtilis for chiral pure meso-2,3-butanediol production. Biotechnol. Biofuels 2016, 9, 90. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain | Substrate | 2,3-BD (g/L) | Yield (g/g) | Productivity (g/L/h) | Process Mode | Reference |
|---|---|---|---|---|---|---|---|
| B. licheniformis | |||||||
| NCIMB 8059 | Corn starch hydrolysate | 6.44 | - | 0.126 | Batch | [68] | |
| Isolate | Glucose | 8.70 | - | - | Batch | [66] | |
| 10-1-A | Glucose | 115.7 | 0.47 | 2.4 | Fed-batch | [70] | |
| ATCC 14580 | Inulin | 103.0 | - | 3.4 | Fed-batch | [88] | |
| DSM 8785 | Glucose | 76.2 | 0.42 | 0.86 | Batch | [71] | |
| DSM 8785 | Glucose | 144.7 | 0.4 | 1.14 | Fed-batch | [71] | |
| NCIMB 8059 | Apple pomace hydrolysate/glucose | 87.71 | 0.36 | 0.55 | Fed-batch | [89] | |
| 24 | Glucose | 91.23 | 0.49 | 1.94 | Batch | [67] | |
| 24 | Glucose | 138.8 | 0.48 | 1.16 | Fed-batch | [67] | |
| X10 | Corn stover hydrolysate | 74 | 0.47 | 2.1 | Fed-batch | [73] | |
| GSC3102 | Glucose | 95.9 | 0.431 | 1.50 | Fed-batch | [75] | |
| B. subtilis | |||||||
| AJ1992 | Glucose | 2.5 | 0.33 | 0.38 | Batch | [90] | |
| 35 | Glycerol | 11.10 | 0.27 | 0.12 | Batch in flasks | [69] | |
| CS13 | Sucrose | 132.4 | 0.45 | 2.45 | Fed-batch | [58] | |
| B. amyloliquefaciens | |||||||
| B10-127 | Glucose | 92.3 | - | 0.96 | Fed-batch | [63] | |
| 18025 | Molasses | 48.7 | 0.4 | 0.83 | Fed-batch | [78] | |
| Wild type | Bakery waste hydrolysate | 55.2 | 0.42 | 1.19 | Batch | [5] | |
| B10-127 | Glycerol+molasses | 83.3 | 0.42 | 0.87 | Fed-batch | [91] | |
| Other Bacillus spp. | |||||||
| B. velezensis | 5RB | Glucose | 18.1 | - | - | Batch in flasks | [69] |
| B. safensis | 14A | Glucose | 13.9 | - | - | Batch in flasks | [69] |
| B. toyonensis | 11RA | Glucose | 12.0 | - | - | Batch in flasks | [69] |
| B. atrophaeus | NRS-213 | Glucose | 29.9 | 0.33 | 1.0 | Batch | [76] |
| B. mojavensis | B-14698 | Glucose | 37.8 | 0.26 | 1.1 | Batch | [76] |
| B. vallismortis | B-14891 | Glucose | 59.1 | 0.37 | 1.18 | Batch | [76] |
| P. polymyxa | |||||||
| ZJ-9 | Inulin | 36.92 | 0.51 | 0.89 | Batch | [92] | |
| CJX518 | Glucose | 71.71 | - | 1.33 | Fed-batch | [93] | |
| DSM 365 | Glucose | 68.5 | 0.34 | 0.70 | Fed-batch | [61] | |
| DSM 365 | Glucose | 111.0 | - | - | Batch | [62] | |
| ICGEB2008 | Cellulose hydrolysates | 16.50 | 0.33 | 2.01 | Batch | [94] | |
| ATCC 12321 | Glucose/xylose | 18.8 | 0.31 | 1.13 | CRCF * | [95] | |
| P. brasiliensis | |||||||
| PB24 | Glucose | 27.0 | 0.43 | 0.68 | Batch in flasks | [96] | |
| Species | Strain | Substrate | Acetoin (g/L) | Yield (g/g) | Productivity (g/L/h) | Process Mode | Reference |
|---|---|---|---|---|---|---|---|
| B. licheniformis | |||||||
| MEL09 | Glucose | 41.3 | 0.42 | - | Batch | [35] | |
| B. subtilis | |||||||
| JNA 3-10 | Glucose | 42.2 | 0.32 | - | Batch in flasks | [7] | |
| CICC10025 | Molasses | 35.4 | 0.41 | 0.63 | Batch | [97] | |
| SF4-3 | Glucose | 48.9 | 0.39 | 0.56 | Batch | [2] | |
