Exploring the Biodiversity of Red Yeasts for In Vitro and In Vivo Phenotypes Relevant to Agri-Food-Related Processes

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Culture Media
2.2. Adhesion to Plastic Materials
2.3. Cell MAT Formation
2.4. Invasive Growth
2.5. Growth at Different Temperatures
2.6. Biocontrol In Vivo
2.7. Statistical Analysis
3. Results and Discussion
3.1. Biofilm Forming Ability
3.2. Invasive Growth and Temperature Growth Interval
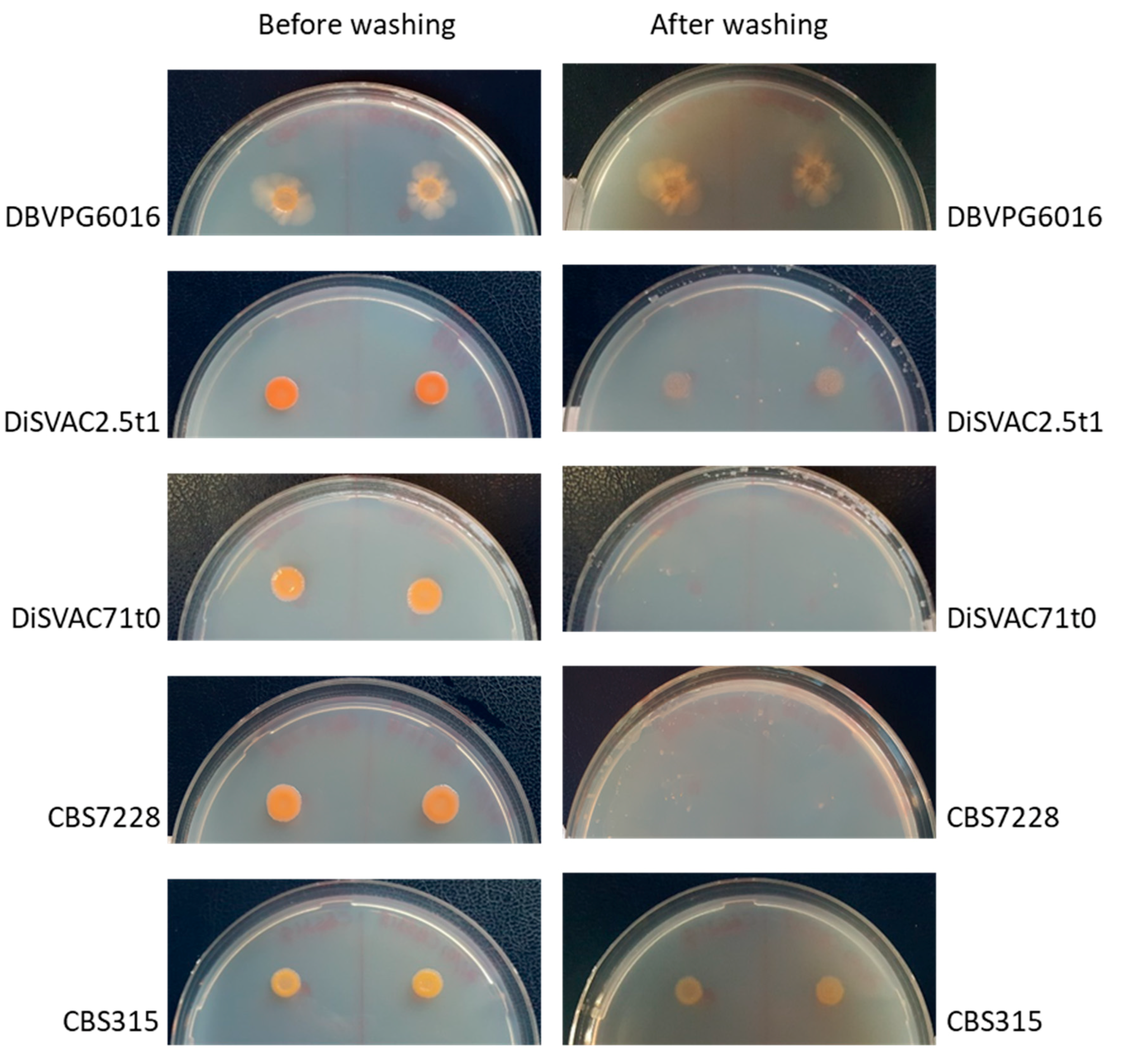
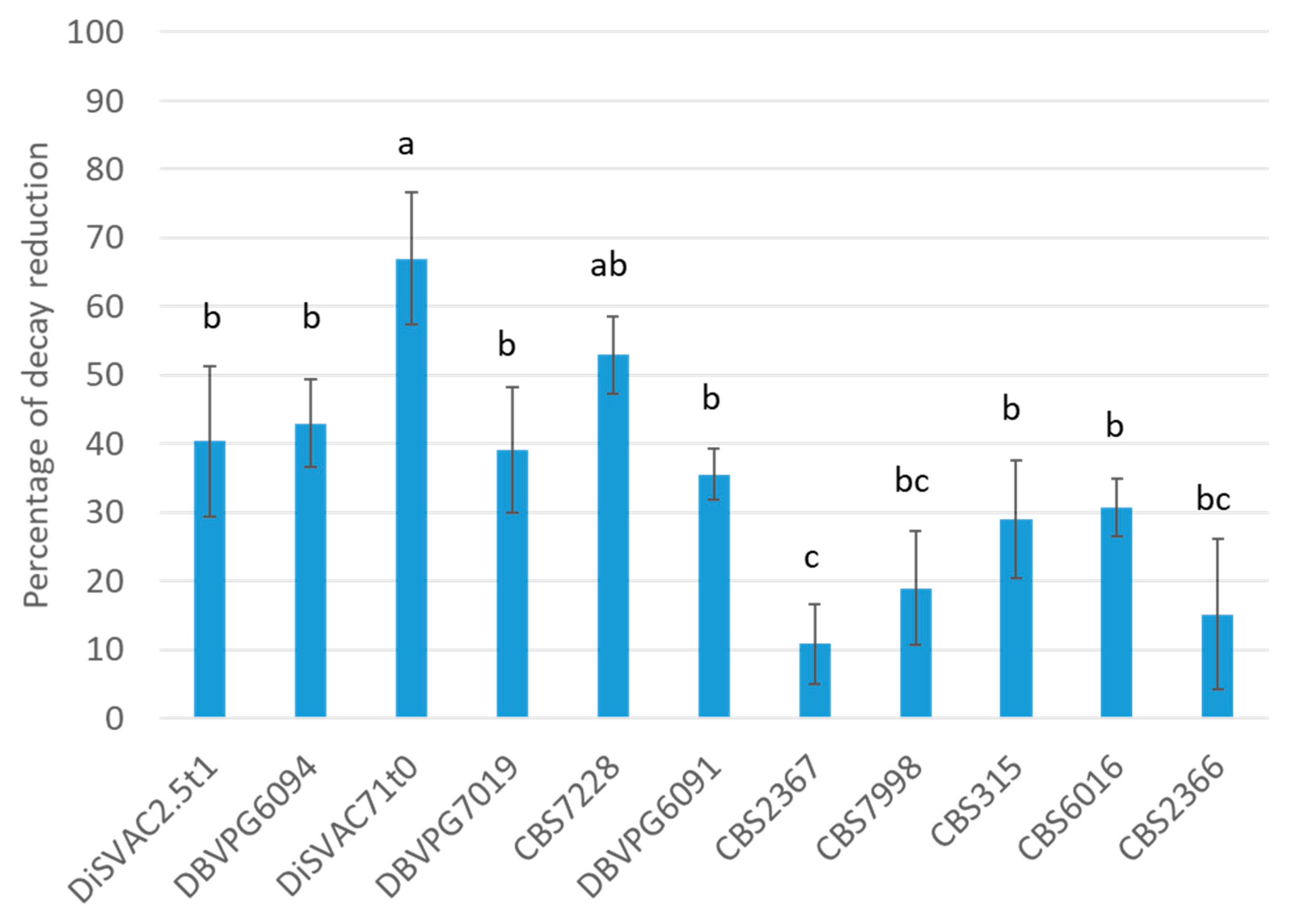
3.3. Biocontrol Activity on Apple Fruit
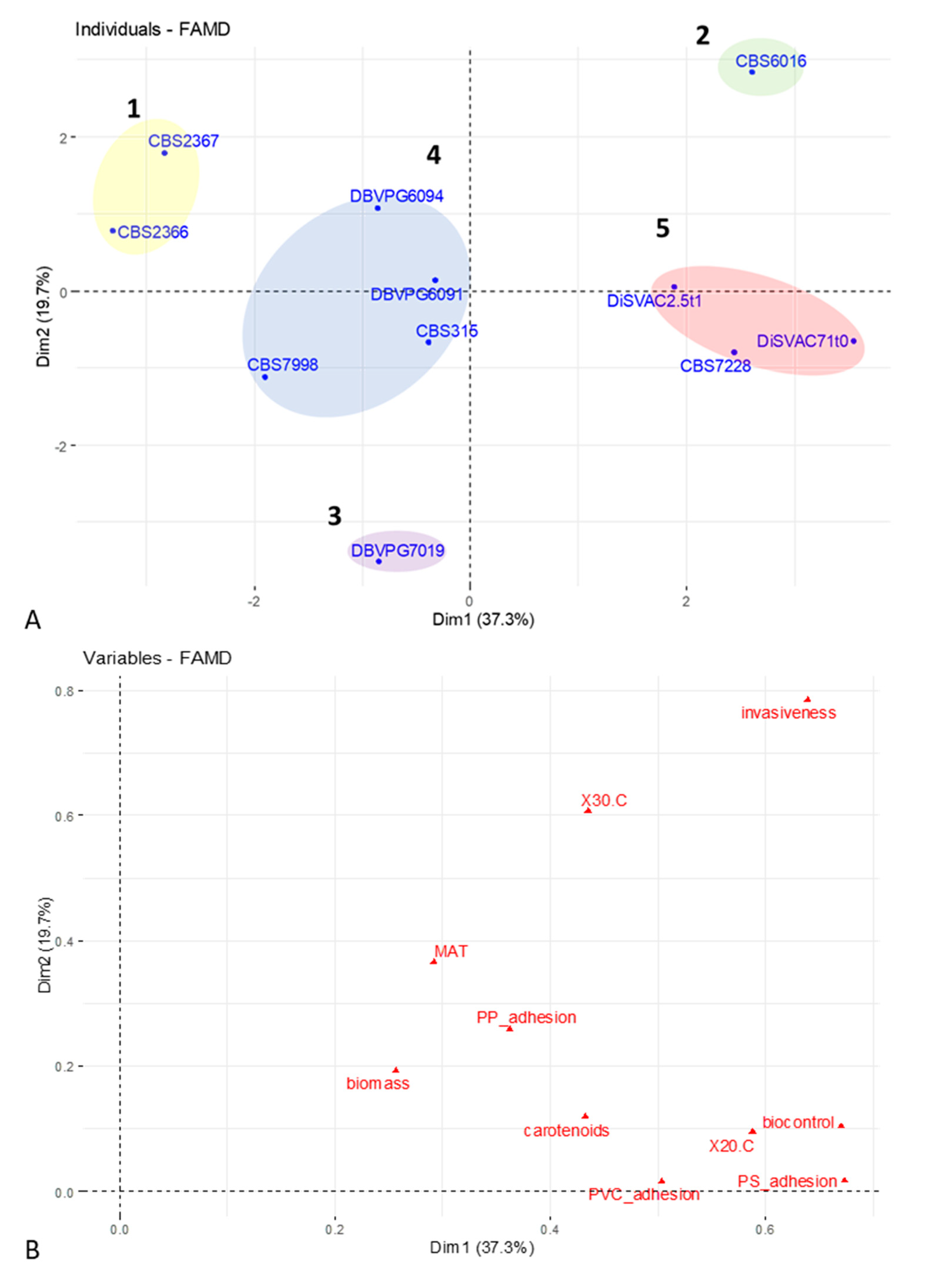
3.4. FAMD and Cluster Analysis of Phenotypic Data
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Molla, G.; Motteran, L.; Piubelli, L.; Pilone, M.S.; Pollegioni, L. Regulation of D-amino acid oxidase expression in the yeast Rhodotorula gracilis. Yeast 2003, 20, 1061–1069. [Google Scholar] [CrossRef] [Green Version]
- Csutak, O.; Stoica, I.; Vassu, T. Evaluation of production, stability and activity of biosurfactants from yeasts with application of bioremediation of oil polluted environment. Rev. Chim. 2012, 67, 973–977. [Google Scholar]
- Johnson, E.A. Biotechnology of non-Saccharomyces yeasts—The basidiomycetes. Appl. Microbiol. Biotechnol. 2013, 97, 7563–7577. [Google Scholar] [CrossRef] [PubMed]
- Moliné, M.; Libkind, D.; del Carmen Diéguez, M.; van Broock, M. Photoprotective role of carotenoids in yeasts: Response to UV-B of pigmented and naturally-occurring albino strains. J. Photochem. Photobiol. B Biol. 2009, 95, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Fiedor, J.; Burda, K. Potential role of carotenoids as antioxidants in human health and disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishino, H.; Murakoshi, M.; Tokuda, H.; Satomi, Y. Cancer prevention by carotenoids. Arch. Biochem. Biophys. 2009, 483, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Zhao, X.; Zhao, J.; Wu, S.; Zhao, Z.K. Effect of biomass hydrolysis by-products on oleaginous yeast Rhodosporidium toruloides. Biores. Technol. 2009, 100, 4843–4847. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhang, H.; Liu, W.; Zheng, X. Biocontrol of postharvest gray and blue mold decay of apples with Rhodotorula mucilaginosa and possible mechanisms of action. Int. J. Food Microbiol. 2011, 146, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Ge, L.; Chen, K.; Zhao, L.; Zhang, X. Enhanced biocontrol activity of Rhodotorula mucilaginosa cultured in media containing chitosan against postharvest diseases in strawberries: Possible mechanisms underlying the effect. J. Agric. Food Chem. 2014, 62, 4214–4224. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, L.; Ma, L.; Dong, Y.; Jiang, S.; Xu, B.; Zheng, X. Biocontrol of major post-harvest pathogens on apple using Rhodotorula glutinis and its effects on post-harvest quality parameters. Biol. Control 2009, 48, 79–83. [Google Scholar] [CrossRef]
- Ge, L.; Zhang, H.; Chen, K.; Ma, L.; Xu, Z. Effect of chitin on the antagonistic activity of Rhodotorula glutinis against Botrytis cinerea in strawberries and the possible mechanisms involved. Food Chem. 2010, 120, 490–495. [Google Scholar] [CrossRef]
- Li, B.; Peng, H.; Tian, S. Attachment capability of antagonistic yeast Rhodotorula glutinis to Botrytis cinerea contributes to biocontrol efficacy. Front. Microbiol. 2016, 7, 601. [Google Scholar] [CrossRef]