| DL01 | Molasses | 61.2 | 0.34 | 0.807 | Batch | [98] | |
| B. pumilus | |||||||
| DSM 16187 | Glucose | 63.0 | - | - | Batch in flasks | [99] | |
| B. amyloliquefaciens | |||||||
| FMME044 | Glucose | 51.2 | 0.43 | 1.42 | Batch | [64] | |
| Wild type | Bakery waste hydrolysate | 65.9 | 0.31 | 1.57 | Fed-batch | [5] | |
| P. polymyxa | |||||||
| CS107 | Glucose | 55.3 | - | 1.32 | Fed-batch | [100] | |
| Species | Strain | Substrate | 2,3-BD (g/L) | Yield (g/g) | Productivity (g/L/h) | Modification | Reference |
|---|---|---|---|---|---|---|---|
| B. licheniformis | |||||||
| MW3 | Glucose | 123.7 (D(−)) | 0.508 | 2.95 | ΔbudC | [45] | |
| MW3 | Glucose | 90.1 (meso) | 0.424 | 2.82 | Δgdh | [45] | |
| WX-02 | Glucose | 30.7 (D(−)) | - | 1.28 | ΔbudC | [101] | |
| WX-02 | Glucose | 98.0 (meso) | - | 0.94 | ΔgdhΔacoR | [102] | |
| B. subtilis | |||||||
| LOCK 1086 | Sugarbeet molasses, glucose | 75.73 | - | 0.66 | bdhA overexpression, vhb cloning | [103] | |
| B. amyloliquefaciens | |||||||
| B10-127 | Glucose | [64] | |||||
| B10-127 | Crude glycerol+molasses | 102.3 | 0.44 | 1.16 | NADH/NAD+ regeneration system introduction, alsR under pbdhA promoter control | [104] | |
| B 10-127 | Glucose | 132.9 | - | 2.95 | GAPDH, BDH overexpression | [105] | |
| P. polymyxa | |||||||
| ZJ-9 | Glucose | 25.88 | - | - | ΔdudA | [106] | |
| XG-1 | Glucose | 51.3 | - | - | NAD+-dependent fdh gene cloning | [107] | |
| Species | Strain | Substrate | Acetoin (g/L) | Yield (g/g) | Productivity (g/L/h) | Modification | Reference |
|---|---|---|---|---|---|---|---|
| B. licheniformis | |||||||
| WX-02 | Glucose | 78.79 | 0.31 | 0.58 | ΔbudCΔacoR | [111] | |
| B. subtilis | |||||||
| ZB02 | Xylose/glucose/arabinose | 62.2 | 0.29 | 0.86 | ARSRCPΔacoAΔbdhA, cloned araE, xylA, xylB | [8] | |
| ZB02 | Lignocellulose hydrolysate | 11.2 | - | - | ARSRCPΔacoAΔbdhA, cloned araE, xylA, xylB | [8] | |
| 168 | 2,3-BD | 91.8 | 0.765 | 2.30 | NADH oxidase and BDH coexpression | [108] | |
| BMN | Glucose | 56.7 | 0.378 | 0.639 | ΔbdhA, yodC overexpression | [109] | |
| 168 | Glucose | 24.6 | - | - | ΔbdhA | [110] | |
| PAR | Glucose | 41.5 | 0.35 | 0.43 | alsR promoter control | [112] | |
| BSUW06 | Glucose | 20 | - | 0.273 | ΔbdhA, ΔacoA, Δpta, overexpression of alsSD | [113] | |
| TH-49 | Glucose | 56.9 | - | - | Mutagenesis | [114] | |
| JNA-UD(−)6 | Glucose | 53.9 | - | - | Mutagenesis | [115] | |
| IPE5-4-UD(−)4 | Lignocellulose hydrolysate | 22.76 | 0.46 | 0.38 | Mutagenesis | [116] | |
| B. amyloliquefaciens | |||||||
| E-11 | Glucose | 71.5 | 0.41 | 1.63 | Adaptive evolution | [117] | |
| Species | Strain | Temperature (°C) | pH | Agitation (rpm) | Aeration (vvm) | Reference |
|---|---|---|---|---|---|---|
| B. licheniformis | ||||||
| 10-1-A | 50 * | 7.0 * | 400/200 * | 1.0 | [70] | |
| NCIMB 8059 | 37 | 6.5 † | 250 | 1.2 | [89] | |
| 24 | 37.8 * | 6.23 * | 500 | 3.68 * | [67] | |
| X10 | 50 | 7.0 | 400/200 | 1.0 | [73] | |