- Chen, J.; Li, B.; Qin, G.; Tian, S. Mechanism of H2O2-induced oxidative stress regulating viability and biocontrol ability of Rhodotorula glutinis. Int. J. Food Microbiol. 2015, 193, 152–158. [Google Scholar] [CrossRef]
- Yan, F.; Xu, S.; Chen, Y.; Zheng, X. Effect of rhamnolipids on Rhodotorula glutinis biocontrol of Alternaria alternata infection in cherry tomato fruit. Postharvest Biol.Technol. 2014, 97, 32–33. [Google Scholar] [CrossRef]
- Zhang, H.; Ma, L.; Turner, M.; Xu, H.; Zheng, X.; Dong, Y.; Jiang, S. Salicylic acid enhances biocontrol efficacy of Rhodotorula glutinis agains tpost-harvest Rhizopus rot of strawberries and the possible mechanisms involved. Food Chem. 2010, 122, 577–583. [Google Scholar] [CrossRef]
- Filonow, A.B. Role of competition for sugars by yeasts in the biocontrol of gray mold of apple. Biocontrol Sci. Technol. 1998, 8, 243–256. [Google Scholar] [CrossRef]
- Zheng, X.; Yang, Q.; Zhang, X.; Apaliya, M.T.; Ianiri, G.; Zhang, H.; Castoria, R. Biocontrol agents increase the specific rate of patulin production by Penicillium expansum but decrease the disease and total patulin contamination of apples. Front. Microbiol. 2017, 8, 1240. [Google Scholar] [CrossRef] [PubMed]
- Droby, S.; Wisniewski, M.; Macarisin, D.; Wilson, C. Twenty years of postharvest biocontrol research: Is it time for a new paradigm? Postharvest Biol. Technol. 2009, 52, 137–145. [Google Scholar] [CrossRef]
- Freimoser, F.M.; Rueda-Mejla, M.P.; Tilocca, B.; Migheli, Q. Biocontrol yeasts: Mechanisms and application. World J. Microbiol. Biotechnol. 2019, 35, 154. [Google Scholar] [CrossRef] [Green Version]
- Buck, J.W.; Andrews, J.H. Localized, positive charge mediates adhesion of Rhodosporidiun toruloides to barley leaves and polystyrene. Appl. Environ. Microbiol. 1999, 65, 2179–2183. [Google Scholar] [CrossRef] [Green Version]
- Pandin, C.; Le Coq, D.; Canette, A.; Symerich, S.; Briandet, R. Should the biofilm mode of life be taken into consideration for microbial biocontrol agents? Microbial Biotechnol. 2017, 10, 719–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raut, J.S.; Doke, S.K.; Karuppayil, S.M. Yeast biofilms in the context of human health and disease. In Yeast Diversity in Human Welfare; Satyanarayana, T., Kunze, G., Eds.; Springer: Singapore, 2017; pp. 57–82. [Google Scholar]
- Parvu, M.; Paraschivescu, M.T. Feeding Rhodotorula rubra yeast in egg yolk pigmentation. Rom. Biotechnol. Lett. 2014, 19, 4–14. [Google Scholar]
- Elwan, H.A.M.; Elnesr, S.S.; Abdallah, Y.; Hamdy, A.; El-Bogdady, A.H. Red yeast (Phaffia rhodozyma) as a source of astaxanthin and its impacts on productive performance and physiological responses of poultry. World’s Poultry Sci. J. 2019, 75, 273–284. [Google Scholar] [CrossRef]
- Camiolo, S.; Toome-Heller, M.; Aime, M.C.; Haridas, S.; Grigoriev, I.V.; Porceddu, A.; Mannazzu, I. An analysis of codon bias in six red yeasts species. Yeast 2018, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cutzu, R.; Coi, A.; Rosso, F.; Bardi, L.; Ciani, M.; Budroni, M.; Zara, G.; Zara, S.; Mannazzu, I. From crude glycerol to carotenoids by using a Rhodotorula glutinis mutant. World J. Microbiol. Biotechnol. 2013, 29, 1009–1017. [Google Scholar] [CrossRef]