| GSC3102 | 50 | 7.0 | 500/400 | 1.0 | [75] | |
| DSM 13T | 37 | 6.0 * | Anaerobic | [86] | ||
| DSM 394 | 37 | 6.0–6.5 * | Anaerobic | [86] | ||
| NCIB 8069 | 37 | 6.0 * | Anaerobic | [86] | ||
| NCIB 8062 | 37 | 6.0 * | Anaerobic | [86] | ||
| NCIB 8549 | 37 | 6.0 * | Anaerobic | [86] | ||
| DSM 392 | 37 | 6.0–6.5 * | Anaerobic | [86] | ||
| DSM 603 | 37 | 6.0 * | Anaerobic | [86] | ||
| B. subtilis | ||||||
| CS13 | 37 | 6.5 | 500 * | 2.0 * | [58] | |
| B. amyloliquefaciens | ||||||
| B10-127 | 37 * | 6.5 † * | 100 f * | [63] | ||
| B10-127 | 37 | 6.5 ‡ | 350/400/350 * | 0.66 | [104] | |
| pBG | 37 | 6.5 † | 350 | 0.33 | [105] | |
| 18025 | 40 | 6.0 | 300 * | 1.0 | [5] | |
| 18025 | 40 * | 6.0 * | kL a = 49 h−1 * | 1.0 | [78] | |
| Other Bacillus spp. | ||||||
| B. velezensis | 5RB | 37 | 6.5 † | 200 f | - | [69] |
| B. safensis | 14A | 37 | 6.5 † | 200 f | - | [69] |
| B. toyonensis | 11RA | 37 | 6.5 † | 200 f | - | [69] |
| B. atropaeus | NRS-213 | 39 | 6.7 † | 700 mp | - | [76] |
| B. mojavensis | B-14698 | 39 | 6.7 † | 700 mp | - | [76] |
| B. vallismortis | B-14891 | 39 | 6.7 † | 700 mp | - | [76] |
| P. polymixa | ||||||
| DSM 365 | 35 * | 6.5 † | 300 | 0.075 | [61] | |
| DSM 365 | 37 | 6.0 | 500 * | 0.2 * | [62] | |
| ATCC 12321 | 37 | 6.5 | 500 | 0.2 | [95] | |
| CJX518 | 37 * | 5.2 | 400 * | 0.1 * | [93] | |
| P. brasiliensis | ||||||
| PB24 | 32 | 200 f | [96] |
| Species | Strain | Temperature (°C) | pH | Agitation (rpm) | Aeration (vvm) | Reference |
|---|---|---|---|---|---|---|
| B. licheniformis | ||||||
| 10-1-A | 50 * | 7.0 * | 400 * | 1.0 | [70] | |
| B. subtilis | ||||||
| CS13 | 37 | 6.5 | 500 * | 2.0 * | [58] | |
| CICC10025 | 37 | 7.0 † | 700 | 1.0 | [97] | |
| SF4-3 | 37 | 7.0 † | 300 * | 0.5 | [2] | |
| DL1 | 37 | 6.5 † | 200/300/400 * | 0.4 * | [98] | |
| JNA-310 | 37 * | 4.0 † * | 160 f | [7] | ||
| ZB02 | 37 | 7.0 | 400 * | 1.0 | [8] | |
| B. amyloliquefaciens | ||||||
| B10-127 | 25 * | 4.5 † * | 200 f * | [63] | ||
| 18025 | 40 | 6.0 | 600 * | 1.0 | [5] | |
| 18025 | 40 | 6.0 * | kL a = 104 h−1 * | 1.0 | [78] | |
| FMME044 | 37 | - | 350/500 * | 1.0 | [64] | |
| B. pumilus | ||||||
| DSM 16187 | 37 | - | 180 f | - | [99] | |
| P. polymixa | ||||||
| DSM 365 | 37 | 6.0 | 800 * | 0.2 * | [62] | |
| CS107 | 37 | 6.0 | 500 * | 0.5 | [100] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrov, K.; Petrova, P. Current Advances in Microbial Production of Acetoin and 2,3-Butanediol by Bacillus spp. Fermentation 2021, 7, 307. https://doi.org/10.3390/fermentation7040307
Petrov K, Petrova P. Current Advances in Microbial Production of Acetoin and 2,3-Butanediol by Bacillus spp. Fermentation. 2021; 7(4):307. https://doi.org/10.3390/fermentation7040307
Chicago/Turabian StylePetrov, Kaloyan, and Penka Petrova. 2021. "Current Advances in Microbial Production of Acetoin and 2,3-Butanediol by Bacillus spp." Fermentation 7, no. 4: 307. https://doi.org/10.3390/fermentation7040307