- Deligios, M.; Fraumene, C.; Abbondio, M.; Mannazzu, I.; Tanca, A.; Addis, M.F.; Uzzau, S. Draft genome sequence of Rhodotorula mucilaginosa, an emergent opportunistic pathogen. Genome Announc. 2015, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landolfo, S.; Ianiri, G.; Camiolo, S.; Porceddu, A.; Mulas, G.; Chessa, R.; Zara, G.; Mannazzu, I. CAR gene cluster and transcript levels of carotenogenic genes in Rhodotorula mucilaginosa. Microbiology 2018, 164, 78–87. [Google Scholar] [CrossRef]
- Addis, M.F.; Tanca, A.; Abbondio, M.; Cutzu, R.; Biosa, G.; Pagnozzi, D.; Uzzau, S.; Mannazzu, I. Proteomic analysis of Rhodotorula mucilaginosa: Dealing with the issues of a non-conventional yeast. Yeast 2016, 33, 433–449. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, T.B.; Fink, G.R. Bakers’ yeast, a model for fungal biofilm formation. Science 2001, 291, 878–881. [Google Scholar] [CrossRef]
- Zara, G.; Mannazzu, I.; Sanna, M.L.; Orro, D.; Farris, G.A.; Budroni, M. Exploitation of the semi-homothallic life cycle of Saccharomyces cerevisiae for the development of breeding strategies. FEMS Yeast Res. 2008, 8, 1147–1154. [Google Scholar] [CrossRef] [Green Version]
- Bou Zeidan, M.; Zara, G.; Viti, C.; Decorosi, F.; Mannazzu, I.; Budroni, M.; Giovannetti, L.; Zara, S. L-histidine inhibits biofilm formation and FLO11-associated phenotypes in Saccharomyces cerevisiae flor yeasts. PLoS ONE 2014, 9, e112141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Soft. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Kassambara, A.; Mundt, F. factoextra: Extract and Visualize the Results of Multivariate Data Analyses. 2020. R Package Version 1.0.7. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 21 December 2020).
- Kumamoto, C.A. A contact-activated kinase signals Candida albicans invasive growth and biofilm development. Proc. Natl. Acad. Sci. USA 2005, 102, 5576–5581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, C.H.; Koh, K.S.; Xie, C.; Zhou, Y.; Williams, R.; Ng, W.J.; Rice, S.A.; Kjelleberg, S. The role of quorum sensing signalling in EPS production and the assembly of a sludge community into aerobic granules. ISME J. 2014, 8, 1186–1197. [Google Scholar] [CrossRef] [Green Version]
- Gow, N.; van de Veerdonk, F.; Brown, A.; Netea, M.G. Candida albicans morphogenesis and host defence: Discriminating invasion from colonization. Nat. Rev. Microbiol. 2012, 10, 112–122. [Google Scholar] [CrossRef] [Green Version]
- Abdallah, M.; Benoliel, C.; Drider, D.; Dhulster, P.; Chihib, N.-E. Biofilm formation and persistence on abiotic surfaces in the context of food and medical environments. Arch. Microbiol. 2014, 196, 453–472. [Google Scholar] [CrossRef]
- Gomes, I.B.; Simões, M.; Simões, L. An overview on the reactors to study drinking water biofilms. Water Res. 2014, 62, 63–87. [Google Scholar] [CrossRef] [Green Version]
- Zara, G.; Budroni, M.; Mannazzu, I.; Fancello, F.; Zara, S. Yeast biofilm in food realms: Occurrence and control. Worl. J. Microbiol. Biotechnol. 2020, 36, 134–145. [Google Scholar] [CrossRef]
- Licitra, G.; Ogier, J.C.; Parayre, S.; Pediliggieri, C.; Carnemolla, T.M.; Falentin, H. Variability of bacterial biofilms of the “tina” wood vats used in the ragusano cheese-making process. Appl. Environ. Microbiol. 2007, 73, 6980–6987. [Google Scholar] [CrossRef] [Green Version]
- Feofilova, E.P.; Galanina, L.A.; Sergeeva, Y.E.; Mysyakina, I.S. Strategies of food substrate colonization by mycelial fungi. Microbiology 2013, 82, 11–14. [Google Scholar] [CrossRef]
- Giaouris, E.; Heir, E.; Hébraud, M.; Chorianopoulos, N.; Langsrud, S.; Møretrø, T.; Habimana, O.; Desvaux, M.; Renier, S.; Nychas, G.-J. Attachment and biofilm formation by foodborne bacteria in meat processing environments: Causes, implications, role of bacterial interactions and control by alternative novel methods. Meat Sci. 2014, 97, 298–309. [Google Scholar] [CrossRef] [PubMed]
- Legras, J.-L.; Moreno-Garcia, J.; Zara, S.; Zara, G.; Garcia-Martinez, T.; Mauricio, J.C.; Mannazzu, I.; Coi, A.L.; Bou Zeidan, M.; Dequin, S.; et al. Flor Yeast: New Perspectives Beyond Wine Aging. Front. Microbiol. 2016, 7, 503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashraf, M.A.; Ullah, S.; Ahmad, I.; Qureshi, A.K.; Balkhair, K.S.; Abdur Rehman, M. Green biocides, a promising technology: Current and future applications to industry and industrial processes. J. Sci. Food Agric. 2014, 94, 388–403. [Google Scholar] [CrossRef] [PubMed]
- Grujić, S.; Vasić, S.; Radojević, I.; Čomić, L.; Ostojić, A. Comparison of the Rhodotorula mucilaginosa biofilm and planktonic culture on heavy metal susceptibility and removal potential. Water Air Soil Pollut. 2017, 228, 73. [Google Scholar] [CrossRef]
- Reynods, T. The Opi1p transcription factor affects expression of FLO11, mat formation, and invasive growth in Saccharomyces cerevisiae. Eukaryot. Cell 2006, 5, 1266–1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cullen, P.J. Biofilm/MAT assays for budding yeasts. Cold Spring Harb. Protoc. 2015, 2, 172–175. [Google Scholar]
- Zara, G.; Zara, S.; Pinna, C.; Marceddu, S.; Budroni, M. FLO11 gene length and transcriptional level affect biofilm forming ability of wild flor strains of Saccharomyces cerevisiae. Microbiology 2009, 155, 3838–3846. [Google Scholar] [CrossRef] [Green Version]
- Šťovíček, V.; Váchová, L.; Begany, M.; Wilkinson, D.; Palková, Z. Global changes in gene expression associated with phenotypic switching of wild yeast. BMC Genomics 2014, 15, 136. [Google Scholar] [CrossRef] [Green Version]
- Vopálenská, I.; St’ovícek, V.; Janderová, B.; Palkovà, Z. Role of distinct dimorphic transitions in territory colonizing and formation of yeast colony architecture. Environ. Microbiol. 2010, 12, 264–277. [Google Scholar] [CrossRef]
- Zara, S.; Bakalinsky, A.T.; Zara, G.; Pirino, G.; Demontis, M.A.; Budroni, M. FLO11-based model for air-liquid interfacial biofilm formation by Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2005, 71, 2934–2939. [Google Scholar] [CrossRef] [Green Version]
- Giobbe, S.; Marceddu, S.; Scherm, B.; Zara, G.; Mazzarello, V.; Budroni, M.; Migheli, Q. The strange case of a biofilm-forming strain of Pichia fermentans which controls Monilinia brown rot on apple but is pathogenic on peach fruit. FEMS Yeast Res. 2007, 7, 1389–1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saville, S.P.; Lazzell, A.L.; Monteagudo, C.; Lopez-Ribot, J.L. Engineered control of cell morphology in vivo reveals distinct roles for yeast and filamentous forms of Candida albicans during infection. Eukaryot. Cell 2003, 2, 1053–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whiteway, M.; Oberholzer, U. Candida morphogenesis and host-pathogen interactions. Curr. Opin. Microbiol. 2004, 7, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Nally, M.C.; Pesce, V.M.; Maturano, Y.P.; Muñoz, C.J.; Combina, M.; Toro, M.E.; de Figueroa, L.I.C.; Vazquez, F. Biocontrol of Botrytis cinerea in table grapes by non-pathogenic indigenous Saccharomyces cerevisiae yeasts isolated from viticultural environments in Argentina. Postharvest Biol. Technol. 2012, 64, 40–48. [Google Scholar] [CrossRef]
- Parafati, L.; Vitale, A.; Restuccia, C.; Cirvilleri, G. The effect of locust bean gum (LBG)-based edible coatings carrying biocontrol yeasts against Penicillium digitatum and Penicillium italicum causal agents of postharvest decay of mandarin fruit. Food Microbiol. 2016, 58, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Grzegorczyk, M.; Zarowska, B.; Restuccia, C.; Cirvilleri, G. Postharvest biocontrol ability of killer yeasts against Monilinia fructigena and Monilinia fructicola on stone fruit. Food Microbiol. 2017, 61, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Grzegorczyk, M.; Szalewicz, A.; Żarowska, B.; Połomska, X.; Wątorek, W.; Wojtatowicz, M. Microorganisms in biological control of phytopathogenic fungi. Acta Sci. Pol. Biotechnol. 2015, 14, 19–42. [Google Scholar]
- Liu, J.; Sui, Y.; Wisniewski, M.; Liu, Y. Review: Utilization of antagonistic yeasts to manage postharvest fungal diseases of fruit. Int. J. Food Microbiol. 2013, 167, 153–160. [Google Scholar] [CrossRef]
- Taavoni, S.; Habibi, A.; Varmira, K.; Alipour, S. Kinetics of continuous production of β-carotene in an airlift bioreactor. Asia-Pac. J. Chem. Eng. 2018, 13, e2160. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Species |
|---|---|
| DBVPG6091 | Rhodotorula mucilaginosa |
| DBVPG6094 | Rhodotorula mucilaginosa |
| DiSVAC2.5t1 | Rhodotorula mucilaginosa |
| DBVPG7019 | Rhodotorula mucilaginosa |
| DiSVAC71t0 | Rhodotorula mucilaginosa |
| CBS2366 | Rhodotorula glutinis |
| CBS2367 | Rhodotorula glutinis |
| CBS6016 | Rhodotorula toruloides |
| CBS315 | Rhodotorula toruloides |
| CBS7228 | Sporobolomyces oryzicola |
| CBS7998 | Rhodotorula cresolica |
| Strain | Abs570 (Mean ± Std) | ||
|---|---|---|---|
| PS | PP | PVC | |
| DBVPG6091 | 2.67 ± 0.32 | 3.43 ± 0.10 | 0.82 ± 0.13 |
| DBVPG6094 | 1.21 ± 0.05 | 1.32 ± 0.07 | 1.64 ± 0.21 |
| DBVPG7019 | 2.10 ± 0.47 | 0.88 ± 0.13 | 2.30 ± 0.56 |
| DiSVAC2.5t1 | 2.75 ± 0.13 | 2.93 ± 0.32 | 2.56 ± 0.12 |
| DiSVAC71t0 | 3.16 ± 0.13 | 3.17 ± 0.06 | 3.08 ± 0.28 |
| CBS2366 | 1.26 ± 0.12 | 1.47 ± 0.15 | 1.57 ± 0.14 |
| CBS2367 | 1.50 ± 0.63 | 2.55 ± 0.30 | 1.77 ± 0.15 |
| CBS6016 | 3.44 ± 0.04 | 3.40 ± 0.04 | 3.50 ± 0.00 |
| CBS315 | 2.18 ± 0.33 | 2.05 ± 0.18 | 1.85 ± 0.13 |
| CBS7228 | 2.11 ± 0.14 | 2.23 ± 0.40 | 2.14 ± 0.32 |
| CBS7998 | 0.88 ± 0.32 | 1.21 ± 0.09 | 1.70 ± 0.35 |
| UNISSM25 | 2.68 ± 0.06 | 2.71 ± 0.12 | 3.21 ± 0.21 |
| UNISS3238-32 Δflo11 | 0.48 ± 0.03 | 1.02 ± 0.12 | 1.11 ± 0.19 |
| Strain | Temperature (°C) | ||||
|---|---|---|---|---|---|
| 20 | 25 | 30 | 37 | 40 | |
| DBVPG7019 | + | + | + | - | - |
| CBS7228 | + | + | + | - | - |
| CBS315 | + | + | + | - | - |
| CBS2366 | - | + | - | - | - |
| DBVPG6094 | - | + | w | - | - |
| DBVPG6091 | - | + | + | - | - |
| CBS7899 | - | + | + | - | - |
| CBS2367 | - | + | - | - | - |
| CBS6016 | + | + | w | - | - |
| DiSVAC71t0 | + | + | + | - | - |
| DiSVAC2.5t1 | + | + | + | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zara, G.; Farbo, M.G.; Multineddu, C.; Migheli, Q.; Budroni, M.; Zara, S.; Mannazzu, I. Exploring the Biodiversity of Red Yeasts for In Vitro and In Vivo Phenotypes Relevant to Agri-Food-Related Processes. Fermentation 2021, 7, 2. https://doi.org/10.3390/fermentation7010002
Zara G, Farbo MG, Multineddu C, Migheli Q, Budroni M, Zara S, Mannazzu I. Exploring the Biodiversity of Red Yeasts for In Vitro and In Vivo Phenotypes Relevant to Agri-Food-Related Processes. Fermentation. 2021; 7(1):2. https://doi.org/10.3390/fermentation7010002
Chicago/Turabian StyleZara, Giacomo, Maria Grazia Farbo, Chiara Multineddu, Quirico Migheli, Marilena Budroni, Severino Zara, and Ilaria Mannazzu. 2021. "Exploring the Biodiversity of Red Yeasts for In Vitro and In Vivo Phenotypes Relevant to Agri-Food-Related Processes" Fermentation 7, no. 1: 2. https://doi.org/10.3390/fermentation7010002